Anti-Cancer Effects of Xanthones from Pericarps of Mangosteen

Abstract
1. Overview
2. Introduction
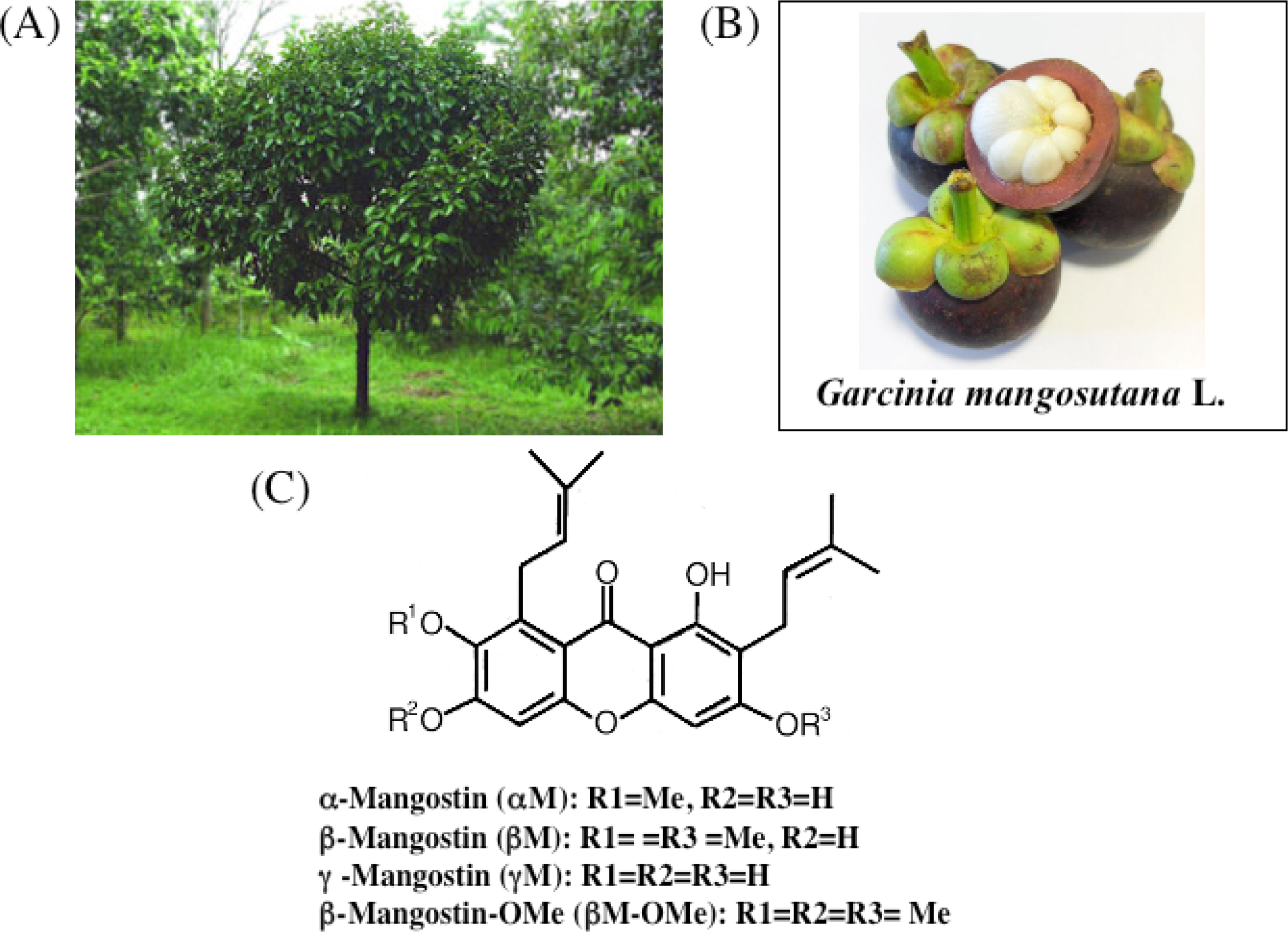
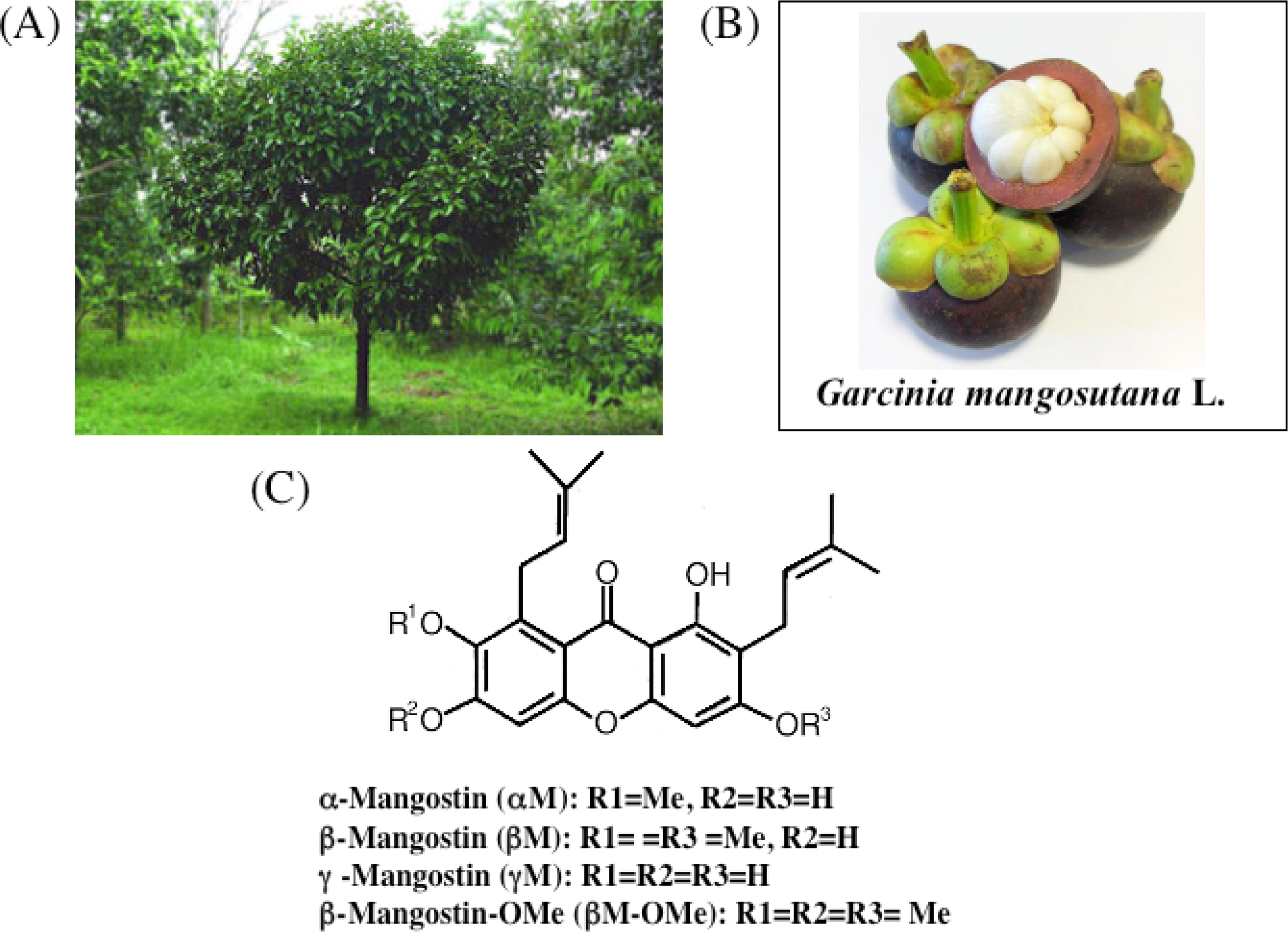
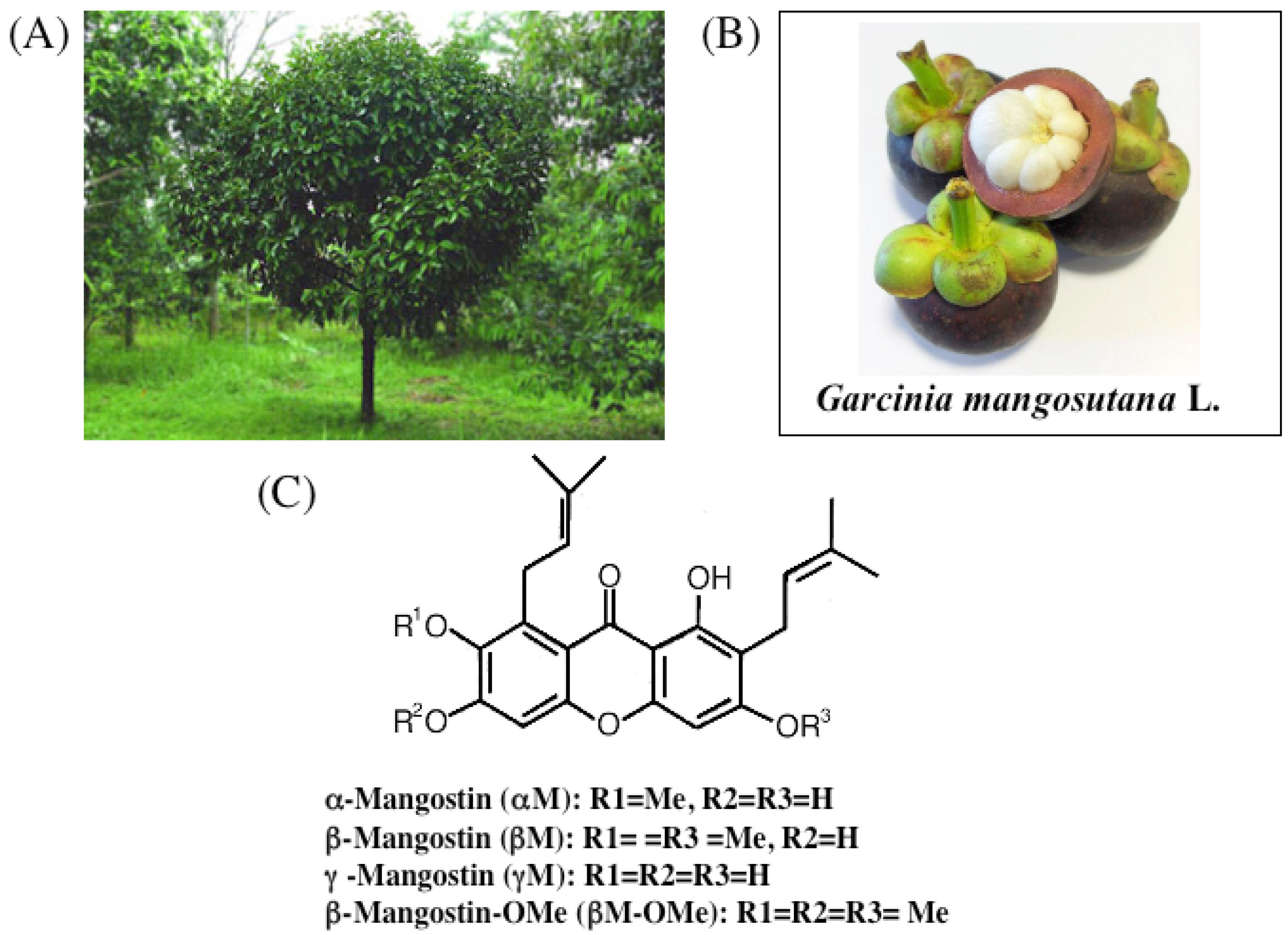
3. Chemistry of Xanthones
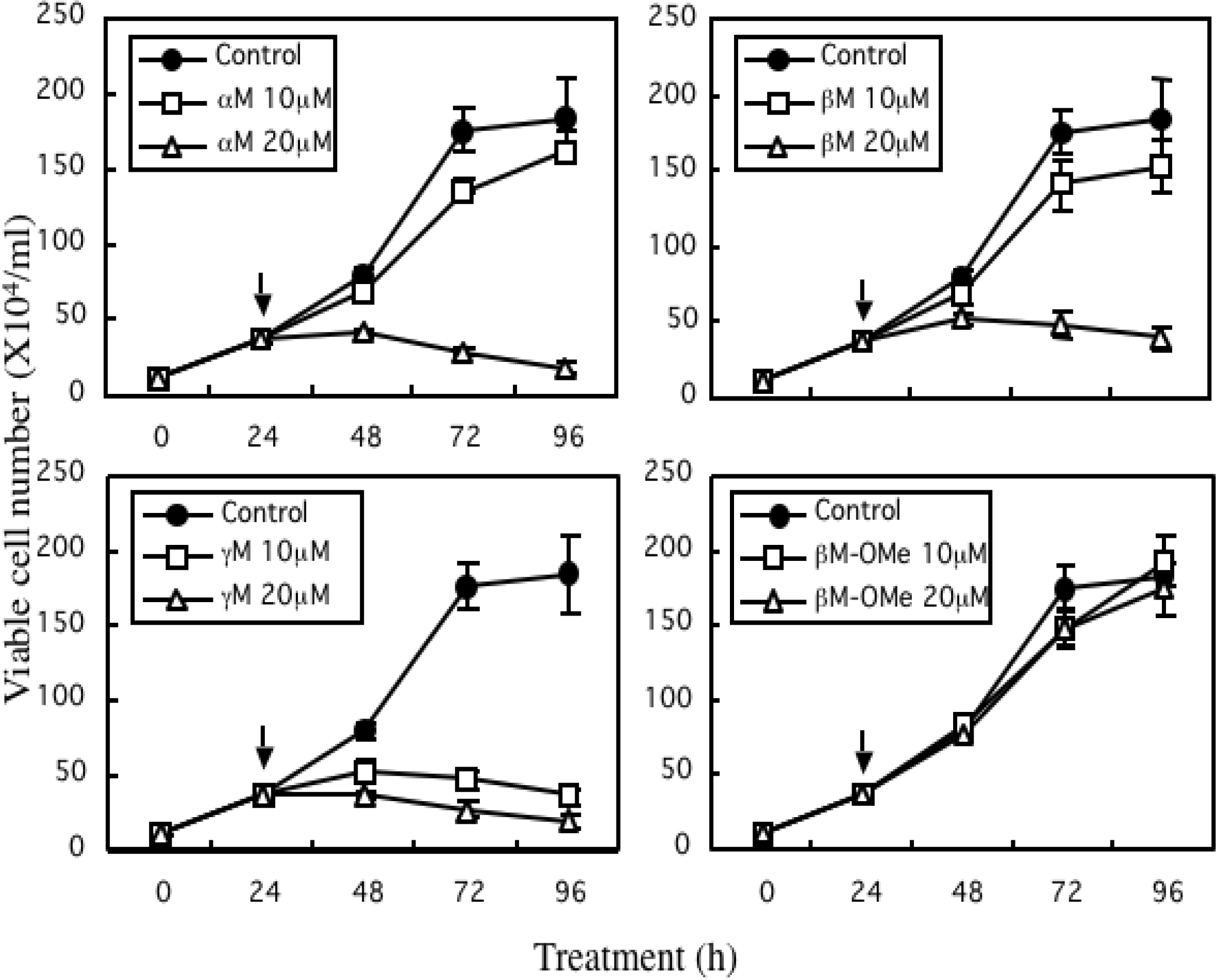
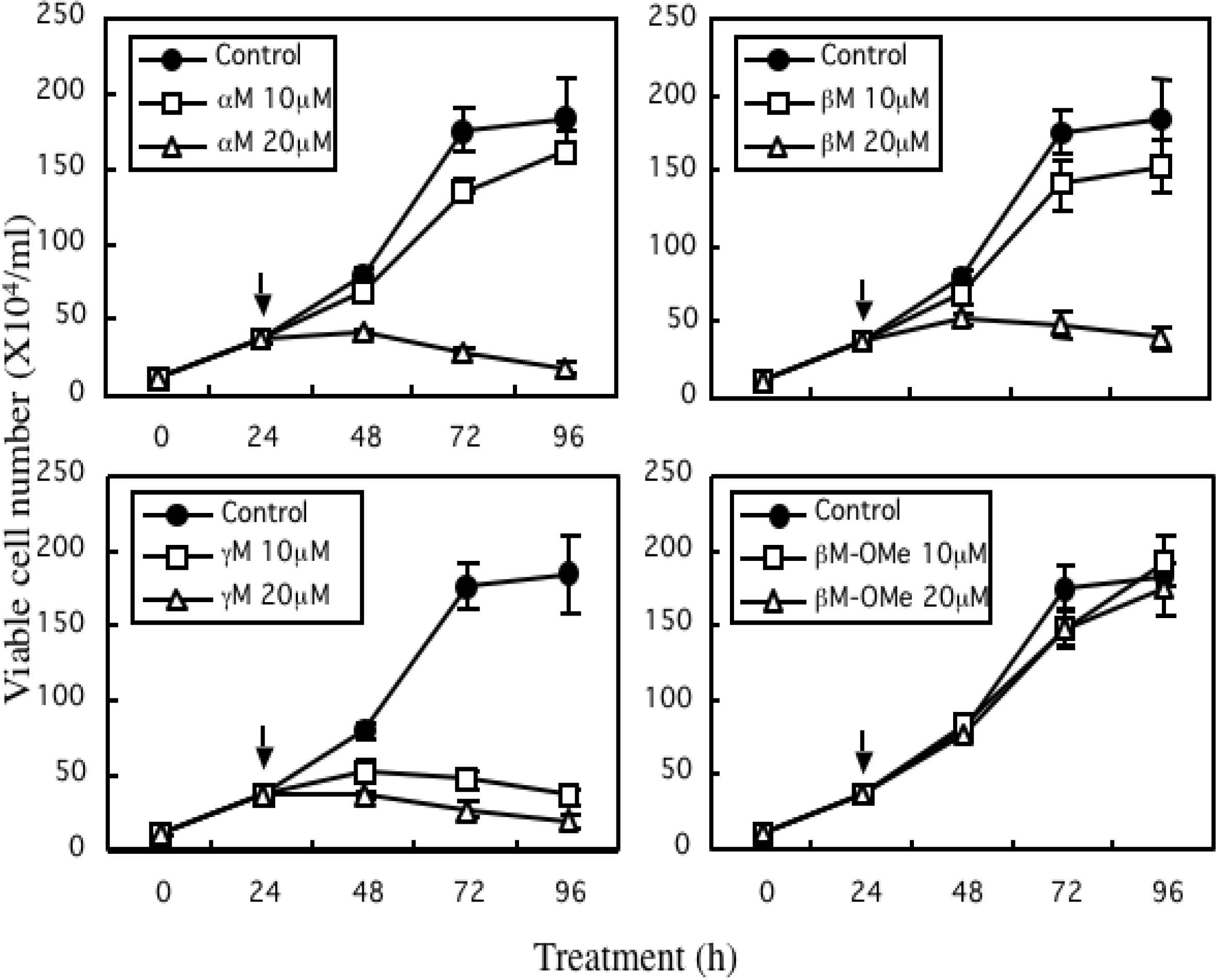
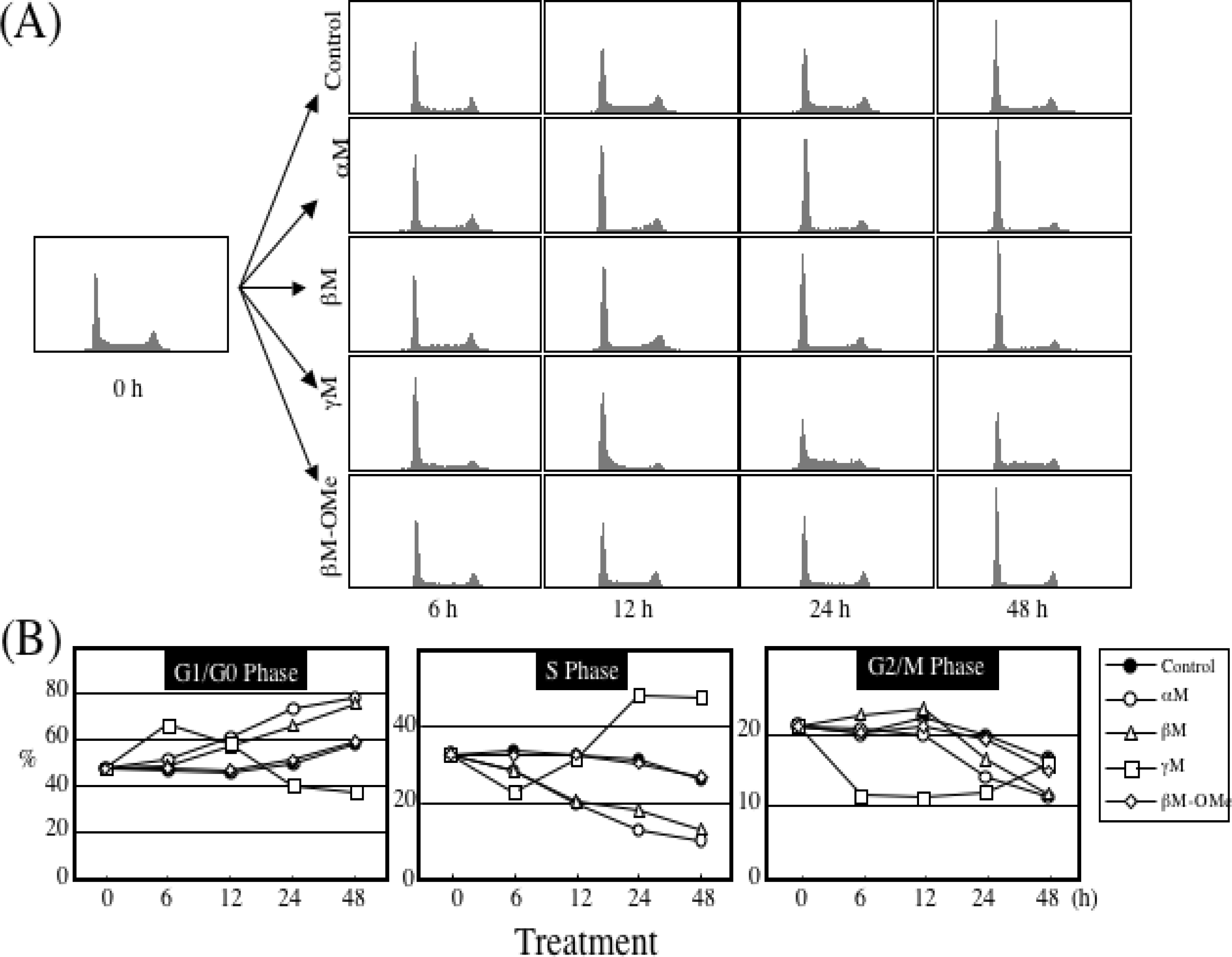
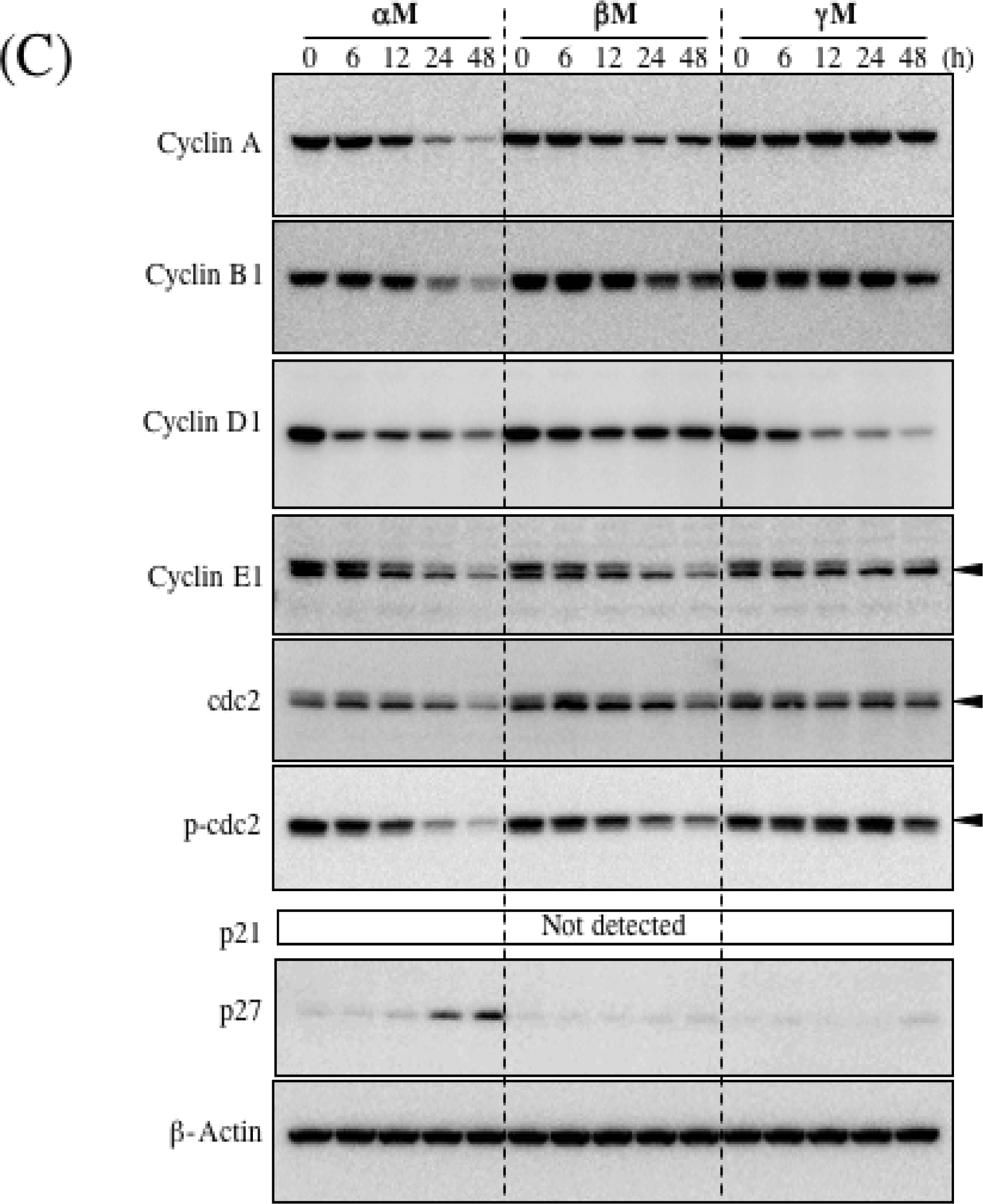
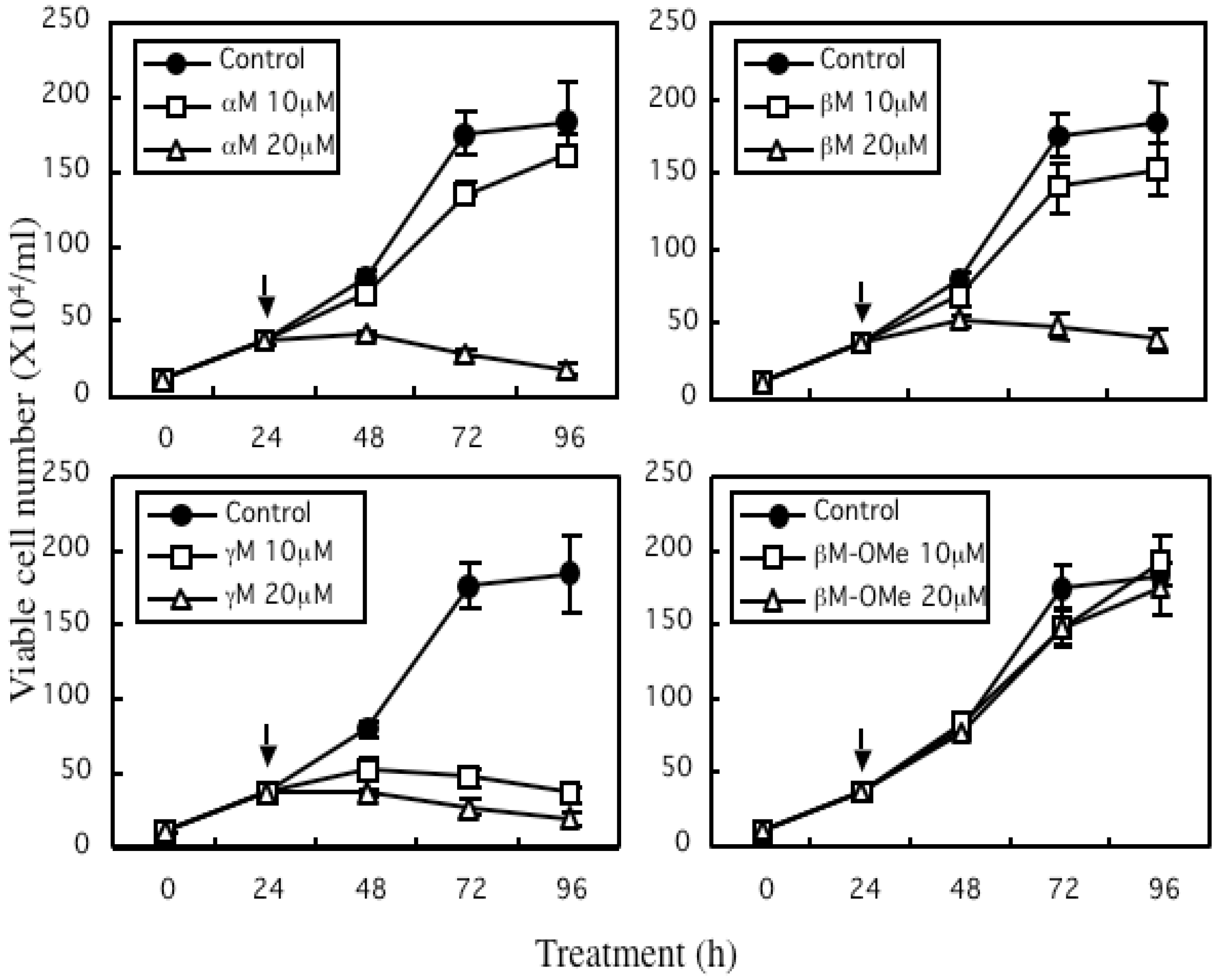
4. Growth Inhibitory Effect of Prenylated Xanthones
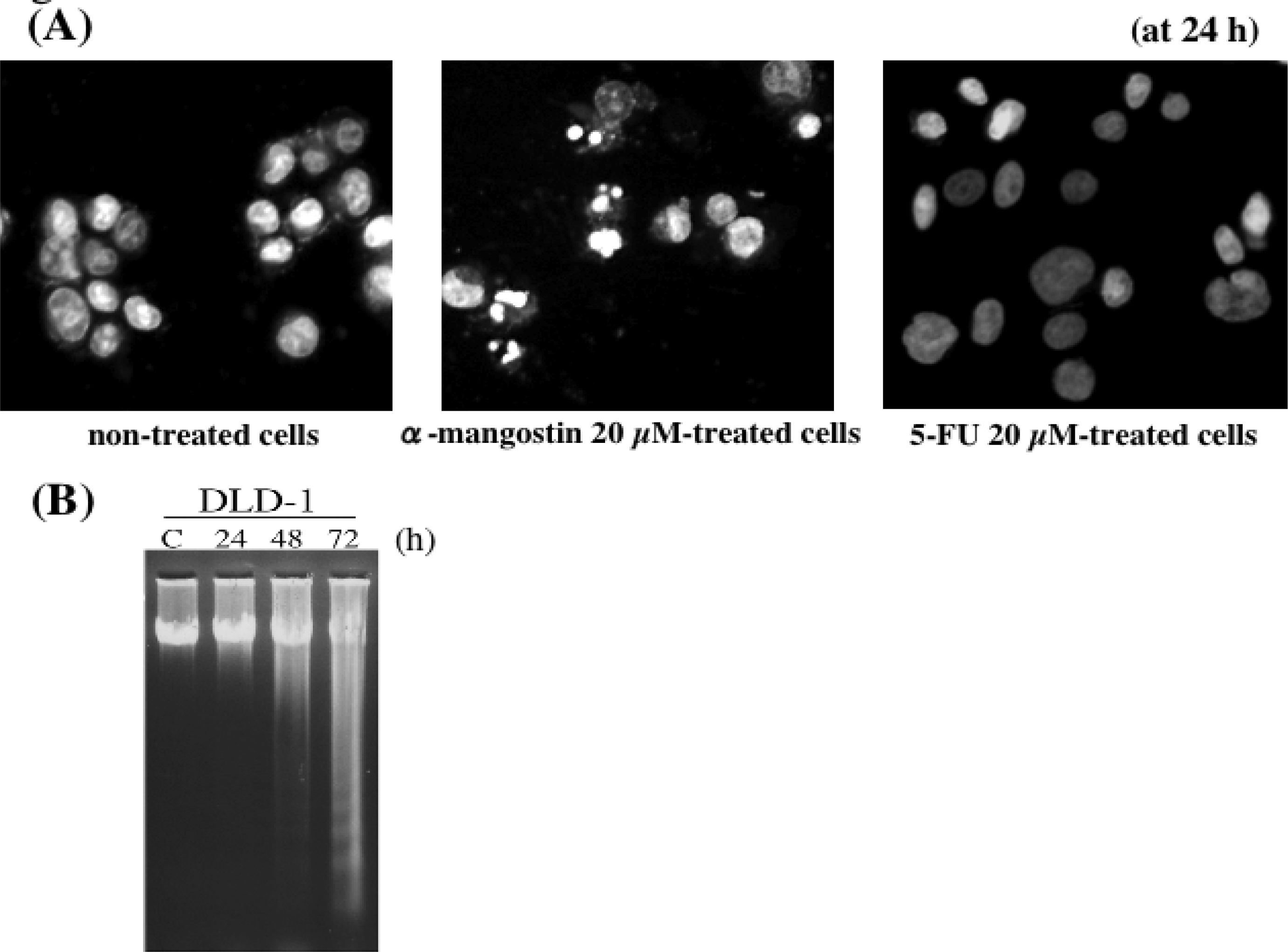
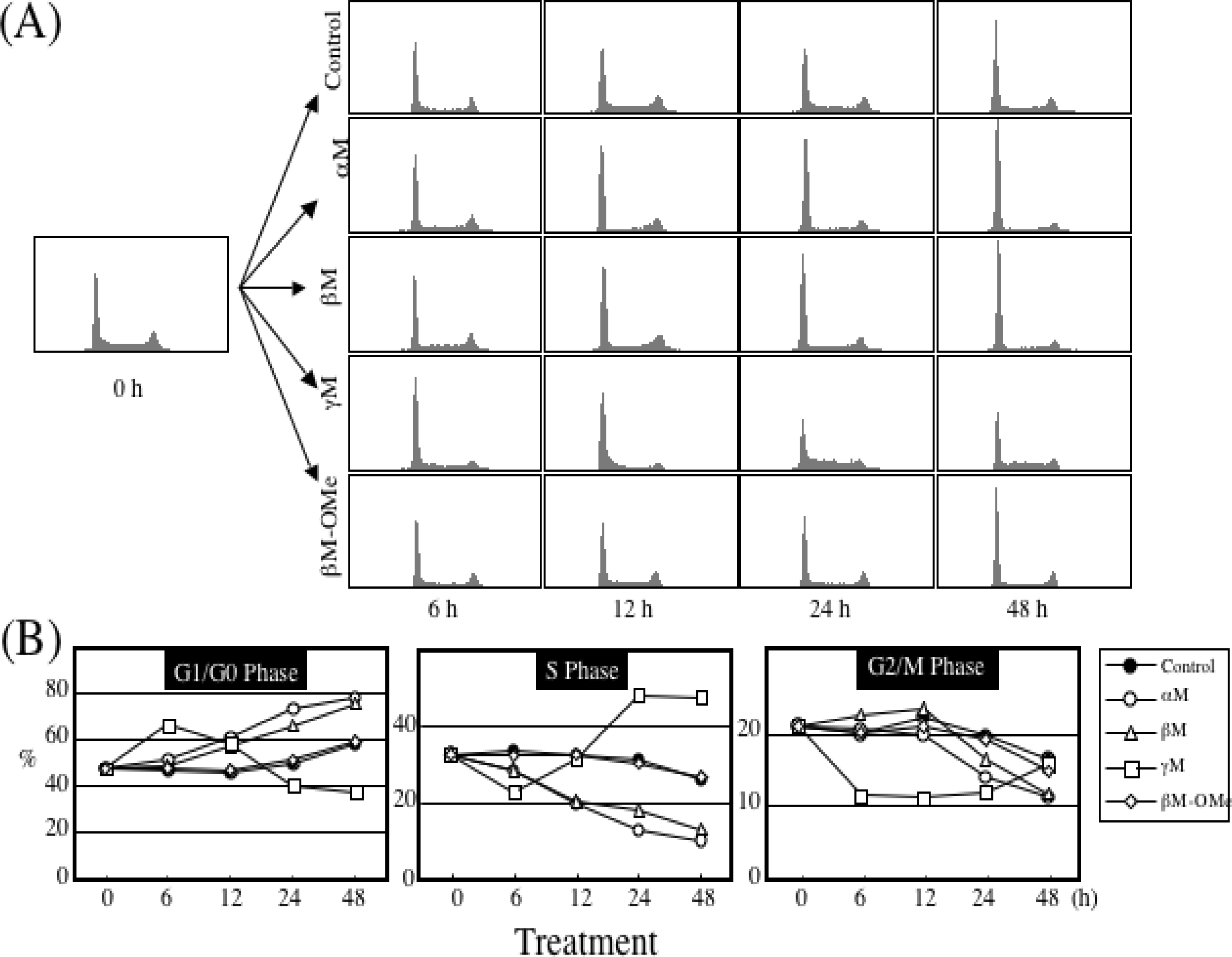
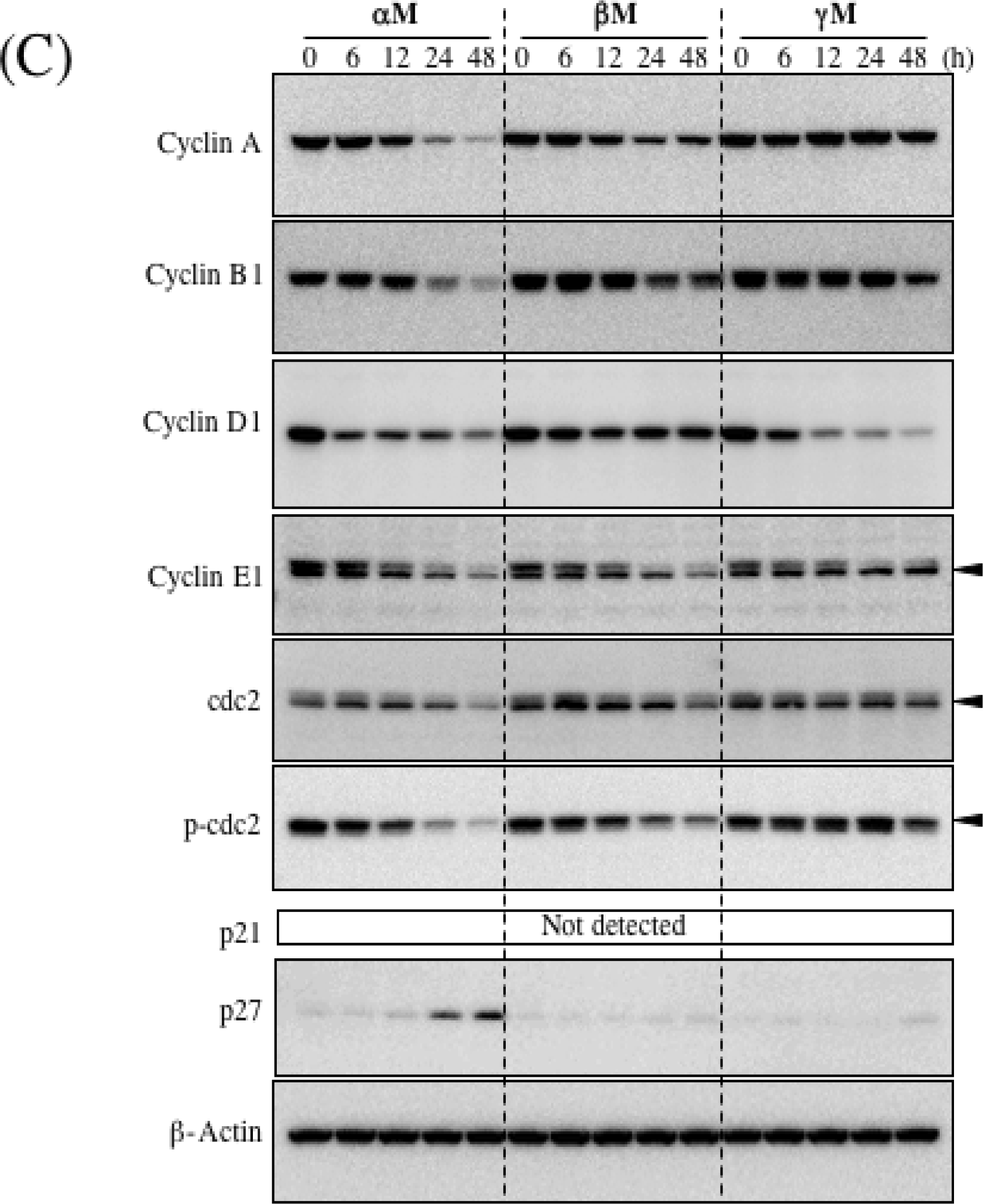
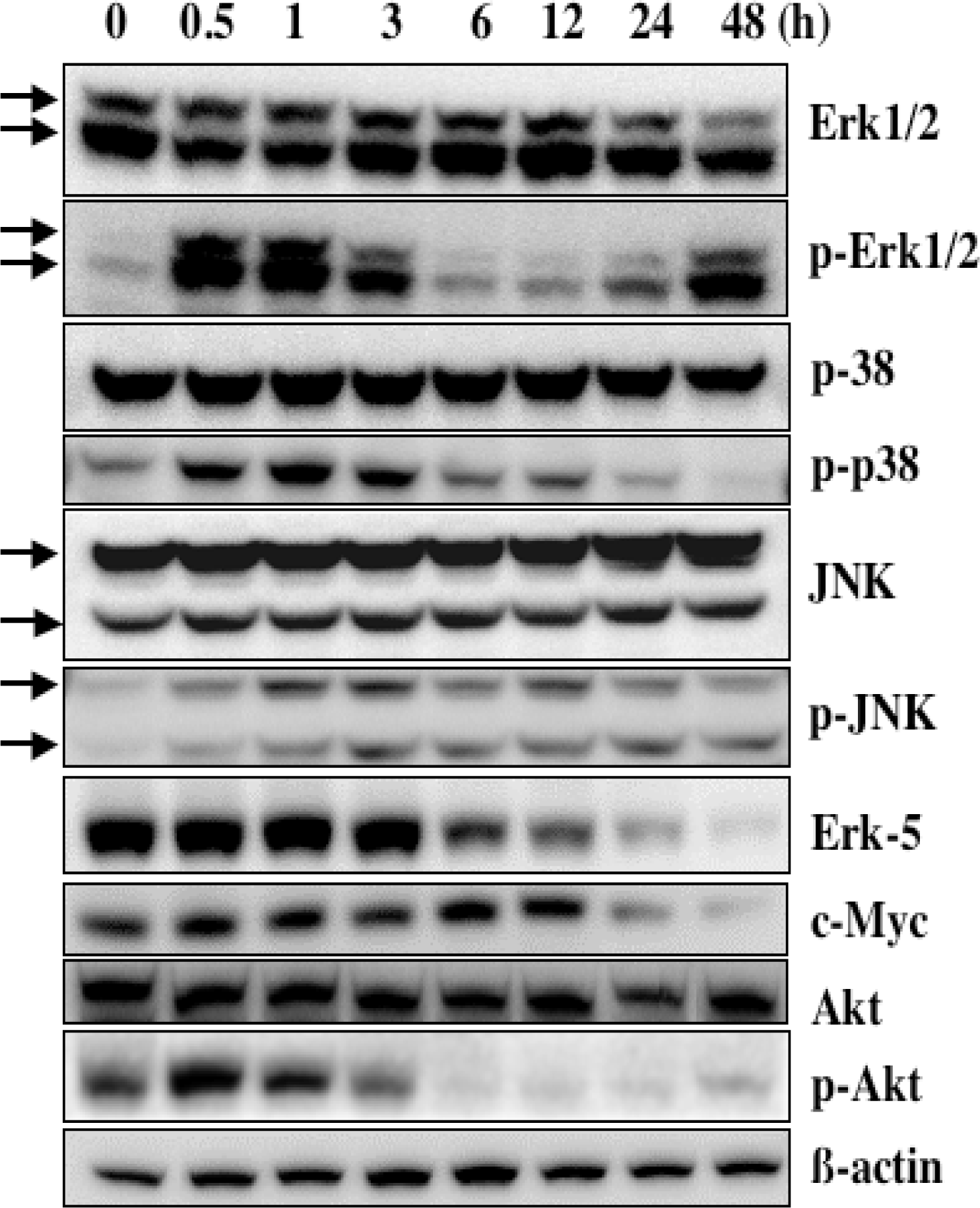
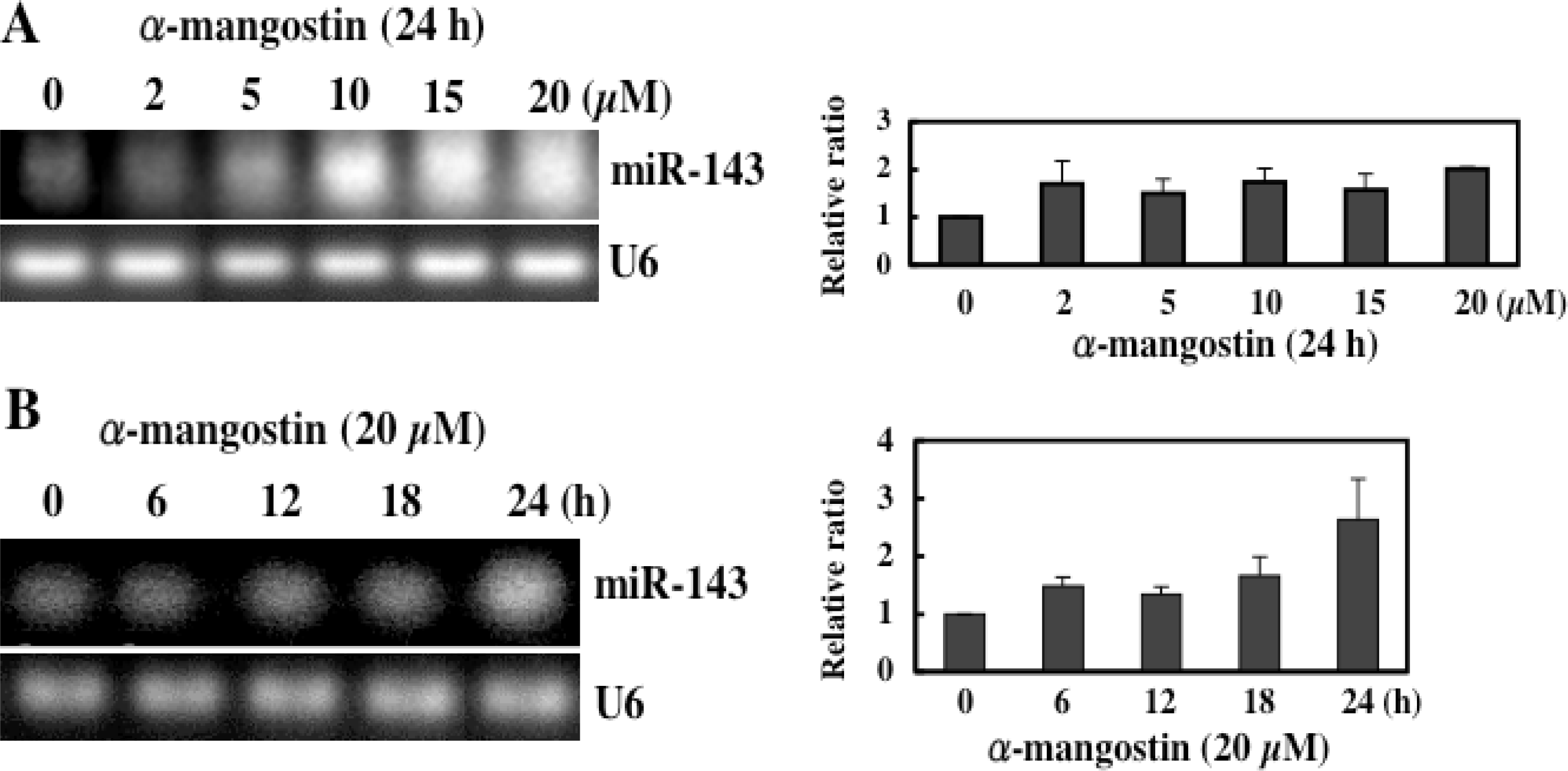
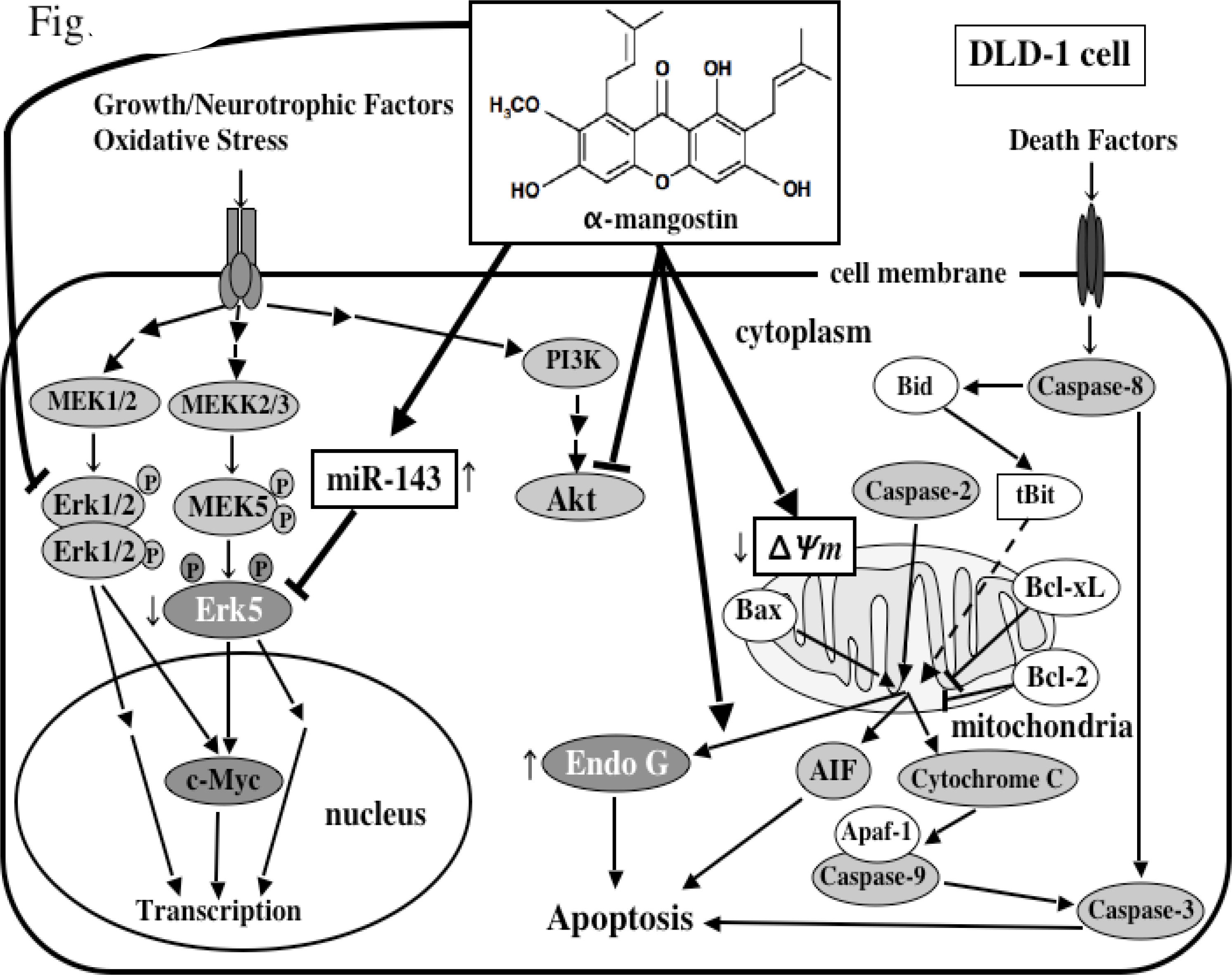
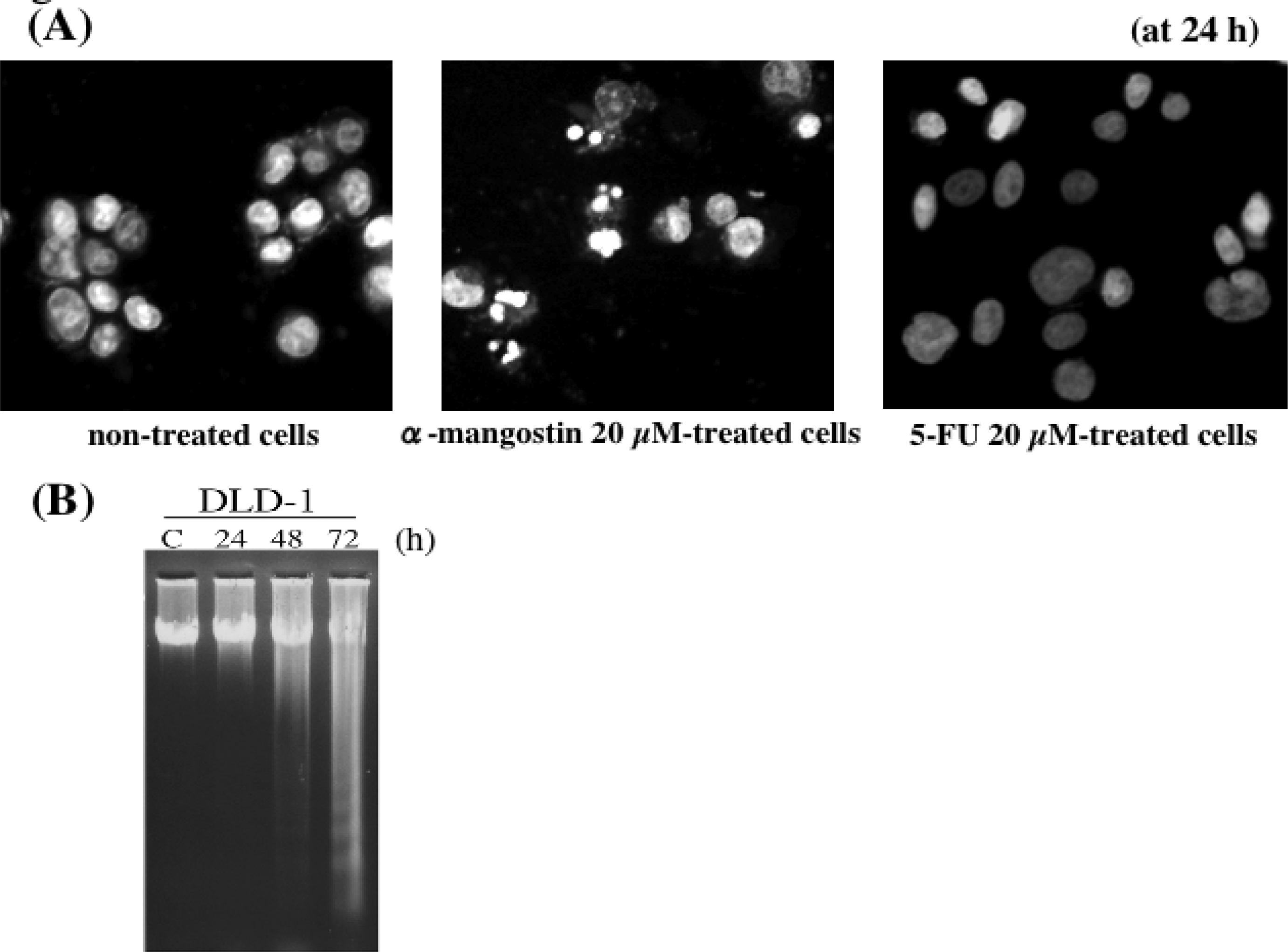
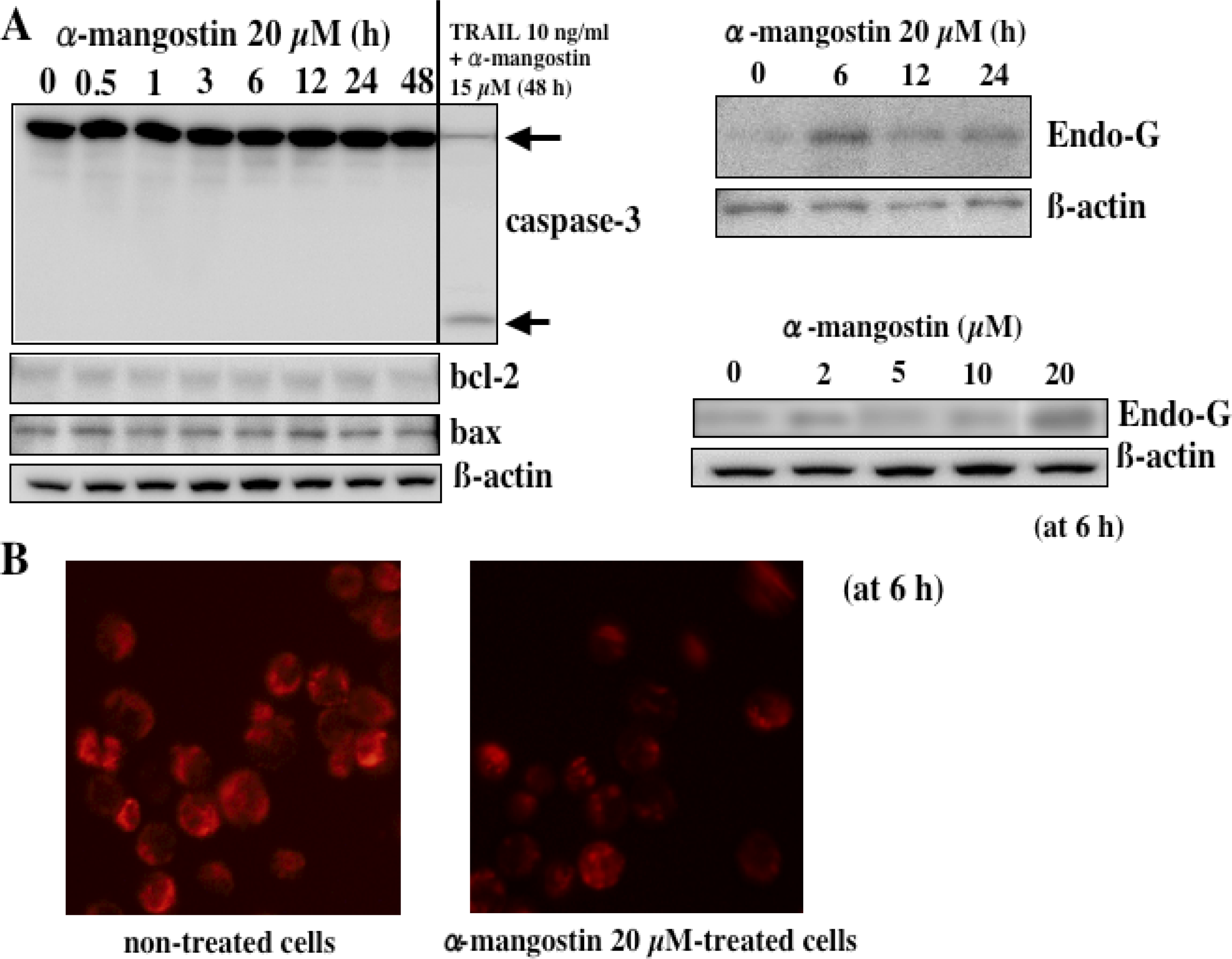
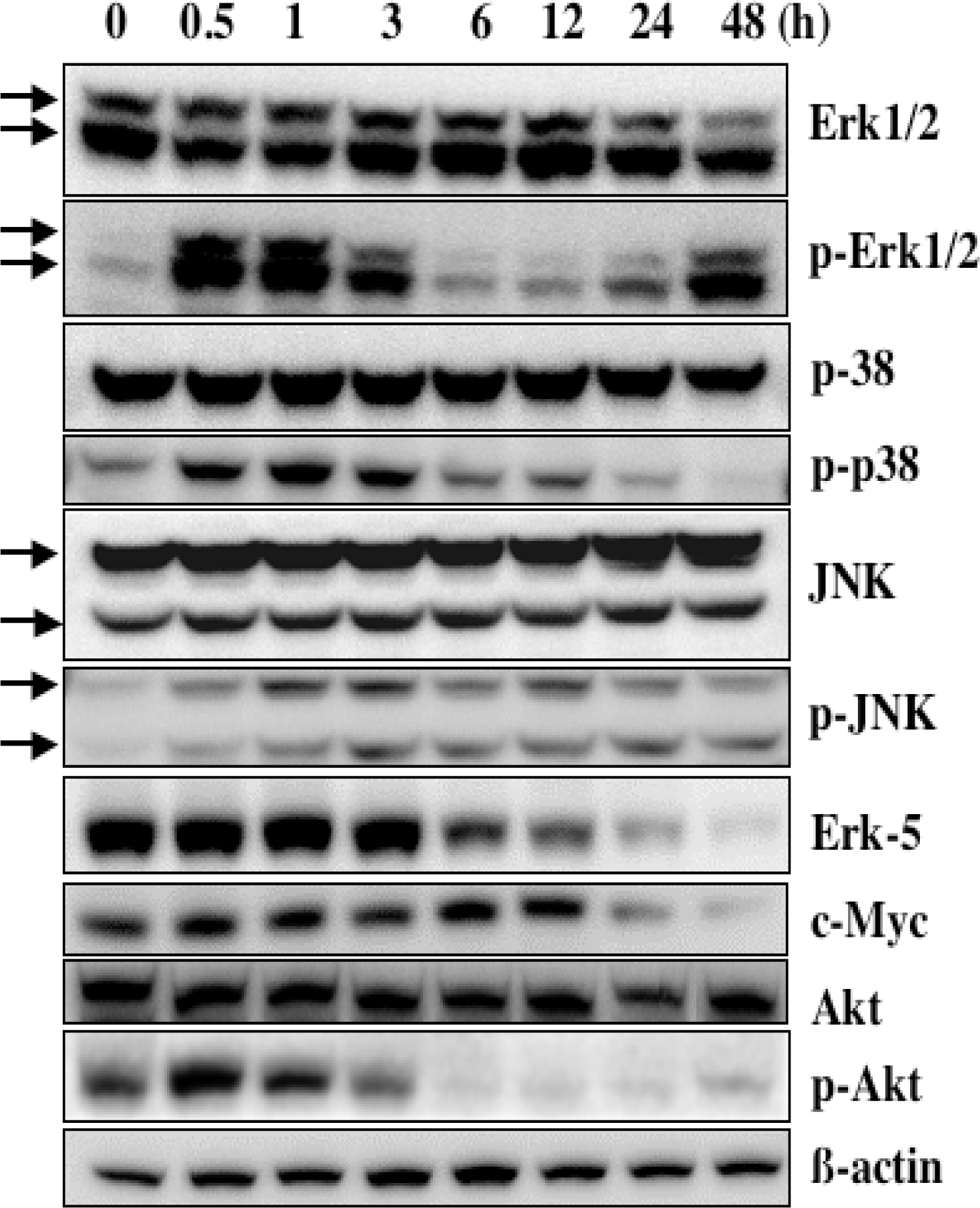
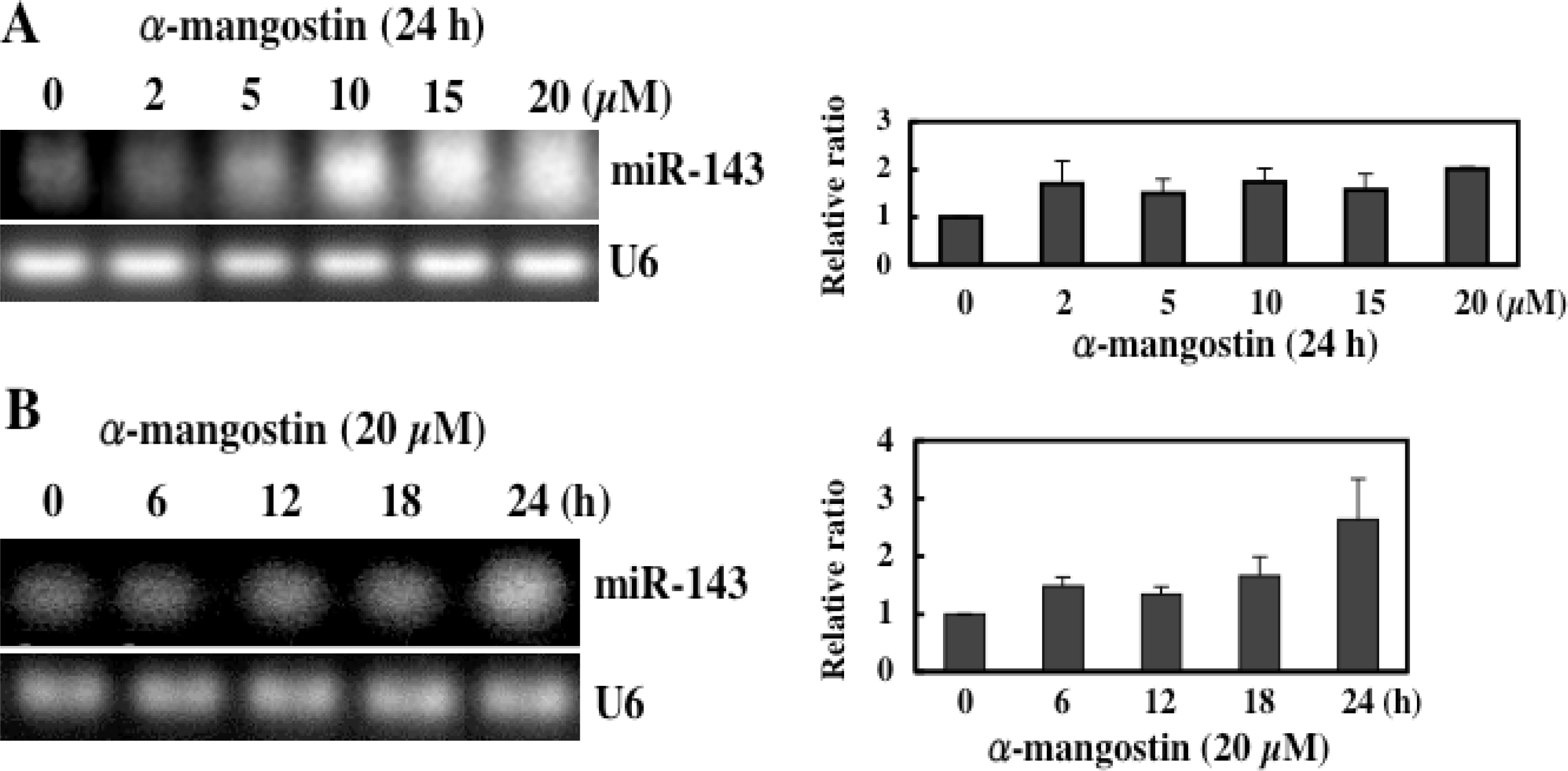
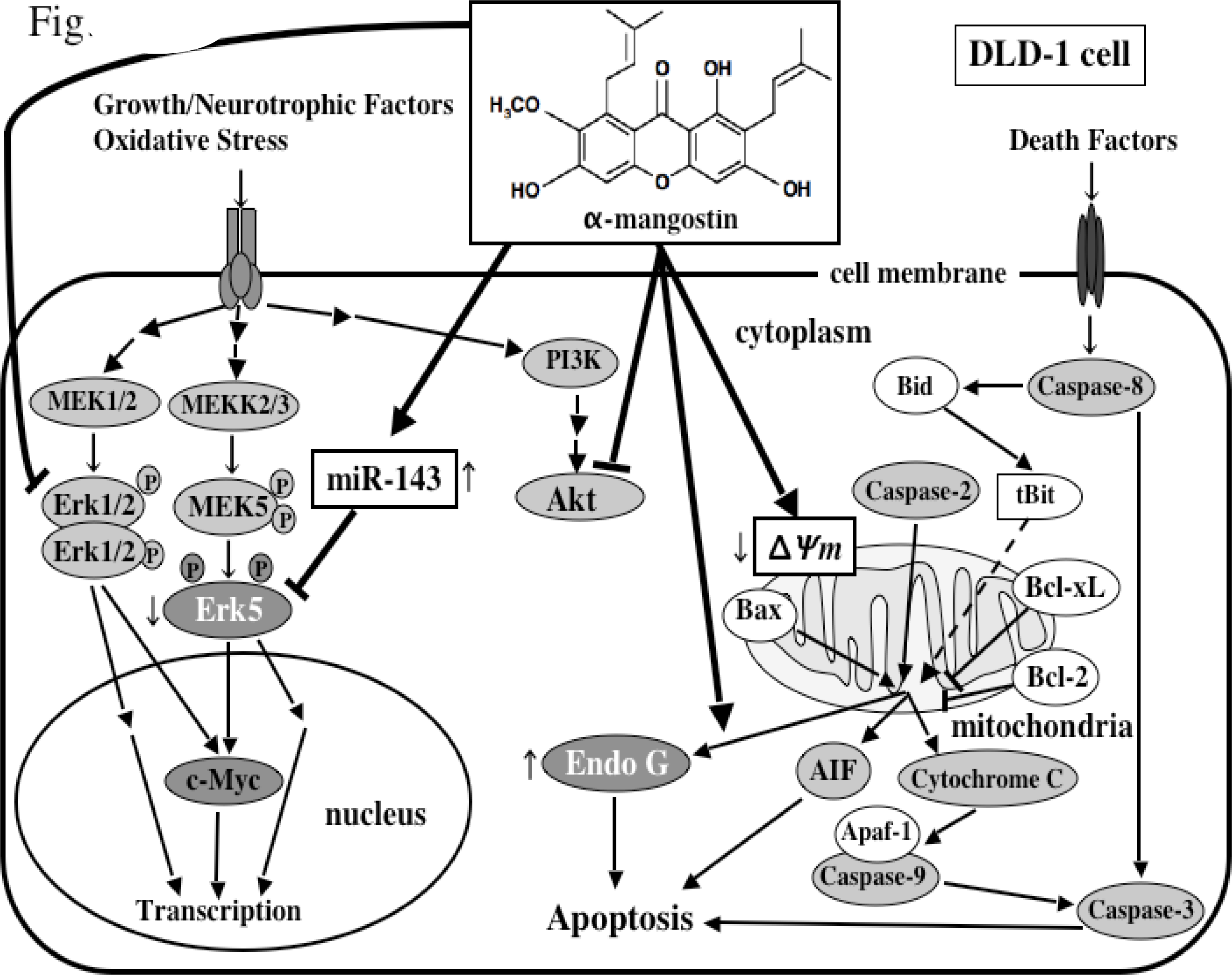
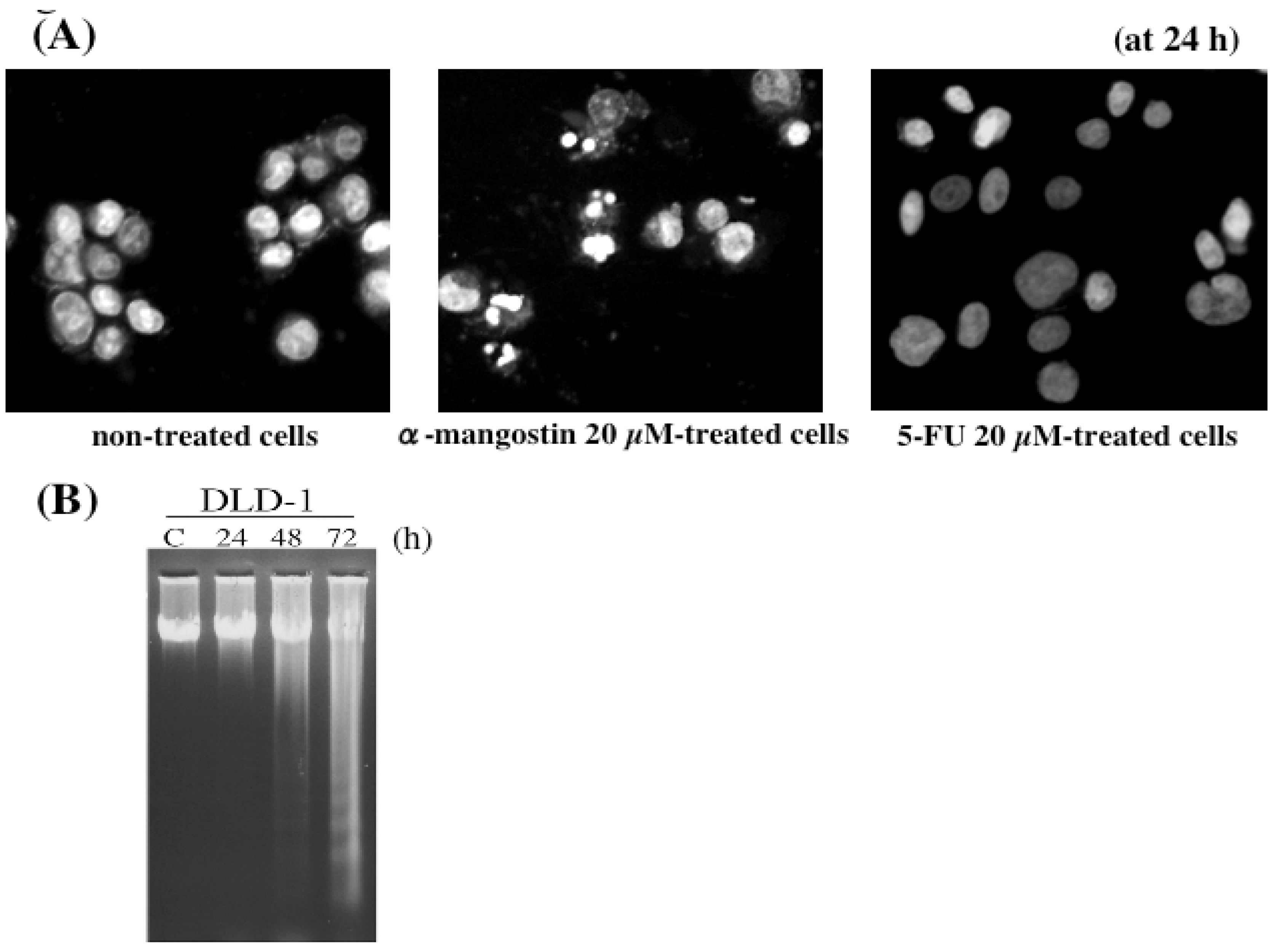
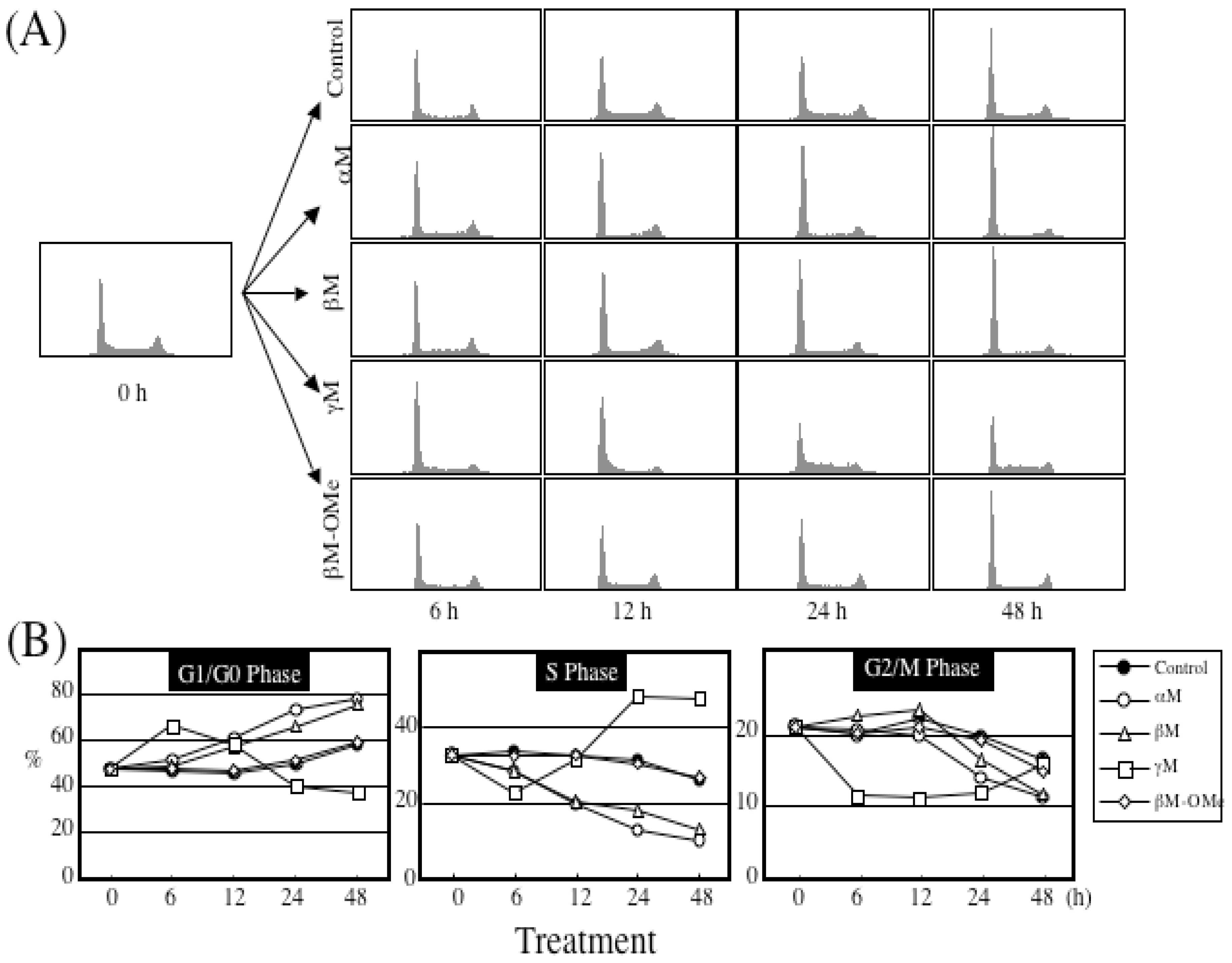
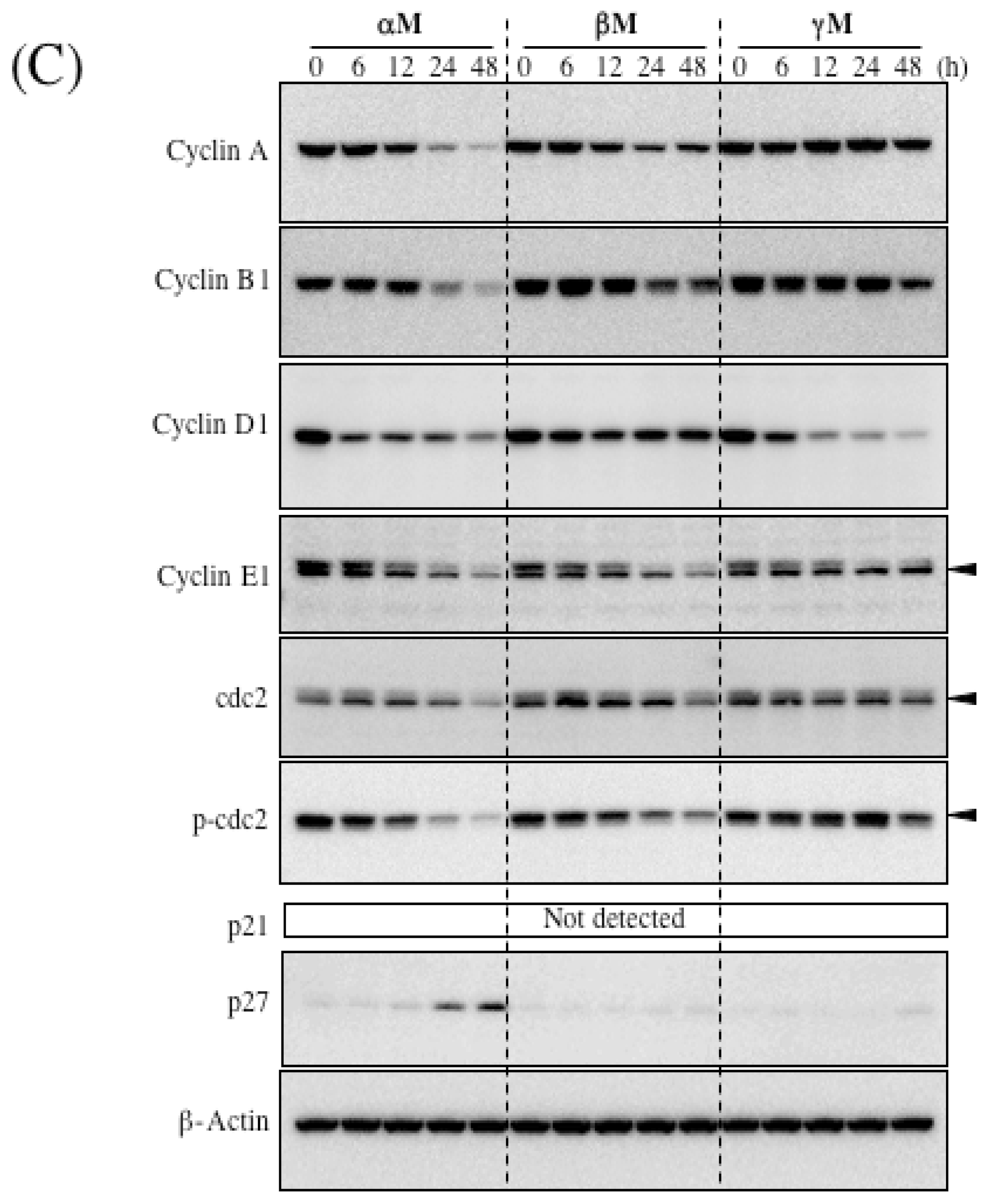
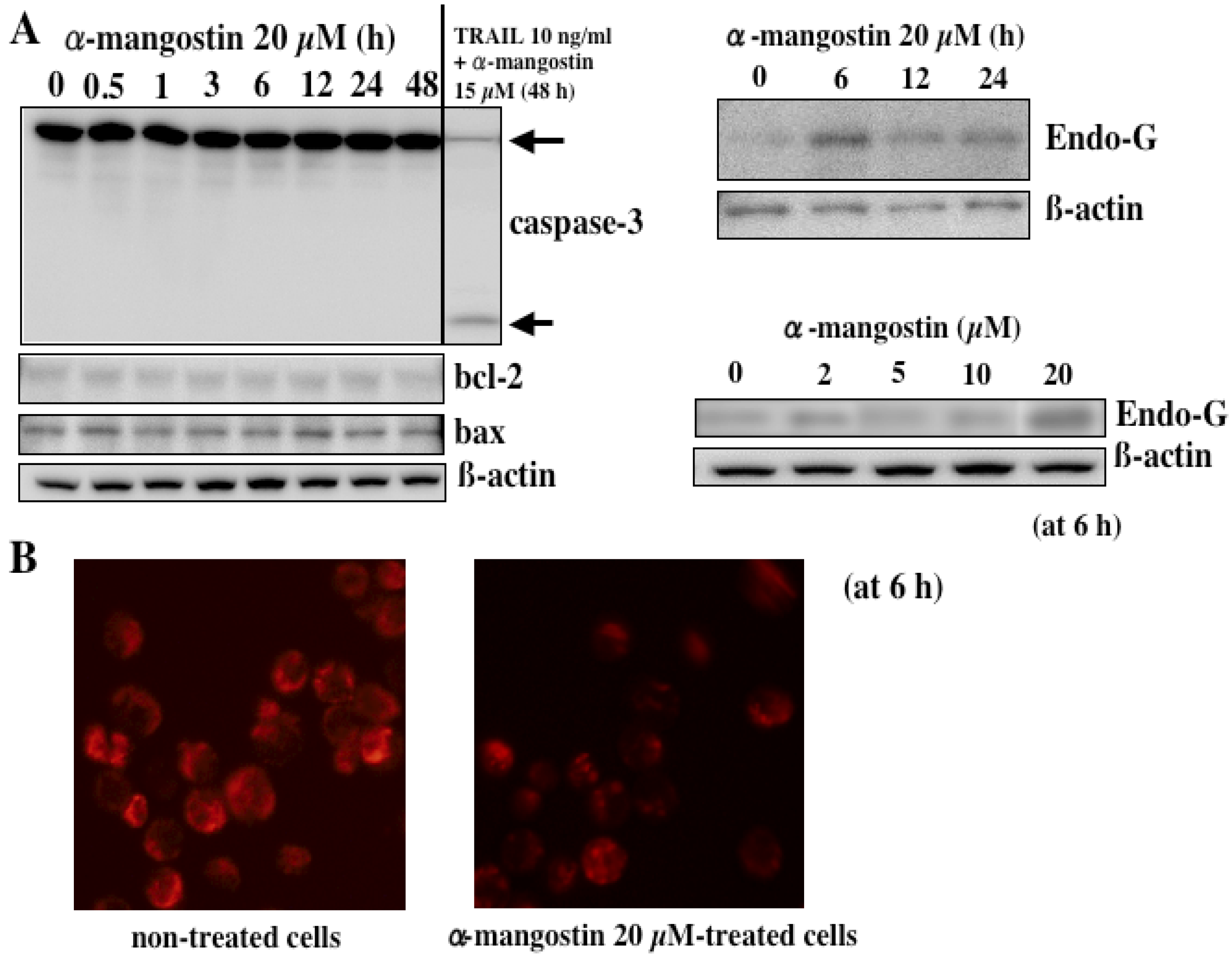
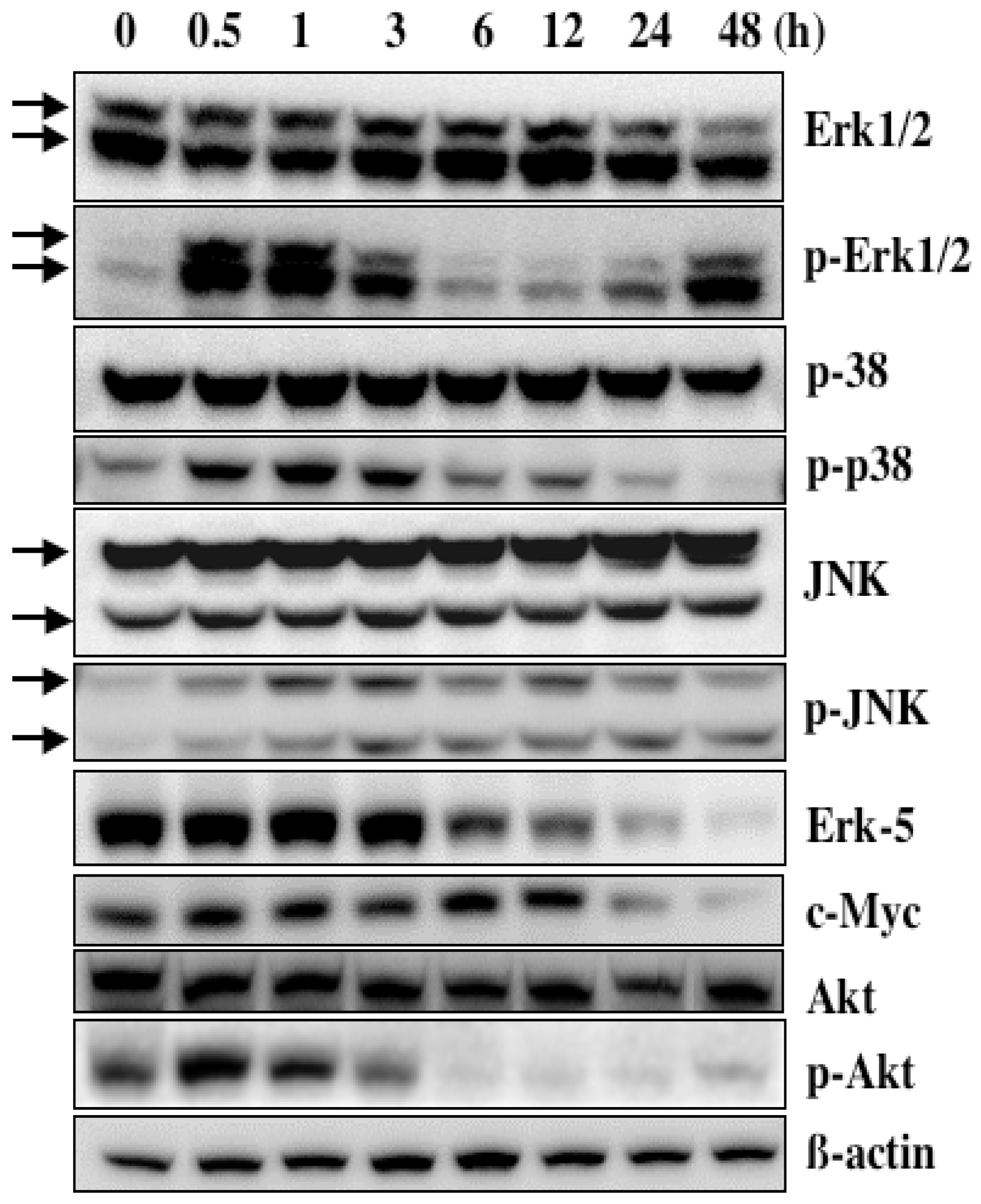
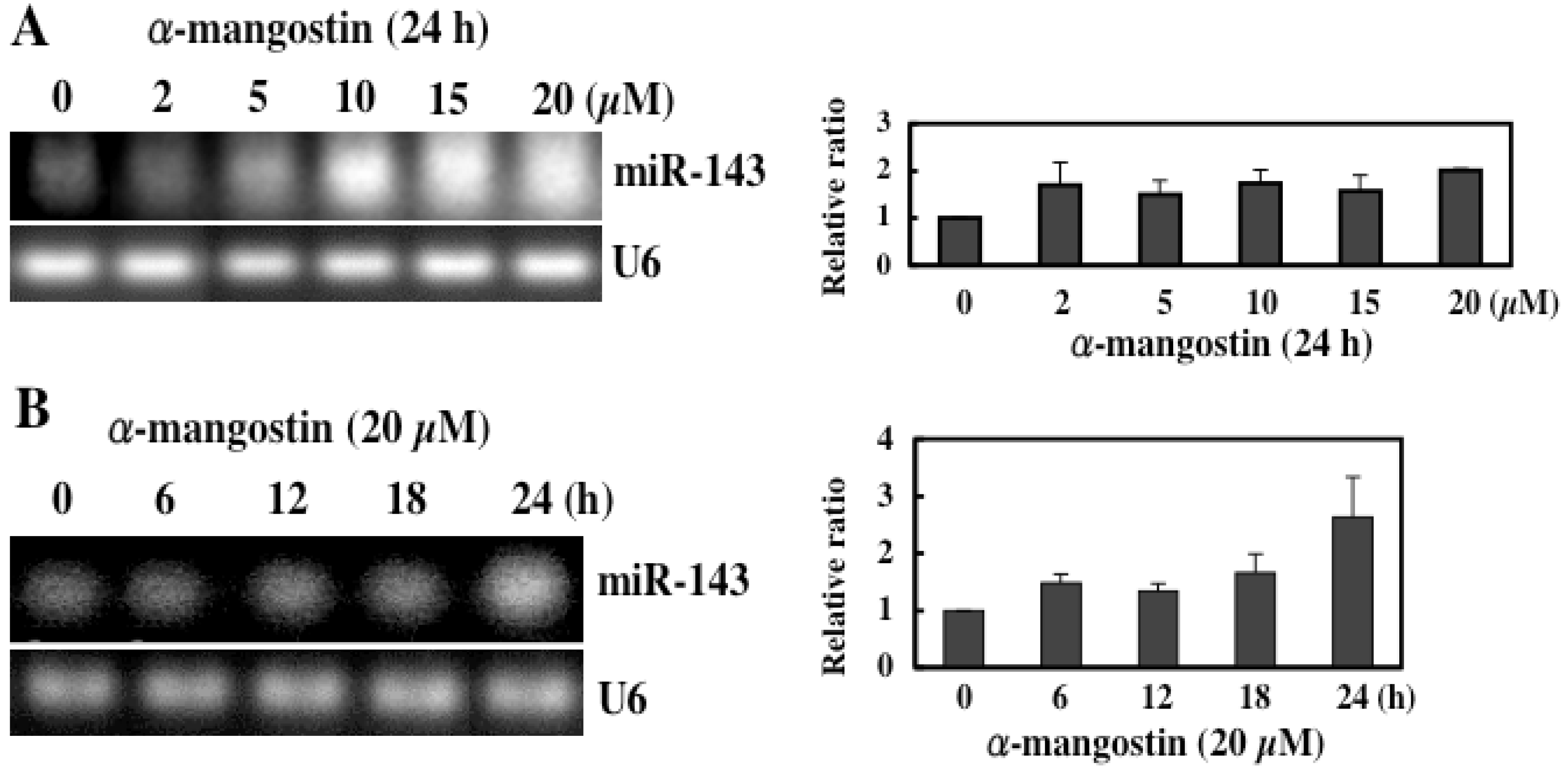
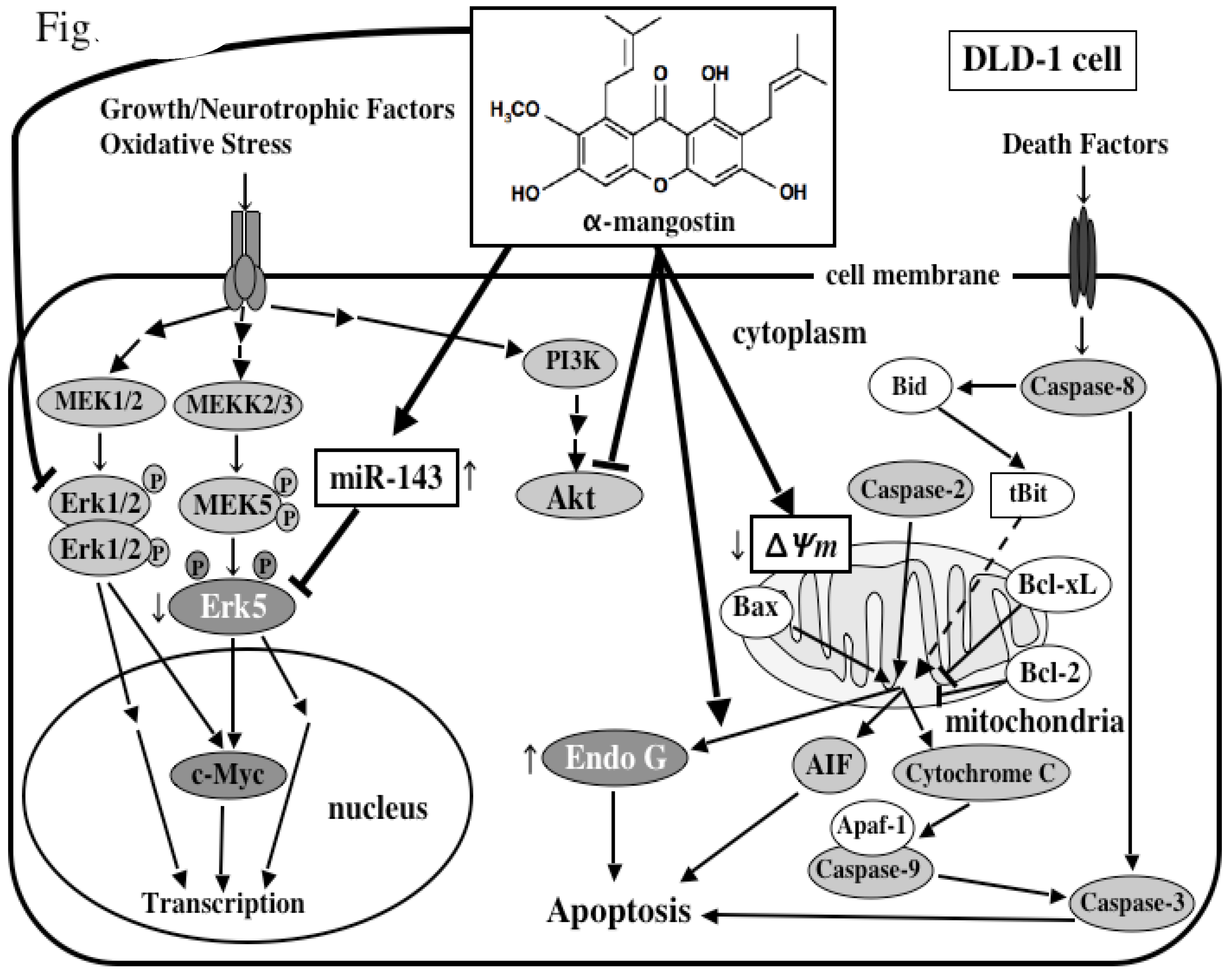
5. Mechanism of α-Mangostin-inducing Apoptosis
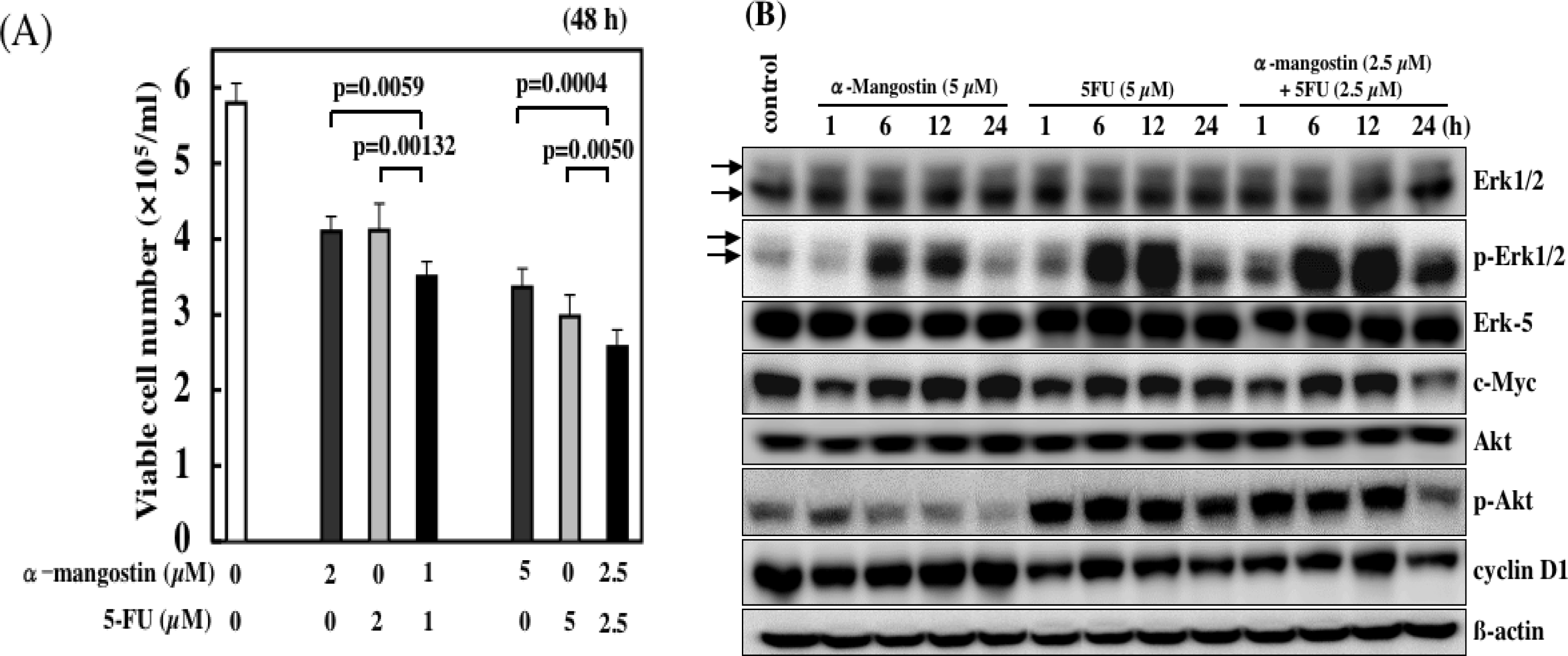
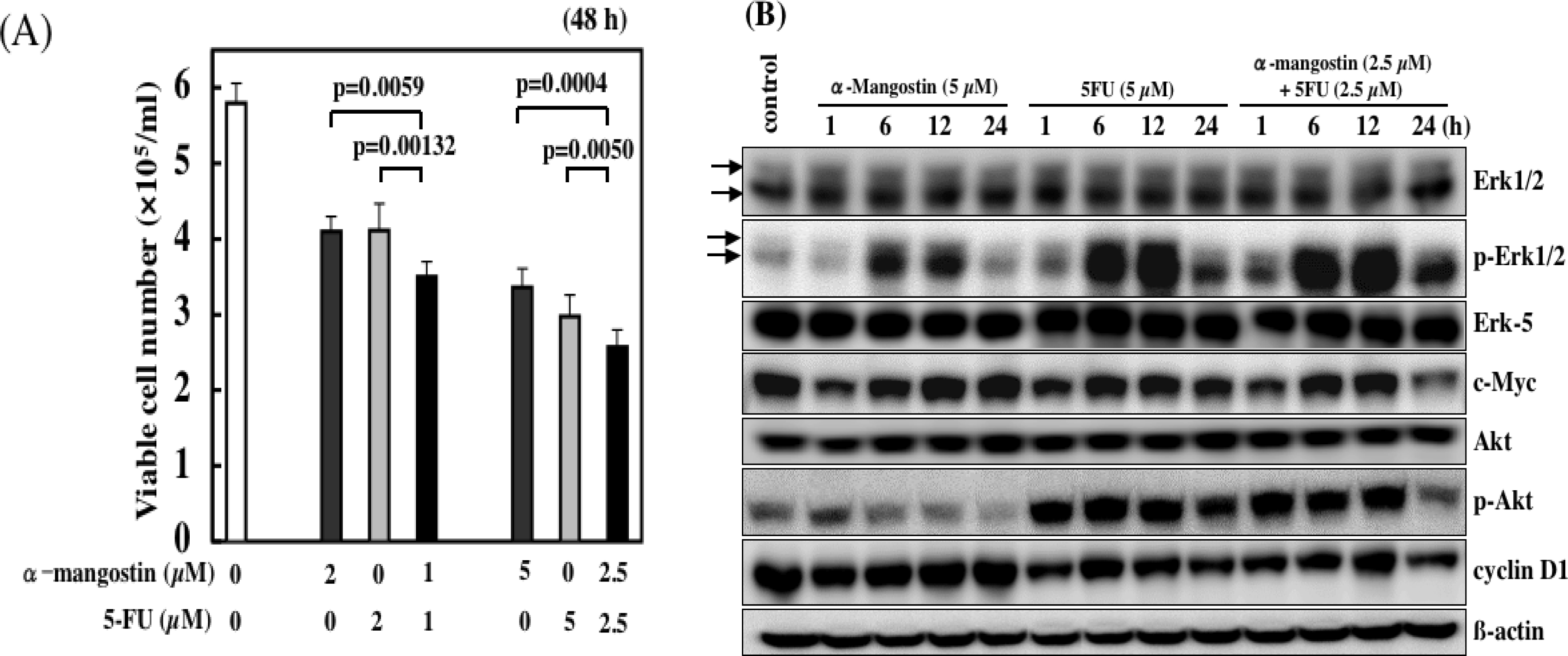
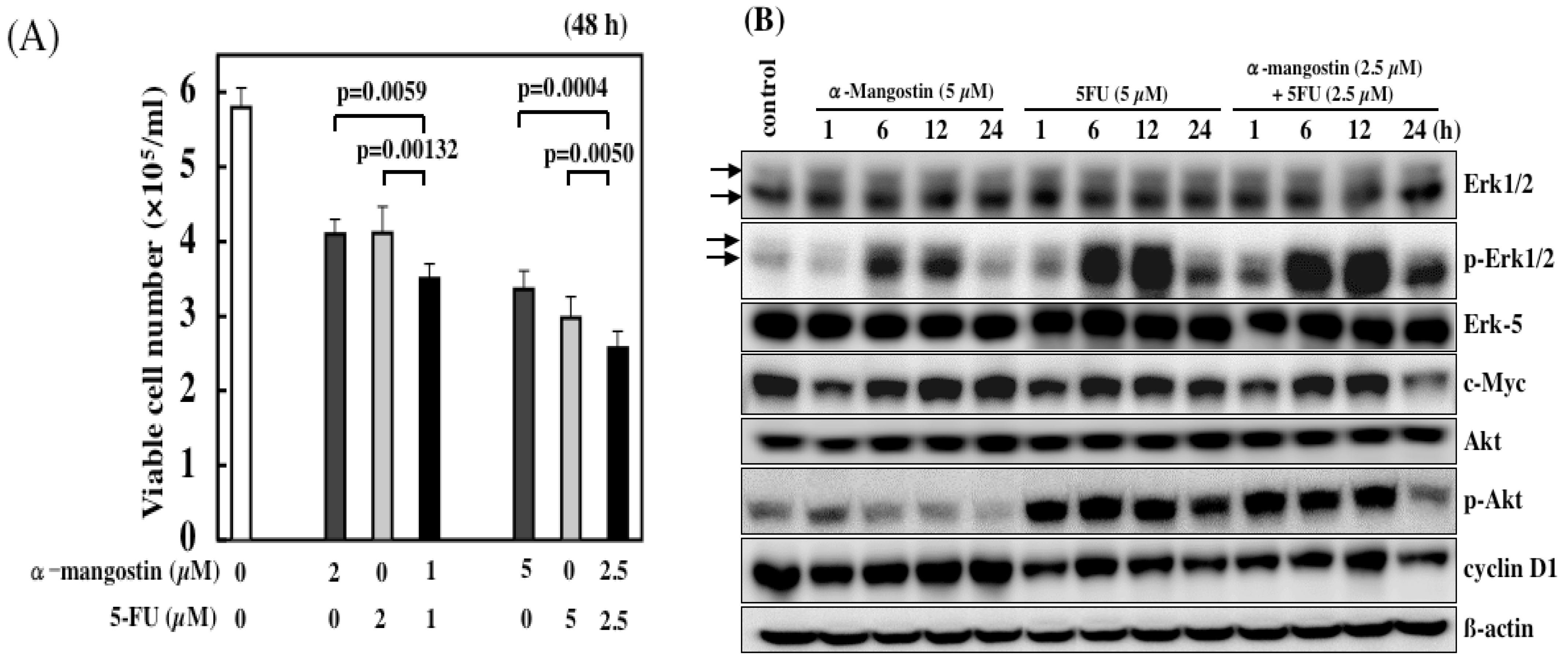
6. Combined Treatment of α-Mangostin with Anticancer Drugs
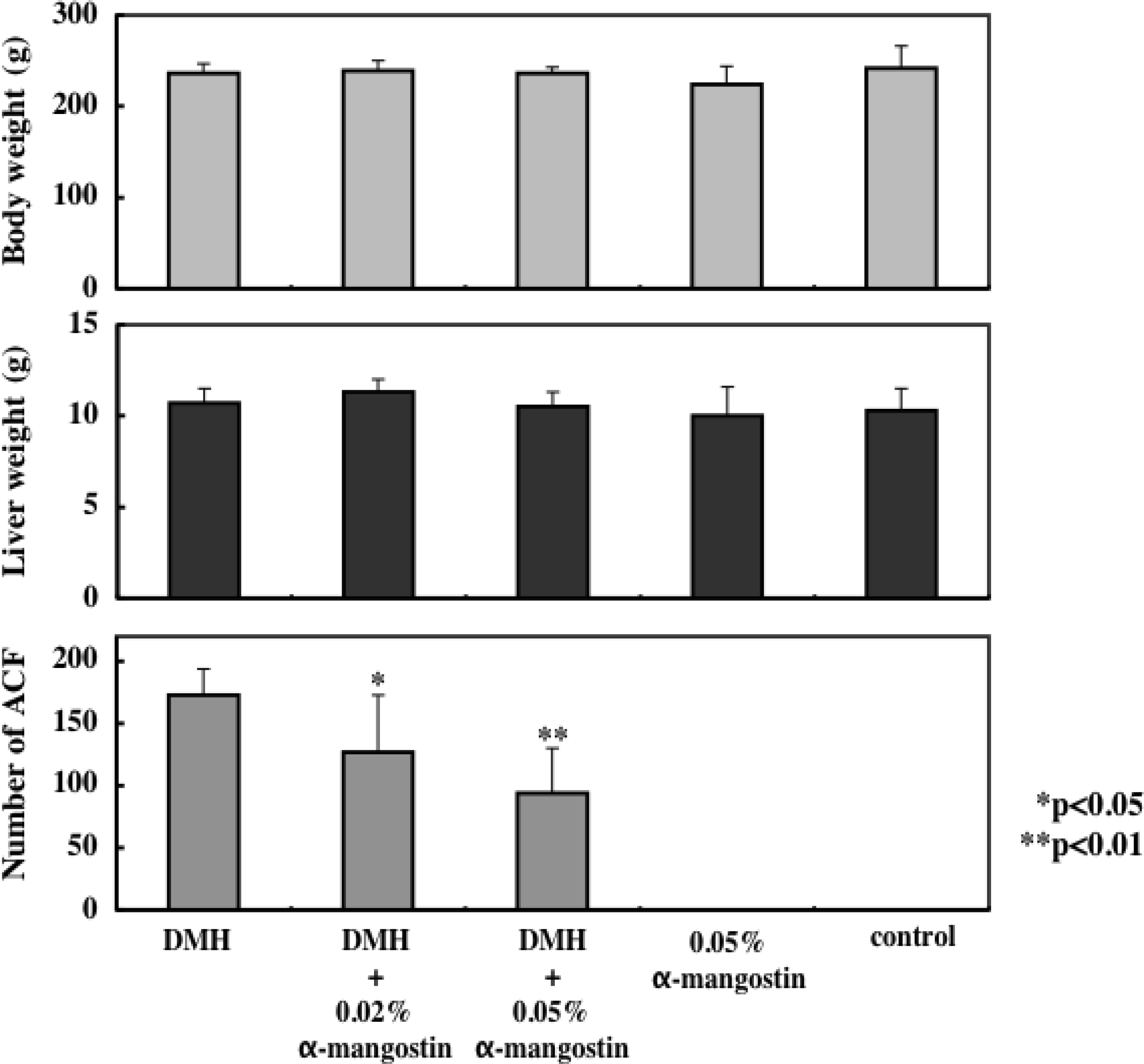
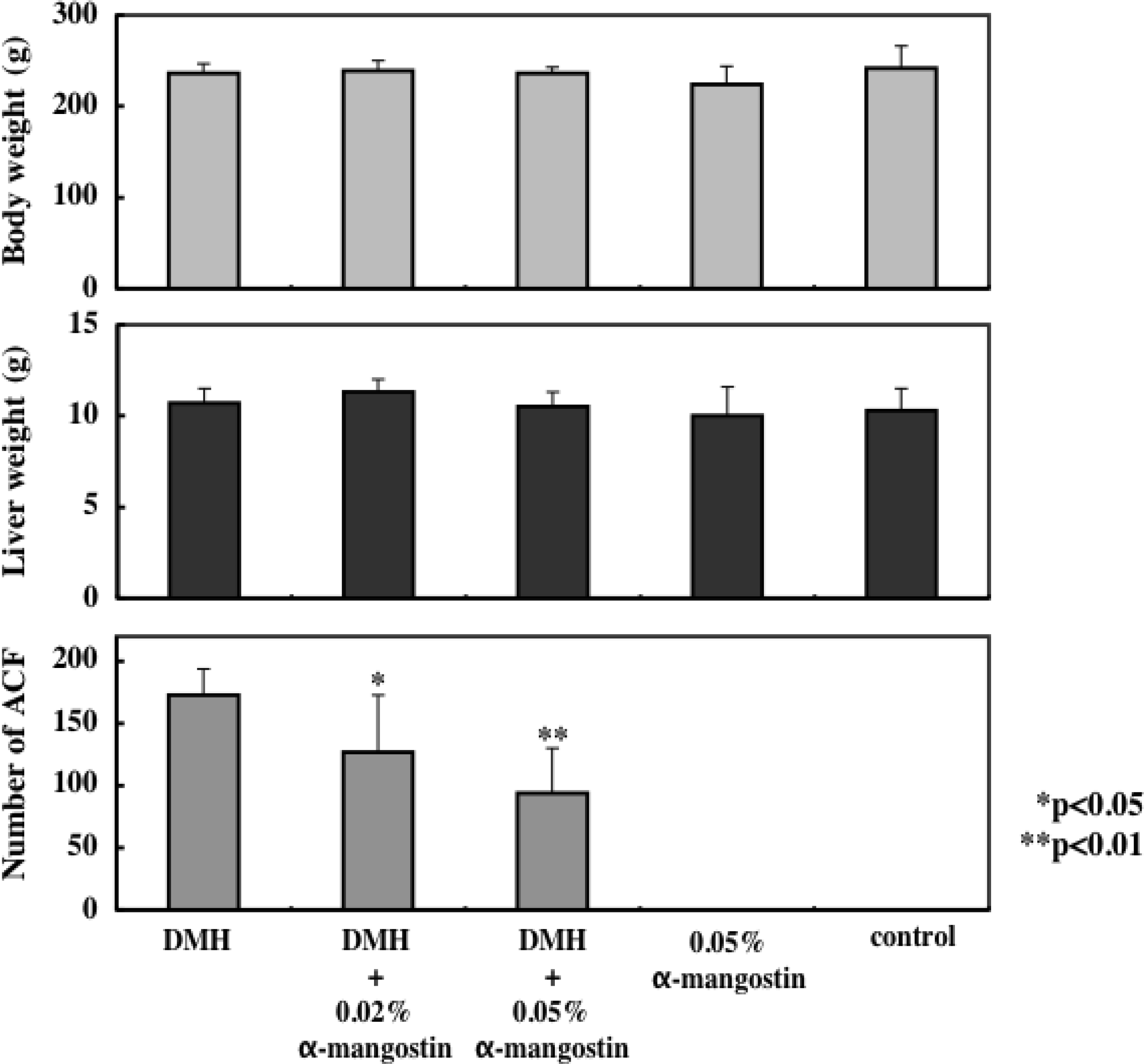
