Forecasting Extinctions: Uncertainties and Limitations

Abstract
:1. Introduction
“Prediction is very difficult, especially about the future”Nils Bohr, Nobel Prize winning physicist
2. A Typology of Extinction Forecasting
- How many species are likely to go extinct in area x over time t?
- What are the identities of the species with a high probability of extinction in area x over time t?
- What is the probability of species y going extinct in habitat x over time t?
- What is the probability that species y is already extinct in habitat x?

| Specificity | Key Question | Data available | Models |
|---|---|---|---|
| General | How many species will go extinct in area x over time t? | Estimates of species richness/endemism Estimates of habitat loss | Species-area relationshipNeutral theory |
| Environment-species richness correlations Extinction rate estimates | Extrapolation Simple deterministic relationships | ||
| What are the identities of the species with a high probability of extinction in area x over time t? | Species inventory Biogeographic data (e.g., distribution, dispersal, etc.) Ecological data (e.g., demography, etc.) | Threatened species lists Multispecies metapopulation models Expert judgement Ecosystem models Species distribution models Biocultural models | |
| What is the probability of species y going extinct in habitat x over time t? | Species inventory Biogeographic data Ecological data | Threatened species lists Metapopulation models Population viability analysis Expert judgement Ecosystem models Species distribution models Biocultural models Simple Extrapolation Deterministic models | |
| What is the probability that species y is already extinct in habitat x? | Historical records/sightings | Extrapolation based on sightings |

{kind=link}
{kind=link}
| Category of Model | Main Extinction Drivers | Key assumptions |
|---|---|---|
| Trend extrapolation | N/A | 1. Trend of decline will continue into the future 2. If trend is in sightings, observer efforts are temporally constant |
| Parametric/non-parametric models | Various simple deterministic | 1. Key causes of population decline are known, ongoing and will continue into the future 2. The model is correctly parameterized (e.g., the spatial distribution of the environmental variable/s is accurately modeled at an appropriate spatial resolution) |
