
CRISPR/Cas9 Ribonucleoprotein Delivery Enhanced by Lipo-Xenopeptide Carriers and Homology-Directed Repair Modulators: Insights from Reporter Cell Lines


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
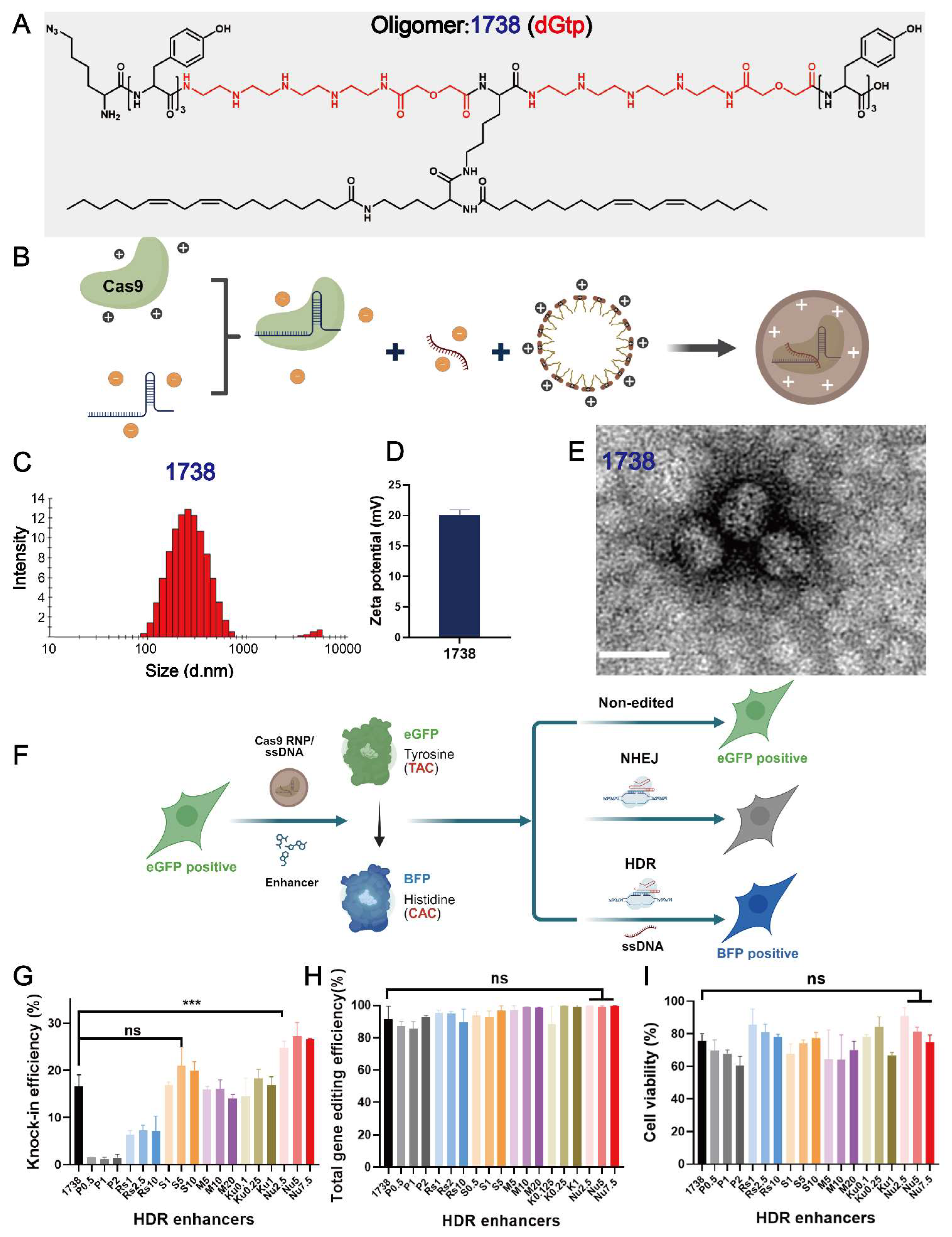
2.1. Characterization of T-Shaped Cas9 RNP/ssDNA Complexes and Selection of Enhancers
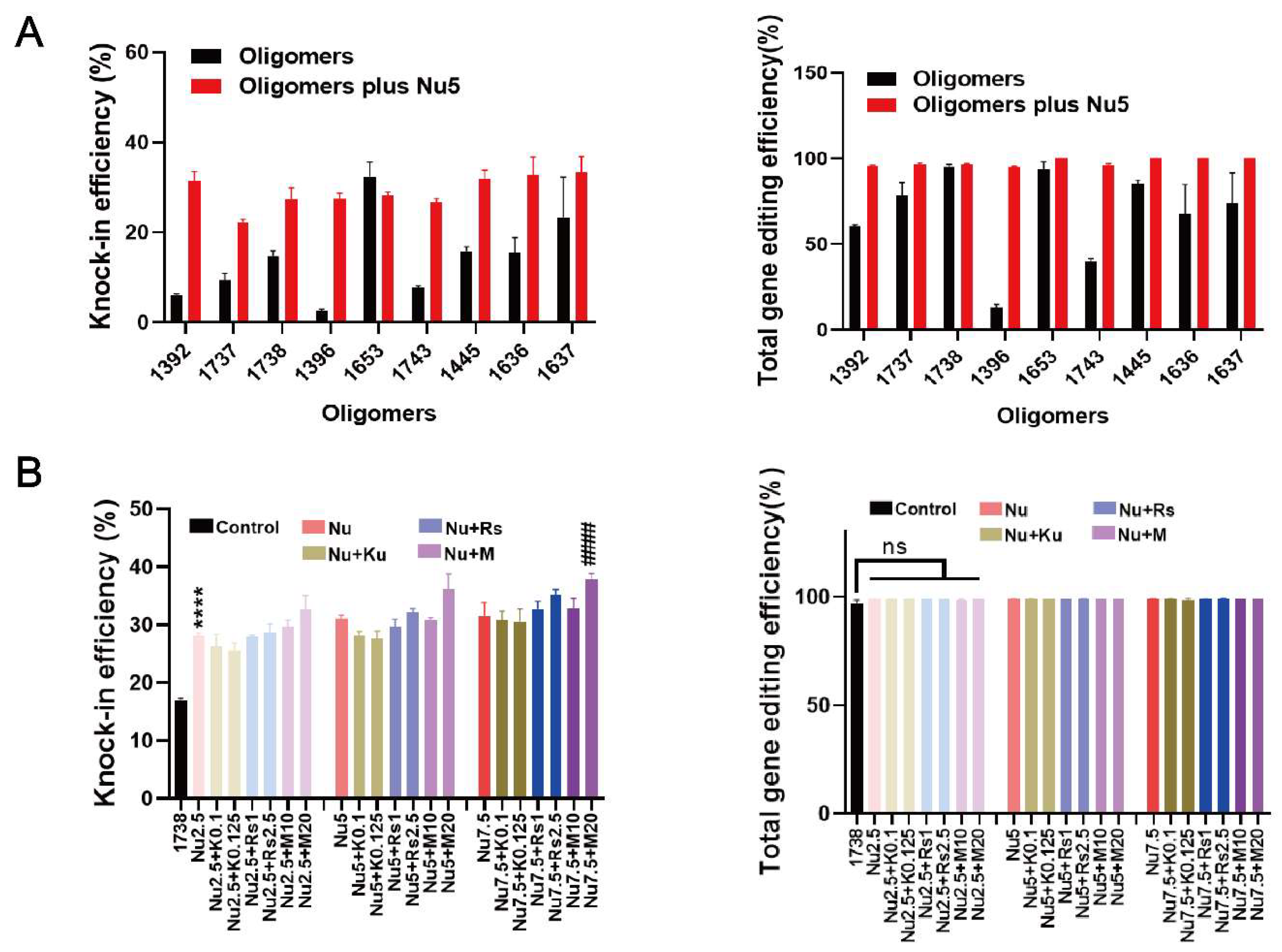
2.2. HDR and Gene Editing Efficiency with Various T-Shaped XPs or Enhancer Combinations
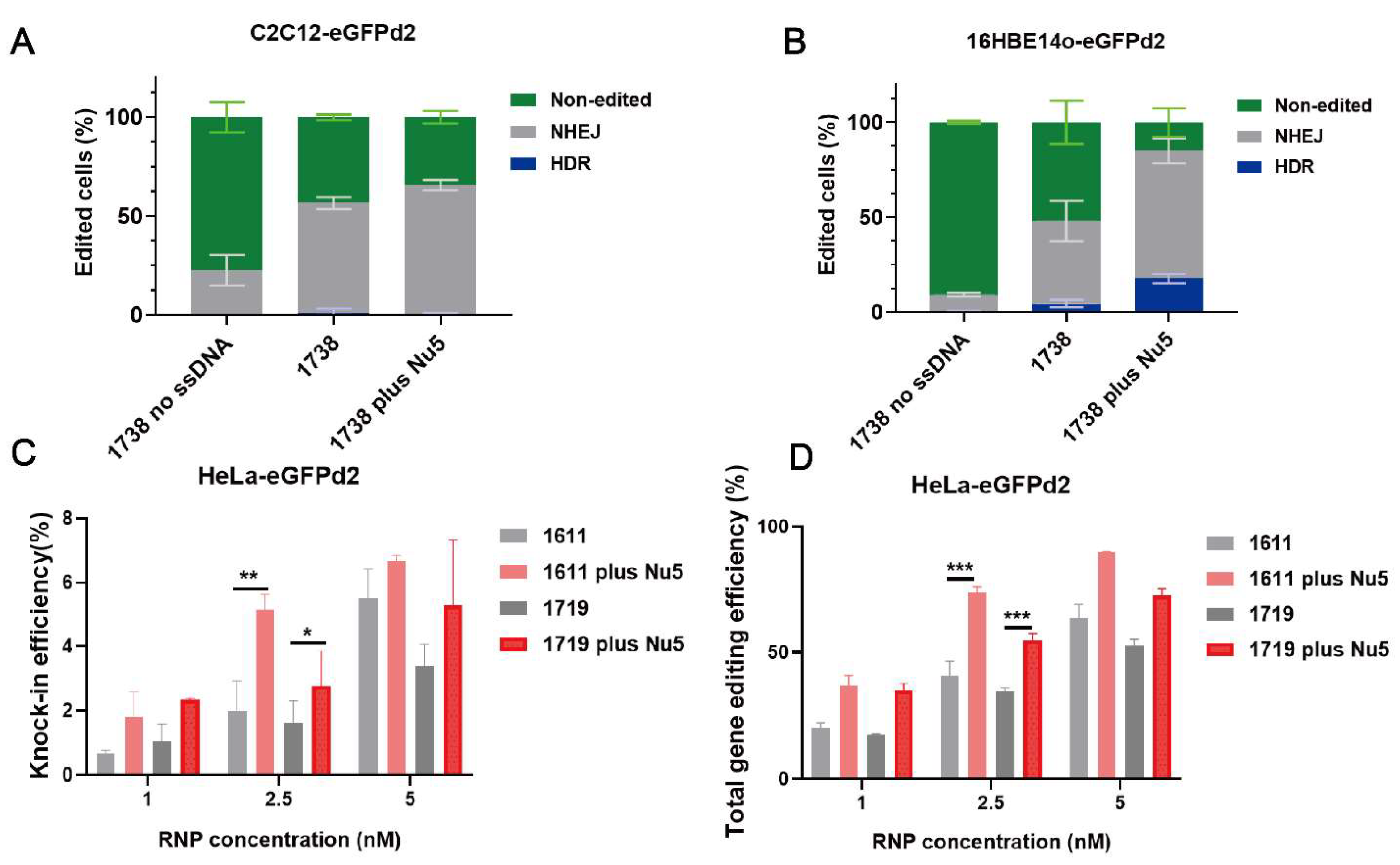
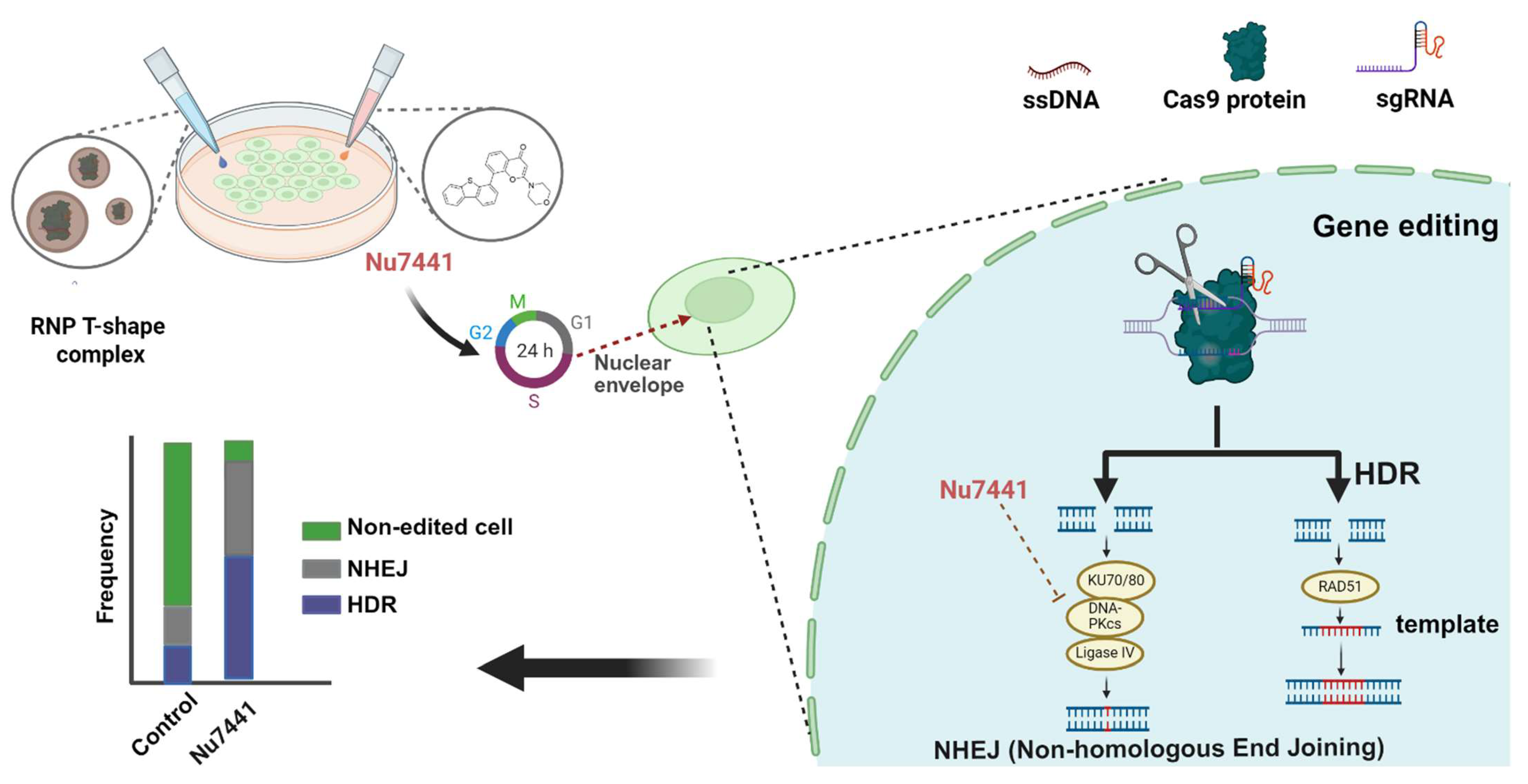
2.3. Mechanistic Investigation of Nu7441 for Gene Editing Improvement
2.4. Exon Skipping Efficiency of 1738 Cas9 RNP Complexes with 5 nM Nu7441 in DMD Exon 23 Reporter Cell Model
2.5. General Applicability of Nu7441 in Different Settings
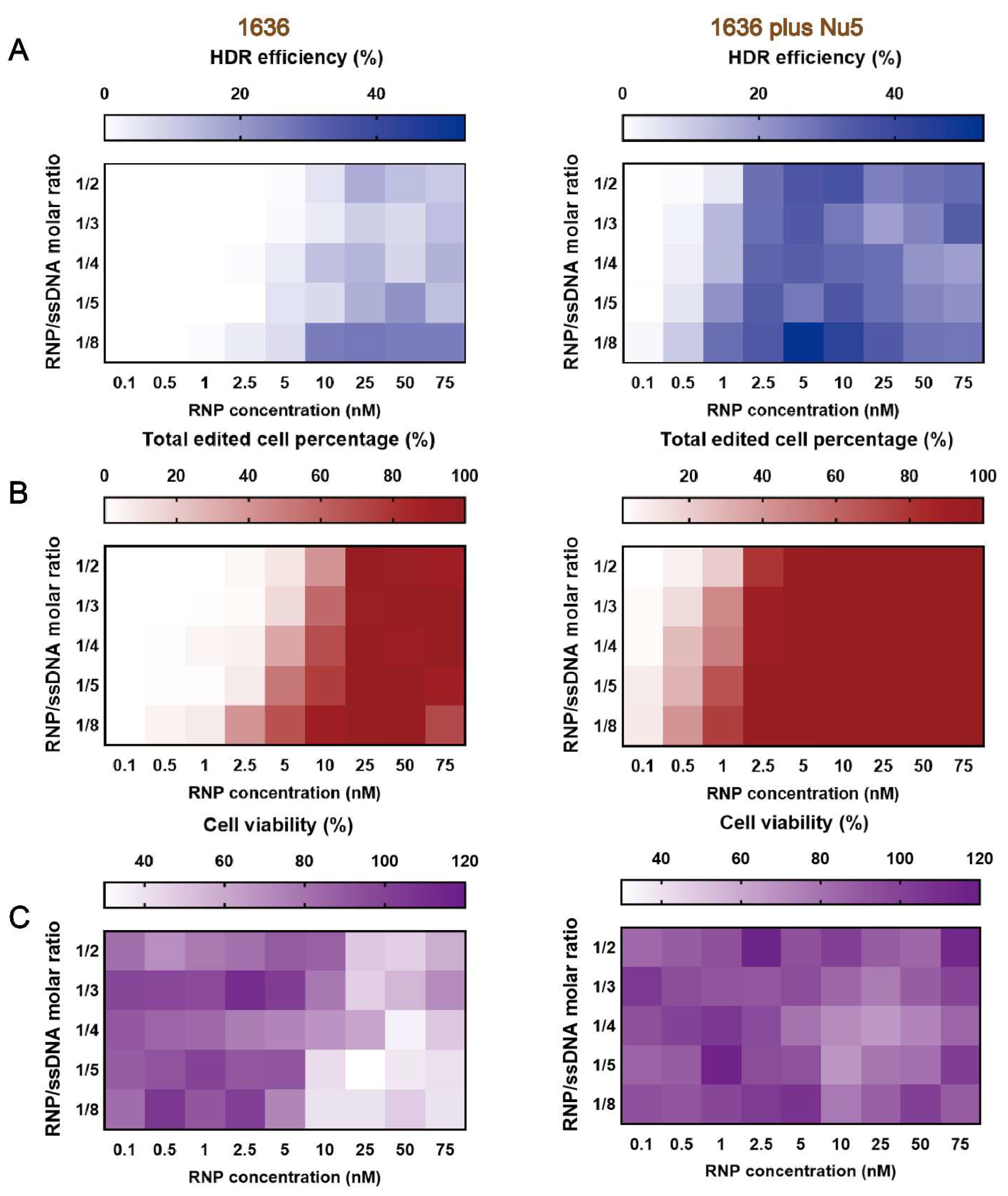
2.6. Gene Correction Efficiency of Optimized T-Shaped Carrier 1636-Based Cas9 RNP/ssDNA Formulation with Nu7441
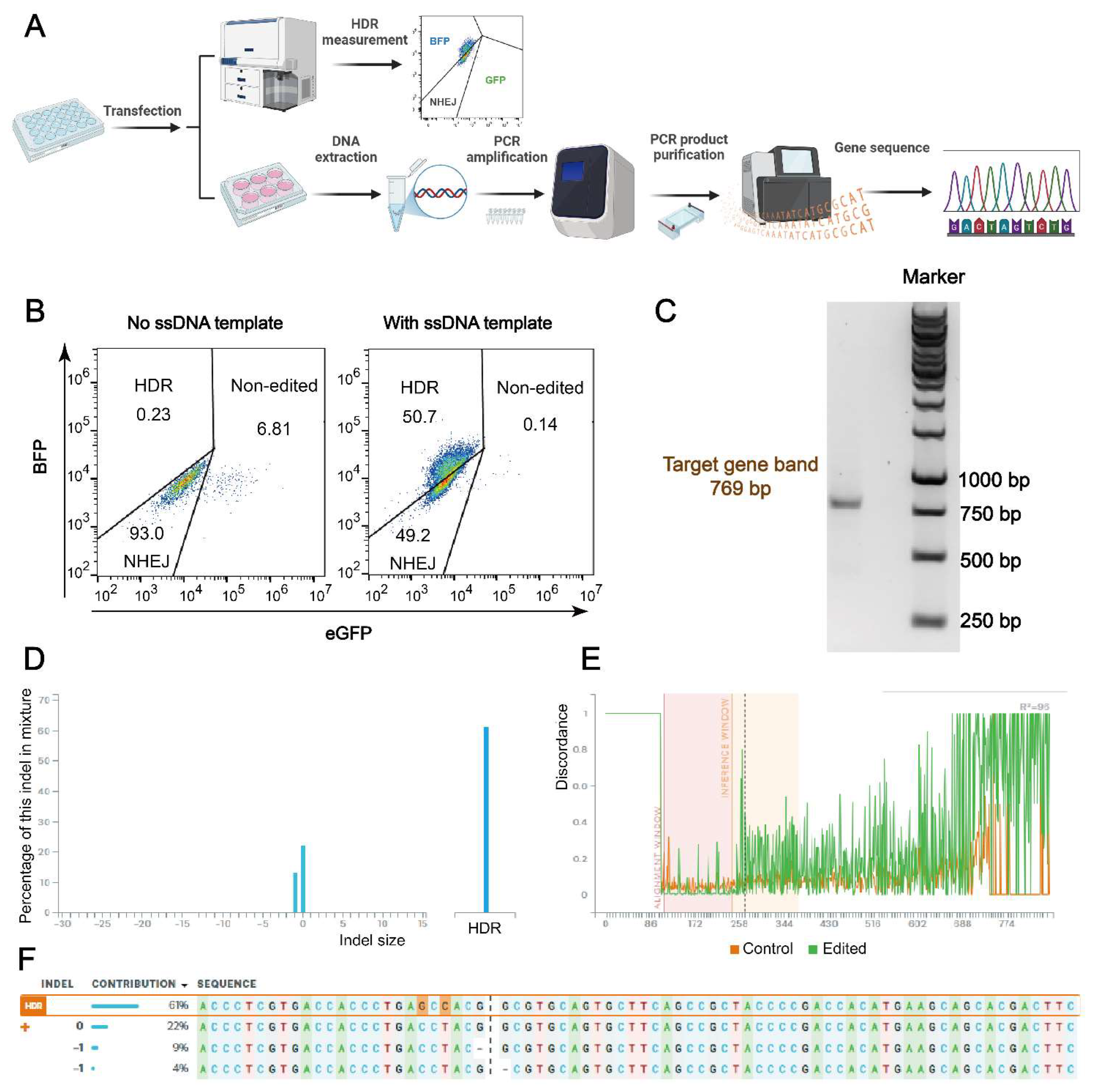
2.7. Comparison of HDR Efficiency in Flow Cytometry and Gene Sequence Measurement
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Fabrication of Cas9 Ribonucleoprotein (RNP) Complexes or mRNA/sgRNA Polyplexes
4.3. Characterization of Cas9 RNP/ssDNA Complexes
4.4. GFP-to-BFP Conversion Mediated via Homology-Directed Repair (HDR)
4.5. Cellular Uptake and Endosomal Escape of Cas9 RNP/ssDNA Complexes
4.6. Cellular Cell Cycle Assay
4.7. Gene Editing-Mediated Exon Skipping of Cas9 RNP Complexes with or Without Nu7441
4.8. Gene Editing-Mediated Exon Skipping by Cas9 RNP Complexes Visualized by CLSM
4.9. Genome Extraction
4.10. PCR Amplification and Sequencing
4.11. Assessment of Cytotoxicity of Cells via MTT Assay
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Cas9 | CRISPR-associated protein 9 |
| chGtp | Cyclohexyl-glutaryl-tetraethylene pentamine |
| CLSM | Confocal laser scanning microscopy |
| CRISPR | Clustered regularly interspaced short palindromic repeat |
| DAPI | 4′,6-Diamidino-2-phenylindole |
| DSBs | Double-strand breaks |
| DNA-PKcs | DNA-dependent protein kinase |
| dGtp | Diglycoloyl tetraethylene pentamine |
| DLS | Dynamic light scattering |
| DMD | Duchenne muscular dystrophy |
| DMEM | Dulbecco’s modified Eagle’s medium |
| DMSO | Dimethyl sulfoxide |
| FACS | Flow cytometry staining |
| FBS | Fetal bovine serum |
| LAF | Lipo-amino fatty acid |
| GEIPA | Glutaryl-ethylenediImine-dipropylamine |
| Gtt | Glutaryl-triethylene tetramine |
| HBG | HEPES-buffered glucose solution |
| HEPES | 4-(2-Hydroxyethyl)-1-piperazineethanesulfonic acid |
| HDR | Homology-directed repair |
| N/P | Nitrogen-to-phosphate |
| NHEJ | Non-homologous end joining |
| NLRP3 | NOD-like receptor family pyrin domain containing 3 |
| MEM | Minimum essential medium |
| MFI | Median fluorescence intensity |
| MTT | 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| sgRNA | Single guide RNA |
| ssDNA | Single-stranded DNA |
| PD-L1 | Programmed cell death ligand 1 |
| PI | Propidium iodide |
| RNP | Ribonucleoprotein |
| Stp | Succinyl-tetraethylene pentamine |