7. Cancer Preventive Effect in vivo
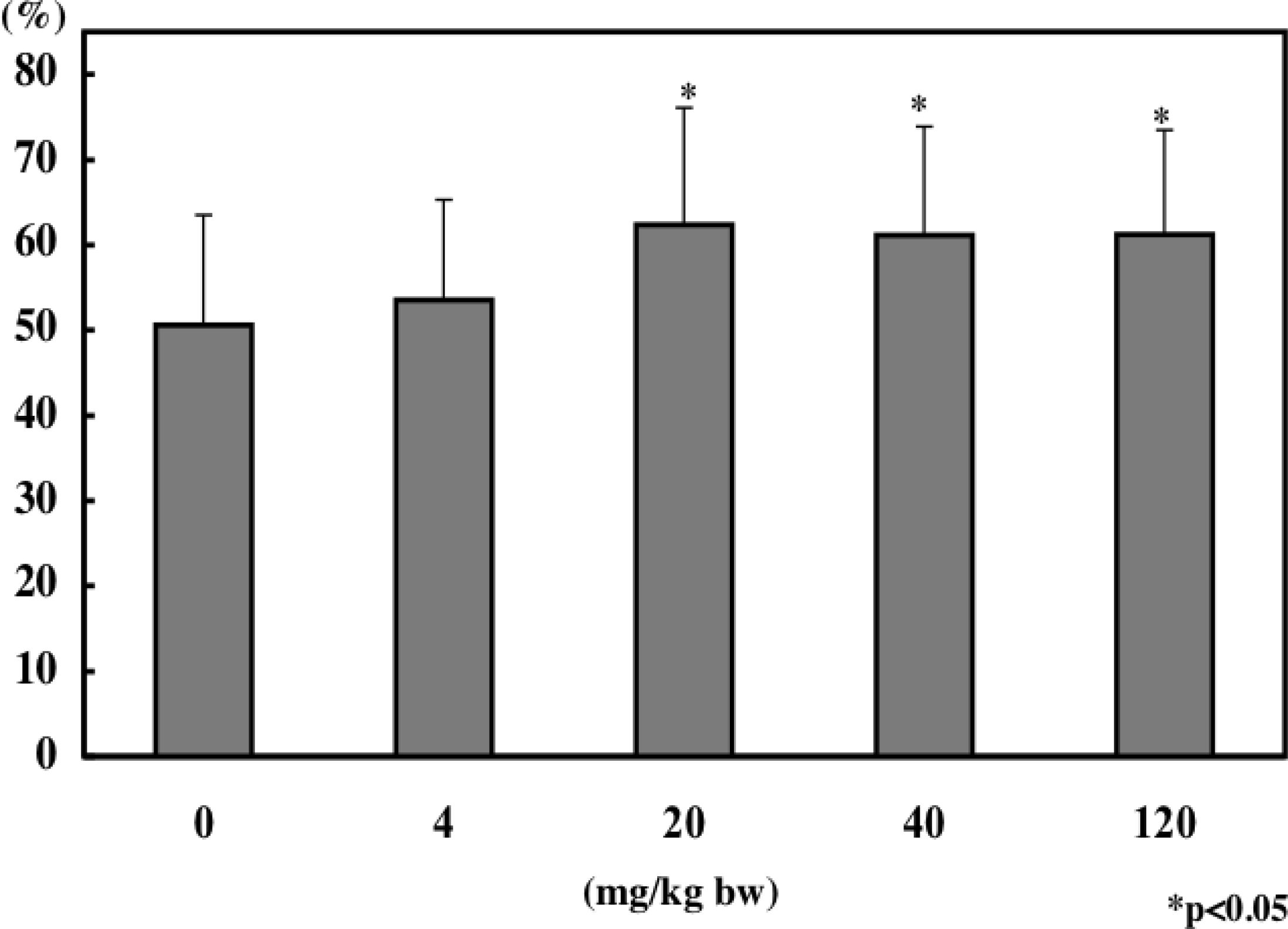
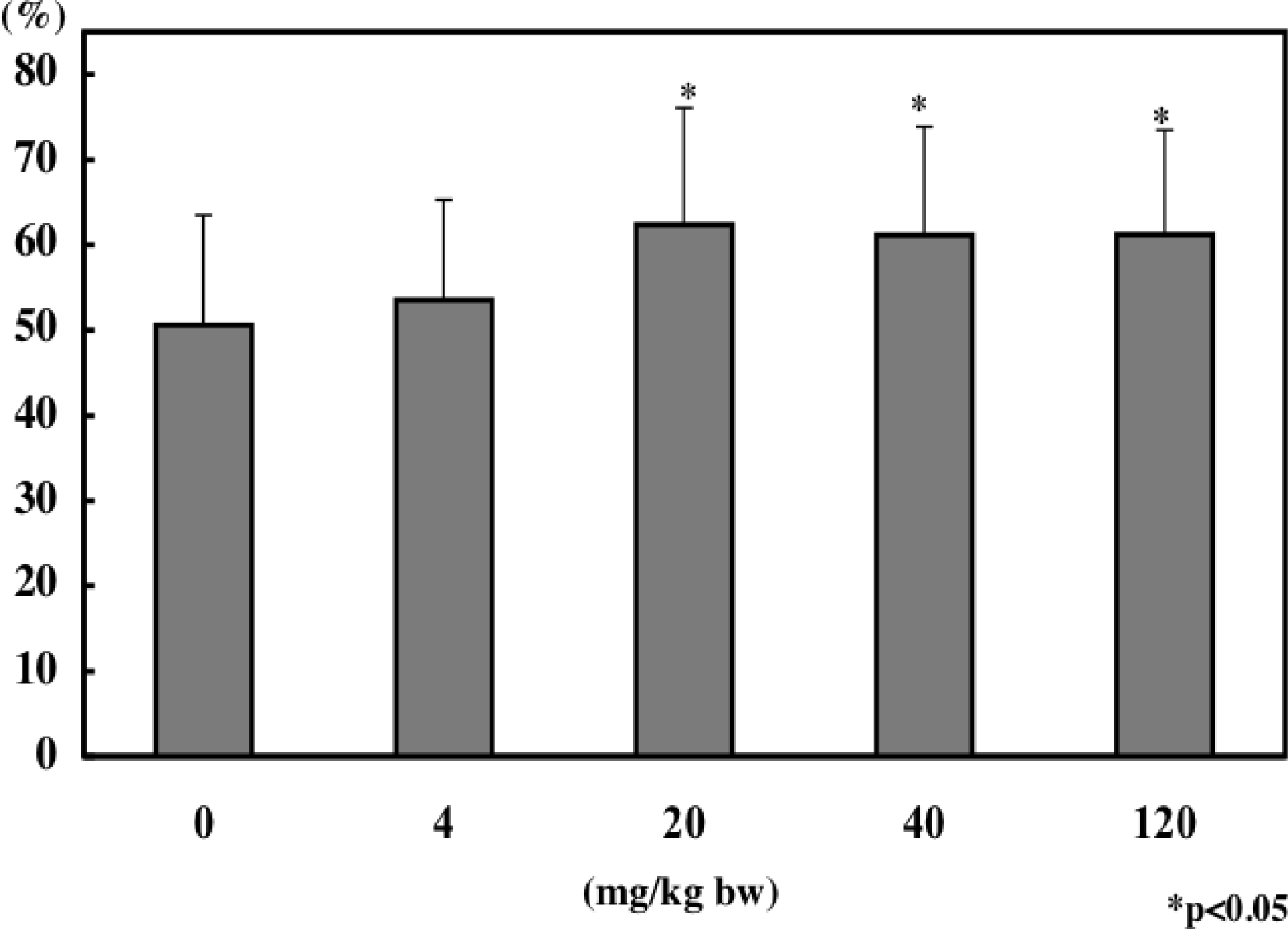
8. Immunomodulatory Effect
9. Conclusion and Future Perspectives
Acknowledgments
References and Notes
- Sun, J; Chu, Y-F; Wu, X; Liu, RH. Antioxidant and antiproliferative activities of common fruits. J. Agric. Food Chem. 2002, 50, 7449–7454. [Google Scholar]
- Chu, YF; Sun, J; Wu, X; Liu, RH. Antioxidant and antiproliferative activities of common vegetables. J. Agric. Food Chem. 2002, 50, 6910–6916. [Google Scholar]
- Block, G; Patterson, B; Subar, A. Fruit, vegetables, and cancer prevention: a review of the epidemiological evidence. Nutr. Cancer 1992, 18, 1–29. [Google Scholar]
- Temple, NJ; Gladwin, KK. Fruit, vegetables, and the prevention of cancer: research challenges. Nutrition 2003, 19, 467–470. [Google Scholar]
- Ames, BN; Gold, LS. Endogenous mutagens and the causes of aging and cancer. Mutat. Res. 1991, 250, 3–16. [Google Scholar]
- Liu, RH; Hotchkiss, JH. Potential genotoxicity of chronically elevated nitric oxide: a review. Mutat. Res. 1995, 339, 73–89. [Google Scholar]
- Ikai, T; Akao, Y; Nakagawa, Y; Ohguchi, K; Sakai, Y; Nozawa, Y. Magnolol-induced apoptosis is mediated via the intrinsic pathway with release of AIF from mitochondria in U937 cells. Biol. Pharm. Bull. 2006, 29, 2498–2501. [Google Scholar]
- Matsumoto, K; Akao, Y; Ohguchi, K; Ito, T; Tanaka, T; Iinuma, M; Nozawa, Y. Xanthones induce cell-cycle arrest and apoptosis in human colon cancer DLD-1 cells. Bioorg. Med. Chem. 2005, 13, 6064–6069. [Google Scholar]
