Global Amphibian Extinction Risk Assessment for the Panzootic Chytrid Fungus

Abstract
:1. Introduction
2. Material and Methods
2.1. Prediction of Bd Distribution
2.2. Identification of the Most Threatened Species
2.3. Bd Risk Factor for Anuran Amphibians
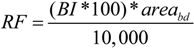
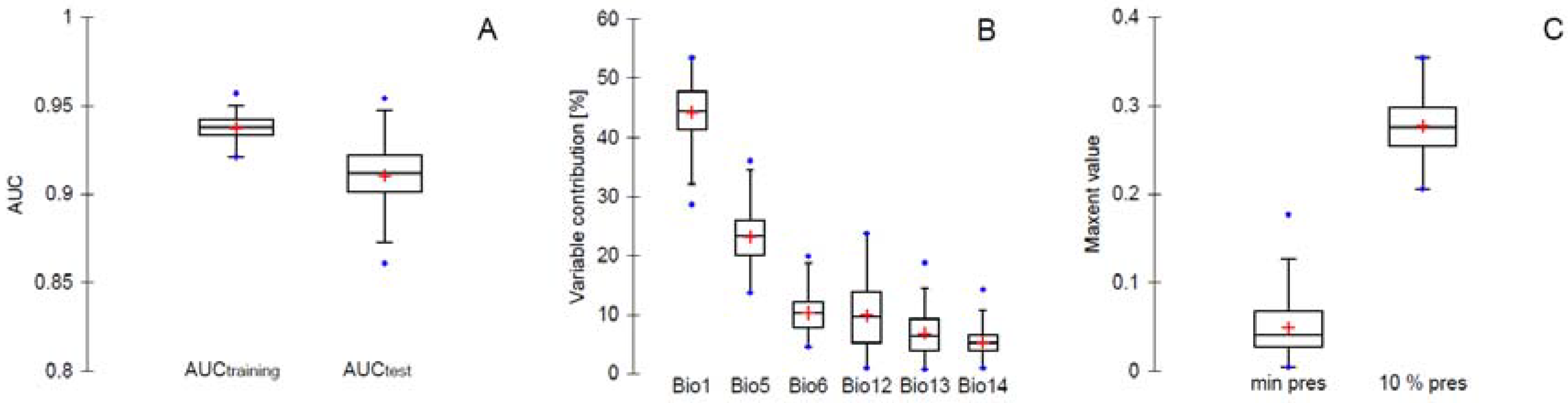
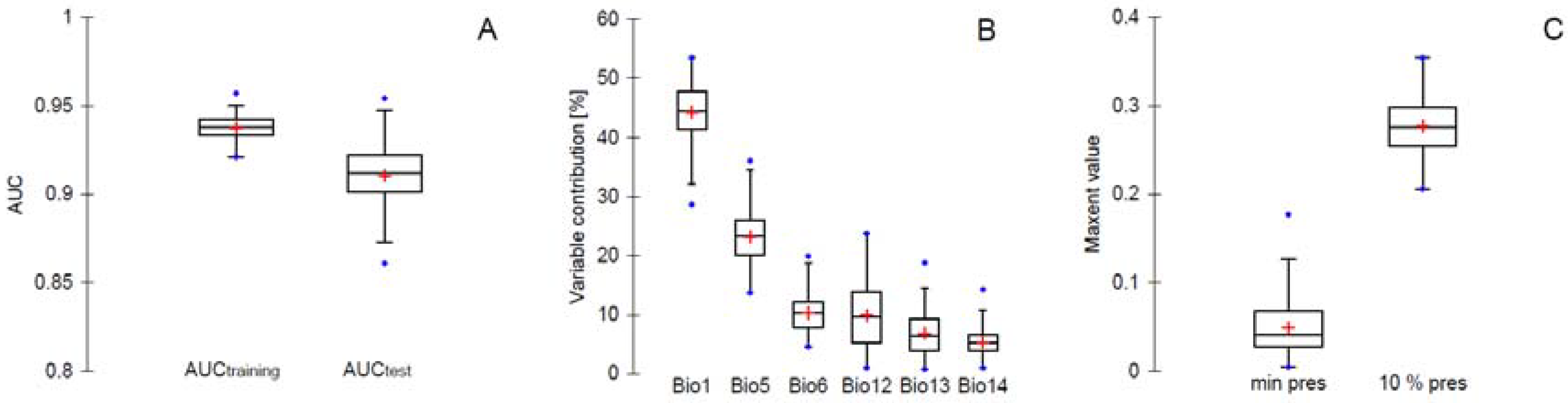
3. Results and Discussion
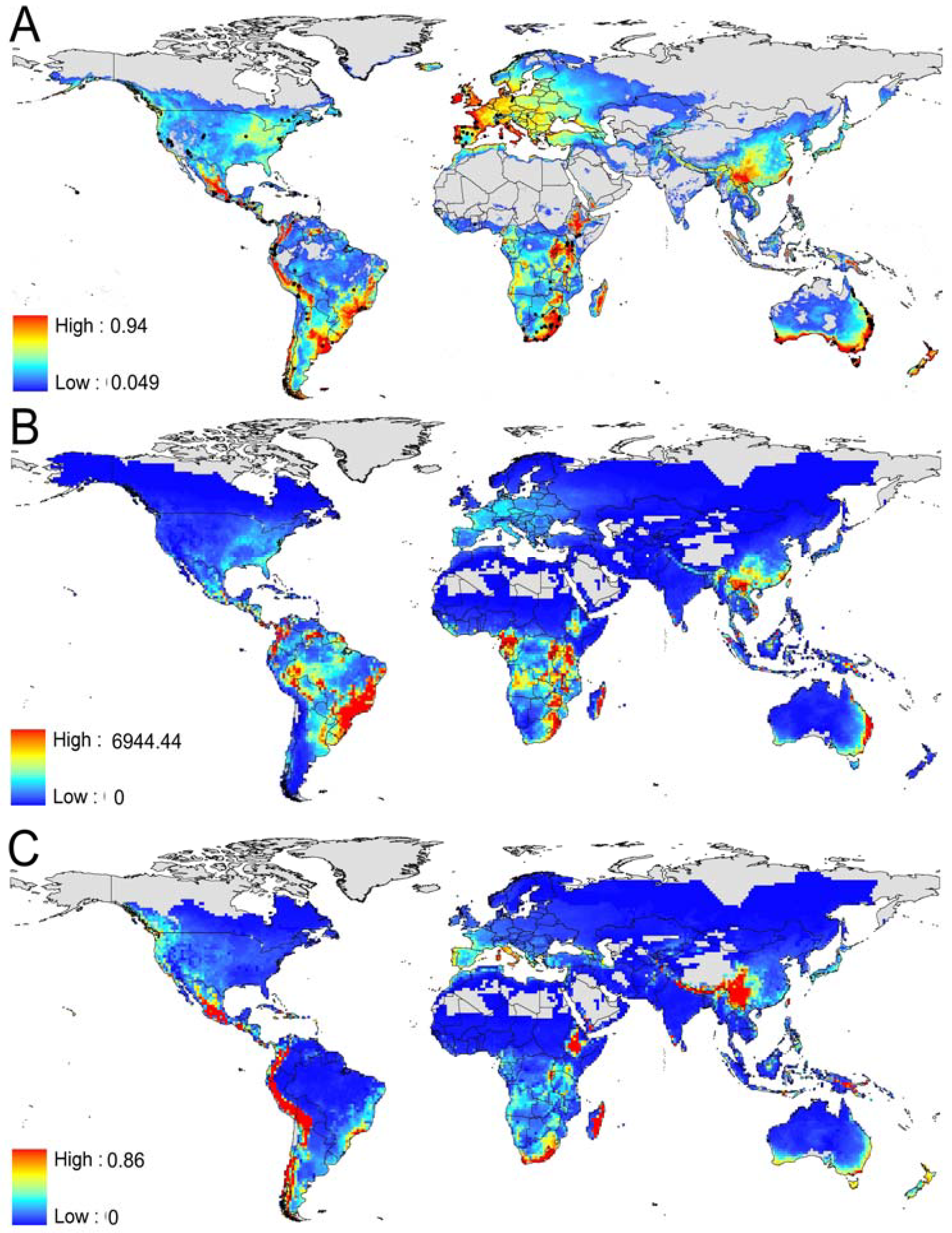
3.1. Prediction of Bd Distribution
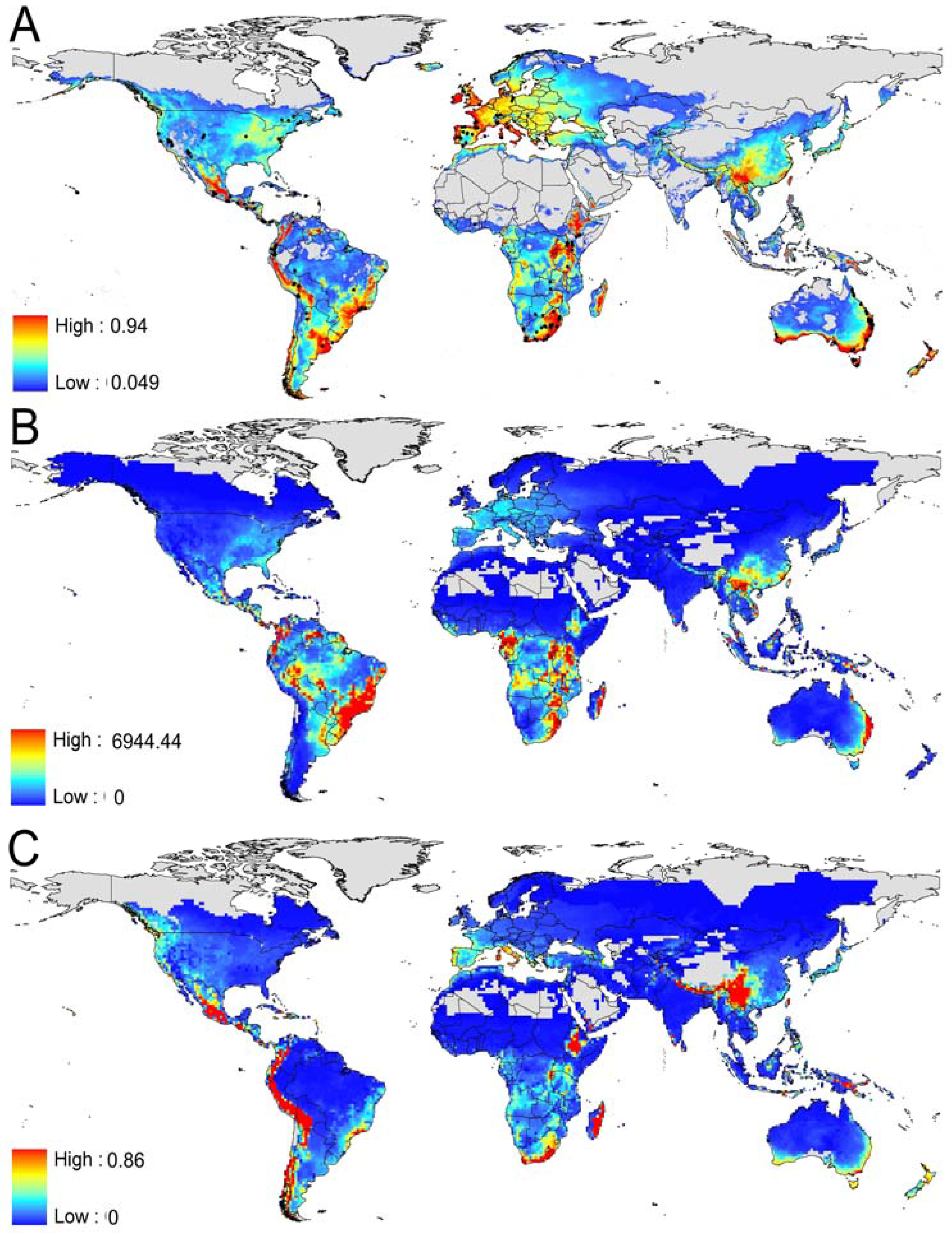
3.2. Identification of the Most Threatened Species

{kind=link}
{kind=link}
{kind=link}
| Portion of species' range environmentally suitable (>0.5/minimum observed training presence) | |||
|---|---|---|---|
| 100% | 75% | 50% | |
| Caecilians (172 species) | 10/90 | 27/132 | 38/138 |
| Salamanders (551 species) | 129/498 | 195/543 | 226/545 |
| Frogs and toads (5,532 species) | 961/3,905 | 1,756/4,953 | 2,259/5,211 |
| Total (6,142 species) | 1,100/4,493 | 1,978/5,628 | 2,523/5,894 |
3.3. Bd Risk Factor for Anuran Amphibians
