
From Synaptic Plasticity to Neurodegeneration: BDNF as a Transformative Target in Medicine

Abstract
1. Introduction
1.1. Unveiling BDNF: A Pillar of Neurobiology
1.2. The Dual Promise of BDNF: Biology and Clinical Relevance
1.3. Objectives of This Review
2. Molecular Structure and Mechanisms of BDNF
2.1. BDNF Gene Encoding and Precision Expression
2.2. The Multifaceted Protein: BDNF Isoforms and Structural Precision
2.3. Receptors and Pathway Dynamics: Translating Signals into Action
2.4. Dynamic Regulation of BDNF Signaling: Sustaining Neural Health
3. BDNF in Neurodevelopment
3.1. Orchestrating Neurogenesis and Neuronal Differentiation
3.2. Sculpting Neural Circuits: Axonal and Dendritic Growth
3.3. Activity-Dependent Plasticity and Synaptogenesis
3.4. Implications for Neurodevelopmental Disorders
4. BDNF and Synaptic Plasticity
4.1. Mechanisms of LTP and LTD
4.2. Role in Learning, Memory, and Cognitive Adaptability
4.3. Pre- and Postsynaptic Modulation in Synaptic Transmission
4.4. Therapeutic Applications and Innovations in Plasticity Modulation
4.4.1. Pharmacological Advances
4.4.2. Lifestyle Interventions
4.4.3. Technological Innovations
4.4.4. Key Innovations in the Pipeline
5. BDNF Polymorphisms and Cognitive Function
5.1. Val66Met Polymorphism: Molecular Disruptions and Functional Implications
5.2. Structural and Connectivity Alterations in Val66Met Carriers
5.3. Neuropsychiatric Vulnerabilities in Val66Met Carriers
5.4. Emerging Therapies and Precision Medicine for Val66Met Carriers
6. BDNF in Neurodegenerative Diseases
6.1. Alzheimer’s Disease: Synaptic Preservation and Mitochondrial Regulation
6.2. Parkinson’s Disease: Protecting Dopaminergic Neurons and Axonal Integrity
6.3. Huntington’s Disease: Counteracting mHTT Toxicity and Synaptic Dysfunction
6.4. ALS, MS, and Expanding the Therapeutic Scope of BDNF
7. BDNF as a Biomarker
7.1. Peripheral and Central BDNF Levels: Mechanisms and Implications
7.2. Diagnostic Value of BDNF in Early Disease Detection
7.3. Monitoring Disease Progression and Evaluating Therapeutic Efficacy
7.4. Advances in BDNF Detection Technologies
8. BDNF and Physical Activity
8.1. Molecular Mechanisms: Exercise-Induced Regulation of BDNF Expression
8.2. Cognitive Benefits of Exercise-Induced BDNF
8.3. Neuroprotection and Recovery in Neurological Disorders
8.4. Tailoring Exercise Protocols for Maximum BDNF Modulation
9. BDNF and Diet
9.1. Molecular Mechanisms of Nutrient-Induced BDNF Regulation
9.2. Cognitive and Neuroprotective Benefits of BDNF-Boosting Diets
10. BDNF and Stress
10.1. Stress Suppression of BDNF: Mechanisms and Consequences
10.2. BDNF’s Role in Resilience to Stress
10.3. Stress-Related Psychiatric Disorders: BDNF Dysregulation as a Mechanistic Link
10.4. Interventions to Restore BDNF and Mitigate Stress Effects
11. BDNF and Pharmacological Interventions
11.1. TrkB Agonists: Directly Enhancing Neurotrophic Signaling
11.2. BDNF Modulation by Antidepressants: Broadening Mechanistic Understanding
11.3. Neurodegenerative Applications: Addressing Synaptic and Cellular Loss
11.4. Emerging Therapies: mRNA Delivery and Gene Editing
12. BDNF as a Therapeutic Target for Precision Medicine
13. Challenges and Future Directions in BDNF Research and Therapeutics
14. Conclusions and Future Directions—The Roadmap for BDNF-Centered Therapies
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| AAV | Adeno-Associated Virus |
| AD | Alzheimer’s Disease |
| Akt | Protein Kinase B |
| ALS | Amyotrophic Lateral Sclerosis |
| AMPK | AMP-Activated Protein Kinase |
| BBB | Blood–Brain Barrier |
| BDNF | Brain-Derived Neurotrophic Factor |
| CNS | Central Nervous System |
| CREB | cAMP Response Element-Binding Protein |
| CRISPR | Clustered Regularly Interspaced Short Palindromic Repeats |
| CSF | Cerebrospinal Fluid |
| DHA | Docosahexaenoic Acid |
| ERK | Extracellular Signal-Regulated Kinase |
| GABA | Gamma-Aminobutyric Acid |
| GPx | Glutathione Peroxidase |
| HD | Huntington’s Disease |
| HDAC | Histone Deacetylase |
| Hsp70 | Heat Shock Protein 70 |
| IL-6 | Interleukin-6 |
| LTP | Long-Term Potentiation |
| MAPK | Mitogen-Activated Protein Kinase |
| MDD | Major Depressive Disorder |
| MEK | MAPK/ERK Kinase |