- Ohguchi, K; Akao, Y; Matsumoto, K; Tanaka, T; Ito, T; Iinuma, M; Nozawa, Y. Vaticanol C-induced cell death is associated with inhibition of pro-survival signaling in HL60 human leukemia cell line. Biosci. Biotechnol. Biochem. 2005, 69, 353–356. [Google Scholar]
- Nakagawa, Y; Iinuma, M; Matsuura, N; Yi, K; Naoi, M; Nakayama, T; Nozawa, Y; Akao, Y. A potent apoptosis-inducing activity of a sesquiterpene lactone, arucanolide, in HL60 cells: a crucial role of apoptosis-inducing factor. J. Pharmacol. Sci. 2005, 97, 242–252. [Google Scholar]
- Matsumoto, K; Akao, Y; Yi, H; Ohguchi, K; Ito, T; Tanaka, T; Kobayashi, E; Iinuma, M; Nozawa, Y. Preferential target is mitochondria in α-mangostin-induced apoptosis in human leukemia HL60 cells. Bioorg. Med. Chem. 2004, 12, 5799–5806. [Google Scholar]
- Ito, T; Akao, Y; Yi, H; Ohguchi, K; Matsumoto, K; Tanaka, T; Iinuma, M; Nozawa, Y. Anti-tumor effect of resveratrol oligomers against human cancer cell lines and the molecular mechanism of apoptosis induced by vaticanol C. Carcinogenesis 2003, 24, 1489–1497. [Google Scholar]
- Nakagawa, Y; Iinuma, M; Naoe, T; Nozawa, Y; Akao, Y. Characterized mechanism of α-mangostin-induced cell death: Caspase-independent apoptosis with release of endonuclease-G from mitochondria and increased miR-143 expression in human colorectal cancer DLD-1 cells. Bioorg. Med. Chem. 2007, 16, 5620–5628. [Google Scholar]
- Matsumoto, K; Akao, Y; Ohguchi, K; Ito, T; Tanaka, T; Iinuma, M; Nozawa, Y. Xanthones induce cell-cycle arrest and apoptosis in human colon cancer DLD-1 cells. Bioorg. Med. Chem. 2005, 13, 6064–6069. [Google Scholar]