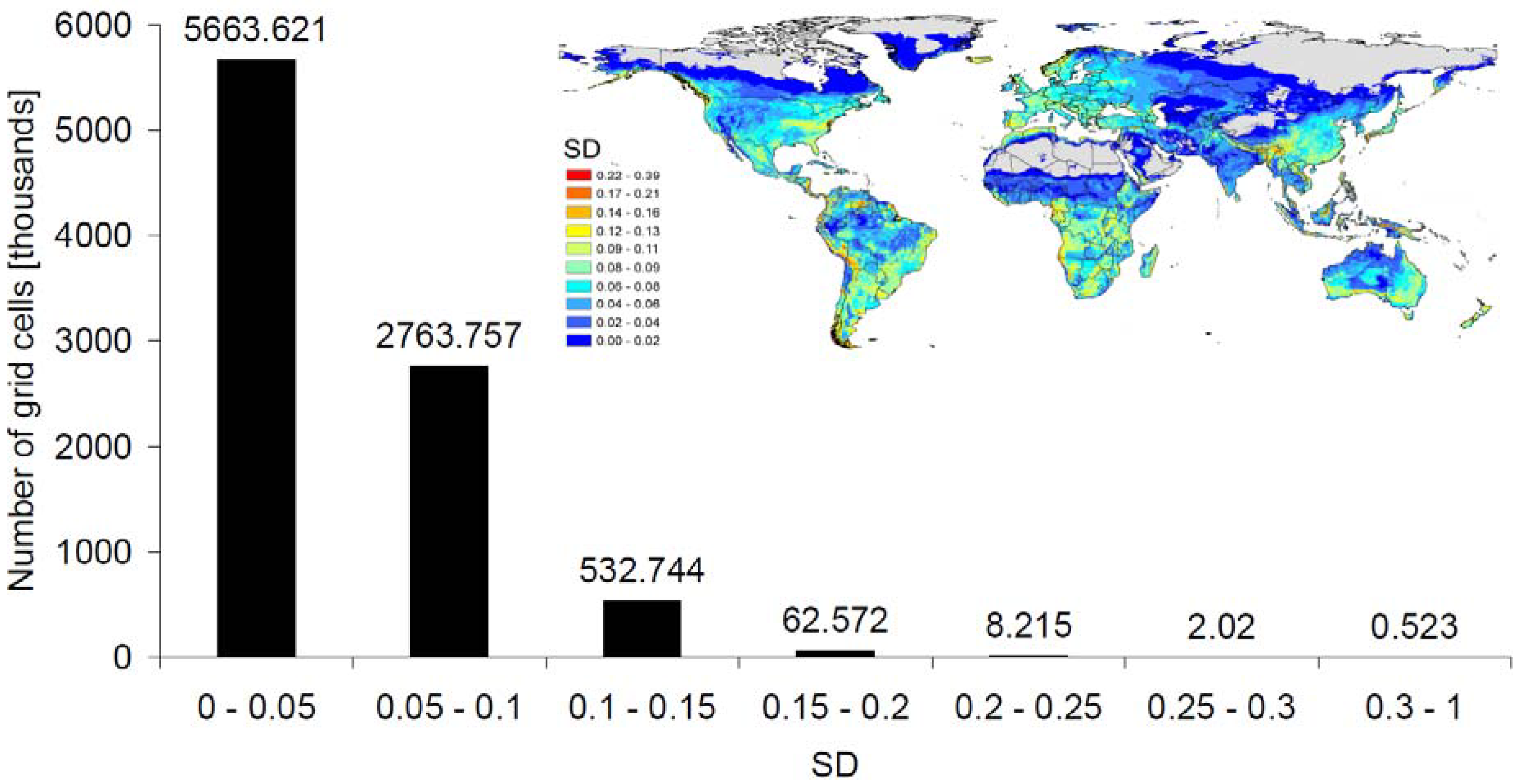
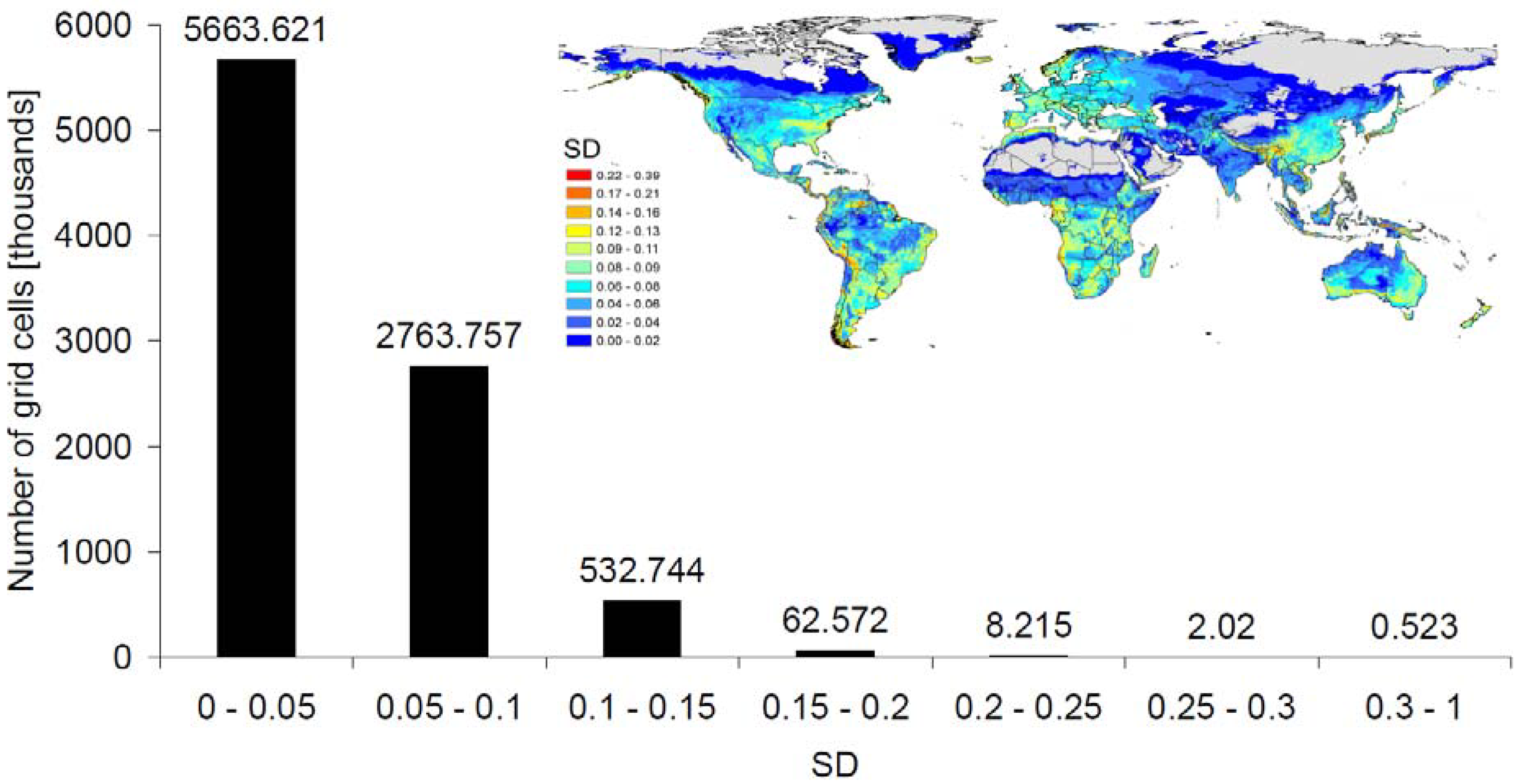
3.4. Methodological Considerations
4. Conclusions
Acknowledgements
Supplementary Material
References
- Stuart, S.; Chanson, J.S.; Cox, N.A.; Young, B.E.; Rodrigues, A.S.L.; Fischman, D.L.; Waller, R.W. Status and trends of amphibian declines and extinctions worldwide. Science 2004, 306, 1783–1786. [Google Scholar] [CrossRef] [PubMed]
- Berger, L.; Speare, R.; Daszak, P.; Green, D.E.; Cunningham, A.A.; Goggin, C.L.; Slocombe, R.; Ragan, M.A.; Hyatt, A.D.; McDonald, K.R.; Hines, H.B.; Lips, K.R.; Marantelli, G.; Parkes, H.T. Chytridiomycosis causes amphibian mortality associated with population declines in the rain forests of Australia and Central America. Proc. Natl. Acad. Sci. USA 1998, 95, 9031–9036. [Google Scholar] [CrossRef] [PubMed]
- Mendelson, J.R., III; Lips, K.R.; Gagliardo, R.W.; Rabb, G.B.; Collins, J.P.; Diffendorfer, J.E.; Daszack, P.; Ibáñez, R.; Zippel, K.C.; Lawson, D.P.; Wright, K.M.; Stuart, S.N.; Gascon, C.; Silva, H.R.; Burrowes, P.A.; Joglar, R.L.; La