| TEM | Transmission electron microscopy |
| TFE | Trifluoroethyl-iminodiacetyl-tetraethylene pentamine |
| XP | Xenopeptide |
References
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA–guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Mali, P.; Yang, L.; Esvelt, K.M.; Aach, J.; Guell, M.; DiCarlo, J.E.; Norville, J.E.; Church, G.M. RNA-guided human genome engineering via Cas9. Science 2013, 339, 823–826. [Google Scholar] [CrossRef]
- Chapman, J.R.; Taylor, M.R.; Boulton, S.J. Playing the end game: DNA double-strand break repair pathway choice. Mol. Cell. 2012, 47, 497–510. [Google Scholar] [CrossRef] [PubMed]
- Chiruvella, K.K.; Liang, Z.; Wilson, T.E. Repair of double-strand breaks by end joining. Cold Spring Harb. Perspect. Biol. 2013, 5, a012757. [Google Scholar] [CrossRef] [PubMed]
- An, S.Y.; Ka, D.; Kim, I.; Kim, E.-H.; Kim, N.-K.; Bae, E.; Suh, J.-Y. Intrinsic disorder is essential for Cas9 inhibition of anti-CRISPR AcrIIA5. Nucleic Acids Res. 2020, 48, 7584–7594. [Google Scholar] [CrossRef] [PubMed]
- Farbiak, L.; Cheng, Q.; Wei, T.; Álvarez-Benedicto, E.; Johnson, L.T.; Lee, S.; Siegwart, D.J. All-in-one dendrimer-based lipid nanoparticles enable precise HDR-mediated gene editing in vivo. Adv. Mater. 2021, 33, 2006619. [Google Scholar] [CrossRef]
- Wei, T.; Sun, Y.; Cheng, Q.; Chatterjee, S.; Traylor, Z.; Johnson, L.T.; Coquelin, M.L.; Wang, J.; Torres, M.J.; Lian, X. Lung SORT LNPs enable precise homology-directed repair mediated CRISPR/Cas genome correction in cystic fibrosis models. Nat. Commun. 2023, 14, 7322. [Google Scholar] [CrossRef]
- Chen, K.; Han, H.; Zhao, S.; Xu, B.; Yin, B.; Lawanprasert, A.; Trinidad, M.; Burgstone, B.W.; Murthy, N.; Doudna, J.A. Lung and liver editing by lipid nanoparticle delivery of a stable CRISPR–Cas9 ribonucleoprotein. Nat. Biotechnol. 2024, 1–13. [Google Scholar] [CrossRef]
- Shy, B.R.; Vykunta, V.S.; Ha, A.; Talbot, A.; Roth, T.L.; Nguyen, D.N.; Pfeifer, W.G.; Chen, Y.Y.; Blaeschke, F.; Shifrut, E. High-yield genome engineering in primary cells using a hybrid ssDNA repair template and small-molecule cocktails. Nat. Biotechnol. 2023, 41, 521–531. [Google Scholar] [CrossRef]
- Selvaraj, S.; Feist, W.N.; Viel, S.; Vaidyanathan, S.; Dudek, A.M.; Gastou, M.; Rockwood, S.J.; Ekman, F.K.; Oseghale, A.R.; Xu, L. High-efficiency transgene integration by homology-directed repair in human primary cells using DNA-PKcs inhibition. Nat. Biotechnol. 2024, 42, 731–744. [Google Scholar] [CrossRef]
- Fu, Y.-W.; Dai, X.-Y.; Wang, W.-T.; Yang, Z.-X.; Zhao, J.-J.; Zhang, J.-P.; Wen, W.; Zhang, F.; Oberg, K.C.; Zhang, L. Dynamics and competition of CRISPR–Cas9 ribonucleoproteins and AAV donor-mediated NHEJ, MMEJ and HDR editing. Nucleic Acids Res. 2021, 49, 969–985. [Google Scholar] [CrossRef] [PubMed]
- Komor, A.C.; Kim, Y.B.; Packer, M.S.; Zuris, J.A.; Liu, D.R. Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature 2016, 533, 420–424. [Google Scholar] [CrossRef]