| MVP analysis PVA | Small population size | 1. Demographic/population data is extensive and reliable 2. Distribution of vital rates between individuals and years is stationary in the future, or any changes can be predicted 3. Probability of catastrophes has been accurately assessed and incorporated |
| Meta-population models | Habitat fragmentation | 1. Accurate knowledge of existing sub-populations 2. Sub-populations are temporally and spatially stable 3. Dispersal potential accurately captured 4. Relative competitive abilities accurately captured |
| SARs | Habitat loss | 1. z value is accurately estimated 2. Habitat fragments act as true islands 3. No ‘small island effect’ 4. Current or future habitat loss is accurately estimated 5. System will eventually reach a phase that exhibits the same z-value as the past (this time period being difficult to predict) 6. Fragments have all been defined in the same way |
| Neutral Theory | 1. Correct species-distribution relationship identified 2. Range sizes are known or can be accurately estimated from abundance data 3. Ranges can be geographically located or realistically modelled 4. Species react in predictable way to different degrees of habitat transformation 5. Current or future habitat loss is accurately estimated 6. Fragments have all been defined in the same way | |
| Species Distribution Models | Climate change | 1. Minimal influence of evolution and phenotypic plasticity 2. Realistic spatially explicit climate scenario(s) used 3. Observed species distribution (if used) accurate and at equilibrium 4. Dispersal potential accurately captured 5. If SARs used see assumptions above |
| Ecosystem models | Trophic cascades | 1. Trophic relationships accurately mapped 2. Causal relationships between trophic interactions well understood |
| Threatened species lists | Multiple interacting | 1. Extinction risk criteria appropriately applied 2. Sub-criteria equally weighted 3. Subjective biases of experts understood and controlled for |
| Expert judgement | 1. Experts biases understood and controlled 2. Interacting factors appropriately identified and weighted | |
| Institutional capacity and ecological assessment | Biocultural | 1. Key institutions/actors identified 2. Relative capacity of institutions/actors to mount conservation interventions understood and assessed 3. Ecological extinction-risk assessment accurate |