- Suksamrarn, S; Komutiban, O; Ratananukul, P; Chimnoi, N; Lartpornmatulee, N; Suksamrarn, A. Cytotoxic prenylated xanthones from the young fruit of Garcinia mangostana. Chem. Pharm. Bull. (Tokyo) 2006, 3, 301–305. [Google Scholar]
- Suksamrarn, S; Suwannapoch, N; Ratananukul, P; Aroonlerk, N; Suksamrarn, A. Xanthones from the green fruit hulls of Garcinia mangostana. J. Nat. Prod. 2002, 5, 761–763. [Google Scholar]
- Jung, HA; Su, BN; Keller, WJ; Mehta, RG; Kinghorn, AD. Antioxidant xanthones from the pericarp of Garcinia mangostana (Mangosteen). J. Agric. Food Chem. 2006, 6, 2077–2082. [Google Scholar]
- Wätjen, W; Weber, N; Lou, YJ; Wang, ZQ; Chovolou, Y; Kampkötter, A; Kahl, R; Proksch, P. Prenylation enhances cytotoxicity of apigenin and liquiritigenin in rat H4IIE hepatoma and C6 glioma cells. Food Chem. Toxicol. 2007, 1, 119–124. [Google Scholar]
- Bae, EY; Na, M; Njamen, D; Mbafor, JT; Fomum, ZT; Cui, L; Choung, DH; Kim, BY; Oh, WK; Ahn, JS. Inhibition of protein tyrosine phosphatase 1B by prenylated isoflavonoids isolated from the stem bark of Erythrina addisoniae. Planta. Med. 2006, 10, 945–948. [Google Scholar]
- Roux, PP; Blenis, J. ERK and p38 MAPK-activated protein kinases: a family of protein kinases with diverse biological functions. Microbiol. Mol. Biol. Rev. 2004, 68, 320–344. [Google Scholar]
- Pap, M; Cooper, GM. Role of translation initiation factor 2B in control of cell survival by the phosphatidylinositol 3-kinase/Akt/glycogen synthase kinase 3β signaling pathway. J. Biol. Chem. 1998, 273, 19929–19932. [Google Scholar]