- Gaudelli, N.M.; Komor, A.C.; Rees, H.A.; Packer, M.S.; Badran, A.H.; Bryson, D.I.; Liu, D.R. Programmable base editing of A• T to G• C in genomic DNA without DNA cleavage. Nature 2017, 551, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Scholefield, J.; Harrison, P.T. Prime editing–an update on the field. Gene Ther. 2021, 28, 396–401. [Google Scholar] [CrossRef]
- Nambiar, T.S.; Baudrier, L.; Billon, P.; Ciccia, A. CRISPR-based genome editing through the lens of DNA repair. Mol. Cell. 2022, 82, 348–388. [Google Scholar] [CrossRef] [PubMed]
- Schumann, K.; Lin, S.; Boyer, E.; Simeonov, D.R.; Subramaniam, M.; Gate, R.E.; Haliburton, G.E.; Ye, C.J.; Bluestone, J.A.; Doudna, J.A. Generation of knock-in primary human T cells using Cas9 ribonucleoproteins. Proc. Natl. Acad. Sci. USA 2015, 112, 10437–10442. [Google Scholar] [CrossRef]
- Kim, K.; Ryu, S.-M.; Kim, S.-T.; Baek, G.; Kim, D.; Lim, K.; Chung, E.; Kim, S.; Kim, J.-S. Highly efficient RNA-guided base editing in mouse embryos. Nat. Biotechnol. 2017, 35, 435–437. [Google Scholar] [CrossRef]
- Merienne, N.; Vachey, G.; de Longprez, L.; Meunier, C.; Zimmer, V.; Perriard, G.; Canales, M.; Mathias, A.; Herrgott, L.; Beltraminelli, T. The self-inactivating KamiCas9 system for the editing of CNS disease genes. Cell Rep. 2017, 20, 2980–2991. [Google Scholar] [CrossRef]
- Xu, L.; Park, K.H.; Zhao, L.; Xu, J.; El Refaey, M.; Gao, Y.; Zhu, H.; Ma, J.; Han, R. CRISPR-mediated genome editing restores dystrophin expression and function in mdx mice. Mol. Ther. 2016, 24, 564–569. [Google Scholar] [CrossRef]
- Ibraheim, R.; Tai, P.W.; Mir, A.; Javeed, N.; Wang, J.; Rodríguez, T.C.; Namkung, S.; Nelson, S.; Khokhar, E.S.; Mintzer, E. Self-inactivating, all-in-one AAV vectors for precision Cas9 genome editing via homology-directed repair in vivo. Nat. Commun. 2021, 12, 6267. [Google Scholar] [CrossRef]
- Cheng, H.; Zhang, F.; Ding, Y. CRISPR/Cas9 delivery system engineering for genome editing in therapeutic applications. Pharmaceutics 2021, 13, 1649. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Ping, Y. Development of CRISPR/Cas delivery systems for in vivo precision genome editing. Acc. Chem. Res. 2023, 56, 2185–2196. [Google Scholar] [CrossRef]
- Lächelt, U.; Wagner, E. Nucleic acid therapeutics using polyplexes: A journey of 50 years (and beyond). Chem. Rev. 2015, 115, 11043–11078. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Wagner, E. Polymeric carriers for nucleic acid delivery: Current designs and future directions. Biomacromolecules 2019, 20, 3613–3626. [Google Scholar] [CrossRef]
- Kumar, R.; Santa Chalarca, C.F.; Bockman, M.R.; Bruggen, C.V.; Grimme, C.J.; Dalal, R.J.; Hanson, M.G.; Hexum, J.K.; Reineke, T.M. Polymeric delivery of therapeutic nucleic acids. Chem. Rev. 2021, 121, 11527–11652. [Google Scholar] [CrossRef] [PubMed]
- Paunovska, K.; Sago, C.D.; Monaco, C.M.; Hudson, W.H.; Castro, M.G.; Rudoltz, T.G.; Kalathoor, S.; Vanover, D.A.; Santangelo, P.J.; Ahmed, R. A direct comparison of in vitro and in vivo nucleic acid delivery mediated by hundreds of nanoparticles reveals a weak correlation. Nano Lett. 2018, 18, 2148–2157. [Google Scholar] [CrossRef]