2.1. Simple Extrapolations
2.2. Simple Deterministic Relationships Models
2.3. Population Viability Analysis
2.4. Metapopulation Models
2.5. Species-Area Models
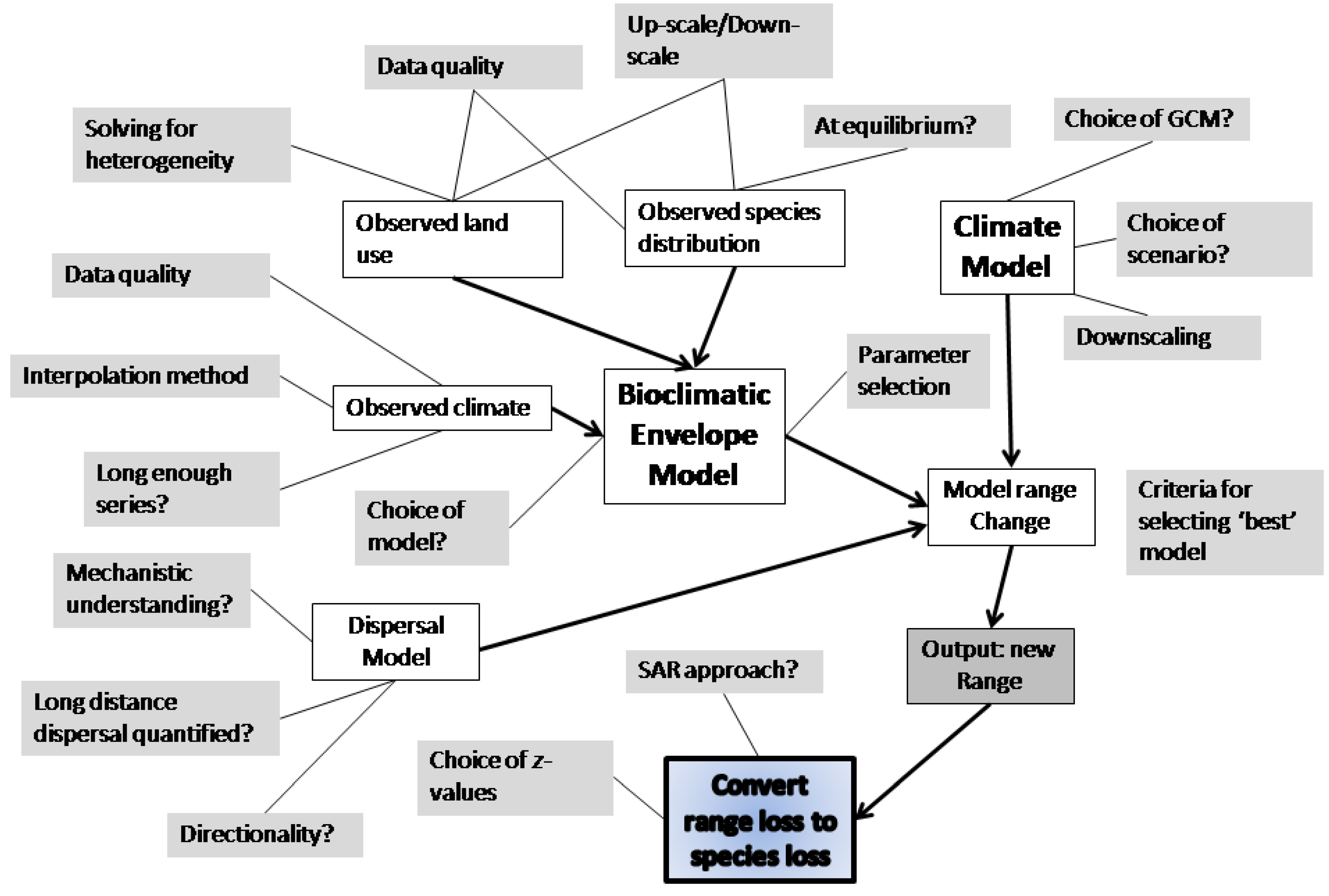
2.6. Species Distribution Models
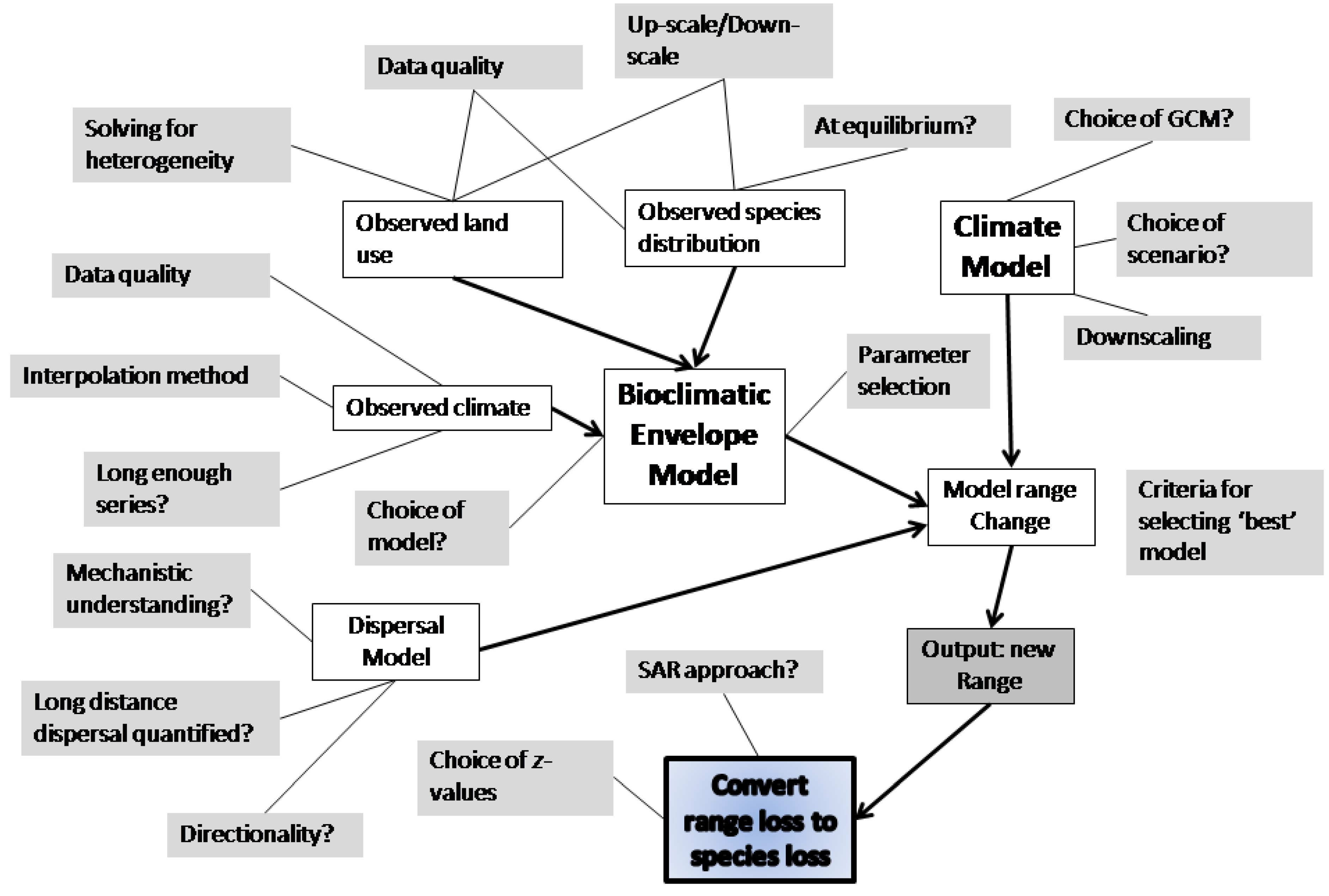
2.7. Ecosystem Models
2.8. Changes in Extinction Risk Categorization
2.9. Expert Judgement
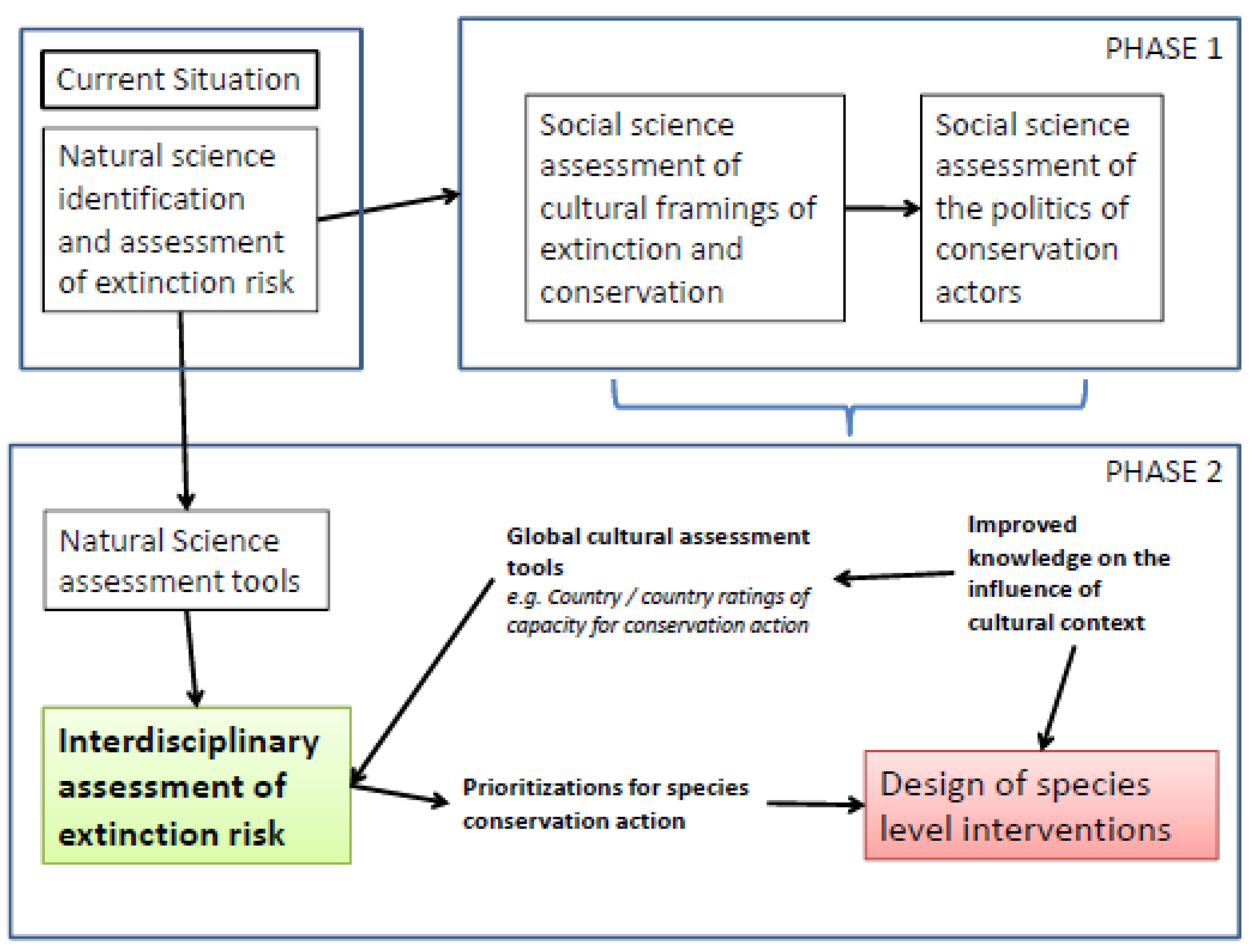
2.10. Biocultural Models
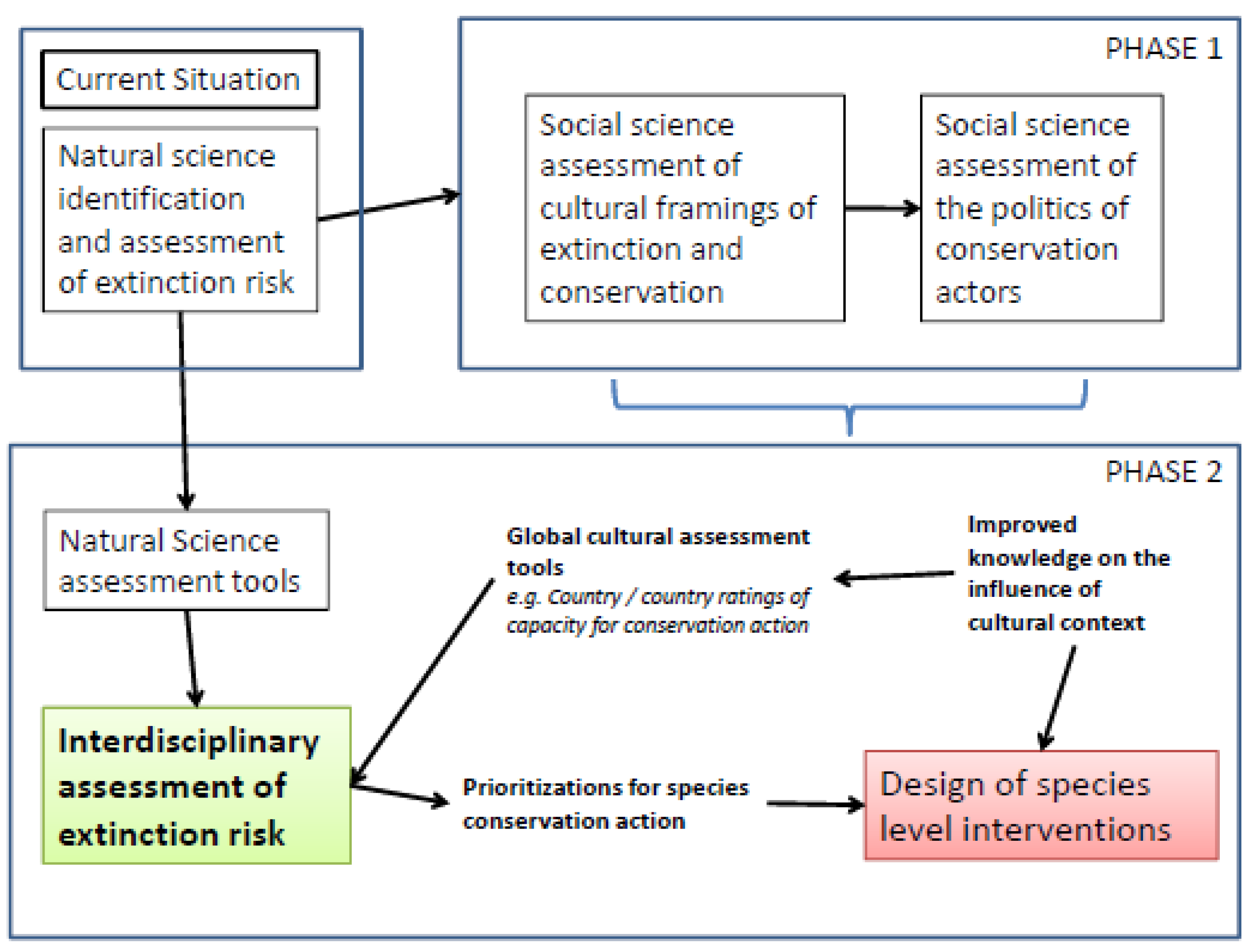
3. Conclusions
Acknowledgements
References and Notes
- Myers, N. The Sinking Ark; Pergamon: Oxford, UK, 2009. [Google Scholar]
- Lomborg, B. The Skeptical Environmentalist: Measuring the State of the World; Cambridge University: Cambridge, UK, 2001; p. 252. [Google Scholar]
- Ladle, R.J.; Jepson, P.; Araujo, M.B.; Whittaker, R.J. Dangers of crying wolf over risk of extinctions. Nature 2004, 482, 799. [Google Scholar]
- Ladle, R.J.; Jepson, P. Towards a biocultural theory of avoided extinction. Conserv. Lett. 2008, 1, 111–118. [Google Scholar] [CrossRef]
- Red List Categories and Criteria: Version 3.1; IUCN: Gland, Switzerland, Cambridge, UK; 2001.
