Fumonisins, Trichothecenes and Zearalenone in Cereals

Abstract
:
1. Introduction
2. Fumonisins
Short history and synthesis
Stability
Products
Mode of action
Toxicity
3. Trichothecenes
Short history and synthesis
- Type A: functional group other than a ketone at C8 position (e.g.; T-2, HT-2, DAS);
- Type B: carbonyl functions at C8 position (e.g.; DON, NIV, FUS-X, 3-acetyl-deoxynivalenol, 15-acetyl-deoxynivalenol);
- Type C: second epoxide group at C7, 8 or C9, 10; (e.g.; crotocin and baccharin);
Stability
Products
Mode of action
Toxicity
3.1. T-2 and HT-2
Short history and synthesis
Stability
Products
Mode of action
Toxicity
3.2. DON
Short history and synthesis
Stability
Products
Mode of action
Toxicity
3.3. NIV
Short history and synthesis
Stability
Products
The mode of action
Toxicity
3.4. DAS
Short history and synthesis
Stability
Products
Mode of action
Toxicity
3.5. FUS-X
Short history and synthesis
Stability
Products
Mode of action
Toxicity
4. ZEA
Short history and synthesis
Stability
Products
The mode of action
Toxicity
5. Conclusions
References
- Magan, N; Aldred, D. Post-harvest control strategies: Minimizing mycotoxins in the food chain. Int. J. Food Microbiol 2007, 119, 131–139. [Google Scholar]
- Gelderblom, WC; Marasas, WF; Vleggaar, R; Thiel, PG; Cawood, ME. Fumonisins: isolation, chemical characterization and biological effects. Mycopathologia 1992, 117, 11–16. [Google Scholar]
- Jackson, LS; Bullerman, LB. Effect of processing on Fusarium mycotoxins. Adv. Exp. Med. Biol 1999, 459, 243–261. [Google Scholar]
- FDA. Fumonisin levels in human foods and animal feeds, . Available at: http://www.cfsan.fda.gov/∼dms/fumongui.html. Accessed 9 November 2000.
- Creppy, EE. Update of survey, regulation and toxic effects of mycotoxins in Europe. Toxicol. Lett 2002, 127, 19–28. [Google Scholar]
- Omurtag, GZ; Yazıcıoglu, D; Beyoglu, D; Tozan, A; Atak, G. A review on fumonisin and trichothecene mycotoxins in foods consumed in Turkey. ARI, ITU-Bulletin 2006, 54, 39–45. [Google Scholar]
- Maragos, C; Bumsan, M; Sugita-Konishi, Y. Production and characterization of a monoclonal antibody that cross-reacts with the mycotoxins nivalenol and 4-deoxynivalenol. Food Addit. Contam 2006, 23, 816–825. [Google Scholar]
- Severino, L; Luongo, D; Bergamo, P; Lucisano, A; Rossi, M. Mycotoxins nivalenol and deoxynivalenol differentially modulate cytokine mRNA expression in Jurkat T cells. Cytokine 2006, 36, 75–82. [Google Scholar]
- Goyarts, T; Dänicke, S; Valenta, H; Ueberschär, KH. Carry-over of Fusarium toxins (deoxynivalenol and zearalenone) from naturally contaminated wheat to pigs. Food Addit. Contam 2007, 24, 369–380. [Google Scholar]
- IARC. Toxins derived from Fusarium moniliforme: Fumonisin B1 and B2 and fusarin C 1993.
- Bezuidenhout, SC; Gelderblom, WCA; Gorst-Allman, CP; Horak, RM; Marasas, WFO; Spiteller, G; Vleggaar, R. Structure elucidation of the fumonisins, mycotoxins from Fusarium moniliforme. J. Chem. Soc. Chem. Commun 1988, 743–745. [Google Scholar]
- Bennett, JW; Klich, M. Mycotoxins. Clin. Microbiol. Rev 2003, 16, 497–516. [Google Scholar]
- Soriano, JM; González, L; Catalá, AI. Mechanism of action of splingolipids and their metabolites in the toxicity of fumonisin B1. Prog. Lipid Res 2005, 44, 345–356. [Google Scholar]
- Wang, J; Zhou, Y; Liu, W; Zhu, X; Du, L; Wang, Q. Fumonisin level in corn-based food and feed from Linxian Country, a high-risk area for esophageal cancer in China. Food Chem 2008, 106, 241–246. [Google Scholar]
- Ciacci-Zanella, JR; Merrill, AH, Jr; Wang, E; Jones, C. Characterization of cell-cycle arrest by fumonisin B1 in CV-1 cells. Food Chem. Toxicol 1998, 36, 791–804. [Google Scholar]
- Frisvad, JC; Smedsgaard, J; Samson, RA; Larsen, TO; Thrane, U. Fumonisin B2 production by Aspergillus niger. J. Agric. Food Chem 2007, 55, 9727–9732. [Google Scholar]
- Doyle, E. Fusarium mycotoxins. FRI Briefings 1997, 1–4. [Google Scholar]
- Desjardins, AE; Manandhar, HK; Plattner, RD; Manandhar, GG; Poling, SM; Maragos, CM. Fusarium species from nepalese rice and production of mycotoxins and gibberellic acid by selected species. Appl. Environ. Microbiol 2000, 66, 1020–1025. [Google Scholar]
- Marasas, WFO. Discovery and occurrence of the fumonisins: A historical perspective. Environ. Health Perspect 2001, 109, 239–243. [Google Scholar]
- Haschek, WM; Gumprecht, LA; Smith, G; Tumbleson, ME; Constable, PD. Fumonisin toxicosis in swine: An overview of porcine pulmonary edema and current perspectives. Environ. Health Perspect 2001, 109, 251–257. [Google Scholar]
- Castelo, MM; Sumner, SS; Bullerman, LB. Stability of fumonisins in thermally processed corn products. J. Food Prot 1998, 161, 1030–1033. [Google Scholar]
- Alberts, JF; Gelderblom, CA; Thiel, PG; Marasas, WFO; Van Schalkwyk, DJ; Behrend, Y. Effects of temperature and incubation period on protection of fumonisin B1 by Fusarium moniliforme. Appl. Environ. Microbiol 1990, 56, 1729–1733. [Google Scholar]
- Jackson, LS; Hlywka, JJ; Senthil, KR; Bullerman, LB. Effect of thermal processing on the stability of fumonisins. Adv. Exp. Med. Biol 1996, 392, 345–353. [Google Scholar]
- Marin, S; Sanchis, V; Vinas, I; Canela, R; Magan, N. Effect of water activity and temperature on growth and fumonisin B1 and B2 production by Fusarium proliferatum and F. moniliforme on maize grain. Lett. Appl. Microbiol 1995, 21, 298–301. [Google Scholar]
- Pacin, AM; Broggi, LE; Resnik, SL; González, HHL. Mycoflora and mycotoxins natural occurrence in corn from Entre Rios province, Argentina. Myco. Res 2001, 17, 31–38. [Google Scholar]
- Broggi, LE; Resnik, SL; Pacin, AM; González, HH; Cano, G; Taglieri, D. Distribution of fumonisins in dry-milled corn fractions in Argentina. Food Addit. Contam 2002, 19, 465–469. [Google Scholar]
- Ho, JA; Durst, RA. Detection of fumonisin B1: Comparison of flow-injection liposome immunoanalysis with high-performance liquid chromatography. Anal. Biochem 2003, 312, 7–13. [Google Scholar]
- Soriano, JM; Dragacci, S. Occurrence of fumonisins in foods. Food Res. Intern 2004, 37, 985–1000. [Google Scholar]
- Domijan, A-M; Želježić, D; Milić, M; Peraica, M. Fumonisin B1: Oxidative status and DNA damage in rats. Toxicology 2007, 232, 163–169. [Google Scholar]
- International Programme on Chemical Safety (IPCS), Fumonisin B1, . Environ. Health Criteria 219. World Health Organization: Geneva, Switzerland. Available at: http://www.inchem.org/documents/ehc/ehc/ehc219.htm, 12 February 2000.