- Lin, Y.; Wagner, E.; Lächelt, U. Non-viral delivery of the CRISPR/Cas system: DNA versus RNA versus RNP. Biomater. Sci. 2022, 10, 1166–1192. [Google Scholar] [CrossRef]
- Cullis, P.; Felgner, P. The 60-year evolution of lipid nanoparticles for nucleic acid delivery. Nat. Rev. Drug Discov. 2024, 23, 709–722. [Google Scholar] [CrossRef]
- Cabral, H.; Miyata, K.; Osada, K.; Kataoka, K. Block copolymer micelles in nanomedicine applications. Chem. Rev. 2018, 118, 6844–6892. [Google Scholar] [CrossRef]
- Abbasi, S.; Uchida, S.; Toh, K.; Tockary, T.A.; Dirisala, A.; Hayashi, K.; Fukushima, S.; Kataoka, K. Co-encapsulation of Cas9 mRNA and guide RNA in polyplex micelles enables genome editing in mouse brain. J. Control. Release. 2021, 332, 260–268. [Google Scholar] [CrossRef]
- Öktem, M.; Mastrobattista, E.; de Jong, O.G. Amphipathic cell-penetrating peptide-aided delivery of Cas9 RNP for in vitro gene editing and correction. Pharmaceutics 2023, 15, 2500. [Google Scholar] [CrossRef] [PubMed]
- Onuma, H.; Shimizu, R.; Suzuki, Y.; Sato, M.; Harashima, H.; Sato, Y. Engineering branched ionizable lipid for hepatic delivery of clustered regularly interspaced short palindromic repeat-Cas9 ribonucleoproteins. iScience 2024, 27, 110928. [Google Scholar] [CrossRef]
- Onuma, H.; Sato, Y.; Harashima, H. Lipid nanoparticle-based ribonucleoprotein delivery for in vivo genome editing. J. Control. Release. 2023, 355, 406–416. [Google Scholar] [CrossRef]
- Zhang, D.; Wang, G.; Yu, X.; Wei, T.; Farbiak, L.; Johnson, L.T.; Taylor, A.M.; Xu, J.; Hong, Y.; Zhu, H. Enhancing CRISPR/Cas gene editing through modulating cellular mechanical properties for cancer therapy. Nat. Nanotechnol. 2022, 17, 777–787. [Google Scholar] [CrossRef] [PubMed]
- Tan, E.; Wan, T.; Pan, Q.; Duan, J.; Zhang, S.; Wang, R.; Gao, P.; Lv, J.; Wang, H.; Li, D. Dual-responsive nanocarriers for efficient cytosolic protein delivery and CRISPR-Cas9 gene therapy of inflammatory skin disorders. Sci. Adv. 2024, 10, eadl4336. [Google Scholar] [CrossRef] [PubMed]
- Schaffert, D.; Troiber, C.; Salcher, E.E.; Fröhlich, T.; Martin, I.; Badgujar, N.; Dohmen, C.; Edinger, D.; Kläger, R.; Maiwald, G. Solid-phase synthesis of sequence-defined T-, i-, and U-shape polymers for pDNA and siRNA delivery. Angew. Chem. Int. Ed. 2011, 50, 8986. [Google Scholar] [CrossRef] [PubMed]
- Freitag, F.; Wagner, E. Optimizing synthetic nucleic acid and protein nanocarriers: The chemical evolution approach. Adv. Drug Deliv. Rev. 2021, 168, 30–54. [Google Scholar] [CrossRef]
- Lin, Y.; Wilk, U.; Pöhmerer, J.; Hörterer, E.; Höhn, M.; Luo, X.; Mai, H.; Wagner, E.; Lächelt, U. Folate receptor-mediated delivery of Cas9 RNP for enhanced immune checkpoint disruption in cancer cells. Small 2023, 19, 2205318. [Google Scholar] [CrossRef]
- Lin, Y.; Luo, X.; Burghardt, T.; Dorrer, S.; Höhn, M.; Wagner, E.; Lächelt, U. Chemical evolution of amphiphilic xenopeptides for potentiated Cas9 ribonucleoprotein delivery. J. Am. Chem. Soc. 2023, 145, 15171–15179. [Google Scholar] [CrossRef]