- Roberts, D.L. Extinct or possibly extinct? Science 2005, 5776, 997–998. [Google Scholar]
- Butchart, S.H.M.; Statterfield, A.J.; Brooks, T.M. Going or gone: defining ‘Possibly Extinct’ species to give a truer picture of recent extinctions. Bull. B.O.C. 2006, 126a, 7–24. [Google Scholar]
- Wilson, E.O. The Diversity of Life; Belknap Press: Cambridge, MA, USA, 1992. [Google Scholar]
- Solow, A.J. Inferring extinction from a sighting record. Math. Biosci. 2005, 195, 47–55. [Google Scholar] [CrossRef]
- Fitzpatrick, J.W.; Lammertink, M.; Luneau, M.D., Jr; Gallagher, T.W.; Harrison, B.R.; Sparling, G.M.; Rosenberg, K.V.; Rohrbaugh, R.W.; Swarthout, E.C.H.; Wrege, P.H.; Swarthout, S.B.; Dantzker, M.S.; Charif, R.A.; Barksdale, T.R.; Remsen, J.V.; Simon, S.D.; Zollner, D. Ivory-billed woodpecker (Campephilus principalis) persists in continental North America. Science 2005, 5727, 1460–1462. [Google Scholar]
- Lozier, J.D.; Aniello, P.; Hickerson, M.J. Predicting the distribution of Sasquatch in western North America: anything goes with ecological niche modeling. J. Biogeog. 2009, 36, 1623–1627. [Google Scholar] [CrossRef]
- Braithewaite, R.W.; Muller, W.J. Rainfall, groundwater and refuges: predicting extinctions of Australian tropical mammal species. Aust. J. Ecol. 1997, 22, 57–67. [Google Scholar] [CrossRef]
- Norris, K. Managing threatened species: the ecological toolbox, evolutionary theory, and the declining population paradigm. J. Appl. Ecol. 2004, 41, 413–426. [Google Scholar] [CrossRef]
- Caughley, G. Directions in conservation biology. J. Anim. Ecol. 1994, 63, 215–244. [Google Scholar] [CrossRef]
- Akçakaya, H.R.; Sjögren-Gulve, P. Population viability analysis in conservation planning: an overview. Ecol. Bull. 2000, 48, 9–21. [Google Scholar]
- Gilpin, M.E.; Soulé, M.E. Minimum viable populations: process of extinction. In Conservation Biology: The Science of Scarcity and Diversity; Soulé, M.E., Ed.; Sinauer Associates: Sunderland, UK, 1986; pp. 19–34. [Google Scholar]
- Fagan, W.F.; Holmes, E.E. Quantifying the extinction vortex. Ecol. Lett. 2006, 9, 51–60. [Google Scholar]
- Hanski, I. A practical model of metapopulation dynamics. J. Anim. Ecol. 1994, 63, 151–162. [Google Scholar] [CrossRef]
- Bulman, C.R.; Wilson, R.J.; Holt, A.R.; Bravo, L.G.; Early, R.I.; Warren, M.S.; Thomas, C.D. Minimum viable metapopulation size, extinction debt, and the conservation of a declining species. Ecol. Appl. 2007, 17, 1460–1473. [Google Scholar] [CrossRef]
- Tilman, D.; May, R.M.; Lehman, C.L.; Nowak, M.A. Habitat destruction and the extinction debt. Nature 1994, 371, 65–66. [Google Scholar] [CrossRef]
- Malanson, G.P. Extinction debt: origins, developments and applications of a biogeographical trope. Prog. Phys. Geog. 2008, 32, 277–291. [Google Scholar] [CrossRef]
- Rosenzweig, M.L. Species Diversity in Space and Time; Cambridge University: Cambridge, UK, 1995; p. 9. [Google Scholar]
