
Molecular Dynamics Simulations of the SPRED2Leu100Pro EVH-1 Domain Complexed with the GAP-Related Domain of Neurofibromin
 , ,
, ,
Abstract
1. Introduction
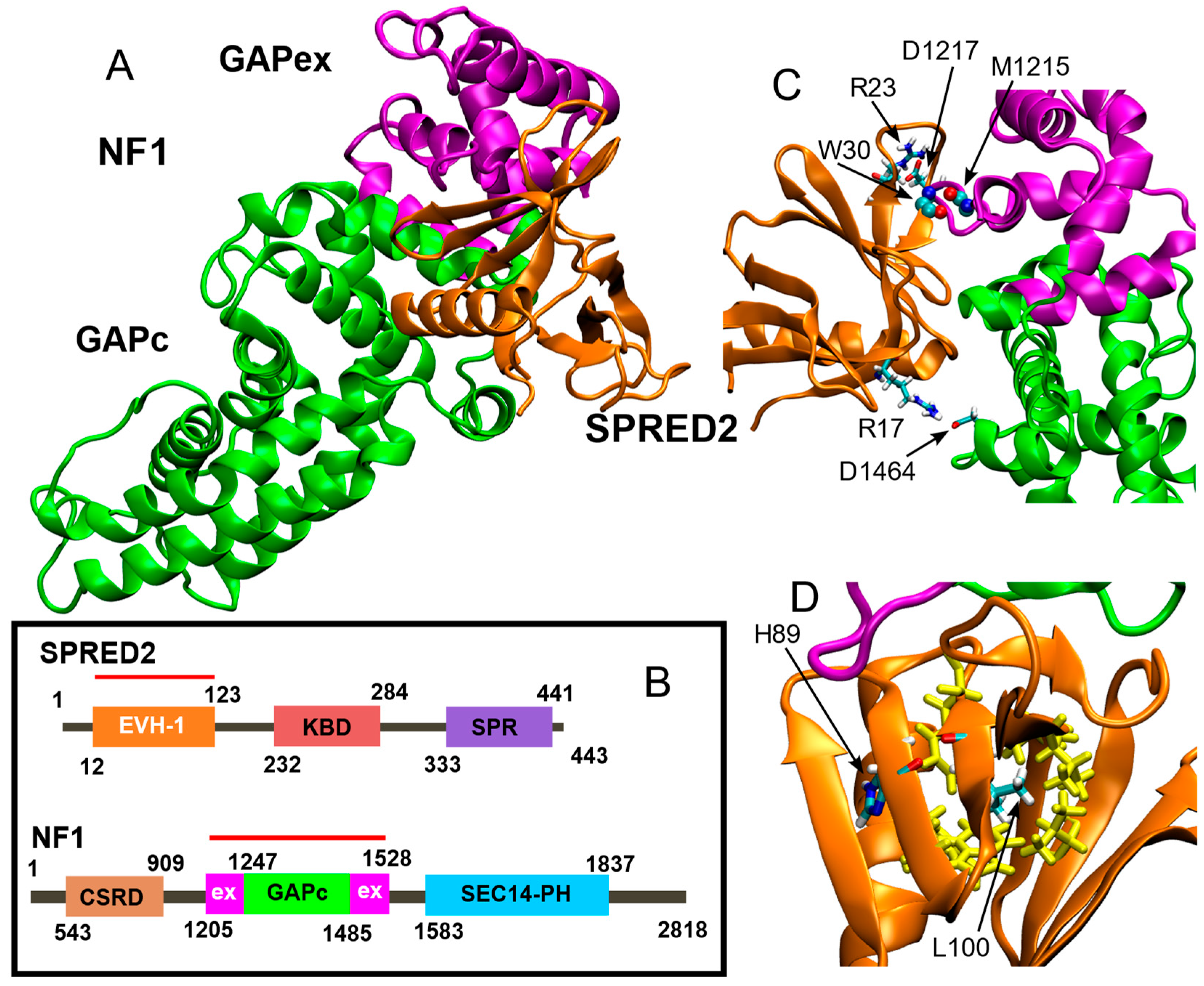
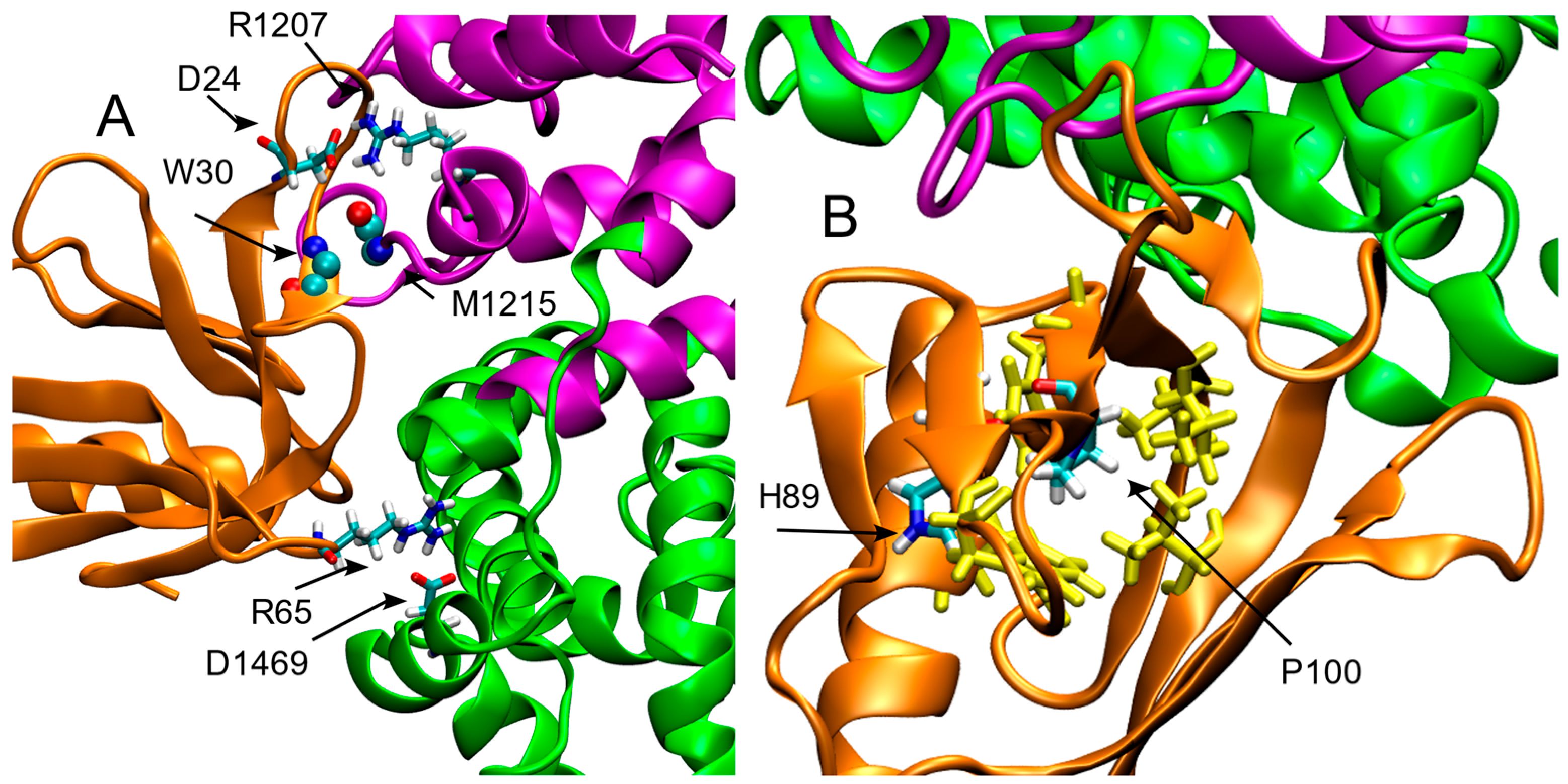
2. Results
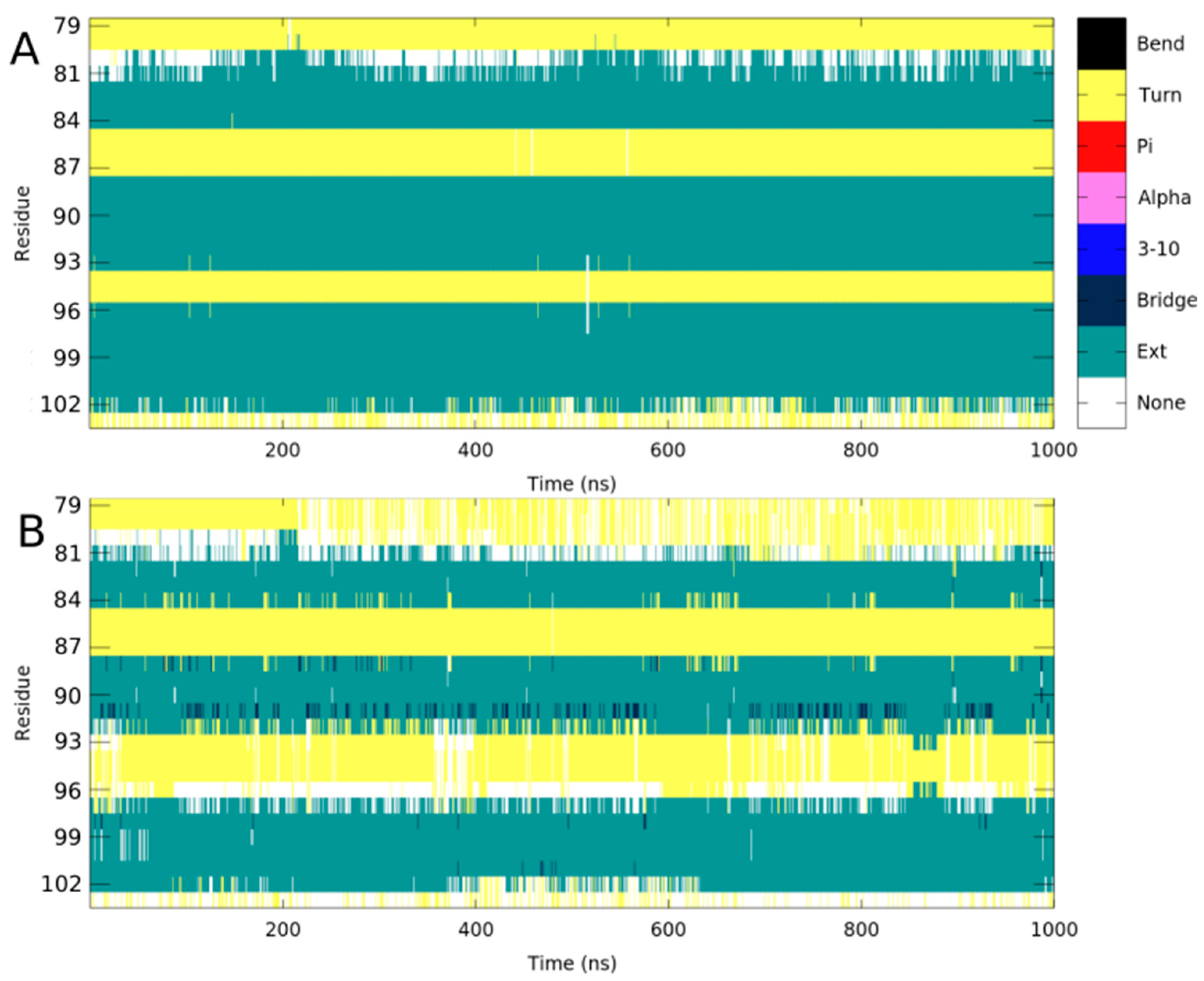
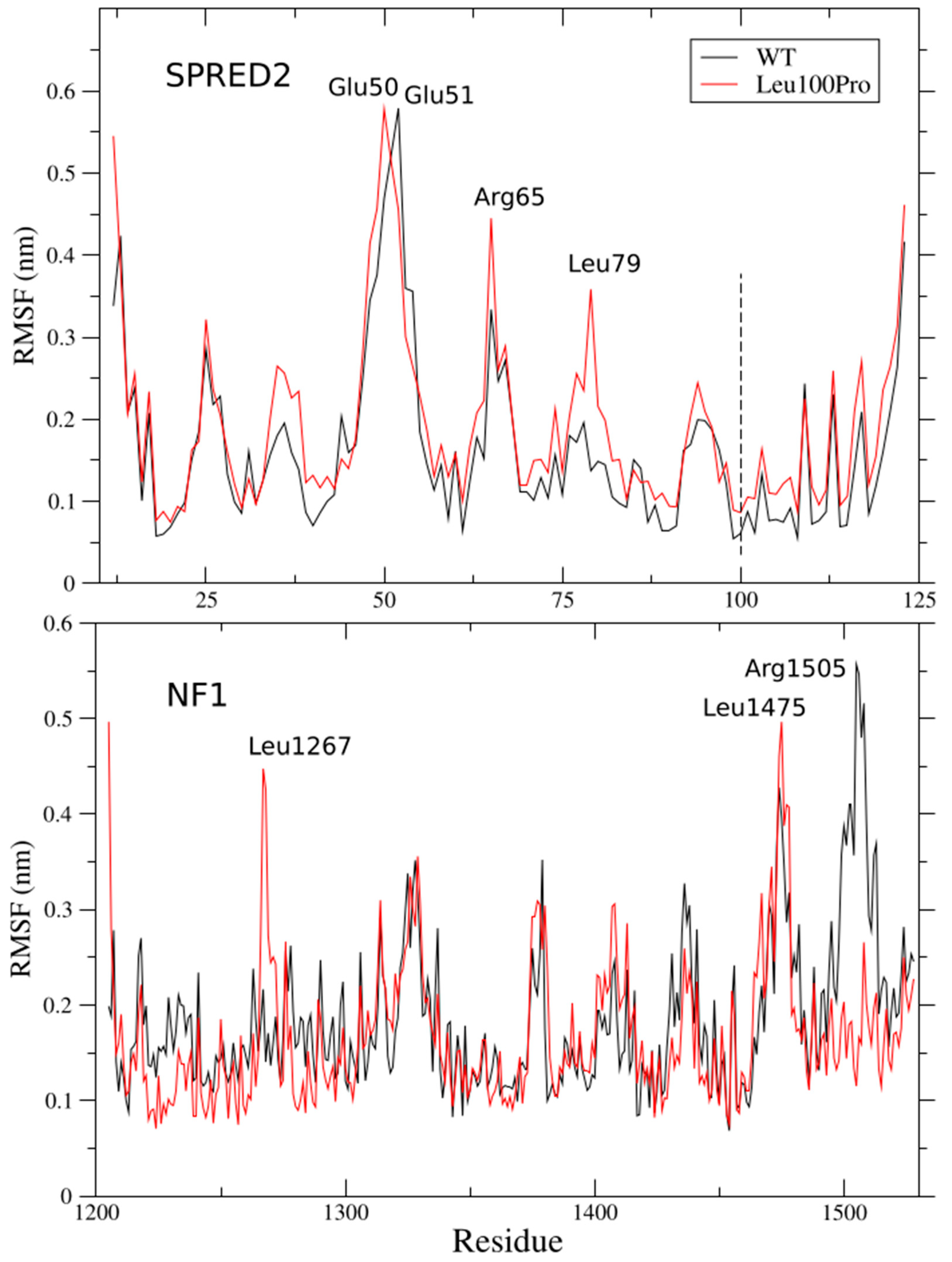
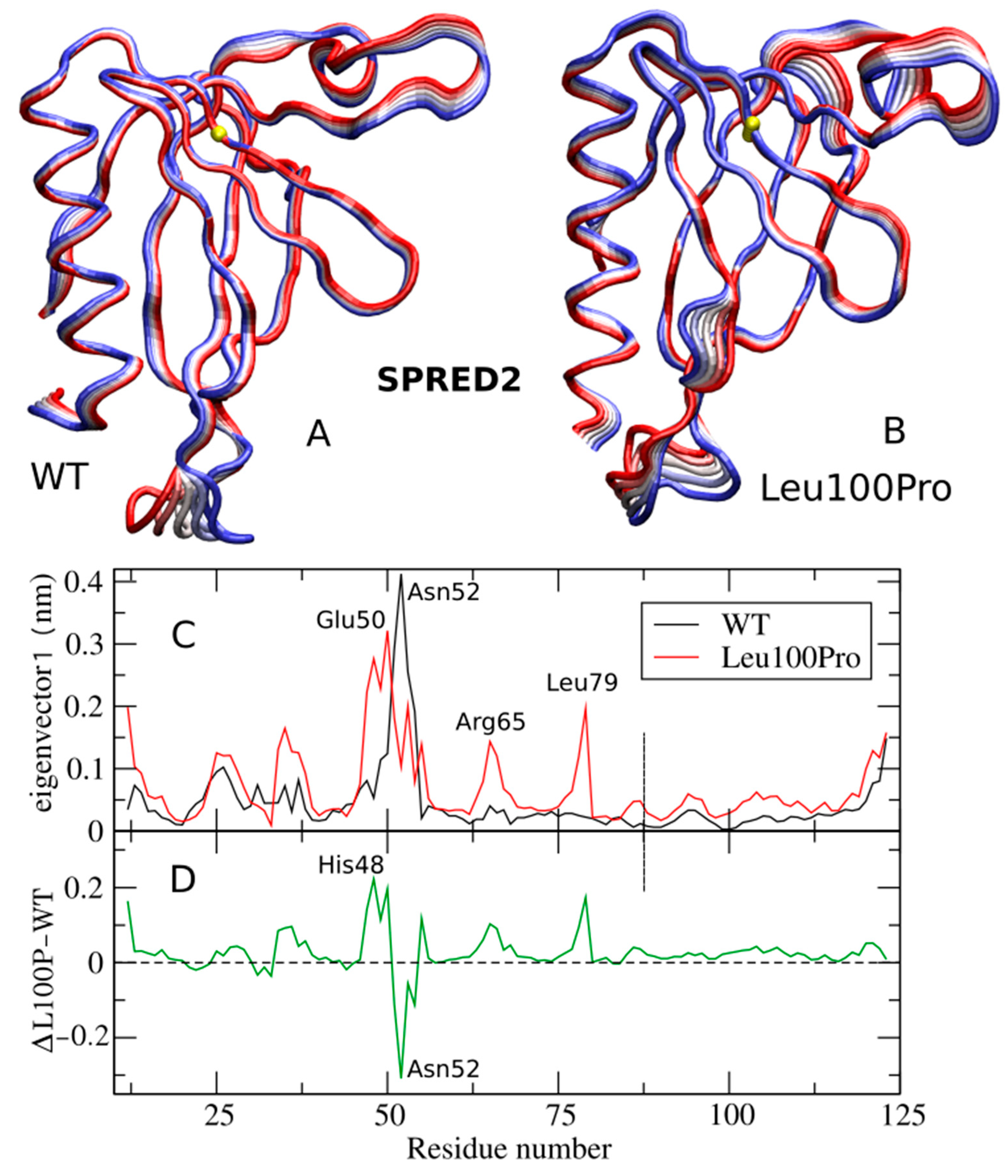
2.1. Per-Residue Root Mean Square Fluctuations
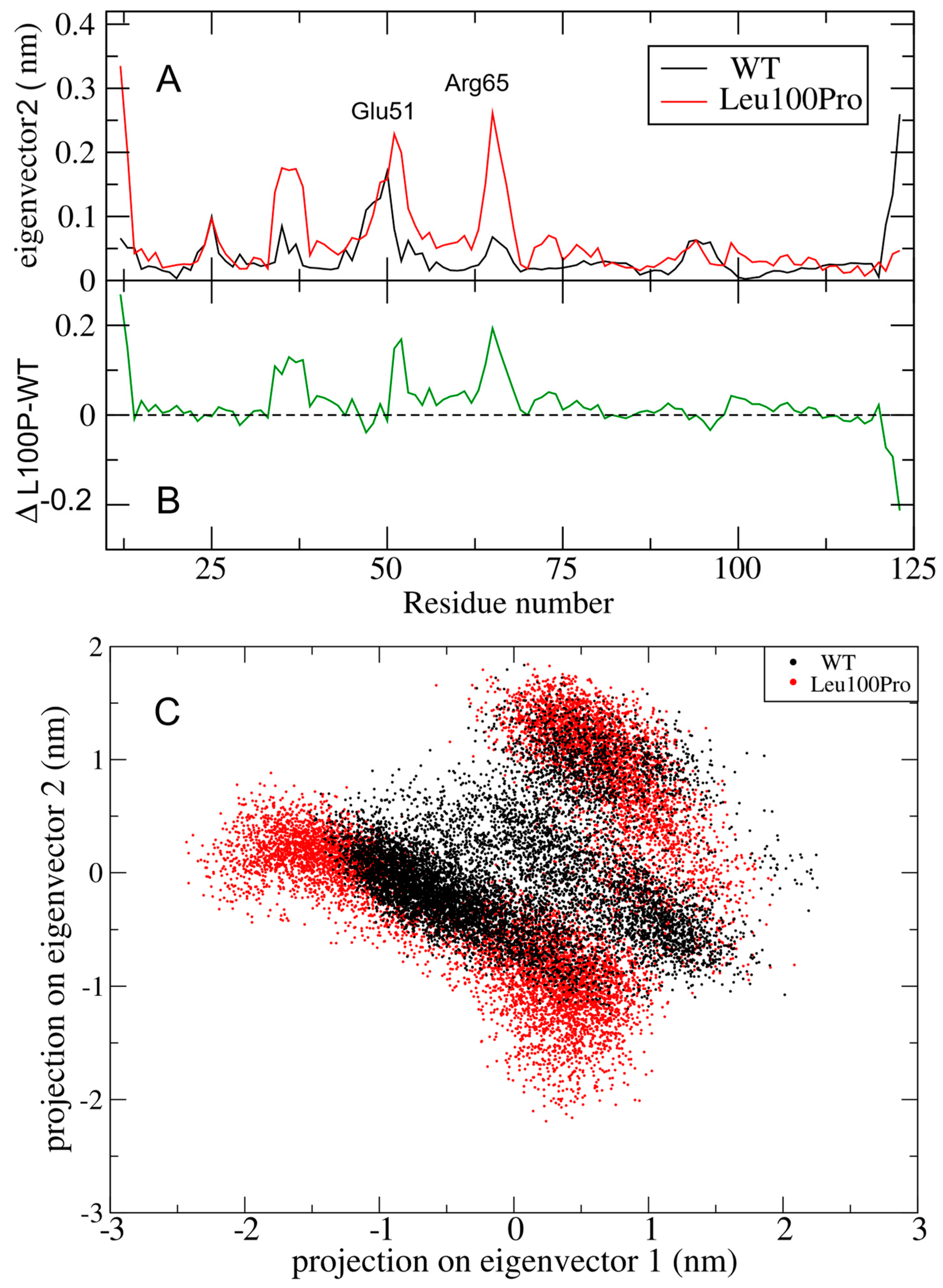
2.2. Large Collective Protein Movements
3. Discussion
4. Materials and Methods
4.1. Molecular Model
4.2. Molecular Dynamics Simulation of the Wild-Type System for Equilibration
4.3. Mutation Insertion and Trajectory Production
4.4. Molecular Dynamics Analyses
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SPRED | Sprouty-related EVH-1 domain-containing proteins |
| EVH-1 | Enabled/Vasodilator-stimulated phosphoprotein homology-1 |
| KBD | c-KIT Binding Domain |
| SPR | Sprouty-Related |
| LoF | Loss-of-Function |
| LS | Legius Syndrome |