- Shams, F.; Bayat, H.; Mohammadian, O.; Mahboudi, S.; Vahidnezhad, H.; Soosanabadi, M.; Rahimpour, A. Advance trends in targeting homology-directed repair for accurate gene editing: An inclusive review of small molecules and modified CRISPR-Cas9 systems. BioImpacts 2022, 12, 371. [Google Scholar] [CrossRef]
- Waters, C.A.; Strande, N.T.; Wyatt, D.W.; Pryor, J.M.; Ramsden, D.A. Nonhomologous end joining: A good solution for bad ends. DNA Repair 2014, 17, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Radhakrishnan, S.K.; Lees-Miller, S.P. DNA requirements for interaction of the C-terminal region of Ku80 with the DNA-dependent protein kinase catalytic subunit (DNA-PKcs). DNA Repair 2017, 57, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Weterings, E.; Gallegos, A.C.; Dominick, L.N.; Cooke, L.S.; Bartels, T.N.; Vagner, J.; Matsunaga, T.O.; Mahadevan, D. A novel small molecule inhibitor of the DNA repair protein Ku70/80. DNA Repair 2016, 43, 98–106. [Google Scholar] [CrossRef]
- Kostyushev, D.; Kostyusheva, A.; Brezgin, S.; Zarifyan, D.; Utkina, A.; Goptar, I.; Chulanov, V. Suppressing the NHEJ pathway by DNA-PKcs inhibitor NU7026 prevents degradation of HBV cccDNA cleaved by CRISPR/Cas9. Sci. Rep. 2019, 9, 1847. [Google Scholar] [CrossRef]
- Maruyama, T.; Dougan, S.K.; Truttmann, M.C.; Bilate, A.M.; Ingram, J.R.; Ploegh, H.L. Increasing the efficiency of precise genome editing with CRISPR-Cas9 by inhibition of nonhomologous end joining. Nat. Biotechnol. 2015, 33, 538–542. [Google Scholar] [CrossRef]
- Robert, F.; Barbeau, M.; Éthier, S.; Dostie, J.; Pelletier, J. Pharmacological inhibition of DNA-PK stimulates Cas9-mediated genome editing. Genome Med. 2015, 7, 93. [Google Scholar] [CrossRef]
- Budke, B.; Lv, W.; Kozikowski, A.P.; Connell, P.P. Recent developments using small molecules to target RAD51: How to best modulate RAD51 for anticancer therapy? ChemMedChem 2016, 11, 2468–2473. [Google Scholar] [CrossRef]
- Riesenberg, S.; Maricic, T. Targeting repair pathways with small molecules increases precise genome editing in pluripotent stem cells. Nat. Commun. 2018, 9, 2164. [Google Scholar] [CrossRef] [PubMed]
- Jayavaradhan, R.; Pillis, D.M.; Malik, P. A versatile tool for the quantification of CRISPR/Cas9-induced genome editing events in human hematopoietic cell lines and hematopoietic stem/progenitor cells. J. Mol. Biol. 2019, 431, 102–110. [Google Scholar] [CrossRef]
- Song, J.; Yang, D.; Xu, J.; Zhu, T.; Chen, Y.E.; Zhang, J. RS-1 enhances CRISPR/Cas9-and TALEN-mediated knock-in efficiency. Nat. Commun. 2016, 7, 10548. [Google Scholar] [CrossRef]
- Hu, Z.; Shi, Z.; Guo, X.; Jiang, B.; Wang, G.; Luo, D.; Chen, Y.; Zhu, Y.-S. Ligase IV inhibitor SCR7 enhances gene editing directed by CRISPR–Cas9 and ssODN in human cancer cells. Cell Biosci. 2018, 8, 12. [Google Scholar] [CrossRef] [PubMed]
- Greco, G.E.; Matsumoto, Y.; Brooks, R.C.; Lu, Z.; Lieber, M.R.; Tomkinson, A.E. SCR7 is neither a selective nor a potent inhibitor of human DNA ligase IV. DNA Repair 2016, 43, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Gutschner, T.; Haemmerle, M.; Genovese, G.; Draetta, G.F.; Chin, L. Post-translational regulation of Cas9 during G1 enhances homology-directed repair. Cell Rep. 2016, 14, 1555–1566. [Google Scholar] [CrossRef]