- Sah, JF; Balasubramanian, S; Eckert, RL; Rorke, EA. Epigallocatechin-3-gallate inhibits epidermal growth factor receptor signaling pathway. Evidence for direct inhibition of ERK1/2 and AKT kinases. J. Biol. Chem. 2004, 279, 12755–12762. [Google Scholar]
- Aggarwal, BB; Bhardwaj, A; Aggarwal, RS; Seeram, NP; Shishodia, S; Takada, Y. Role of resveratrol in prevention and therapy of cancer: preclinical and clinical studies. Anticancer Res. 2004, 24, 2783–2840. [Google Scholar]
- Luo, Y; DeFranco, DB. Opposing roles for ERK1/2 in neuronal oxidative toxicity: distinct mechanisms of ERK1/2 action at early versus late phases of oxidative stress. J. Biol. Chem. 2006, 281, 16436–16442. [Google Scholar]
- Akao, Y; Nakagawa, Y; Naoe, T. MicroRNAs 143 and 145 are possible common onco-microRNAs in human cancers. Oncol. Rep. 2006, 4, 845–850. [Google Scholar]
- Akao, Y; Nakagawa, Y; Naoe, T. MicroRNA-143 and -145 in colon cancer. DNA Cell Biol. 2007, 5, 311–320. [Google Scholar]
- Bava, SV; Puliappadamba, VT; Deepti, A; Nair, A; Karunagaran, D; Anto, RJ. Sensitization of taxol-induced apoptosis by curcumin involves down-regulation of nuclear factor-κB and the serine/threonine kinase Akt and is independent of tubulin polymerization. J. Biol. Chem. 2005, 280, 6301–6308. [Google Scholar]
- Wein, A; Riedel, C; Bruckl, W; Kastl, S; Reingruber, B; Hohenberger, W; Hahn, EG. Weekly 24-h infusion of high-dose 5-fluorouracil (5-FU) with folinic acid (FA) in adjuvant therapy of colon cancer. Z. Gastroenterol. 2001, 39, 153–156. [Google Scholar]