| mRNA | Messenger Ribonucleic Acid |
| mTOR | Mammalian Target of Rapamycin |
| Nrf2 | Nuclear Factor Erythroid 2-Related Factor 2 |
| PD | Parkinson’s Disease |
| PI3K | Phosphatidylinositol 3-Kinase |
| PLC-γ | Phospholipase C Gamma |
| p-tau181 | Phosphorylated Tau 181 |
| PTEN | Phosphatase and Tensin Homolog |
| PTSD | Post-Traumatic Stress Disorder |
| ROS | Reactive Oxygen Species |
| SIRT1 | Sirtuin 1 |
| SCFA | Short-Chain Fatty Acid |
| SHP2 | Src Homology 2 Domain-Containing Protein Tyrosine Phosphatase 2 |
| SOD | Superoxide Dismutase |
| TNF-α | Tumor Necrosis Factor Alpha |
| TrkB | Tropomyosin Receptor Kinase B |
| UPDRS | Unified Parkinson’s Disease Rating Scale |
| Val66Met | BDNF Gene Polymorphism |
References
- You, S.; Ma, Z.; Zhang, P.; Xu, W.; Zhan, C.; Sang, N.; Xu, J.; Wang, F.; Zhang, J. Neuroprotective effects of the salidroside derivative SHPL-49 via the BDNF/TrkB/Gap43 pathway in rats with cerebral ischemia. Biomed. Pharmacother. 2024, 174, 116460. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Xu, J.; Cui, Q.; Lin, G.; Wang, F.; Ding, X.; You, S.; Sang, N.; Tan, J.; Xu, W.; et al. Multi-pathway neuroprotective effects of a novel salidroside derivative SHPL-49 against acute cerebral ischemic injury. Eur. J. Pharmacol. 2023, 949, 175716. [Google Scholar] [CrossRef] [PubMed]
- Ávila-Gámiz, F.; Pérez-Cano, A.M.; Pérez-Berlanga, J.M.; Mullor-Vigo, R.M.; Zambrana-Infantes, E.N.; Santín, L.J.; Ladrón de Guevara-Miranda, D. Sequential treadmill exercise and cognitive training synergistically increase adult hippocampal neurogenesis in mice. Physiol. Behav. 2023, 266, 114184. [Google Scholar] [CrossRef]
- Yan, P.; Xue, Z.; Li, D.; Ni, S.; Wang, C.; Jin, X.; Zhou, D.; Li, X.; Zhao, X.; Chen, X.; et al. Dysregulated CRTC1-BDNF signaling pathway in the hippocampus contributes to Aβ oligomer-induced long-term synaptic plasticity and memory impairment. Exp. Neurol. 2021, 345, 113812. [Google Scholar] [CrossRef] [PubMed]
- Abellaneda-Pérez, K.; Martin-Trias, P.; Cassé-Perrot, C.; Vaqué-Alcázar, L.; Lanteaume, L.; Solana, E.; Babiloni, C.; Lizio, R.; Junqué, C.; Bargalló, N.; et al. BDNF Val66Met gene polymorphism modulates brain activity following rTMS-induced memory impairment. Sci. Rep. 2022, 12, 176. [Google Scholar] [CrossRef]
- Soler, C.T.; Kanders, S.H.; Olofsdotter, S.; Vadlin, S.; Åslund, C.; Nilsson, K.W. Exploration of the Moderating Effects of Physical Activity and Early Life Stress on the Relation between Brain-Derived Neurotrophic Factor (BDNF) rs6265 Variants and Depressive Symptoms among Adolescents. Genes 2022, 13, 1236. [Google Scholar] [CrossRef]
- Białecka, M.; Kurzawski, M.; Roszmann, A.; Robowski, P.; Sitek, E.J.; Honczarenko, K.; Mak, M.; Deptuła-Jarosz, M.; Gołąb-Janowska, M.; Droździk, M.; et al. BDNF G196A (Val66Met) polymorphism associated with cognitive impairment in Parkinson’s disease. Neurosci. Lett. 2014, 561, 86–90. [Google Scholar] [CrossRef]
- Hayat, M.R.; Umair, M.; Ikhtiar, H.; Wazir, S.; Palwasha, A.; Shah, M. The Relationship Between Brain-Derived Neurotrophic Factor and Serotonin in Major Depressive and Bipolar Disorders: A Cross-Sectional Analysis. Cureus 2024, 16, e70728. [Google Scholar] [CrossRef]
- Sakhawat, A.; Khan, M.U.; Rehman, R.; Khan, S.; Shan, M.A.; Batool, A.; Javed, M.A.; Ali, Q. Natural compound targeting BDNF V66M variant: Insights from in silico docking and molecular analysis. AMB Express 2023, 13, 134. [Google Scholar] [CrossRef]
- Colliva, A.; Tongiorgi, E. Distinct role of 5′UTR sequences in dendritic trafficking of BDNF mRNA: Additional mechanisms for the BDNF splice variants spatial code. Mol. Brain 2021, 14, 10. [Google Scholar] [CrossRef]
- Esvald, E.-E.; Tuvikene, J.; Kiir, C.S.; Avarlaid, A.; Tamberg, L.; Sirp, A.; Shubina, A.; Cabrera-Cabrera, F.; Pihlak, A.; Koppel, I.; et al. Revisiting the expression of BDNF and its receptors in mammalian development. Front. Mol. Neurosci. 2023, 16, 1182499. [Google Scholar] [CrossRef] [PubMed]
- Veltri, A.; Nicolì, V.; Marino, R.; Rea, F.; Corsi, M.; Chiumiento, M.; Giangreco, M.; Caldi, F.; Guglielmi, G.; Foddis, R.; et al. Plasma Brain-Derived Neurotrophic Factor (BDNF) Levels and BDNF Promoters’ DNA Methylation in Workers Exposed to Occupational Stress and Suffering from Psychiatric Disorders. Brain Sci. 2024, 14, 1106. [Google Scholar] [CrossRef]