- Marasas, WF; Kellerman, TS; Gelderblom, WC; Coetzer, JA; Thiel, PG; van der Lugt, JJ. Leukoencephalomalacia in a horse induced by fumonisin B1 isolated from Fusarium moniliforme. Onderstepoort J. Vet. Res 1988, 55, 197–203. [Google Scholar]
- Wilson, TM; Poss, PF; Rice, LG; Osweiler, GD; Nelson, HA; Owens, DL; Plattner, RD; Reggiardo, C; Noon, TH; Pickrell, JW. Fumonisin B1 levels associated with an epizootic of equine leukoencephalomalacia. J. Vet. Diagn. Invest 1990, 2, 213–216. [Google Scholar]
- Kellerman, TS; Marasas, WF; Thiel, PG; Gelderblom, WC; Cawood, M; Coetzer, JA. Leukoencephalomalacia in two horses induced by oral dosing of fumonisin B1. Onderstepoort J. Vet. Res 1990, 57, 269–275. [Google Scholar]
- Bucci, TJ; Howard, PC. Effect of fumonisin mycotoxins in animals. J. Toxicol. Toxin. Rev 1996, 15, 293–302. [Google Scholar]
- Kriek, NP; Kellerman, TS; Marasas, WF. A comparative study of the toxicity of Fusarium verticillioides (= F. moniliforme) to horses, primates, pigs, sheep and rats. Onderstepoort J. Vet. Res 1981, 48, 129–131. [Google Scholar]
- Harrison, LR; Colvin, BM; Grene, JT; Newman, LE; Cole, JR. Pulmonary edema and hydrothorax in swine produced by fumonisin B1, a toxic metabolite of Fusarium moniliforme. J. Vet. Diagn. Invest 1990, 2, 217–221. [Google Scholar]
- Fincham, JE; Marasas, WF; Taljaard, JJ; Kriek, NP; Badenhorst, CJ; Gelderblom, WC; Seier, JV; Smuts, CM; Faber, M; Weight, MJ. Atherogenic effects in a non-human primate of Fusarium moniliforme cultures added to a carbohydrate diet. Atherosclerosis 1992, 94, 13–25. [Google Scholar]
- Bucci, TJ; Hansen, DK; LaBorde, JB. Leukoencephalomalacia and hemorrhage in the brain of rabbits gavaged with mycotoxin fumonisin B1. J. Nat. Toxins 1996, 4, 51–52. [Google Scholar]
- Gelderblom, WCA; Jaskiewicz, K; Marasas, WFO; Thiel, PG; Horak, RM; Vleggaar, R; Kriek, NPJ. Fumonisins-Novel mycotoxins with cancer-promoting activity produced by Fusarium moniliforme. Appl. Environ. Microbiol 1988, 54, 1806–1811. [Google Scholar]
- Gelderblom, WCA; Kriek, NPJ; Marasas, WFO; Thiel, PG. Toxicity and carcinogenecity of the Fusarium moniliforme metabolite, fumonisin B1, in rats. Carcinogenesis 1991, 12, 1247–1251. [Google Scholar]
- Hard, GC; Howard, PC; Kovatch, RM; Bucci, TJ. Rat kidney pathology induced by chronic exposure to fumonisin B1 includes rare variants of renal tubule tumor. Toxicol. Pathol 2001, 29, 379–386. [Google Scholar]
- Marasas, WF; Riley, RT; Hendricks, KA; Stevens, VL; Sadler, TW; Gelineauvan Waes, J; Missmer, SA; Cabrera, J; Torres, O; Gelderblom, WC; Allegood, J; Martínerz, C; Maddox, J; Miller, JD; Star, L; Sullards, MC; Roman, AV; Voss, KA; Wang, E; Merril, AH. Fumonisins distrupt sphingolipid metabolism, folate transport, and neuronal tube development in embryo culture and in vivo: A potential risk factor for human neuronal tube defects among populations consuming fumonisin-contaminated maize. J. Nutr 2004, 134, 711–716. [Google Scholar]
- Edrington, TS; Kamps-Holtzapple, CA; Harvey, RB; Kulbena, LF; Elissalde, MH; Rottinghaus, GE. Acute hepatic and renal toxicity in lambs dosed with fumonisin-containing culture material. J. Anim. Sci 1995, 73, 508–515. [Google Scholar]
- Bucci, TJ; Howard, PC; Tolleson, WH; Laborde, JB; Hansen, DK. Renal effects of fumonisin mycotoxins in animals. Toxicol. Pathol 1998, 26, 160–164. [Google Scholar]
- Voss, KA; Riley, RT; Norred, WP; Bacon, CW; Meredith, FI; Howard, PC; Plattner, RD; Collins, TFX; Hansen, DK; Porter, JK. An overview of rodent toxicities: liver and kidney effects of fumonisins and Fusarium moniliforme. Environ. Health Perspect 2001, 109, 259–266. [Google Scholar]
- Dombrink-Kurtzman, MA; Javed, T; Bennett, GA; Richard, JL; Cote, LM; Buck, WB. Lymphocyte cytotoxicity and erythrocytic abnormalities induced in broiler chicks by fumonisin B1 and B2 and moniliformin from Fusarium proliferatum. Mycopathologia 1993, 124, 47–54. [Google Scholar]
- Dombrink-Kurtzman, MA; Bennett, GA; Richard, JL. An optimized MTT bioassay for determination of cytotoxicity of fumonisins in turkey lymphocytes. J. AOAC Int 1994, 77, 512–516. [Google Scholar]
- Qureshi, MA; Hagler, WM, Jr. Effect of fumonisin-B1 exposure on chicken macrophage functions in vitro. Poult. Sci 1992, 71, 104–112. [Google Scholar]
- Rumora, L; Kovačić, S; Rozgaj, R; Čepelak, I; Pepeljnjak, S; Grubisić, TZ. Cytotoxic and genotoxic effects of fumonisin B1 on rabbit kidney RK13 cell line. Arch. Toxicol 2002, 76, 55–61. [Google Scholar]
- Franceschi, S; Bidoli, E; Barón, AE; La Vecchia, C. Maize and risk of cancer of the oral cavity, pharynx, and esophagus in northeastern Italy. J. Natl. Cancer Inst 1990, 82, 1407–1411. [Google Scholar]
- Shephard, GS; Marasas, WF; Leggott, NL; Yazdanpanah, H; Rahimian, H; Safavi, N. Natural occurrence of fumonisins in corn from Iran. J. Agric. Food Chem 2000, 48, 1860–1864. [Google Scholar]
- Rheeder, JP; Marasas, WFO; Thiel, PG; Sydenham, EW; Shephard, GS; van Schalkwyk, DJ. Fusarium moniliforme and fumonisins in corn in relation to human esophageal cancer in Transkei. Phytopathology 1992, 82, 353–357. [Google Scholar]
- Chu, FS; Li, GY. Simultaneous occurrence of fumonisin B1 and other mycotoxins in moldy corn collected from the People’s Republic of China in regions with incidences of esophageal cancer. Appl. Environ. Microbiol 1994, 60, 847–852. [Google Scholar]
- Groves, FD; Zhang, L; Chang, YS; Ross, PF; Casper, H; Norred, WP; You, WC; Fraumeni, JF, Jr. Fusarium mycotoxins in corn and corn products in a high-risk area for gastric in Shandong Province, China. J. AOAC Int 1999, 82, 657–662. [Google Scholar]
- World Health Organization (WHO) International Agency for Research on Cancer (IARC), IARC monographs on the evaluation of carcinogenic risks to humans; , Lyon 2002; ; volumes Volume 82, , pp. pp. 301–366.