Marca, E.; Lötters, S.; Perez, L.H.; Weldon, C.; Hyatt, A.; Rodriguez-Mahecha, J.V.; Hunt, S.; Robertson, H.; Lock, B.; Raxworthy, C.J.; Frost, D.R.; Lacy, R.C.; Alfrod, R.A.; Campbell, J.A.; Parra-Olea, G.; Bolaños, F.; Domingo, J.J.C.; Halliday, T.; Murphy, J.B.; Wake, M.H.; Coloma, L.A.; Kuzmin, S.L.; Price, M.S.; Howell, K.M.; Lau, M.; Pethiyagoda, R.; Boone, M.; Lannoo, M.J.; Blaustein, A.R.; Dobson, A.; Griffiths, A.; Crump, M.L.; Wake, D.B.; Brodie, E.D.J. Confronting amphibian declines and extinctions. Science 2006, 313, 48. [Google Scholar]
- Gascon, C.; Collins, J.P.; Moore, R.D.; Church, D.R.; McKay, J.E.; Mendelson, J.R., III. Amphibian Conservation Action Plan; IUCN: Gland, Switzerland, 2007. [Google Scholar]
- Skerratt, L.F.; Berger, L.; Speare, R.; Cashins, S.; McDonald, K.R.; Phillott, A.D.; Hines, H.B.; Kenyon, N. Spread of chytridiomycosis has caused the rapid global decline and extinction of frogs. EcoHealth 2007, 4, 125–134. [Google Scholar] [CrossRef]