- Oga, K.; Fuchikami, M.; Kobayashi, H.; Miyagi, T.; Fujita, S.; Fujita, S.; Okada, S.; Morinobu, S. Involvement of dysregulated hippocampal histone H3K9 methylation at the promoter of the BDNF gene in impaired memory extinction. Psychopharmacology 2024, 241, 2363–2374. [Google Scholar] [CrossRef]
- Lekk, I.; Cabrera-Cabrera, F.; Turconi, G.; Tuvikene, J.; Esvald, E.-E.; Rähni, A.; Casserly, L.; Garton, D.R.; Andressoo, J.-O.; Timmusk, T.; et al. Untranslated regions of brain-derived neurotrophic factor mRNA control its translatability and subcellular localization. J. Biol. Chem. 2023, 299, 102897. [Google Scholar] [CrossRef]
- Danelon, V.; Garret-Thomson, S.C.; Almo, S.C.; Lee, F.S.; Hempstead, B.L. Immune activation of the p75 neurotrophin receptor: Implications in neuroinflammation. Front. Mol. Neurosci. 2023, 16, 1305574. [Google Scholar] [CrossRef] [PubMed]
- Ilchibaeva, T.; Tsybko, A.; Lipnitskaya, M.; Eremin, D.; Milutinovich, K.; Naumenko, V.; Popova, N. Brain-Derived Neurotrophic Factor (BDNF) in Mechanisms of Autistic-like Behavior in BTBR Mice: Crosstalk with the Dopaminergic Brain System. Biomedicines 2023, 11, 1482. [Google Scholar] [CrossRef]
- Santos, M.; Lima, L.; Carvalho, S.; Mota-Pereira, J.; Pimentel, P.; Maia, D.; Correia, D.; Barroso, M.F.; Gomes, S.; Cruz, A.; et al. The Impact of BDNF, NTRK2, NGFR, CREB1, GSK3B, AKT, MAPK1, MTOR, PTEN, ARC, and SYN1 Genetic Polymorphisms in Antidepressant Treatment Response Phenotypes. Int. J. Mol. Sci. 2023, 24, 6758. [Google Scholar] [CrossRef]
- Moya-Alvarado, G.; Guerra, M.V.; Tiburcio, R.; Bravo, E.; Bronfman, F.C. The Rab11-regulated endocytic pathway and BDNF/TrkB signaling: Roles in plasticity changes and neurodegenerative diseases. Neurobiol. Dis. 2022, 171, 105796. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.K.S.; Coughlan, C.; Heyn, P.C.; Tagawa, A.; Carollo, J.J.; Kua, E.H.; Mahendran, R. Increased plasma brain-derived neurotrophic factor (BDNF) as a potential biomarker for and compensatory mechanism in mild cognitive impairment: A case-control study. Aging 2021, 13, 22666–22689. [Google Scholar] [CrossRef]
- Bouvet, P.; de Gea, P.; Aimard, M.; Chounlamountri, N.; Honnorat, J.; Delcros, J.G.; Salin, P.A.; Meissirel, C. A novel peptide derived from vascular endothelial growth factor prevents amyloid beta aggregation and toxicity. Aging Cell 2023, 22, e13907. [Google Scholar] [CrossRef]
- Ou, Z.-Y.A.; Byrne, L.M.; Rodrigues, F.B.; Tortelli, R.; Johnson, E.B.; Foiani, M.S.; Arridge, M.; De Vita, E.; Scahill, R.I.; Heslegrave, A.; et al. Brain-derived neurotrophic factor in cerebrospinal fluid and plasma is not a biomarker for Huntington’s disease. Sci. Rep. 2021, 11, 3481. [Google Scholar] [CrossRef] [PubMed]
- Want, A.; Nan, X.; Kokkali, E.; Barde, Y.-A.; Morgan, J.E. Brain-derived neurotrophic factor released from blood platelets prevents dendritic atrophy of lesioned adult central nervous system neurons. Brain Commun. 2023, 5, fcad046. [Google Scholar] [CrossRef]
- Varghese, N.; Buergin, D.; Boonmann, C.; Stadler, C.; Schmid, M.; Eckert, A.; Unternaehrer, E. Interplay between stress, sleep, and BDNF in a high-risk sample of young adults. Sci. Rep. 2023, 13, 20524. [Google Scholar] [CrossRef]
- Ge, Y.; Cao, Y.; Zhang, J.; Li, F.; Wang, J.; Sun, M.; Liu, Y.; Long, X.; Guo, W.; Liu, J.; et al. GOS enhances BDNF-mediated mammary gland development in pubertal mice via the gut-brain axis. npj Biofilms Microbiomes 2024, 10, 130. [Google Scholar] [CrossRef]
- Hang, P.-Z.; Ge, F.-Q.; Zhang, M.-R.; Li, Q.-H.; Yu, H.-Q.; Song, Y.-C.; Guo, D.-D.; Zhao, J.; Zhu, H. BDNF mimetic 7,8-dihydroxyflavone rescues rotenone-induced cytotoxicity in cardiomyocytes by ameliorating mitochondrial dysfunction. Free. Radic. Biol. Med. 2023, 198, 83–91. [Google Scholar] [CrossRef]
- Ávila-Gómez, P.; Shingai, Y.; Dash, S.; Liu, C.; Callegari, K.; Meyer, H.; Khodarkovskaya, A.; Aburakawa, D.; Uchida, H.; Faraco, G.; et al. Molecular and Functional Alterations in the Cerebral Microvasculature in an Optimized Mouse Model of Sepsis-Associated Cognitive Dysfunction. eNeuro 2024, 11, ENEURO.0426-23.2024. [Google Scholar] [CrossRef] [PubMed]