- Tai, CJ; Liu, JH; Chen, WS; Lin, JK; Wang, WS; Yen, CC; Chiou, TJ; Chen, PM. Irinotecan (CPT11) plus high-dose 5-fluorouracil (5-FU) and leucovorin (LV) as salvage therapy for metastatic colorectal cancer (MCRC) after failed oxaliplatin plus 5-FU and LV: a pilot study in Taiwan. Jpn. J. Clin. Oncol. 2003, 33, 136–140. [Google Scholar]
- Zalcberg, J; Kerr, D; Seymour, L; Palmer, M. Haematological and non-haematological toxicity after 5-fluorouracil and leucovorin in patients with advanced colorectal cancer is significantly associated with gender, increasing age and cycle number. Eur. J. Cancer 1998, 34, 1871–1875. [Google Scholar]
- Petrelli, N; Douglass, HO, Jr; Herrera, L; Russell, D; Stablein, DM; Bruckner, HW; Mayer, RJ; Schinella, R; Green, MD; Muggia, FM. The modulation of fluorouracil with leucovorin in metastatic colorectal carcinoma: a prospective randomized phase III trial. J. Clin. Oncol. 1989, 7, 1419–1426. [Google Scholar]
- Maindrault-Goebel, F; de Gramont, A; Louvet, C; Andre, T; Carola, E; Mabro, M; Artru, P; Gilles, V; Lotz, JP; Izrael, V; Krulik, M. High-dose intensity oxaliplatin added to the simplified bimonthly leucovorin and 5-fluorouracil regimen as second-line therapy for metastatic colorectal cancer (FOLFOX 7). Eur. J. Cancer 2001, 37, 1000–1005. [Google Scholar]