- Wake, D.B.; Vredenburg, V.T. Are we in the midst of the sixth mass extinction? A review from the world of amphibians. Proc. Natl. Acad. Sci. USA 2008, 105, 11466–11473. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.N.; Lauer, A.; Simon, M.A.; Banning, J.L.; Alford, R.A. Addition of antifungal skin bacteria to salamanders ameliorates the effects of chytridiomycosis. Dis. Aquat. Org. 2009, 83, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Deutsch, C.A.; Tweksbury, J.J.; Huey, R.B.; Sheldon, K.S.; Ghalambor, C.K.; Haak, D.C.; Martin, P.R. Impacts of climate warming on terrestrial ectotherms across latitude. Proc. Natl. Acad. Sci. USA 2008, 105, 6668–6672. [Google Scholar] [CrossRef] [PubMed]
- Lips, K.R.; Diffendorfer, J.; Mendelson, J.R., III; Sears, M.W. Riding the wave: reconciling the roles of disease and climate change in amphibian declines. PLoS Biol. 2008, 6, 441–454. [Google Scholar]
- Woodhams, D.C.; Alford, R.A.; Briggs, C.J.; Johnson, M.; Rollins-Smith, L.A. Life-history trade-offs influence disease in changing climates: strategies of an amphibian pathogen. Ecology 2008, 89, 1627–1639. [Google Scholar] [CrossRef] [PubMed]