| NS | Noonan Syndrome |
| MD | Molecular Dynamics |
| MAPK | Mitogen-Activated Protein Kinase |
| GAP | GTPase-Activating Protein |
| GAPex | GAP External region |
| RMSF | Root Mean Square Fluctuation |
| ED | Essential Dynamics |
| WT | Wild-type |
| Lincs | Linear Constraint Solver |
| PME | Particle Mesh Ewald |
References
- Gong, J.; Yan, Z.; Liu, Q. Progress in experimental research on SPRED protein family. J. Int. Med. Res. 2020, 48, 0300060520929170. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, C.; McCormick, F. SPRED proteins and their roles in signal transduction, development, and malignancy. Genes Dev. 2020, 34, 1410–1421. [Google Scholar] [CrossRef] [PubMed]
- Legius, E.; Stevenson, D. Legius syndrome. In GeneReviews; Adam, M.P., Feldman, J., Mirzaa, G.M., Pagon, R.A., Wallace, S.E., Amemiya, A., Eds.; Updated 2020; University of Washington: Seattle, WA, USA, 1993–2025. [Google Scholar]
- Brems, H.; Chmara, M.; Sahbatou, M.; Denayer, E.; Taniguchi, K.; Kato, R.; Somers, R.; Messiaen, L.; De Schepper, S.; Fryns, J.P.; et al. Germline loss-of-function mutations in SPRED1 cause a neurofibromatosis 1-like phenotype. Nat. Genet. 2007, 39, 1120–1126. [Google Scholar] [CrossRef]
- Friedman, J.M. Neurofibromatosis 1. In GeneReviews; Adam, M.P., Feldman, J., Mirzaa, G.M., Pagon, R.A., Wallace, S.E., Amemiya, A., Eds.; Updated 2025; University of Washington: Seattle, WA, USA, 1993–2025. [Google Scholar]
- Bergoug, M.; Doudeau, M.; Godin, F.; Mosrin, C.; Vallée, B.; Bénédetti, H. Neurofibromin Structure, Functions and Regulation. Cells. 2020, 9, 2365. [Google Scholar] [CrossRef]
- Motta, M.; Fasano, G.; Gredy, S.; Brinkmann, J.; Bonnard, A.A.; Simsek-Kiper, P.O.; Gulec, E.Y.; Essaddam, L.; Utine, G.E.; Guarnetti Prandi, I.; et al. SPRED2 loss-of-function causes a recessive Noonan syndrome-like phenotype. Am. J. Hum. Genet. 2021, 108, 2112–2129. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.E.; Allanson, J.E.; Tartaglia, M.; Gelb, B.D. Noonan syndrome. Lancet. 2013, 381, 333–342. [Google Scholar] [CrossRef]