- Piancatelli, D.; Aureli, A.; Sebastiani, P.; Colanardi, A.; Del Beato, T.; Del Cane, L.; Sucapane, P.; Marini, C.; Di Loreto, S. Gene- and Gender-Related Decrease in Serum BDNF Levels in Alzheimer’s Disease. Int. J. Mol. Sci. 2022, 23, 14599. [Google Scholar] [CrossRef]
- Riolo, G.; Ricci, C.; De Angelis, N.; Marzocchi, C.; Guerrera, G.; Borsellino, G.; Giannini, F.; Battistini, S. BDNF and Pro-BDNF in Amyotrophic Lateral Sclerosis: A New Perspective for Biomarkers of Neurodegeneration. Brain Sci. 2022, 12, 617. [Google Scholar] [CrossRef] [PubMed]
- Gaur, A.; Varatharajan, S.; Balan, Y.; Taranikanti, M.; John, N.A.; Umesh, M.; Ganji, V.; Medala, K. Brain-derived neurotrophic factor (BDNF) and other neurotrophic factors in type 2 diabetes mellitus and their association with neuropathy. Ir. J. Med. Sci. 2024, 193, 2287–2292. [Google Scholar] [CrossRef]
- El-Khatib, S.M.; Vagadia, A.R.; Le, A.C.D.; Baulch, J.E.; Ng, D.Q.; Du, M.; Johnston, K.G.; Tan, Z.; Xu, X.; Chan, A.; et al. BDNF augmentation reverses cranial radiation therapy-induced cognitive decline and neurodegenerative consequences. Acta Neuropathol. Commun. 2024, 12, 190. [Google Scholar] [CrossRef]
- Li, G.; Wu, Q.; Wang, C.; Deng, P.; Li, J.; Zhai, Z.; Li, Y. Curcumin reverses cognitive deficits through promoting neurogenesis and synapse plasticity via the upregulation of PSD95 and BDNF in mice. Sci. Rep. 2025, 15, 1135. [Google Scholar] [CrossRef] [PubMed]
- Tong, T.; Chen, Y.; Hao, C.; Shen, J.; Chen, W.; Cheng, W.; Yan, S.; Li, J.; Li, Y.; Gulizhaerkezi, T.; et al. The effects of acupuncture on depression by regulating BDNF-related balance via lateral habenular nucleus BDNF/TrkB/CREB signaling pathway in rats. Behav. Brain Res. 2023, 451, 114509. [Google Scholar] [CrossRef]
- Mercado, N.M.; Szarowicz, C.; Stancati, J.A.; Sortwell, C.E.; Boezwinkle, S.A.; Collier, T.J.; Caulfield, M.E.; Steece-Collier, K. Advancing age and the rs6265 BDNF SNP are permissive to graft-induced dyskinesias in parkinsonian rats. npj Park. Dis. 2024, 10, 163. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Chu, S.; Wang, Z.; Li, F.; Yuan, Y.; Chen, N. Ginsenoside Rg1 exerts neuroprotective effects in 3-nitropronpionic acid-induced mouse model of Huntington’s disease via suppressing MAPKs and NF-κB pathways in the striatum. Acta Pharmacol. Sin. 2021, 42, 1409–1421. [Google Scholar] [CrossRef] [PubMed]
- Calvo-Enrique, L.; Lisa, S.; Vicente-García, C.; Deogracias, R.; Arévalo, J.C. Enhanced TrkA signaling impairs basal forebrain-dependent behavior. Front. Mol. Neurosci. 2023, 16, 1266983. [Google Scholar] [CrossRef]
- Fabbretti, E.; Antognolli, G.; Tongiorgi, E. Amyloid-β Impairs Dendritic Trafficking of Golgi-Like Organelles in the Early Phase Preceding Neurite Atrophy: Rescue by Mirtazapine. Front. Mol. Neurosci. 2021, 14, 661728. [Google Scholar] [CrossRef]
- Moya-Alvarado, G.; Tiburcio-Felix, R.; Ibáñez, M.R.; Aguirre-Soto, A.A.; Guerra, M.V.; Wu, C.; Mobley, W.C.; Perlson, E.; Bronfman, F.C. BDNF/TrkB signaling endosomes in axons coordinate CREB/mTOR activation and protein synthesis in the cell body to induce dendritic growth in cortical neurons. eLife 2023, 12, e77455. [Google Scholar] [CrossRef]
- Sumbul-Sekerci, B.; Sekerci, A.; Pasin, O.; Durmus, E.; Yuksel-Salduz, Z.I. Cognition and BDNF levels in prediabetes and diabetes: A mediation analysis of a cross-sectional study. Front. Endocrinol. 2023, 14, 1120127. [Google Scholar] [CrossRef]
- Dunne, V.L.; Ghita-Pettigrew, M.; Redmond, K.M.; Small, D.M.; Weldon, S.; Taggart, C.C.; Prise, K.M.; Hanna, G.G.; Butterworth, K.T. PTEN Depletion Increases Radiosensitivity in Response to Ataxia Telangiectasia-Related-3 (ATR) Inhibition in Non-Small Cell Lung Cancer (NSCLC). Int. J. Mol. Sci. 2024, 25, 7817. [Google Scholar] [CrossRef]
- Li, T.; Li, X.; Huang, X.; Yu, H.; Li, S.; Zhang, Z.; Xie, Y.; Song, X.; Liu, J.; Yang, X.; et al. Mitochondriomics reveals the underlying neuroprotective mechanism of TrkB receptor agonist R13 in the 5×FAD mice. Neuropharmacology 2022, 204, 108899. [Google Scholar] [CrossRef]