- Piotrowski, J.S.; Annis, S.L.; Longcore, J.E. Physiology of Batrachochytrium dendrobatidis, a chytrid pathogen of amphibians. Mycologia 2004, 96, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Kriger, K.M.; Hero, J.-M. Large-scale seasonal variation in the prevalence and severity of chytridiomycosis. J. Zool. 2007, 271, 352–359. [Google Scholar]
- Kriger, K.M.; Peregolou, F.; Hero, J.-M. Latitudinal variation in the prevalence and intensity of chytrid (Batrachochytrium dendrobatidis) infection in eastern Australia. Conserv. Biol. 2007, 21, 1280–1290. [Google Scholar] [CrossRef] [PubMed]
- Rödder, D.; Veith, M.; Lötters, S. Environmental gradients explaining prevalence and intensity of infection with the amphibian chytrid fungus: The host's perspective. Anim. Conserv. 2008, 11, 513–517. [Google Scholar] [CrossRef]
- Woodhams, D.C.; Alford, R.A.; Marantelli, G. Ecology of chytridiomycosis in rainforest stream frog assemblages of tropical Queensland. Conserv. Biol. 2005, 19, 1449–1459. [Google Scholar] [CrossRef]
- Kielgast, J.; Rödder, D.; Veith, M.; Lötters, S. Widespread occurrence of the amphibian chytrid fungus in Kenya. Animal Cons. 2009, (in press). [Google Scholar]
- Blackburn, J.K.; McNyset, K.M.; Curtins, A.; Hugh-Jones, M.E. Modeling the geographic distribution of Bacillus anthracis, the causative agent of Anthrax disease, for the contiguous United States using predictive ecological niche modeling. Am. J. Trop. Med. Hyg. 2007, 77, 1103–1110. [Google Scholar] [PubMed]
- Ostfeld, R.S.; Glass, G.E.; Keesing, F. Spatial epidemiology: an emerging (or re-emerging) discipline. TREE 2005, 20, 328–336. [Google Scholar] [PubMed]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Hutchinson, M.F. Anusplin version 4.3, Centre for Resource and Environment Studies; The Australian National University: Canberra, Australia, 2004.
- Hijmans, R.J.; Cruz, J.M.; Rojas, E.; Guarino, L. Diva-GIS, version 1.4. A Geographic Information System for the Management and Analysis of Genetic Resources Data. Manual, International Potato Center and International Plant Genetic Resources Institute: Lima, Peru, 2001.