- Duvick, J. Prospects for reducing fumonisin contamination of maize through genetic modification. Environ. Health Perspect 2001, 109, 337–342. [Google Scholar]
- Keck, BB; Bodini, AB. The effects of fumonisin B1 on viability and mitogenic response of avian immune cells. Poult. Sci 2006, 85, 1020–1024. [Google Scholar]
- Aranda, M; Pérez-Alzola, LP; Ellahueñe, MF; Sepúlveda, C. Assesment of in vitro mutagenicity in Salmonella and in vivo genotoxicity in mice of the mycotoxin fumonisin B1. Mutagenesis 2000, 15, 469–471. [Google Scholar]
- Turner, PC; Nikiema, P; Wild, CP. Fumonisin contamination of food: progress in development of biomarkers to better assess human health risks. Mutat. Res 1999, 443, 81–93. [Google Scholar]
- Humpf, HU; Voss, KA. Effects of thermal food processing on the chemical structure and toxicity. Mol. Nutr. Food Res 2004, 48, 255–269. [Google Scholar]
- FDA. Background paper in support of fumonisin levels in corn and corn products intended for human consumption, , Available at: http://www.micotoxinas.com.br/boletim36.pdf, accessed 9 November 2001.
- Council for Agricultural Science and Technology, Mycotoxins: Risks in plant, animal, and human system; ; Task Force Report, No.139 Ames, Iowa, USA, 2003; pp. 48–58.
- Pestka, JJ; Smolinski, AT. Deoxynivalenol: Toxicology and potential effects on humans. J. Environ. Sci. Health B 2005, 8, 39–69. [Google Scholar]
- Eriksen, GS. Metabolism and Toxicity of Trichothecenes; , Doctoral thesis, Uppsala, Sweden, 2003.
- Sudakin, DL. Trichothecenes in the environment: Relevance to human health. Toxicol. Lett 2003, 143, 97–107. [Google Scholar]
- Hsia, CC; Wu, JL; Lu, XQ; Li, YS. Natural occurrence and clastogenic effects of nivalenol, deoxynivalenol, 3-acetyl-deoxynivalenol, 15-acetyl-deoxynivalenol, and zearalenone in corn from a high-risk area of esophageal cancer. Cancer Detect. Prev 1988, 13, 79–86. [Google Scholar]
- Eriksen, GS; Pettersson, H. Toxicological evaluation of trichothecenes in animal feed. Anim. Feed Sci. Technol 2004, 114, 205–239. [Google Scholar]
- Rocha, O; Ansar, K; Doohan, FM. Effects of trichothecene mycotoxins on eukaryotic cells: A review. Food Addit. Contam 2005, 22, 369–378. [Google Scholar]
- Koch, P. State of the art of trichothecenes analysis. Toxicol. Lett 2004, 153, 109–112. [Google Scholar]
- Pestka, JJ. Deoxynivalenol:toxicity, mechanisms and animal health risks. Anim. Feed Sci. Technol 2007, 137, 283–298. [Google Scholar]
- Schrödter, R. Influence of harvest and storage conditions on trichothecenes levels in various cereals. Toxicol. Lett 2004, 153, 47–49. [Google Scholar]
- Widestrand, J; Pettersson, H. Effect of time, temperature and solvent on the stability of T-2 toxin, HT-2 toxin, deoxynivalenol and nivalenol calibrants. Food Addit. Contam 2001, 18, 987–992. [Google Scholar]
- Lauren, DR; Smith, WA. Stability of the Fusarium mycotoxins nivalenol, deoxynivalenol and zearalenone in ground maize under typical cooking environments. Food Addit. Contam 2001, 18, 1011–1016. [Google Scholar]
- Larsen, JC; Hunt, J; Perin, I; Ruckenbauer, P. Workshop on trichothecenes with a focus on DON: Summary report. Toxicol. Lett 2004, 1–22. [Google Scholar]
- Osweiler, GD. Diagnosis of mycotoxicoses. In Occurrence and clinical manifestations of trichothecene toxicoses and zearalenone toxicoses; Richard, JL, Thurston, JR, Eds.; ., Eds.; National Animal Disease Center: Ames, Iowa, USA, 1986; pp. 31–42. [Google Scholar]
- D’Mello, JPF; Placinta, CM; Macdonald, AMC. Fusarium mycotoxins: A review of global implications for animal health, welfare and productivity. Anim. Feed Sci. Technol 1999, 80, 183–205. [Google Scholar]
- Thuvander, A; Wikman, C; Gadhasson, I. In vitro exposure of human lymphocytes to trichothecenes: individual variation in sensitivity and effects of combined exposure on lymphocyte function. Food Chem. Toxicol 1999, 37, 639–648. [Google Scholar]
- Mateo, JJ; Mateo, R; Jiménez, M. Accumulation of type A trichothecenes in maize, wheat and rice by Fusarium sporotrichioides isolates under diverse culture conditions. Int. J. Food Microbiol 2002, 72, 115–123. [Google Scholar]
- Holt, PS; Buckley, S; Norman, JO; Deloach, JR. Cytotoxic effect of T-2 mycotoxin on cells in culture as determined by a rapid colorimetric bioassay. Toxicon 1988, 26, 549–558. [Google Scholar]
- Rabie, CJ; Sydenham, EW; Thiel, PG; Lübben, A; Marasas, WFO. T-2 toxin production by Fusarium acuminatum isolated from oats and barley. Appl. Environ. Microbiol 1986, 52, 594–596. [Google Scholar]
- Omurtag, GZ; Yazicioglu, D. Occurrence of T-2 toxin in processed cereals and pulses in Turkey determined by HPLC and TLC. Food Addit. Contam 2001, 18, 844–849. [Google Scholar]
- Trusal, LR. Stability of T-2 mycotoxin in aqueous media. Appl. Environ. Microbiol 1985, 50, 1311–1312. [Google Scholar]
- Pace, JG; Matson, CF. Stability of T-2, HT-2 and T-2 tetraol in biological fluids. J. Anal. Toxicol 1988, 12, 48–50. [Google Scholar]
- Scientific Committee on Food. Opinion on Fusarium Toxins. Part 5: T-2 toxin and HT-2 toxin.