- Muscat, S.M.; Deems, N.P.; Butler, M.J.; Scaria, E.A.; Bettes, M.N.; Cleary, S.P.; Bockbrader, R.H.; Maier, S.F.; Barrientos, R.M. Selective TLR4 Antagonism Prevents and Reverses Morphine-Induced Persistent Postoperative Cognitive Dysfunction, Dysregulation of Synaptic Elements, and Impaired BDNF Signaling in Aged Male Rats. J. Neurosci. 2023, 43, 155–172. [Google Scholar] [CrossRef]
- Tan, L.L.; Alfonso, J.; Monyer, H.; Kuner, R. Neurogenesis in the adult brain functionally contributes to the maintenance of chronic neuropathic pain. Sci. Rep. 2021, 11, 18549. [Google Scholar] [CrossRef]
- Lin, T.-C.; Tsai, Y.-C.; Chen, Y.-A.; Young, T.-H.; Wu, C.-C.; Chiang, Y.-H.; Kao, C.-H.; Huang, A.P.-H.; Hsu, Y.-H.; Chen, K.-Y.; et al. Brain-derived neurotrophic factor contributes to neurogenesis after intracerebral hemorrhage: A rodent model and human study. Front. Cell. Neurosci. 2023, 17, 1170251. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-W.; Chen, S.-J.; Tsai, K.-J.; Hsu, K.-S.; Chen, Y.-F.; Chang, C.-H.; Lin, H.-H.; Hsueh, W.-Y.; Hsieh, H.-P.; Lee, Y.-F.; et al. Targeting cathepsin S promotes activation of OLF1-BDNF/TrkB axis to enhance cognitive function. J. Biomed. Sci. 2024, 31, 46. [Google Scholar] [CrossRef] [PubMed]
- Sochal, M.; Ditmer, M.; Binienda, A.; Tarasiuk, A.; Białasiewicz, P.; Turkiewicz, S.; Karuga, F.F.; Jakub, F.; Gabryelska, A. Interactions between neurotrophins, mood, and physical activity under the conditions of sleep deprivation. Transl. Psychiatry 2024, 14, 158. [Google Scholar] [CrossRef]
- Thapak, P.; Smith, G.; Ying, Z.; Paydar, A.; Harris, N.; Gomez-Pinilla, F. The BDNF mimetic R-13 attenuates TBI pathogenesis using TrkB-related pathways and bioenergetics. Biochim. Biophys. Acta Mol. Basis Dis. 2023, 1869, 166781. [Google Scholar] [CrossRef]
- Wu, W.; Yao, H.; Negraes, P.D.; Wang, J.; Trujillo, C.A.; de Souza, J.S.; Muotri, A.R.; Haddad, G.G. Neuronal hyperexcitability and ion channel dysfunction in CDKL5-deficiency patient iPSC-derived cortical organoids. Neurobiol. Dis. 2022, 174, 105882. [Google Scholar] [CrossRef]
- Swain, M.; Soman, S.K.; Tapia, K.; Dagda, R.Y.; Dagda, R.K. Brain-derived neurotrophic factor protects neurons by stimulating mitochondrial function through protein kinase A. J. Neurochem. 2023, 167, 104–125. [Google Scholar] [CrossRef]
- Bertacchi, M.; Maharaux, G.; Loubat, A.; Jung, M.; Studer, M. FGF8-mediated gene regulation affects regional identity in human cerebral organoids. eLife 2024, 13, e98096. [Google Scholar] [CrossRef]
- Alnakhli, A.M.; Saleh, A.; Kabel, A.M.; Estfanous, R.S.; Borg, H.M.; Alsufyani, K.M.; Sabry, N.M.; Gomaa, F.A.M.; Abd Elmaaboud, M.A. Perindopril Ameliorates Sodium Valproate-Induced Rat Model of Autism: Involvement of Sirtuin-1, JAK2/STAT3 Axis, PI3K/Akt/GSK-3β Pathway, and PPAR-Gamma Signaling. Medicina 2024, 60, 1802. [Google Scholar] [CrossRef]
- Niu, W.; Deng, L.; Mojica-Perez, S.P.; Tidball, A.M.; Sudyk, R.; Stokes, K.; Parent, J.M. Abnormal cell sorting and altered early neurogenesis in a human cortical organoid model of Protocadherin-19 clustering epilepsy. Front. Cell. Neurosci. 2024, 18, 1339345. [Google Scholar] [CrossRef] [PubMed]
- Camussi, D.; Naef, V.; Brogi, L.; Della Vecchia, S.; Marchese, M.; Nicoletti, F.; Santorelli, F.M.; Licitra, R. Delving into the Complexity of Valproate-Induced Autism Spectrum Disorder: The Use of Zebrafish Models. Cells 2024, 13, 1349. [Google Scholar] [CrossRef] [PubMed]
- Chandía-Cristi, A.; Stuardo, N.; Trejos, C.; Leal, N.; Urrutia, D.; Bronfman, F.C.; Álvarez Rojas, A. c-Abl Tyrosine Kinase Is Required for BDNF-Induced Dendritic Branching and Growth. Int. J. Mol. Sci. 2023, 24, 1944. [Google Scholar] [CrossRef]
- Raudzus, F.; Schöneborn, H.; Neumann, S.; Secret, E.; Michel, A.; Fresnais, J.; Brylski, O.; Ménager, C.; Siaugue, J.-M.; Heumann, R. Magnetic spatiotemporal control of SOS1 coupled nanoparticles for guided neurite growth in dopaminergic single cells. Sci. Rep. 2020, 10, 22452. [Google Scholar] [CrossRef]