- Williams, M.R.; Lamont, B.B.; Henstridge, J.D. Species-area functions revisited. J. Biogeog. 2009, 36, 1994–2004. [Google Scholar] [CrossRef]
- Brooks, T.M.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Rylands, A.B.; Konstant, W.R.; Flick, P.; Pilgrim, J.; Oldfield, S.; Magin, G.; Hilton-Taylor, C. Habitat loss and extinction in the hotspots of biodiversity. Conserv. Biol. 2002, 16, 909–923. [Google Scholar] [CrossRef]
- Thomas, C.D.; Cameron, A.; Green, R.E.; Bakkenes, M.; Beaumont, L.J.; Collingham, Y.C.; Erasmus, B.F.N.; de Siqueira, M.F.; Grainger, A.; Hannah, L.; Hughes, L.; Huntley, B.; van Jaarsveld, A.S.; Midgley, G.F.; Miles, L.; Ortega-Huerta, M.; Peterson, A.T.; Phillips, O.L.; Williams, S.E. Extinction risk from climate change. Nature 2004, 427, 145–148. [Google Scholar] [CrossRef]
- Whittaker, R.J.; Fernández-Palacios, J.M. Island Biogeography: Ecology, Evolution, and Conservation, 2nd ed.; Oxford University: Oxford, UK, 2007. [Google Scholar]
- Hubbell, S.P.; He, F.; Condit, R.; Borda-de-Água, L.; Kellneri, J.; ter Steege, H. How many tree species are there in the Amazon and how many of them will go extinct? Proc. Natl. Acad. Sci. USA 2008, 105, 11498–11504. [Google Scholar]
- Wisz, M.S.; Guisan, A. Do pseudo-absence selection strategies influence species distribution models and their predictions? An information-theoretic approach based on simulated data. BMC. Ecol. 2009. Available online: http://www.biomedcentral.com/1472-6785/9/8 (accessed September 30,2009).
- Whittaker, R.J.; Araújo, M.B.; Jepson, P.; Ladle, R.J.; Watson, J.E.M.; Willis, K.J. Conservation biogeography: assessment and prospect. Divers. Distrib. 2005, 11, 3–23. [Google Scholar] [CrossRef]
- Pearson, R.G.; Dawson, T.P. Predicting the impacts of climate change on the distribution of species: are bioclimate envelope models useful? Glob. Ecol. Biogeog. 2003, 12, 361–371. [Google Scholar] [CrossRef]
- Dunn, R.R.; Nyeema, C.H.; Colwell, R.K.; Koh, L.P.; Sodhi, N.S. The sixth mass coextinction: are most endangered species parasites and mutualists? Proc. R. Soc. B. 2009, 276, 3037–3045. [Google Scholar] [CrossRef]
- Temple, S.A. Plant-Animal mutualism: co-evolution with dodo leads to near extinction of plant. Science 1977, 197, 885–886. [Google Scholar]
- Wittmer, M.C.; Cheke, A.S. The dodo and the tambalacoque tree: on obligate mutualism reconsidered. Oikos 1991, 61, 133–137. [Google Scholar] [CrossRef]
- da Silva, J.M.C.; Tabarelli, M. Tree species impoverishment and the future flora of the Atlantic forest of northeast Brazil. Nature 2000, 404, 72–74. [Google Scholar] [CrossRef]
- Dunne, J.A.; Williams, R.J. Cascading extinctions and community collapse in model food webs. Philos. Trans. R. Soc. B. 2009, 364, 1711–1723. [Google Scholar] [CrossRef]
- 2001 IUCN Red List Categories and Criteria Version 3.1; The International Union for Conservation of Nature: Gland, Switzerland. Available online: www.iucnredlist.org/info/ categories criteria2001 (accessed September 30, 2009).