- Riesenberg, S.; Chintalapati, M.; Macak, D.; Kanis, P.; Maricic, T.; Pääbo, S. Simultaneous precise editing of multiple genes in human cells. Nucleic Acids Res. 2019, 47, e116. [Google Scholar] [CrossRef] [PubMed]
- Bermudez-Cabrera, H.C.; Culbertson, S.; Barkal, S.; Holmes, B.; Shen, M.W.; Zhang, S.; Gifford, D.K.; Sherwood, R.I. Small molecule inhibition of ATM kinase increases CRISPR-Cas9 1-bp insertion frequency. Nat. Commun. 2021, 12, 5111. [Google Scholar] [CrossRef]
- Zhang, J.-P.; Li, X.-L.; Li, G.-H.; Chen, W.; Arakaki, C.; Botimer, G.D.; Baylink, D.; Zhang, L.; Wen, W.; Fu, Y.-W. Efficient precise knockin with a double cut HDR donor after CRISPR/Cas9-mediated double-stranded DNA cleavage. Genome Biol. 2017, 18, 35. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, X.; Liu, C.; Jiang, C.; Guo, X.; Xu, Q.; Yin, Z.; Liu, Z.; Mu, Y. Improving the Efficiency of CRISPR Ribonucleoprotein-Mediated Precise Gene Editing by Small Molecules in Porcine Fibroblasts. Animals 2024, 14, 719. [Google Scholar] [CrossRef]
- Maurissen, T.L.; Woltjen, K. Synergistic gene editing in human iPS cells via cell cycle and DNA repair modulation. Nat. Commun. 2020, 11, 2876. [Google Scholar] [CrossRef]
- Rui, Y.; Wilson, D.R.; Tzeng, S.Y.; Yamagata, H.M.; Sudhakar, D.; Conge, M.; Berlinicke, C.A.; Zack, D.J.; Tuesca, A.; Green, J.J. High-throughput and high-content bioassay enables tuning of polyester nanoparticles for cellular uptake, endosomal escape, and systemic in vivo delivery of mRNA. Sci. Adv. 2022, 8, eabk2855. [Google Scholar] [CrossRef]
- Luo, X.; Germer, J.; Burghardt, T.; Grau, M.; Lin, Y.; Höhn, M.; Lächelt, U.; Wagner, E. Dual pH-responsive CRISPR/Cas9 ribonucleoprotein xenopeptide complexes for genome editing. Eur. J. Pharm. Sci. 2025, 205, 106983. [Google Scholar] [CrossRef]
- Wittrup, A.; Ai, A.; Liu, X.; Hamar, P.; Trifonova, R.; Charisse, K.; Manoharan, M.; Kirchhausen, T.; Lieberman, J. Visualizing lipid-formulated siRNA release from endosomes and target gene knockdown. Nat. Biotechnol. 2015, 33, 870–876. [Google Scholar] [CrossRef] [PubMed]
- Grau, M.; Wagner, E. Strategies and mechanisms for endosomal escape of therapeutic nucleic acids. Curr. Opin. Chem. Biol. 2024, 81, 102506. [Google Scholar] [CrossRef]
- Heyer, W.-D.; Ehmsen, K.T.; Liu, J. Regulation of homologous recombination in eukaryotes. Annu. Rev. Genet. 2010, 44, 113–139. [Google Scholar] [CrossRef] [PubMed]
- Hustedt, N.; Durocher, D. The control of DNA repair by the cell cycle. Nat. Cell Biol. 2017, 19, 1–9. [Google Scholar] [CrossRef]
- Brunner, S.; Sauer, T.; Carotta, S.; Cotten, M.; Saltik, M.; Wagner, E. Cell cycle dependence of gene transfer by lipoplex, polyplex and recombinant adenovirus. Gene Ther. 2000, 7, 401–407. [Google Scholar] [CrossRef]