- Harnett, D.; Ambrozkiewicz, M.C.; Zinnall, U.; Rusanova, A.; Borisova, E.; Drescher, A.N.; Couce-Iglesias, M.; Villamil, G.; Dannenberg, R.; Imami, K.; et al. A critical period of translational control during brain development at codon resolution. Nat. Struct. Mol. Biol. 2022, 29, 1277–1290. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.-Z.; Wang, S.-H.; Zhang, R.-H.; Lin, J.-H.; Tian, Y.-H.; Yang, Y.-Q.; Liu, J.; Ma, Y.-X. Neuroprotective effect of astragalin via activating PI3K/Akt-mTOR-mediated autophagy on APP/PS1 mice. Cell Death Discov. 2023, 9, 15. [Google Scholar] [CrossRef]
- Urrutia-Ruiz, C.; Rombach, D.; Cursano, S.; Gerlach-Arbeiter, S.; Schoen, M.; Bockmann, J.; Demestre, M.; Boeckers, T.M. Deletion of the Autism-Associated Protein SHANK3 Abolishes Structural Synaptic Plasticity after Brain Trauma. Int. J. Mol. Sci. 2022, 23, 6081. [Google Scholar] [CrossRef]
- Jakimovski, D.; Dorn, R.P.; Regno, M.D.; Bartnik, A.; Bergsland, N.; Ramanathan, M.; Dwyer, M.G.; Benedict, R.H.B.; Zivadinov, R.; Szigeti, K. Human restricted CHRFAM7A gene increases brain efficiency. Front. Neurosci. 2024, 18, 1359028. [Google Scholar] [CrossRef]
- Zeidán-Chuliá, F.; de Oliveira, B.-H.; Salmina, A.B.; Casanova, M.F.; Gelain, D.P.; Noda, M.; Verkhratsky, A.; Moreira, J.C. Altered expression of Alzheimer’s disease-related genes in the cerebellum of autistic patients: A model for disrupted brain connectome and therapy. Cell Death Dis. 2014, 5, e1250. [Google Scholar] [CrossRef]
- Jang, K.; Garraway, S.M. TrkB Agonist (7,8-DHF)-Induced Responses in Dorsal Root Ganglia Neurons Are Decreased after Spinal Cord Injury: Implication for Peripheral Pain Mechanisms. eNeuro 2025, 12, ENEURO.0219-24.2024. [Google Scholar] [CrossRef]
- McEachern, E.P.; Coley, A.A.; Yang, S.-S.; Gao, W.-J. PSD-95 deficiency alters GABAergic inhibition in the prefrontal cortex. Neuropharmacology 2020, 179, 108277. [Google Scholar] [CrossRef] [PubMed]
- Miyasaka, Y.; Yamamoto, N. Neuronal Activity Patterns Regulate Brain-Derived Neurotrophic Factor Expression in Cortical Cells via Neuronal Circuits. Front. Neurosci. 2021, 15, 699583. [Google Scholar] [CrossRef]
- Costa, G.A.; de Gusmão Taveiros Silva, N.K.; Marianno, P.; Chivers, P.; Bailey, A.; Camarini, R. Environmental Enrichment Increased Bdnf Transcripts in the Prefrontal Cortex: Implications for an Epigenetically Controlled Mechanism. Neuroscience 2023, 526, 277–289. [Google Scholar] [CrossRef]
- Martinez, J.D.; Donnelly, M.J.; Popke, D.S.; Torres, D.; Wilson, L.G.; Brancaleone, W.P.; Sheskey, S.; Lin, C.; Clawson, B.C.; Jiang, S.; et al. Enriched binocular experience followed by sleep optimally restores binocular visual cortical responses in a mouse model of amblyopia. Commun. Biol. 2023, 6, 408. [Google Scholar] [CrossRef] [PubMed]
- Qi, S.; Yu, J.; Li, L.; Dong, C.; Ji, Z.; Cao, L.; Wei, Z.; Liang, Z. Advances in non-invasive brain stimulation: Enhancing sports performance function and insights into exercise science. Front. Hum. Neurosci. 2024, 18, 1477111. [Google Scholar] [CrossRef] [PubMed]
- Consorti, A.; Sansevero, G.; Torelli, C.; Di Marco, I.; Berardi, N.; Sale, A. Visual Perceptual Learning Induces Long-Lasting Recovery of Visual Acuity, Visual Depth Perception Abilities and Binocular Matching in Adult Amblyopic Rats. Front. Cell. Neurosci. 2022, 16, 840708. [Google Scholar] [CrossRef]
- Toscano, C.V.A.; Ferreira, J.P.; Quinaud, R.T.; Silva, K.M.N.; Carvalho, H.M.; Gaspar, J.M. Exercise improves the social and behavioral skills of children and adolescent with autism spectrum disorders. Front. Psychiatry 2022, 13, 1027799. [Google Scholar] [CrossRef]
- Gogate, A.; Kaur, K.; Khalil, R.; Bashtawi, M.; Morris, M.A.; Goodspeed, K.; Evans, P.; Chahrour, M.H. The genetic landscape of autism spectrum disorder in an ancestrally diverse cohort. npj Genom. Med. 2024, 9, 62. [Google Scholar] [CrossRef]
- Nagappan-Chettiar, S.; Yasuda, M.; Johnson-Venkatesh, E.M.; Umemori, H. The molecular signals that regulate activity-dependent synapse refinement in the brain. Curr. Opin. Neurobiol. 2023, 79, 102692. [Google Scholar] [CrossRef]
- Gumus, C.; Yazici, I.P.; Yazici, K.U.; Ustundag, B. Increased Serum Brain-derived Neurotrophic Factor, Nerve Growth Factor, Glial-derived Neurotrophic Factor and Galanin Levels in Children with Attention Deficit Hyperactivity Disorder, and the Effect of 10 Weeks Methylphenidate Treatment. Clin. Psychopharmacol. Neurosci. 2022, 20, 635–648. [Google Scholar] [CrossRef]