- Gyongyossy-Issa, MIC; Khanna, V; Khachatourians, GC. Characterisation of hemolysis induced by T-2 toxin. Biochim. Biophys. Acta 1985, 838, 252–256. [Google Scholar]
- Holt, PS; Buckley, S; Norman, JO; Deloach, JR. Cytotoxic effect of T-2 mycotoxin on cells in culture as determined by a rapid colorimetric bioassay. Toxicon 1988, 26, 549–558. [Google Scholar]
- Moss, MO. Mycotoxin review-2. Fusarium. Mycologist 2002, 16, 158–161. [Google Scholar]
- Pacin, A; Reale, C; Mirengui, H; Orellana, L; Boente, G. Subclinic effect of the administration of T-2 toxin and nivalenol in mice. Mycotoxin Research 1994, 10, 34–46. [Google Scholar]
- Wang, ZG; Feng, JN; Tong, Z. Human toxicosis caused by moldy rice contaminated with Fusarium and T-2 toxin. Biomed. Environ. Sci 1993, 6, 65–70. [Google Scholar]
- Samar, MM; Resnik, SL; González, HHL; Pacin, AM; Castillo, MD. Deoxynivalenol reduction during the frying process of turnover pie covers. Food Control 2007, 18, 1295–1299. [Google Scholar]
- Instanes, C; Hetland, G. Deoxynivalenol (DON) is toxic to human colonic, lung and monocytic cell lines, but does not increase the IgE response in a mouse model for allergy. Toxicology 2004, 204, 13–21. [Google Scholar]
- González, HH; Moltó, GA; Pacin, A; Resnik, SL; Zelaya, MJ; Masana, M; Martínez, EJ. Trichothecenes and mycoflora in wheat harvested in nine locations in Buenos Aires province, Argentina. Mycopathologia 2008, 165, 105–114. [Google Scholar]
- Wijnands, LM; van Leusden, FM. An overview of adverse health effects caused by mycotoxins and biossays for their detection; , RIVM report 257852 004, Bilthoven, 2000.
- Rotter, BA; Prelusky, DB; Pestka, JJ. Invited Review: Toxicology of Deoxynivalenol (vomitoxin). J. Toxicol. Environ. Health Part A 1996, 48, 1–34. [Google Scholar]
- Pestka, JJ; Zhou, H; Moon, Y; Chung, YJ. Cellular and molecular mechanisms for immune modulation by deoxynivalenol and other trichothecenes: unraveling a paradox. Toxicol. Lett 2004, 153, 61–73. [Google Scholar]
- Ito, Y; Ohtsubo, K; Ishii, K; Ueno, Y. Effects of nivalenol on pregnancy and fetal development of mice. Mycotoxin Research 1986, 2, 71–77. [Google Scholar]
- Larsen, JC; Hunt, J; Perin, I; Ruckenbauer, P. Workshop on trichothecenes with a focus on DON: Summary report. Toxicol. Lett 2004, 153, 1–22. [Google Scholar]
- Tep, J; Videmann, B; Mazallon, M; Balleydier, S; Cavret, S; Lecoeur, S. Transepithelial transport of fusariotoxin nivalenol: Mediation of secretion by ABC transporters. Toxicol. Lett 2007, 170, 248–258. [Google Scholar]
- Scientific Committee on Food, (SCF). Opinion of the Scientific Committee on food on Fusarium Toxins. Part 4: Nivalenol.
- Maragos, C; Bumsan, M; Sugita-Konish, Y. Production and characterization of a monoclonal antibody that cross-react with the mycotoxins nivalenol and 4-deoxynivalenol. Food Addit. Contam 2006, 23, 816–825. [Google Scholar]
- Ueno, Y; Umemori, K; Nimi, E; Tanuma, S; Nagata, S; Sugamata, M; Ihara, T; Sekijima, M; Kawai, K; Ueno, I. Induction of apoptosis by T-2 toxin and other natural toxins in HL-60 human promyelotic leukemia cells. Nat. Toxins 1995, 3, 129–137. [Google Scholar]
- Pronk, MEJ; Schothorst, RC; van Egmond, HP. Toxicology and occurrence of nivalenol, fusarenon X, diacetoxyscirpenol, neosolaniol and 3- and 15-acetylnivalenol: A review of six trichothecenes. 2002, 75.
- Choi, C-Y; Nakajima-Adachi, H; Kaminogawa, S; Sugita-Konishi, Y. Nivalenol inhibits total an antigen-specific IgE prouction in mice. Toxicol. Appl. Pharmacol 2000, 165, 94–98. [Google Scholar]
- Ryu, JC; Ohtsubo, K; Izumiyama, N; Nakamura, K; Tanaka, T; Yamamura, H; Ueno, Y. The acute and chronic toxicities of nivalenol in mice. Fundam Appl. Toxicol 1988, 11, 38–47. [Google Scholar]
- Bauer, J; Bollwahn, W; Gareis, M; Gedek, B; Heinritzi, K. Kinetic profiles of Diacetoxyscirpenol and two of its metabolites in blood serum of pigs. Appl. Environ. Microbiol 1985, 49, 842–845. [Google Scholar]
- Omurtag, GZ; Tozan, A; Sirkecioglu, O; Kumbaraci, V; Rollas, S. Occurrence of diacetoxyscirpenol (anguidine) in processed cereals and pulses in Turkey by HPLC. Food Control 2007, 18, 970–974. [Google Scholar]
- Swanson, SP; Terwel, L; Corley, RA; Buck, WB. Gas chromatographic method fort he determination of diacetoxyscirpenol in swine plasma and urine. J. Chromatogr 1982, 248, 456–460. [Google Scholar]
- Mateo, JJ; Mateo, R; Jimanez, M. Accumulation of type A trichothecenes in maize, wheat and rice by Fusarium sporotrichioides isolates under diverse culture conditions. Int. J. Food Microbiol 2002, 72, 115–123. [Google Scholar]
- Patey, AL; Gilbert, J. Fate of Fusarium mycotoxins in cereals during food processing and methods for their detoxification. Fusarium mycotoxins, Taxonomy and pathogenicity 1989, Volume 2, 399–420. [Google Scholar]
- Wang, J-S; Busby, WF; Wogan, GN. Percutaneous absorption and tissue distribution of [3H]diacetoxyscirpenol (Anguidine) in rats and mice. Toxicol. Appl. Pharmacol 1996, 140, 264–273. [Google Scholar]
- Mirocha, CJ; Pathre, SV; Schauerhamer, B; Christensen, CM. Natural occurrence of Fusarium toxins in feedstuff. Appl. Environ. Microbiol 1976, 32, 553–556. [Google Scholar]
- Swanson, SP; Nicoletti, J; Rood, HD; Buck, WB; Cote, LM. Metabolism of three trichothecene mycotoxins, T-2 toxin, diacetoxyscirpenol and deoxynivalenol, by bovine rumen microorganisms. J. Chromatogr 1987, 414, 335–342. [Google Scholar]
- Wang, J-S; Busby, WF; Wogan, GN. Comparative tissue distribution and excretion of orally administered [3H] diacetoxyscirpenol (anguidine) in rats and mice. Toxicol. Appl. Pharmacol 1990, 103, 430–440. [Google Scholar]
- Mayura, K; Smith, EE; Clement, BA; Harvey, RB; Kubena, LF; Phillips, TD. Developmental toxicity of diacetoxyscirpenol in the mouse. Toxicology 1987, 45, 245–255. [Google Scholar]
- Craddock, VM; Hill, RJ; Henderson, AR. Stimulation of DNA replication in rat esophagus and stomach by the trichothecene mycotoxin diacetoxyscirpenol. Cancer Lett 1987, 38, 199–208. [Google Scholar]
- Masuda, E; Takemoto, T; Tatsuno, T; Obara, T. Induction of suppressor macrophages in mice by Fusarenon-X. Immunology 1982, 47, 701–708. [Google Scholar]
- Weidenbörner, M. Encyclopedia of Food mycotoxins; Springer: Berlin, German, 2000; pp. 1–80. [Google Scholar]
- Carter, CJ; Cannon, M. Inhibition of Eukaryotic ribosomal function by the sesquiterpenoid antibiotic Fusarenon-X, Eur. J. Biochem 1978, 84, 103–111. [Google Scholar]
- Bony, S; Olivier-Loiseau, L; Carcelen, M; Devaux, A. Genotoxic potential associated with low levels of the Fusarium mycotoxins nivalenol and fusarenon-X in a human intestinal cell line. Toxicol. In Vitro 2007, 21, 457–465. [Google Scholar]
- Miura, K; Aminova, L; Murayama, Y. Fusarenon-X induced apoptosis in HL-60 cells depends on caspase activation and cytochrome c release. Toxicology 2002, 172, 103–112. [Google Scholar]
- Tritscher, AM; Page, SW. The risk assessment paradigm and its application for trichothecenes. Toxicol. Lett 2004, 153, 155–163. [Google Scholar]
- Bondy, GS; McCormick, SP; Beremand, MN; Pestka, JJ. Murine lymphocyte proliferation impaired by substituted neosolaniols and calonectrins--Fusarium metabolites associated with trichothecene biosynthesis. Toxicon 1991, 29, 1107–1113. [Google Scholar]
- Krska, R; Pettersson, H; Josephs, RD; Lemmens, M; MacDonald, S; Welzig, E. Zearalenone in maize: stability testing and matrix characterization of a certified reference material. Food Addit. Contam. Part A 2003, 20, 1141–1152. [Google Scholar]
- Placinta, CM; D’Mello, JPF; Macdonald, AMC. A review of worldwide contamination of cereal grains and animal feed with Fusarium mycotoxins. Anim. Feed Sci. Technol 1999, 78, 21–37. [Google Scholar]
- Wood, GE. Mycotoxins in foods and feeds in the United States. J. Anim. Sci 1992, 70, 3941–3949. [Google Scholar]
- Scientific Committee on Food (SCF). Opinion of the Scientific Committee on food on Fusarium Toxins. Part 2: Zearalenone (ZEA).