- Symens, N.; Soenen, S.J.; Rejman, J.; Braeckmans, K.; De Smedt, S.C.; Remaut, K. Intracellular partitioning of cell organelles and extraneous nanoparticles during mitosis. Adv. Drug Deliv. Rev. 2012, 64, 78–94. [Google Scholar] [CrossRef]
- Remaut, K.; Symens, N.; Lucas, B.; Demeester, J.; De Smedt, S. Cell division responsive peptides for optimized plasmid DNA delivery: The mitotic window of opportunity? J. Control. Release. 2014, 179, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Staahl, B.T.; Alla, R.K.; Doudna, J.A. Enhanced homology-directed human genome engineering by controlled timing of CRISPR/Cas9 delivery. eLife 2014, 3, e04766. [Google Scholar] [CrossRef]
- Lessl, A.-L.; Pöhmerer, J.; Lin, Y.; Wilk, U.; Höhn, M.; Hörterer, E.; Wagner, E.; Lächelt, U. mCherry on Top: A Positive Read-Out Cellular Platform for Screening DMD Exon Skipping Xenopeptide–PMO Conjugates. Bioconjug. Chem. 2023, 34, 2263–2274. [Google Scholar] [CrossRef]
- Germer, J.; Lessl, A.-L.; Pöhmerer, J.; Grau, M.; Weidinger, E.; Höhn, M.; Yazdi, M.; Cappelluti, M.A.; Lombardo, A.; Lächelt, U. Lipo-Xenopeptide Polyplexes for CRISPR/Cas9 based Gene editing at ultra-low dose. J. Control. Release. 2024, 370, 239–255. [Google Scholar] [CrossRef]
- Chen, S.; Carneiro, S.P.; Merkel, O.M. Anionic polymer coating for enhanced delivery of Cas9 mRNA and sgRNA nanoplexes. Biomater. Sci. 2025, 13, 659–676. [Google Scholar] [CrossRef] [PubMed]
- Aksoy, Y.A.; Nguyen, D.T.; Chow, S.; Chung, R.S.; Guillemin, G.J.; Cole, N.J.; Hesselson, D. Chemical reprogramming enhances homology-directed genome editing in zebrafish embryos. Commun. Biol. 2019, 2, 198. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.; Lin-Shiao, E.; Trinidad, M.; Saffari Doost, M.; Colognori, D.; Doudna, J.A. Decorating chromatin for enhanced genome editing using CRISPR-Cas9. Proc. Natl. Acad. Sci. USA 2022, 119, e2204259119. [Google Scholar] [CrossRef]
- Babačić, H.; Mehta, A.; Merkel, O.; Schoser, B. CRISPR-cas gene-editing as plausible treatment of neuromuscular and nucleotide-repeat-expansion diseases: A systematic review. PLoS ONE 2019, 14, e0212198. [Google Scholar] [CrossRef]
- Conant, D.; Hsiau, T.; Rossi, N.; Oki, J.; Maures, T.; Waite, K.; Yang, J.; Joshi, S.; Kelso, R.; Holden, K. Inference of CRISPR edits from Sanger trace data. CRISPR J. 2022, 5, 123–130. [Google Scholar] [CrossRef]
- Brinkman, E.K.; Chen, T.; Amendola, M.; Van Steensel, B. Easy quantitative assessment of genome editing by sequence trace decomposition. Nucleic Acids Res. 2014, 42, e168. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Foden, J.A.; Khayter, C.; Maeder, M.L.; Reyon, D.; Joung, J.K.; Sander, J.D. High-frequency off-target mutagenesis induced by CRISPR-Cas nucleases in human cells. Nat. Biotechnol. 2013, 31, 822–826. [Google Scholar] [CrossRef]
- Pacesa, M.; Lin, C.-H.; Cléry, A.; Saha, A.; Arantes, P.R.; Bargsten, K.; Irby, M.J.; Allain, F.H.-T.; Palermo, G.; Cameron, P. Structural basis for Cas9 off-target activity. Cell 2022, 185, 4067–4081.e21. [Google Scholar] [CrossRef]
- Naeem, M.; Majeed, S.; Hoque, M.Z.; Ahmad, I. Latest developed strategies to minimize the off-target effects in CRISPR-Cas-mediated genome editing. Cells 2020, 9, 1608. [Google Scholar] [CrossRef]