- Volnova, A.; Kurzina, N.; Belskaya, A.; Gromova, A.; Pelevin, A.; Ptukha, M.; Fesenko, Z.; Ignashchenkova, A.; Gainetdinov, R.R. Noradrenergic Modulation of Learned and Innate Behaviors in Dopamine Transporter Knockout Rats by Guanfacine. Biomedicines 2023, 11, 222. [Google Scholar] [CrossRef] [PubMed]
- Jaehne, E.J.; Kent, J.N.; Antolasic, E.J.; Wright, B.J.; Spiers, J.G.; Creutzberg, K.C.; De Rosa, F.; Riva, M.A.; Sortwell, C.E.; Collier, T.J.; et al. Behavioral phenotyping of a rat model of the BDNF Val66Met polymorphism reveals selective impairment of fear memory. Transl Psychiatry 2022, 12, 93. [Google Scholar] [CrossRef]
- Kutsarova, E.; Schohl, A.; Munz, M.; Wang, A.; Zhang, Y.Y.; Bilash, O.M.; Ruthazer, E.S. BDNF signaling in correlation-dependent structural plasticity in the developing visual system. PLoS Biol. 2023, 21, e3002070. [Google Scholar] [CrossRef]
- Zhao, R.; Ren, B.; Xiao, Y.; Tian, J.; Zou, Y.; Wei, J.; Qi, Y.; Hu, A.; Xie, X.; Huang, Z.J.; et al. Axo-axonic synaptic input drives homeostatic plasticity by tuning the axon initial segment structurally and functionally. Sci. Adv. 2024, 10, eadk4331. [Google Scholar] [CrossRef] [PubMed]
- Myers, K.R.; Fan, Y.; McConnell, P.; Cooper, J.A.; Zheng, J.Q. Actin capping protein regulates postsynaptic spine development through CPI-motif interactions. Front. Mol. Neurosci. 2022, 15, 1020949. [Google Scholar] [CrossRef]
- Buendía, D.; Guncay, T.; Oyanedel, M.; Lemus, M.; Weinstein, A.; Ardiles, Á.O.; Marcos, J.; Fernandes, A.; Zângaro, R.; Muñoz, P. The Transcranial Light Therapy Improves Synaptic Plasticity in the Alzheimer’s Disease Mouse Model. Brain Sci. 2022, 12, 1272. [Google Scholar] [CrossRef] [PubMed]
- Martin Flores, N.; Podpolny, M.; McLeod, F.; Workman, I.; Crawford, K.; Ivanov, D.; Leonenko, G.; Escott-Price, V.; Salinas, P.C. Downregulation of Dickkopf-3, a Wnt antagonist elevated in Alzheimer’s disease, restores synapse integrity and memory in a disease mouse model. eLife 2024, 12, RP89453. [Google Scholar] [CrossRef]
- Warm, D.; Bassetti, D.; Schroer, J.; Luhmann, H.J.; Sinning, A. Spontaneous Activity Predicts Survival of Developing Cortical Neurons. Front. Cell Dev. Biol. 2022, 10, 937761. [Google Scholar] [CrossRef]
- Papadopoulou, M.A.; Rogdakis, T.; Charou, D.; Peteinareli, M.; Ntarntani, K.; Gravanis, A.; Chanoumidou, K.; Charalampopoulos, I. Neurotrophin Analog ENT-A044 Activates the p75 Neurotrophin Receptor, Regulating Neuronal Survival in a Cell Context-Dependent Manner. Int. J. Mol. Sci. 2023, 24, 11683. [Google Scholar] [CrossRef]
- Piromalli Girado, D.; Miranda, M.; Giachero, M.; Weisstaub, N.; Bekinschtein, P. Endocytosis is required for consolidation of pattern-separated memories in the perirhinal cortex. Front. Syst. Neurosci. 2023, 17, 1043664. [Google Scholar] [CrossRef]
- Carneiro, C.F.D.; Amorim, F.E.; Amaral, O.B. A meta-analysis of the effect of protein synthesis inhibitors on rodent fear conditioning. Peer Community J. 2023, 3, e110. [Google Scholar] [CrossRef]
- Gil-Marti, B.; Barredo, C.G.; Pina-Flores, S.; Trejo, J.L.; Turiegano, E.; Martin, F.A. The elusive transcriptional memory trace. Oxf. Open Neurosci. 2022, 1, kvac008. [Google Scholar] [CrossRef] [PubMed]
- Jaehne, E.J.; Antolasic, E.J.; Creutzberg, K.C.; Begni, V.; Riva, M.A.; van den Buuse, M. Impaired fear memory in a rat model of the brain-derived neurotrophic factor Val66Met polymorphism is reversed by chronic exercise. Neurobiol. Learn. Mem. 2023, 203, 107779. [Google Scholar] [CrossRef] [PubMed]
- Halonen, R.; Kuula, L.; Lahti, J.; Räikkönen, K.; Pesonen, A.-K. The association between overnight recognition accuracy and slow oscillation-spindle coupling is moderated by BDNF Val66Met. Behav. Brain Res. 2022, 428, 113889. [Google Scholar] [CrossRef]
- Vasileva, F.; Font-Lladó, R.; Carreras-Badosa, G.; López-Ros, V.; Ferrusola-Pastrana, A.; López-Bermejo, A.; Prats-Puig, A. Increased Salivary BDNF and Improved Fundamental Motor Skills in Children Following a 3-Month Integrated Neuromuscular Training in Primary School. J. Funct. Morphol. Kinesiol. 2024, 9, 154. [Google Scholar] [CrossRef]