- Diekman, MA; Green, ML. Mycotoxins and reproduction in domestic livestock. J. Anim. Sci 1992, 70, 1615–1627. [Google Scholar]
- González, HH; Martínez, EJ; Pacin, AM; Resnik, SL; Sydenham, EW. Natural co-occurrence of fumonisins, deoxynivalenol, zearalenone and aflatoxins in field trial corn in Argentina. Food Addit. Contam 1999, 16, 565–569. [Google Scholar]
- Ruhr, LP; Osweiler, GD; Foley, CW. Effect of the estrogenic mycotoxin zearalenone on reproductive potential in the boar. Am. J. Vet. Res 1983, 44, 483–485. [Google Scholar]
- Goyarts, T. Effects of the Fusarium toxin deoxynivalenol (DON) on protein synthesis, immunological parameters and DON-kinetics in the pig; . Doctoral Thesis, Hannover, 2006.
- Tanaka, K; Sago, Y; Zheng, Y; Nakagawa, H; Kushiro, M. Mycotoxins in rice. Int. J. Food Microbiol 2007, 119, 59–66. [Google Scholar]
- Schothorst, R; van Egmond, HP. Report from SCOOP task 3.2.10 “collection of occurrence data of Fusarium toxins in food and assessment of dietary intake by the population of EU member states” Subtask: Trichothecenes. Toxicol. Lett 2004, 153, 133–143. [Google Scholar]
- Binder, EM; Tan, LM; Chin, LJ; Handl, J; Richard, J. Worldwide occurrence of mycotoxins in commodities, feeds and feed ingredients. Anim. Feed Sci. Technol 2007, 137, 265–282. [Google Scholar]
- van Egmond, HP; Jonker, MA. Regulations for mycotoxins in food: Focus on the European Union and Turkey. ARI, ITU-Bulletin 2006, 54, 1–17. [Google Scholar]
- International Programme on Chemical Safety (INCHEM). Environmental Health Criteria 105, Selected Mycotoxins: Ochratoxins, Trichothecenes, Ergot.
- Berg, T. How to establish international limits for mycotoxin in food and feed? Food Control 2003, 14, 219–224. [Google Scholar]
- World Health Organization (WHO). Evaluation of certain mycotoxins in Food. 56th Report of the Joint FAO/WHO Expert Committee on Food Additives (JECFA); . WHO Technical Report 906, Geneva, 2002; pp. 16–26. [Google Scholar]
- Arici, M; Daglioglu, O; Gumus, T; Daglioglu, F. Occurrence of fumonisin in processed and low processed corn-based products in Turkey. Acta Aliment 2004, 33, 325–328. [Google Scholar]
- Zinedine, A; Brera, C; Elakhdari, S; Catano, C; Debegnach, F; Angelini, S; De Santis, B; Faid, M; Benlemlih, M; Minardi, V; Miraglia, M. Natural occurrence of mycotoxins in cereals and species commercialized in Morocco. Food Control 2006, 17, 868–874. [Google Scholar]
- El-Sayed, AM; Soher, EA; Sahab, AF. Occurrence of certain mycotoxins in corn and corn-based products and thermostability of fumonisins B1 during processing. Nahrung 2003, 47, 222–225. [Google Scholar]
- Ariño, A; Estopañan, G; Juan, T; Herrera, A. Estimation of dietary intakes of fumonisins B1 and B2 from conventional and organic corn. Food Control 2007, 18, 1058–1062. [Google Scholar]
- Cvetnić, Z; Pepeljnjak, S; Šegvić, M. Toxigenic potential of Fusarium species isolated from non-harvested maize. Arh Hig Rada Toksikol 2005, 56, 275–280. [Google Scholar]
- Broggi, LE; Pacin, AM; Gasparovic, A; Sacchi, C; Rothermel, A; Gallay, A; Resnik, S. Natural occurrence of aflatoxins, deoxynivalenol, fumonisins and zearalenone in maize from Entre Rios Province, Argentina. Myco. Res 2007, 23, 59–64. [Google Scholar]
- Sangare-Tigori, B; Moukha, S; Kouadio, HJ; Betbeder, A-M; Dano, DS; Creppy, EE. Co-occurrence of aflatoxin B1, fumonisin B1, ochratoxin A and zearalenone in cereals and peanuts from Côte d’Ivoire. Food Addit. Contam 2006, 23, 1000–1007. [Google Scholar]
- Domijan, A-M; Peraica, M; Jurjević, Ž; Ivić, D; Cvjetković, B. Fumonisin B1, fumonisin B2, zearalenone and ochratoxin A contamination of maize in Croatia. Food Addit. Contam 2005, 22, 677–680. [Google Scholar]
- de Castro, MFPM; Shephard, GS; Sewram, V; Vicente, E; Mendonça, TA; Jordan, AC. Fumonisins in Brazilian corn-based foods for infant consumption. Food Addit. Contam 2004, 21, 693–699. [Google Scholar]
- Lombaert, GA; Pellaers, P; Roscoe, V; Mankotia, M; Neil, R; Scott, PM. Mycotoxins in infant cereal foods from the Canadian reteal market. Food Addit. Contam 2003, 20, 494–504. [Google Scholar]
- Pacin, AM; Gonzalez, HHL; Resnik, SL; Boca, RT; Burak, R; Broccoli, AM; de Souza, JC. Natural occurrence of mycotoxins and mycoflora of Argentinean popcorn. Myco. Res. 2002, 18–96.