- Mabro, M; Artru, P; Andre, T; Flesch, M; Maindrault-Goebel, F; Landi, B; Lledo, G; Plantade, A; Louvet, C; de Gramont, A. A phase II study of FOLFIRI-3 (double infusion of irinotecan combined with LV5FU) after FOLFOX in advanced colorectal cancer patients. Br. J. Cancer 2006, 94, 1287–1292. [Google Scholar]
- Kawashima, R; Haisa, M; Kimura, M; Takaoka, M; Shirakawa, Y; Takeda, H; Uetsuka, H; Gunduz, M; Nagai, N; Tanaka, N; Naomoto, Y. Cyclin A correlates with the sensitivity of human cancer cells to cytotoxic effects of 5-FU. Int. J. Oncol. 2004, 24, 273–278. [Google Scholar]
- Nabandith, V; Suzui, M; Morioka, T; Kaneshiro, T; Kinjo, T; Matsumoto, K; Akao, Y; Iinuma, M; Yoshimi, N. Inhibitory effects of crude α-mangostin, a xanthone derivative, on two different categories of colon preneoplastic lesions induced by 1, 2-dimethylhydrazine in the rat. Asian Pac. J. Cancer Prev. 2004, 4, 433–438. [Google Scholar]
- Wu, X; Beecher, GR; Holden, JM; Haytowitz, DB; Gebhardt, SE; Prior, RL. Lipophilic and hydrophilic antioxidant capacities of common foods in the United States. J. Agric. Food Chem. 2004, 12, 4026–4037. [Google Scholar]
- Nakatani, K; Yamakuni, T; Kondo, N; Arakawa, T; Oosawa, K; Shimura, S; Inoue, H; Ohizumi, Y. γ-Mangostin inhibits inhibitor-κB kinase activity and decreases lipopolysaccharide-induced cyclooxygenase-2 gene expression in C6 rat glioma cells. Mol. Pharmacol. 2004, 3, 667–674. [Google Scholar]
- Nakatani, K; Nakahata, N; Arakawa, T; Yasuda, H; Ohizumi, Y. Inhibition of cyclooxygenase and prostaglandin E2 synthesis by gamma-mangostin, a xanthone derivative in mangosteen, in C6 rat glioma cells. Biochem. Pharmacol. 2002, 63, 73–79. [Google Scholar]