- Tartaglia, M.; Aoki, Y.; Gelb, B.D. The molecular genetics of RASopathies: An update on novel disease genes and new disorders. Am. J. Med. Genet. Part C Semin. Med. Genet. 2022, 190, 425–439. [Google Scholar] [CrossRef]
- Yan, W.; Markegard, E.; Dharmaiah, S.; Urisman, A.; Drew, M.; Esposito, D.; Scheffzek, K.; Nissley, D.V.; McCormick, F.; Simanshu, D.K. Structural insights into the SPRED1-NF1-KRAS complex and disruption of SPRED1-NF1 interaction by oncogenic EGFR. Cell Rep. 2020, 32, 107909. [Google Scholar] [CrossRef] [PubMed]
- Lopez, J.; Bonsor, D.A.; Sale, M.J.; Urisman, A.; Mehalko, J.L.; Cabanski-Dunning, M.; Castel, P.; Simanshu, D.K.; McCormick, F. The ribosomal S6 kinase 2 (RSK2)–SPRED2 complex regulates the phosphorylation of RSK substrates and MAPK signaling. J. Biol. Chem. 2023, 299, 104789. [Google Scholar] [CrossRef]
- Brems, H.; Pasmant, E.; Van Minkelen, R.; Wimmer, K.; Upadhyaya, M.; Legius, E.; Messiaen, L. Review and update of SPRED1 mutations causing Legius syndrome. Hum. Mutat. 2012, 33, 1538–1546. [Google Scholar] [CrossRef]