- NU, P.K.I. Preclinical Evaluation of a Potent Novel DNA-Dependent. Cancer Res 2006, 66, 5354–5362. [Google Scholar]
- Waldrip, Z.J.; Acharya, B.; Armstrong, D.; Hanafi, M.; Rainwater, R.R.; Amole, S.; Fulmer, M.; Azevedo-Pouly, A.C.; Burns, A.; Burdine, L. Discovery of the DNA-PKcs inhibitor DA-143 which exhibits enhanced solubility relative to NU7441. Sci. Rep. 2024, 14, 19999. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, Y. Development and evolution of DNA-dependent protein kinase inhibitors toward cancer therapy. Int. J. Mol. Sci. 2022, 23, 4264. [Google Scholar] [CrossRef] [PubMed]
- Tichy, A.; Durisova, K.; Salovska, B.; Pejchal, J.; Zarybnicka, L.; Vavrova, J.; Dye, N.A.; Sinkorova, Z. Radio-sensitization of human leukaemic MOLT-4 cells by DNA-dependent protein kinase inhibitor, NU7441. Radiat. Environ. Biophys. 2014, 53, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.Q.; Zheng, Z.; Nguyen, N.T.; Liebers, M.; Topkar, V.V.; Thapar, V.; Wyvekens, N.; Khayter, C.; Iafrate, A.J.; Le, L.P. GUIDE-seq enables genome-wide profiling of off-target cleavage by CRISPR-Cas nucleases. Nat. Biotechnol. 2015, 33, 187–197. [Google Scholar] [CrossRef]
- Wienert, B.; Wyman, S.K.; Richardson, C.D.; Yeh, C.D.; Akcakaya, P.; Porritt, M.J.; Morlock, M.; Vu, J.T.; Kazane, K.R.; Watry, H.L. Unbiased detection of CRISPR off-targets in vivo using DISCOVER-Seq. Science 2019, 364, 286–289. [Google Scholar] [CrossRef]
- Berger, S.; Krhač Levačić, A.; Hörterer, E.; Wilk, U.; Benli-Hoppe, T.; Wang, Y.; öztürk, O.; Luo, J.; Wagner, E. Optimizing pDNA lipo-polyplexes: A balancing act between stability and cargo release. Biomacromolecules 2021, 22, 1282–1296. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, X.; Weidinger, E.; Burghardt, T.; Höhn, M.; Wagner, E. CRISPR/Cas9 Ribonucleoprotein Delivery Enhanced by Lipo-Xenopeptide Carriers and Homology-Directed Repair Modulators: Insights from Reporter Cell Lines. Int. J. Mol. Sci. 2025, 26, 4361. https://doi.org/10.3390/ijms26094361
Luo X, Weidinger E, Burghardt T, Höhn M, Wagner E. CRISPR/Cas9 Ribonucleoprotein Delivery Enhanced by Lipo-Xenopeptide Carriers and Homology-Directed Repair Modulators: Insights from Reporter Cell Lines. International Journal of Molecular Sciences. 2025; 26(9):4361. https://doi.org/10.3390/ijms26094361
Chicago/Turabian StyleLuo, Xianjin, Eric Weidinger, Tobias Burghardt, Miriam Höhn, and Ernst Wagner. 2025. "CRISPR/Cas9 Ribonucleoprotein Delivery Enhanced by Lipo-Xenopeptide Carriers and Homology-Directed Repair Modulators: Insights from Reporter Cell Lines" International Journal of Molecular Sciences 26, no. 9: 4361. https://doi.org/10.3390/ijms26094361
APA StyleLuo, X., Weidinger, E., Burghardt, T., Höhn, M., & Wagner, E. (2025). CRISPR/Cas9 Ribonucleoprotein Delivery Enhanced by Lipo-Xenopeptide Carriers and Homology-Directed Repair Modulators: Insights from Reporter Cell Lines. International Journal of Molecular Sciences, 26(9), 4361. https://doi.org/10.3390/ijms26094361