- Gopalakrishnan, C; Shankaranarayanan, D; Kameswaran, L; Nazimudeen, SK. Effect of mangostin, a xanthone from Garcinia mangostana Linn. in immunopathological & inflammatory reactions. Indian J. Exp. Biol. 1980, 18, 843–846. [Google Scholar]
- Yamakuni, T; Aoki, K; Nakatani, K; Kondo, N; Oku, H; Ishiguro, K; Ohizumi, Y. Garcinone B reduces prostaglandin E2 release and NF–κB-mediated transcription in C6 rat glioma cells. Neurosci. Lett. 2006, 3, 206–210. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| agent | IC50 |
|---|---|
| α-mangostin | 7.5 μM |
| β-managostin | 8.1 μM |
| γ-managostin
| 7.1 μM
|
| 5-FU | 4.5 μM |
| Actinomycin D | 0.34 μM |
| Camptothecin | 7.0 nM |
| Taxol | 5.0 nM |
| Etoposide | 2.7 nM |
Share and Cite
Akao, Y.; Nakagawa, Y.; Nozawa, Y. Anti-Cancer Effects of Xanthones from Pericarps of Mangosteen. Int. J. Mol. Sci. 2008, 9, 355-370. https://doi.org/10.3390/ijms9030355
Akao Y, Nakagawa Y, Nozawa Y. Anti-Cancer Effects of Xanthones from Pericarps of Mangosteen. International Journal of Molecular Sciences. 2008; 9(3):355-370. https://doi.org/10.3390/ijms9030355
Chicago/Turabian StyleAkao, Yukihiro, Yoshihito Nakagawa, and Yoshinori Nozawa. 2008. "Anti-Cancer Effects of Xanthones from Pericarps of Mangosteen" International Journal of Molecular Sciences 9, no. 3: 355-370. https://doi.org/10.3390/ijms9030355
APA StyleAkao, Y., Nakagawa, Y., & Nozawa, Y. (2008). Anti-Cancer Effects of Xanthones from Pericarps of Mangosteen. International Journal of Molecular Sciences, 9(3), 355-370. https://doi.org/10.3390/ijms9030355