- Mashinini, K; Dutton, MF. The incidence of fungi and mycotoxins in South Africa wheat and wheat-based products. J. Environ. Sci. Health B 2006, 41, 285–296. [Google Scholar]
- Park, JW; Choi, S-Y; Hwang, H-J; Kim, YB. Fungal mycoflora and mycotoxins in Korean polished rice destined for humans. Int. J. Food Microbiol 2005, 103, 305–314. [Google Scholar]
- Cirillo, T; Ritieni, A; Galvano, F; Cocchieri, RA. Natural co-occurrence of deoxynivalenol and fumonisins B1 and B2 in Italian marketed foodstuffs. Food Addit. Contam 2003, 20, 566–571. [Google Scholar]
- Molinié, A; Faucet, V; Castegnaro, M; Pfohl-Leszkowicz, A. Analysis of some breakfast cereals on the French market for their contents of ochratoxin A, citrinin and fumonisin B1:development of a method for simultaneous extraction of ochratoxin A and citrinin. Food Chem 2005, 92, 391–400. [Google Scholar]
- Solovey, MM; Somoza, C; Canos, G; Pacin, A; Resnik, S. A survey of fumonisins, deoxynivalenol, zearalenone and aflatoxins contamination in corn-based food products in Argentina. Food Addit. Contam 1999, 16, 325–329. [Google Scholar]
- Milanez, TV; Valente-Soares, LM; Baptista, GG. Occurrence of trichothecene mycotoxins in Brazilian corn-based food products. Food Control 2005, 17, 293–298. [Google Scholar]
- Krysińska-Traczyk, E; Perkowski, J; Dutkiewicz, J. Levels of fungi and mycotoxins in the samples of grain and grain dust collected from five various cereal crops in Eastern Poland. Ann. Agric. Environ. Med 2007, 14, 159–167. [Google Scholar]
- Adejumo, TO; Hettwer, U; Karlovsky, P. Occurrence of Fusarium species and trichothecenes in Nigerian maize. Int. J. Food Microbiol 2007, 116, 350–357. [Google Scholar]
- Cavaliere, C; D’Ascenzo, G; Foglia, P; Pastorini, E; Samperi, R; Laganà, A. Determination of type b trichothecenes and macrocyclic lactone mycotoxins in field contaminated maize. Food Chem 2005, 92, 559–568. [Google Scholar]
- Castillo, M-Á; Montes, R; Navarro, A; Segarra, R; Cuesta, G; Hernández, E. Occurrence of deoxynivalenol and nivalenol in Spanish corn-based food products. J. Food Compost. Anal 2008, 21, 423–427. [Google Scholar]
- Al-Julafi, MZ; Al-Falih, AM. Detection of trichothecenes in animal feeds and foodstuffs during the years 1997 to 2000 in Saudi Arabia. J. Food Prot 2001, 64, 1603–1606. [Google Scholar]
- Schollenberger, M; Jara, HT; Suchy, S; Drochner, W; Müler, HM. Fusarium toxins in wheat flour collected in an area in southwest Germany. Int. J. Food Microbiol 2002, 72, 85–89. [Google Scholar]
- Rasmussen, PH; Ghorbani, F; Berg, T. Deoxynivalenol and other Fusarium toxins in wheat and rye flours on the Danish market. Food Addit. Contam 2003, 20, 396–404. [Google Scholar]
- Mankevičienė, A; Butkutė, B; Dabkevičius, Z; Supronienė, S. Fusarium mycotoxins in Lithuanian cereals from the 2004–2005 harvests. Ann. Agric. Environ. Med 2007, 14, 103–107. [Google Scholar]
- Langseth, W; Rundberget, T. The occurrence of HT-2 toxin and other trichothecenes in Norwegian cereals. Mycopathologia 1999, 147, 157–165. [Google Scholar]
- Schneweis, I; Meyer, K; Engelhardt, G; Bauer, J. Occurrence of zearalenone-4-β-D-glucopyranoside in wheat. J. Agric. Food Chem 2002, 50, 1736–1738. [Google Scholar]
- Pan, D; Bonsignore, F; Rivas, F; Perera, G; Bettucci, L. Deoxynivalenol in barley samples from Uruguay. Int. J. Food Microbiol 2007, 114, 149–152. [Google Scholar]
- Ayalew, A; Fehrmann, H; Lepschy, J; Beck, R; Abate, D. Natural occurrence of mycotoxins in staple cereals from Ethiopia. Mycopathologia 2006, 162, 57–63. [Google Scholar]
- Perkowski, J; Basiński, T. Natural contamination of oat with group A trichothecene mycotoxins in Poland. Food Addit. Contam 2002, 19, 478–482. [Google Scholar]
- Omurtag, GZ; Yazicioglu, D. Determination of T-2 toxin in grain and grain products by HPLC and TLC. J. Environ. Sci. Health B 2000, 35, 797–807. [Google Scholar]
- Li, F-Q; Luo, X-Y; Yoshizawa, T. Mycotoxins (trichothecenes, zearalenone and fumonisins) in cereals associated with human red-mold intoxications stored since 1989 and 1991 in China. Nat. Toxins 1999, 7, 93–97. [Google Scholar]
- Eskola, M; Parikka, P; Rizzo, A. Trichothecenes, ochratoxin A and zearalenone contamination and Fusarium infection in Finnish cereal samples in 1998. Food Addit. Contam 2001, 18, 707–718. [Google Scholar]
- Schollenberger, M; Suchy, S; Jara, HT; Drochner, W; Müller, HM. A survey of Fusarium toxins in cereal-based foods marketed in an area of southwest Germany. Mycopathologia 1999, 147, 49–57. [Google Scholar]
Abbreviations:
| FB | Fumonisin |
| TCs | Trichothecenes |
| DON | Deoxynivalenol |
| NIV | Nivalenol |
| DAS | Diacetoxyscirpenol |
| FUS-X | Fusarenone-X |
| HT-2 | HT-2 toxin |
| T-2 | T-2 toxin |
| ZEA | Zearalenone |

{kind=link}
{kind=link}
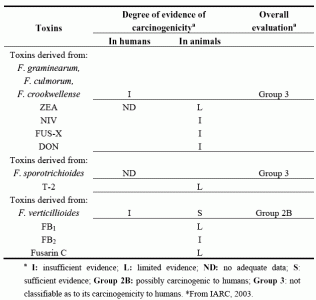
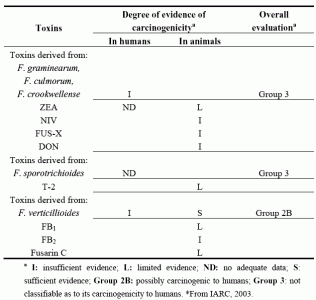
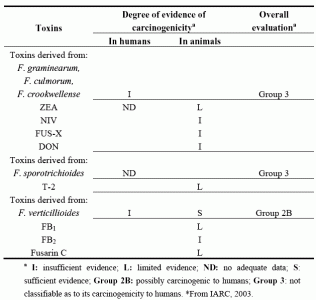