- Brems, H.; Legius, E. Legius syndrome, an Update. Molecular pathology of mutations in SPRED1. Keio J. Med. 2013, 62, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Hollingsworth, S.A.; Dror, R.O. Molecular dynamics simulation for all. Neuron 2018, 99, 1129–1143. [Google Scholar] [CrossRef]
- Nuthakki, V.K.; Barik, R.; Gangashetty, S.B.; Srikanth, G. Advanced molecular modeling of proteins: Methods, breakthroughs, and future prospects. Adv. Pharmacol. 2025, 103, 23–41. [Google Scholar]
- Amadei, A.; Linssen, A.B.M.; Berendsen, H.J.C. Essential dynamics of proteins. Proteins Struct. Funct. Bioinform. 1993, 17, 412–425. [Google Scholar] [CrossRef] [PubMed]
- Moradi, S.; Nowroozi, A.; Aryaei Nezhad, M.; Jalali, P.; Khosravi, R.; Shahlaei, M. A review on description dynamics and conformational changes of proteins using combination of principal component analysis and molecular dynamics simulation. Comput. Biol. Med. 2024, 183, 109245. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Van Der Spoel, D.; Lindahl, E.; Hess, B.; Groenhof, G.; Mark, A.E.; Berendsen, H.J.C. GROMACS: Fast, flexible, and free. J. Comput. Chem. 2005, 26, 1701–1718. [Google Scholar] [CrossRef]
- Mark, P.; Nilsson, L. Structure and Dynamics of the TIP3P, SPC, and SPC/E Water Models at 298 K. J. Phys. Chem. A. 2001, 105, 9954–9960. [Google Scholar] [CrossRef]
- Huang, J.; MacKerell, A.D. CHARMM36 all-atom additive protein force field: Validation based on comparison to NMR data. J. Comput. Chem. 2013, 34, 2135–2145. [Google Scholar] [CrossRef] [PubMed]
- Hess, B.; Bekker, H.; Berendsen, H.J.C.; Fraaije, J.G.E.M. LINCS: A linear constraint solver for molecular simulations. J. Comput. Chem. 1997, 18, 1463–1472. [Google Scholar] [CrossRef]
- Castrignanò, T.; Gioiosa, S.; Flati, T.; Cestari, M.; Picardi, E.; Chiara, M.; Fratelli, M.; Amente, S.; Cirilli, M.; Tangaro, M.A.; et al. ELIXIR-IT HPC@CINECA: High performance computing resources for the bioinformatics community. BMC Bioinform. 2020, 21, 352. [Google Scholar] [CrossRef] [PubMed]