- Brooke, M. de L.; Butchart, S.H.M.; Garnett, S.T.; Crowley, G.M.; Mantilla-Berniers, N.B.; Stattersfield, A.J. Rates of movement of threatened bird species between IUCN Red List categories and toward extinction. Conserv. Biol. 2008, 22, 417–427. [Google Scholar] [CrossRef]
- Mrosovsky, N. IUCN’s credibility critically endangered. Nature 1997, 389, 436. [Google Scholar] [CrossRef]
- Bomhard, B.; Richardson, D.M.; Donaldson, J.S. Potential impacts of future land use and climate change on the Red List status of the Proteaceae in the Cape Floristic region, South Africa. Glob. Change. Biol. 2005, 11, 1452–1468. [Google Scholar] [CrossRef]
- Akçakaya, H.R.; Butchart, S.H.M.; Mace, G.M.; Stuart, S.N.; Hilton-Taylor, C. Use and misuse of IUCN Red List criteria in predicting climate change impacts on biodiversity. Glob. Change. Biol. 2006, 12, 2037–2043. [Google Scholar] [CrossRef]
- Balint, P.J. How ethics shape the policy preferences of environmental scientists: what we can learn from Lomborg and his critics. Polit. Life Sci. 2003, 22, 14–23. [Google Scholar]
- Johns, A.D.; Ayres, J.M. Southern bearded sakis—beyond the brink. Oryx 1987, 21, 164–167. [Google Scholar] [CrossRef]
- Ferrari, S.F; Emidio-Silva, C.; Aparecida Lopes, M.; Bobadilla, U.L. Bearded sakis in south-eastern Amazonia—back from the brink? Oryx 1999, 33, 346–351. [Google Scholar]
- Rowe, G.; Wright, G. The Delphi technique as a forecasting tool: issues and analysis. Int. J. Forecast. 1999, 15, 353–375. [Google Scholar] [CrossRef]
- Courchamp, F.; Angulo, E.; Rivalain, P.; Hall, R.; Signoret, L.; Bull, L.; Meinard, Y. Rarity value and species extinction: the anthropogenic Allee effect. PLOS Biol. 2006, 4, e415. [Google Scholar] [CrossRef]
- Angulo, E.; Deves, A.-L.; Saint Jalmes, M.; Courchamp, F. Fatal attraction: rare species in the spotlight. P. Roy. Soc. B. 2009, 276, 1331–1337. [Google Scholar] [CrossRef]
- Jepson, P.; Ladle, R.J. Governing bird-keeping in Java and Bali: evidence from a household survey. Oryx 2009, 43, 364–374. [Google Scholar] [CrossRef]
- Araújo, M.B.; Whittaker, R.J.; Ladle, R.J.; Erhard, M. Reducing uncertainty in projections of extinction-risk from climate change. Glob. Ecol. Biogeog. 2005, 14, 529–539. [Google Scholar] [CrossRef]
- Keith, D.A.; Akçakaya, H.R.; Thuiller, W.; Midgley, G.F.; Pearson, R.G.; Phillips, S.J.; Regan, H.M.; Araújo, M.B.; Rebelo, T.G. Predicting extinction risks under climate change: coupling stochastic population models with dynamic bioclimatic habitat models. Biol. Lett. 2008, 4, 560–563. [Google Scholar] [CrossRef]
- Pimm, S.L. The dodo went extinct (and other ecological myths). Ann. Mo. Bot. Gard. 2002, 89, 190–198. [Google Scholar] [CrossRef]
- Kuussaari, M.; Bommarco, R.; Heikkinen, R.K.; Helm, A.; Krauss, J.; Lindborg, R.; Öckinger, E.; Pärtel, M.; Pino, J.; Rodà, F.; Stefanescu, C.; Teder, T.; Zobel, M.; Steffan-Dewenter, I. Extinction debt: a challenge for biodiversity conservation. Trends Ecol. Evol. 2009, 24, 564–571. [Google Scholar] [CrossRef]
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ladle, R.J. Forecasting Extinctions: Uncertainties and Limitations. Diversity 2009, 1, 133-150. https://doi.org/10.3390/d1020133
Ladle RJ. Forecasting Extinctions: Uncertainties and Limitations. Diversity. 2009; 1(2):133-150. https://doi.org/10.3390/d1020133
Chicago/Turabian StyleLadle, Richard J. 2009. "Forecasting Extinctions: Uncertainties and Limitations" Diversity 1, no. 2: 133-150. https://doi.org/10.3390/d1020133
APA StyleLadle, R. J. (2009). Forecasting Extinctions: Uncertainties and Limitations. Diversity, 1(2), 133-150. https://doi.org/10.3390/d1020133