| Toxins | Degree of evidence of carcinogenicitya | Overall evaluationa | |
|---|---|---|---|
| In humans | In animals | ||
| Toxins derived from: | |||
| F. graminearum, | |||
| F. culmorum, | |||
| F. crookwellense | I | Group 3 | |
| ZEA | ND | L | |
| NIV | I | ||
| FUS-X | I | ||
| DON | I | ||
| Toxins derived from: | |||
| F. sporotrichioides | ND | Group 3 | |
| T-2 | L | ||
| Toxins derived from: | |||
| F. verticillioides | I | S | Group 2B |
| FB1 | L | ||
| FB2 | I | ||
| Fusarin C | L | ||
| Product | Detected/total | Range | Country | References |
|---|---|---|---|---|
| FB1: 22/110 | FB1: nd–2.66 ppm | |||
| Corn | Turkey | 6 | ||
| FB2: 0/110 | FB2: nd | |||
| Corn | TFBs: 30/92 | TFBs: 0.3–273 mg/kg | Turkey | 138 |
| Corn | FB1: 10/20 | FB1: 10–5960 μg/kg | Morocco | 139 |
| Corn | FB1: 19/57 | FB1: 10–780 μg/kg | Egypt | 140 |
| FB1: 6.99/60 | FB1: 127–359 ng/g | |||
| Corn | Spain | 141 | ||
| FB2: 5.01/60 | FB2: 60–153 ng/g | |||
| Corn | FB1: 18/27 | FB1: 48–918 mg/L | Croatia | 142 |
| Corn | FB1: 80/184 | FB1: 0.21–3.30 μg/g | China | 14 |
| Corn | TFBs: 24/31 | TFBs: nd–34700 μg/kg | Argentina | 143 |
| Corn | FB1: 10/10 | FB1: 0.3–1.5 mg/kg | Côte d’Ivoire | 144 |
| FB1: 49/49 | FB1: 142.2–1377.6 μg/kg | |||
| Corn | Croatia | 145 | ||
| FB2: 3/49 | FB2: 68.4–3084.0 μg/kg | |||
| Corn | TFBs: 96/196 | TFBs: 2242 μg/kg (max) | Brazil | 146 |
| Corn | FB: 0/6 | FB: nd | Canada | 147 |
| Corn | FB: 41/42 | FB: 0.012–0.84 ppm | Argentina | 148 |
| FB1: 0/19 | FB1: nd | |||
| Wheat | Turkey | 6 | ||
| FB2: 0/19 | FB2: nd | |||
| Wheat | FB1: 4/210 | FB1: 1–2 ppm | S. Africa | 149 |
| Barley | FB: 0/10 | FB: nd | Canada | 147 |
| FB1: 0/1 | FB1: nd | |||
| Oat | Turkey | 6 | ||
| FB2: 0/1 | FB2: nd | |||
| Oat | FB: 0/5 | FB: nd | Canada | 147 |
| FB1: 0/31 | FB1: nd | |||
| Rice | Turkey | 6 | ||
| FB2: 0/31 | FB2: nd | |||
| Rice | FB1: 0/10 | FB1: nd | Côte d’Ivoire | 144 |
| Rice | FB1: 2/88 | FB1: 48.2–60.6 ng/g | Korea | 150 |
| Rice | FB: 1/25 | FB: 10 ng/g | Canada | 147 |
| FB1: 52.52/202 | FB1: 0.010–2.870 μg/g | |||
| Cereal | Italy | 151 | ||
| FB2: 70.7/202 | FB2: 0.010–0.790 μg/g | |||
| Cereal | FBs: 30.08/32 | FBs: 1–1110 μg/kg | France | 152 |
| Products | Total Fumonisins(FB1 + FB2 + FB3) (ppm) |
|---|---|
| Degermed dry milled corn products (e.g., flaking grits, corn grits, corn meal, corn flour with fat content of < 2.25 %, dry weight basis) | 2 |
| Whole or partially degermed dry milled corn products (e.g., flaking grits, corn grits, corn meal, corn flour with fat content of ≥2.25 %, dry weight basis) | 4 |
| Dry milled corn bran | 4 |
| Cleaned corn intended for masa production | 4 |
| Cleaned corn intended for popcorn | 3 |
| Country | Intake (μg/kg of body weight per day)
| |
|---|---|---|
| Mean or median | 90th percentile | |
| Argentina | 0.2 | nd |
| Canada | 0.02 | 0.08 |
| Netherlands | 0.06, 1.0a | nd |
| Switzerland | 0.03 | nd |
| United Kingdom | 0.03 | 0.1 |
| United States | 0.08 | nd |
| Product | Detected/total | Range | Countries | References |
|---|---|---|---|---|
| DON: 0/38 | DON: nd | |||
| Corn | Argentina | 153 | ||
| ZEA: 0/38 | ZEA: nd | |||
| DON: 0/78 | DON: nd–60 ng/g | |||
| NIV: 0/78 | NIV: nd–40 ng/g | |||
| Corn | DAS: 0/78 | DAS: 20–120 ng/g | Brazil | 154 |
| T-2: 1/78 | T-2: 20–100 ng/g | |||
| HT-2: 1/78 | HT-2: 20–100 ng/g | |||
| DON: 3/4 | DON: 0.065 μg/g | |||
| Corn | Poland | 155 | ||
| NIV: 3/4 | NIV: 0.13 μg/g | |||
| DON: 8/40 | DON: 204–745 μg/kg | |||
| Corn | Nigeria | 156 | ||
| DAS: 1.5/17 | DAS: 23–51 μg/kg | |||
| NIV: 6/31 | NIV: 12–2440 ng/g | |||
| DON: 6/31 | DON: 5–2060 ng/g | |||
| Corn | Italy (Central Italy) | 157 | ||
| FUS-X: 5/31 | FUS-X: 26–420 ng/g | |||
| ZEA: 5/31 | ZEA: nd–384 ng/g | |||
| NIV: 1/11 | NIV: 200 ng/g | |||
| DON: 11/11 | DON: 45–3430 ng/g | Italy (North-western Italy) | ||
| Corn | 157 | |||
| FUS-X: 1/11 | FUS-X: 34 ng/g | |||
| ZEA: 17/11 | ZEA: nd–969 ng/g | |||
| NIV: 0/4 | NIV: nd | |||
| DON: 3/4 | DON: 68–967 ng/g | |||
| Corn | Italy (Po valley) | 157 | ||
| FUS-X: 0/4 | FUS-X: nd | |||
| ZEA: 0/4 | ZEA: nd | |||
| DON: 4/21 | DON: nd- 834.4 μg/kg | |||
| Corn | Argentina | 143 | ||
| ZEA: 11/23 | ZEA: nd- 2564.8 μg/kg | |||
| Corn | T-2: 8/30 | T-2: 0.45–1.70 ppm | Turkey | 81 |
| DON: 47/175 | DON: 26.1–131.7 μg/kg | |||
| Corn | Spain | 158 | ||
| NIV: 7/175 | NIV: 51.1–106.5 μg/kg | |||
| Corn | ZEA: 3/20 | ZEA: 12–17 μg/kg | Morocco | 139 |
| Corn | TTCs: 42/196 | TTCs: <2–600 μg/kg | Saudi Arabia | 159 |
| Corn | ZEA: 10/10 | ZEA: 20–50 μg/kg | Côte d’Ivoire | 144 |
| Corn | ZEA: 41/49 | ZEA: 0.9–2.54 μg/kg | Croatia | 145 |
| DON: 0/6 | DON: nd | |||
| Corn | ZEA: 0/6 | ZEA: nd | Canada | 147 |
| NIV-HT-2: 0/6 | NIV-HT-2: nd | |||
| DON: 58.8/60 | DON: 15–1379 μg/kg | |||
| NIV: 7.2/60 | NIV: 25–40 μg/kg | |||
| HT-2: 4.2/60 | HT-2: 12 μg/kg | |||