- Abraham, M.J.; Gready, J.E. Optimization of parameters for molecular dynamics simulation using smooth particle-mesh Ewald in GROMACS 4.5. J. Comput. Chem. 2011, 32, 2031–2040. [Google Scholar] [CrossRef]
- Bussi, G.; Donadio, D.; Parrinello, M. Canonical sampling through velocity rescaling. J. Chem. Phys. 2007, 126, 014101. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Donor | Acceptor | Occupancy | |
|---|---|---|---|
| WT | His89-Main-N | Leu100-Main-O | 44.0% |
| Leu100-Main-N | His89-Main-O | 43.4% | |
| L100P | His89-Main-N | Pro100-Main-O | 6.2% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Terrusa, M.; Sangiovanni, E.; Motta, M.; Tartaglia, M.; Prandi, I.G.; Chillemi, G. Molecular Dynamics Simulations of the SPRED2Leu100Pro EVH-1 Domain Complexed with the GAP-Related Domain of Neurofibromin. Int. J. Mol. Sci. 2025, 26, 4342. https://doi.org/10.3390/ijms26094342
Terrusa M, Sangiovanni E, Motta M, Tartaglia M, Prandi IG, Chillemi G. Molecular Dynamics Simulations of the SPRED2Leu100Pro EVH-1 Domain Complexed with the GAP-Related Domain of Neurofibromin. International Journal of Molecular Sciences. 2025; 26(9):4342. https://doi.org/10.3390/ijms26094342
Chicago/Turabian StyleTerrusa, Martina, Elisa Sangiovanni, Marialetizia Motta, Marco Tartaglia, Ingrid Guarnetti Prandi, and Giovanni Chillemi. 2025. "Molecular Dynamics Simulations of the SPRED2Leu100Pro EVH-1 Domain Complexed with the GAP-Related Domain of Neurofibromin" International Journal of Molecular Sciences 26, no. 9: 4342. https://doi.org/10.3390/ijms26094342
APA StyleTerrusa, M., Sangiovanni, E., Motta, M., Tartaglia, M., Prandi, I. G., & Chillemi, G. (2025). Molecular Dynamics Simulations of the SPRED2Leu100Pro EVH-1 Domain Complexed with the GAP-Related Domain of Neurofibromin. International Journal of Molecular Sciences, 26(9), 4342. https://doi.org/10.3390/ijms26094342