- Heikkinen, R.K.; Luoto, M.; Araújo, M.B.; Virkkala, R.; Thuiller, W.; Sykes, M.T. Methods and uncertainties in bioclimatic envelope modeling under climate change. Progr. Phys. Geogr. 2006, 30, 751–777. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Graham, C.H. The ability of climate envelope models to predict the effect of climate change on species distributions. Glob. Change Biol. 2006, 12, 2272–2281. [Google Scholar] [CrossRef]
- Ficetola, G.F.; Thuiller, W.; Miaud, C. Prediction and validation of the potential global distribution of a problematic alien species—the American bullfrog. Divers. Distrib. 2007, 13, 476–485. [Google Scholar] [CrossRef]
- Fitzpatrick, M.C.; Weltzin, J.F.; Sanders, N.J.; Dunn, R.R. The biogeography of prediction error: Why does the introduced range of the fire ant over-predict its native range? Glob. Ecol. Biogeogr. 2007, 16, 24–33. [Google Scholar] [CrossRef]
- Puschendorf, R.; Carnaval, A.C.; Vanderwal, J.; Zumbado-Ulate, H.; Chaves, G.; Bolaños, F.; Alford, R.A. Distribution models for the amphibian chytrid Batrachochytrium dendrobatidis in Costa Rica: proposing climatic refuges as a conservation tool. Divers. Distrib. 2009, 15, 401–408. [Google Scholar] [CrossRef]
- Dormann, C.F.; McPherson, J.; Araújo, M.B.; Bivand, R.; Bollinger, J.; Carl, G.; Davies, R.G.; Hirzel, A.; Jetz, W.; Kissling, W.D.; Kühn, I.; Ohlemüller, R.; Peres-Neto, P.R.; Reineking, B.; Schröder, B.; Schurr, F.M.; Wilson, R. Methods to account for spatial autocorrelation in the analysis of species distributional data: A review. Ecography 2007, 30, 609–628. [Google Scholar] [CrossRef]
- Ron, S.R. Predicting the distribution of the amphibian pathogen Batrachochytrium dendrobatidis in the new world. Biotropica 2005, 37, 209–221. [Google Scholar] [CrossRef]
- Wisz, M.S.; Hijmans, R.J.; Peterson, A.T.; Graham, C.H.; Guisan, A.; NPSDW Group. Effects of sample size on the performance of species distribution models. Divers. Distrib. 2008, 14, 763–773. [Google Scholar]
- Phillips, S.J.; Dudík, M. Modelling of species distributions with Maxent: new extensions and comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Phillips, S.J. Transferability, sample selection bias and background data in presence-only modelling: a response to Peterson et al. (2007). Ecography 2008, 31, 272–278. [Google Scholar]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudik, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; Li, J.; Lohmann, L.G.; Loiselle, B.A.; Manion, G.; Moritz, C.; Nakamura, M.; Nakazawa, Y.; Overton, J.M.M.; Peterson, A.T.; Phillips, S.J.; Richardson, K.; Scachetti-Pereira, R.; Shapire, R.E.; Soberón, J.; Williams, S.; Wisz, M.S.; Zimmermann, N.E. Novel methods improve prediction of species´ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Araújo, M.B.; New, M. Ensemble forecasting of species distributions. TREE 2007, 22, 42–47. [Google Scholar] [PubMed]
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Manel, S.; Williams, H.C.; Ormerod, S.J. Evaluating presence-absence models in ecology: The need to account for prevalence. J. Appl. Ecol. 2001, 38, 921–931. [Google Scholar] [CrossRef]
- Swets, K. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef] [PubMed]
- Vanderwal, J.; Shoo, L.P.; Johnson, C.N.; Williams, S.E. Abundance and the environmental niche: environmental suitability estimated from niche models predicts the upper limit of local abundance. Am. Nat. 2009, 174, 282–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Pam, M.; Dawson, T.P.; Pearson, R.G. Selecting thresholds of occurrence in the prediction of species distributions. Ecography 2005, 28, 385–393. [Google Scholar] [CrossRef]
- Frost, D.R.; Grant, T.; Faivovich, J.; Bain, R.H.; Haas, A.; Haddad, C.F.B.; de Sá, R.O.; Channing, A.; Wilkinson, M.; Donnellan, S.C.; Raxworthy, C.J.; Campbell, J.A.; Blotto, B.L.; Moler, P.; Drewes, R.C.; Nussbaum, R.A.; Lynch, J.D.; Green, M.D.; Wheeler, W.C. The amphibian tree of life. Bull. Am. Mus. Nat. Hist. 2006, 297, 1–370. [Google Scholar] [CrossRef]
- Grant, T.; Frost, D.R.; Caldwell, J.P.; Gagliardo, R.; Haddad, C.F.B.; Kok, P.J.R.; Means, D.B.; Noonan, B.P.; Schargel, W.E.; Wheeler, W.C. Phylogenetic systematics of dart-poison frogs and their relatives (Amphibia, Athesphatanura, Dendrobatidae). Bull. Am. Mus. Nat. Hist. 2006, 299, 1–262. [Google Scholar] [CrossRef]
- Bielby, J.; Cooper, N.; Cunningham, A.A.; Garner, T.W.J.; Purvis, A. Predicting susceptibility to rapid declines in the world’s frogs. Conserv. Lett. 2008, 1, 82–90. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Willmott, C.J.; Kenji, M. Terrestrial Air Temperature and Precipitation: Monthly and Annual Time Series (1950-1999). Available online: http://climate.geog.udel.edu/~climate/html_pages/README.ghcn_ts2.html (access 8 July 2009).
- CIESIN. Center for International Earth Science Information Network; Columbia University: New York, NY, USA, 2000. Available online: http://www.ciesin.columbia.edu/download_data.html (access on 8 July 2009).