| Wheat | Germany | 160 | ||
| T-2: 1.2/60 | T-2: 4 μg/kg | |||
| ZEA: 22.8/60 | ZEA: 1–24 μg/kg | |||
| FUS-X: 0/60 | FUS-X: 0 | |||
| DON: 108/121 | DON: 10–2591 μg/kg | |||
| NIV: 57/120 | NIV: 10–234 μg/kg | |||
| Wheat | HT-2: 6/36 | HT-2: nd–33 μg/kg | Denmark | 161 |
| T-2: 11/38 | T-2: nd–153 μg/kg | |||
| ZEA: 10/30 | ZEA: 1–2 μg/kg | |||
| Wheat | DON: 8/12 | DON: 0.01 μg/g | Poland | 155 |
| DON: 144/150 | DON: Tr-642 μg/kg | |||
| Wheat | Lithuania | 162 | ||
| ZEA: 32/100 | ZEA: Tr-95.6 μg/kg | |||
| Wheat | T-2: 0/3 | T-2: nd | Turkey | 81 |
| Wheat | TTCs: 0/1 | TTCs: nd | Saudi Arabia | 159 |
| DON: 23.17/169 | DON: 350 μg/kg (max) | |||
| NIV: 0/169 | NIV: 10 μg/kg (max) | |||
| Wheat | Norway | 163 | ||
| HT-2: 1.86/169 | HT-2: 20 μg/kg (max) | |||
| T-2: 1.24/169 | T-2: 20 μg/kg (max) | |||
| Wheat | ZEA: 22/24 | ZEA: 11–860 μg/kg | Germany | 164 |
| DON: 10/10 | DON: 0.025 μg/g | |||
| Barley | Poland | 155 | ||
| NIV: 10/10 | NIV: 0.04 μg/g | |||
| DON: 53/55 | DON: Tr-372 μg/kg | |||
| Barley | Lithuania | 162 | ||
| ZEA: 38/66 | ZEA: Tr-193.4 μg/kg | |||
| Barley | DON: 157.45/292 | DON: 500–10.000 μg/kg | Uruguay | 165 |
| Barley | TTCs: 8/93 | TTCs: 3.1–4.000 μg/kg | Saudi Arabia | 159 |
| DON: 17.6/102 | DON: 1440μg/kg (max) | |||
| NIV: 11.2/102 | NIV: 50 μg/kg (max) | |||
| Barley | Norway | 163 | ||
| HT-2: 39.78/102 | HT-2: 440 μg/kg (max) | |||
| T-2: 8.16/102 | T-2: 220 μg/kg (max) | |||
| Barley | DON-NIV: 41/84 | DON-NIV: 40–2340 μg/kg | Ethiopia | 166 |
| DON: 29/50 | DON: 980 ng/g (max) | |||
| Barley | ZEA: 4/29 | ZEA: 22 ng/g (max) | Canada | 147 |
| NIV-HT-2: 0/75 | NIV-HT-2: nd | |||
| HT-2:24/99 | HT-2: 10–47 μg/kg | |||
| Oats | T-2:15/99 | T-2: 10–703 μg/kg | Poland | 167 |
| ZEA:19/99 | ZEA: 10–118 μg/kg | |||
| DON: 12/12 | DON: 0.027 μg/g | |||
| Oats | Poland | 167 | ||
| NIV: 12/12 | NIV: 0.033 μg/g | |||
| DON: 13/14 | DON: Tr-204 μg/kg | |||
| Oats | ZEA: 4/7 | ZEA: Tr-16.3 μg/kg | Lithuania | 162 |
| T-2: 10/10 | T-2: 10.8–121.5 μg/kg | |||
| Oats | T-2: 0/1 | T-2: nd | Turkey | 81 |
| HT-2: 24/99 | HT-2: 10–47 μg/kg | |||
| Oats | T-2: 15/99 | T-2: 10–703 μg/kg | Poland | 167 |
| DAS: 12/99 | DAS: 10–118 μg/kg | |||
| Oats | TTCs: 0/3 | TTCs:nd | Saudi Arabia | 159 |
| DON: 101.96/178 | DON: 849 μg/kg (max) | |||
| NIV: 17.6/178 | NIV: 211 μg/kg (max) | |||
| Oats | Norway | 163 | ||
| HT-2: 124.04/178 | HT-2: 880 μg/kg (max) | |||
| T-2: 53.32/178 | T-2: 380 μg/kg (max) | |||
| DON: 33/53 | DON: 90 ng/g (max) | |||
| Oats | ZEA: 0/6 | ZEA: nd | Canada | 147 |
| NIV-HT-2: 0/53-0/53 | NIV-HT-2: nd | |||
| DON: 10/10 | DON: 0.023 μg/g | |||
| Rye | Poland | 155 | ||
| NIV: 5/10 | NIV: 0.002 μg/g | |||
| DON: 14/16 | DON: Tr-691 μg/kg | |||
| Rye | Lithuania | 162 | ||
| ZEA: 0/10 | ZEA: 0–28.8 μg/kg | |||
| DON: 41/69 | DON: 10–257 μg/kg | |||
| NIV: 9/69 | NIV: nd–48 μg/kg | |||
| Rye | HT-2: 11/26 | HT-2: 10–70 μg/kg | Denmark | 161 |
| T-2: 12/25 | T-2: nd–193 μg/kg | |||
| ZEA: 2/30 | ZEA: 1–2 μg/kg | |||
| DON: 3/88 | DON: 105– 159 ng/g | |||
| Rice | NIV: 5/88 | NIV: 182– 462 ng/g | Korea | 150 |
| ZEA: 3/88 | ZEA: 21.7– 47.0 ng/g | |||
| Rice | TTCs: 0/2 | TTCs: nd | Saudi Arabia | 159 |
| Rice | ZEA: 10/10 | ZEA: 50–200 μg/kg | Côte d’Ivoire | 144 |
| DON: 0/9 | DON: nd | |||
| Rice | ZEA: 1/7 | ZEA: 1 ng/g (max) | Canada | 147 |
| NIV-HT-2: 0/27 | NIV-HT-2: nd | |||
| Cereals | DAS: 0/69 | DAS: nd- 0.8 μg/g | Turkey | 106 |
| Cereals | DON: 169.68/202 | DON: 0.007–0930 μg/g | Italy | 151 |
| Cereals | T-2: 2/30 | T-2: 1.60–33.30 ppm | Turkey | 168 |
| DON: 12/12 | DON: 16–51 450 μg/kg | |||
| Cereals | NIV: 10/12 | NIV: 10–6935 μg/kg | China | 169 |
| ZEA: 10/12 | ZEA: 46–3079 μg/kg | |||
| DON:54/68 | DON: 5–111 μg/kg | |||
| Cereals | NIV:3/68 | NIV: 10–20 μg/kg | Finland | 170 |
| HT-2:2/68 | HT-2: 10–20 μg/kg | |||
| DON: 126/169 | DON: 15–1670 μg/kg | |||
| NIV: 0.21/4 | NIV: 25–231 μg/kg | |||
| Cereals | FUS-X: 0 | FUS-X: nd | Germany | 171 |
| HT2: 11.38/43 | HT-2: 12–51 μg/kg | |||
| T-2: 1.43/10 | T-2: 4–39 μg/kg |
© 2008 by MDPI This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yazar, S.; Omurtag, G.Z. Fumonisins, Trichothecenes and Zearalenone in Cereals. Int. J. Mol. Sci. 2008, 9, 2062-2090. https://doi.org/10.3390/ijms9112062
Yazar S, Omurtag GZ. Fumonisins, Trichothecenes and Zearalenone in Cereals. International Journal of Molecular Sciences. 2008; 9(11):2062-2090. https://doi.org/10.3390/ijms9112062
Chicago/Turabian StyleYazar, Selma, and Gülden Z. Omurtag. 2008. "Fumonisins, Trichothecenes and Zearalenone in Cereals" International Journal of Molecular Sciences 9, no. 11: 2062-2090. https://doi.org/10.3390/ijms9112062
APA StyleYazar, S., & Omurtag, G. Z. (2008). Fumonisins, Trichothecenes and Zearalenone in Cereals. International Journal of Molecular Sciences, 9(11), 2062-2090. https://doi.org/10.3390/ijms9112062