- Fisher, M.C.; Garner, T.W.J. The relationship between the emergence of Batrachochytrium dendrobatidis, the international trade in amphibians and introduced amphibian species. Fungal Biol. Rev. 2007, 21, 2–9. [Google Scholar] [CrossRef]
- Weldon, C.; du Preez, L.H.; Hyatt, A.D.; Muller, R.; Spears, R. Origin of the amphibian chytrid fungus. Emerg. Infect. Dis. 2004, 10, 2100–2105. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, A.A. A walk on the wild side - emerging wildlife diseases. Br. Med. J. 2005, 331, 1214–1215. [Google Scholar] [CrossRef]
- Cunningham, A.A.; Daszak, P.; Rodriguez, J.P. Pathogen pollution: defining a parasitological threat to biodiversity conservation. J. Parasitol. 2003, 9, S78–S83. [Google Scholar]
- Gaston, K.J.; Fuller, R.A. The sizes of species' geographic ranges. J. Appl. Ecol. 2009, 46, 1–9. [Google Scholar]
- Araújo, M.B.; Pearson, R.G. Equilibrium of species' distribution with climate. Ecography 2005, 28, 693–695. [Google Scholar] [CrossRef]
- Guisan, A.; Zimmermann, N. Predictive habitat distribution models in ecology. Ecol. Model. 2000, 135, 147–186. [Google Scholar] [CrossRef]
- Blehert, D.S.; Hicks, A.C.; Behr, M.; Meteyer, C.U.; Berlowski-Zier, B.M.; Buckles, E.L.; Coleman, J.T.H.; Darling, S.R.; Niver, R.; Okoniewski, J.C.; Rudd, R.J.; Stone, W.B. Bat white-nose syndrome: An emerging fungal pathogen? Science 2009, 323, 227. [Google Scholar] [CrossRef] [PubMed]
- Rahbek, C. Disease ecology: the silence of the robins. Nature 2007, 447, 652–653. [Google Scholar] [CrossRef] [PubMed]
- La Marca, E.; Lips, K.R.; Lötters, S.; Puschendorf, R.; Ibáñez, R.; Rueda-Almonacid, J.V.; Schulte, R.; Marty, C.; Castro, F.; Manzanilla-Puppo, J.; García, J.E.; Bolaños, F.; Chaves, G.; Pounds, J.A.; Toral, E.; Young, B.E. Catastrophic population declines and extinctions in Neotropical harlequin frogs (Bufonidae: Atelopus). Biotropica 2005, 37, 190–201. [Google Scholar] [CrossRef]
- Lampo, M.; Rodríguez, A.; La Marca, E.; Daszak, P. A chytridiomycosis epidemic and a severe dry season precede the disappearance of Atelopus species from the Venezuelan Andes. J. Herpetol. 2007, 16, 395–402. [Google Scholar]
- Barrionuevo, S.; Mangione, S. Chytridiomycosis in two species of Telmatobius (Anura: Leptodactylidae) from Argentina. Dis. Aquat. Org. 2006, 73, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Kriger, K.M.; Hero, J.-M. The chytrid fungus Batrachochytrium dendrobatidis is non-randomly distributed across amphibian breeding habitats. Divers. Distrib. 2007, 13, 781–788. [Google Scholar] [CrossRef]
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Rödder, D.; Kielgast, J.; Bielby, J.; Schmidtlein, S.; Bosch, J.; Garner, T.W.J.; Veith, M.; Walker, S.; Fisher, M.C.; Lötters, S. Global Amphibian Extinction Risk Assessment for the Panzootic Chytrid Fungus. Diversity 2009, 1, 52-66. https://doi.org/10.3390/d1010052
Rödder D, Kielgast J, Bielby J, Schmidtlein S, Bosch J, Garner TWJ, Veith M, Walker S, Fisher MC, Lötters S. Global Amphibian Extinction Risk Assessment for the Panzootic Chytrid Fungus. Diversity. 2009; 1(1):52-66. https://doi.org/10.3390/d1010052
Chicago/Turabian StyleRödder, Dennis, Jos Kielgast, Jon Bielby, Sebastian Schmidtlein, Jaime Bosch, Trenton W. J. Garner, Michael Veith, Susan Walker, Matthew C. Fisher, and Stefan Lötters. 2009. "Global Amphibian Extinction Risk Assessment for the Panzootic Chytrid Fungus" Diversity 1, no. 1: 52-66. https://doi.org/10.3390/d1010052
APA StyleRödder, D., Kielgast, J., Bielby, J., Schmidtlein, S., Bosch, J., Garner, T. W. J., Veith, M., Walker, S., Fisher, M. C., & Lötters, S. (2009). Global Amphibian Extinction Risk Assessment for the Panzootic Chytrid Fungus. Diversity, 1(1), 52-66. https://doi.org/10.3390/d1010052