- Geiger, L.S.; Wüstenberg, T.; Zang, Z.; Melzer, M.; Witt, S.H.; Rietschel, M.; Nöthen, M.M.; Herms, S.; Degenhardt, F.; Meyer-Lindenberg, A.; et al. Longitudinal markers of cognitive procedural learning in fronto-striatal circuits and putative effects of a BDNF plasticity-related variant. npj Sci. Learn. 2024, 9, 72. [Google Scholar] [CrossRef]
- Van Hook, M.J. Brain-derived neurotrophic factor is a regulator of synaptic transmission in the adult visual thalamus. J. Neurophysiol. 2022, 128, 1267–1277. [Google Scholar] [CrossRef]
- Rauti, R.; Cellot, G.; D’Andrea, P.; Colliva, A.; Scaini, D.; Tongiorgi, E.; Ballerini, L. BDNF impact on synaptic dynamics: Extra or intracellular long-term release differently regulates cultured hippocampal synapses. Mol. Brain 2020, 13, 43. [Google Scholar] [CrossRef]
- Ripoli, C.; Dagliyan, O.; Renna, P.; Pastore, F.; Paciello, F.; Sollazzo, R.; Rinaudo, M.; Battistoni, M.; Martini, S.; Tramutola, A.; et al. Engineering memory with an extrinsically disordered kinase. Sci. Adv. 2023, 9, eadh1110. [Google Scholar] [CrossRef]
- PA, H.; Basavaraju, N.; Chandran, M.; Jaleel, A.; Bennett, D.A.; Kommaddi, R.P. Mitigation of synaptic and memory impairments via F-actin stabilization in Alzheimer’s disease. Alzheimer’s Res. Ther. 2024, 16, 200. [Google Scholar] [CrossRef]
- Bonilla-Quintana, M.; Wörgötter, F. Exploring new roles for actin upon LTP induction in dendritic spines. Sci. Rep. 2021, 11, 7072. [Google Scholar] [CrossRef] [PubMed]
- Trujillo-Estrada, L.; Vanderklish, P.W.; Nguyen, M.M.T.; Kuang, R.R.; Nguyen, C.; Huynh, E.; da Cunha, C.; Javonillo, D.I.; Forner, S.; Martini, A.C.; et al. SPG302 Reverses Synaptic and Cognitive Deficits Without Altering Amyloid or Tau Pathology in a Transgenic Model of Alzheimer’s Disease. Neurotherapeutics 2021, 18, 2468–2483. [Google Scholar] [CrossRef] [PubMed]
- Choi, I.-A.; Yun, J.H.; Lee, J.; Choi, D.-H. Neuropeptide FF Promotes Neuronal Survival and Enhances Synaptic Protein Expression Following Ischemic Injury. Int. J. Mol. Sci. 2024, 25, 11580. [Google Scholar] [CrossRef] [PubMed]
- Tambaro, S.; Mitra, S.; Gera, R.; Linderoth, B.; Wahlberg, L.U.; Darreh-Shori, T.; Behbahani, H.; Nilsson, P.; Eriksdotter, M. Feasibility and therapeutical potential of local intracerebral encapsulated cell biodelivery of BDNF to AppNL−G−F knock-in Alzheimer mice. Alzheimer’s Res. Ther. 2023, 15, 137. [Google Scholar] [CrossRef]
- Park, K.S.; Buseth, L.; Hong, J.; Etnier, J.L. Music-based multicomponent exercise training for community-dwelling older adults with mild-to-moderate cognitive decline: A feasibility study. Front. Med. 2023, 10, 1224728. [Google Scholar] [CrossRef]
- Muñoz Ospina, B.; Cadavid-Ruiz, N. The effect of aerobic exercise on serum brain-derived neurotrophic factor (BDNF) and executive function in college students. Ment. Health Phys. Act. 2024, 26, 100578. [Google Scholar] [CrossRef]
- Enette, L.; Vogel, T.; Merle, S.; Valard-Guiguet, A.-G.; Ozier-Lafontaine, N.; Neviere, R.; Leuly-Joncart, C.; Fanon, J.L.; Lang, P.O. Effect of 9 weeks continuous vs. interval aerobic training on plasma BDNF levels, aerobic fitness, cognitive capacity and quality of life among seniors with mild to moderate Alzheimer’s disease: A randomized controlled trial. Eur. Rev. Aging Phys. Act. 2020, 17, 2. [Google Scholar] [CrossRef]
- Cheng, N.; Bell, L.; Lamport, D.J.; Williams, C.M. Dietary Flavonoids and Human Cognition: A Meta-Analysis. Mol. Nutr. Food Res. 2022, 66, 2100976. [Google Scholar] [CrossRef]
- Khor, K.L.; Kumarasuriar, V.; Tan, K.W.; Ooi, P.B.; Chia, Y.-C. Effects of fruit and vegetable intake on memory and attention: A systematic review of randomized controlled trials. Syst. Rev. 2024, 13, 151. [Google Scholar] [CrossRef]
- Rakotoarisoa, M.; Angelov, B.; Drechsler, M.; Nicolas, V.; Bizien, T.; Gorshkova, Y.E.; Deng, Y.; Angelova, A. Liquid crystalline lipid nanoparticles for combined delivery of curcumin, fish oil and BDNF: In vitro neuroprotective potential in a cellular model of tunicamycin-induced endoplasmic reticulum stress. Smart Mater. Med. 2022, 3, 274–288. [Google Scholar] [CrossRef]
- Sadat Razavi, Z.; Sina Alizadeh, S.; Sadat Razavi, F.; Souri, M.; Soltani, M. Advancing neurological disorders therapies: Organic nanoparticles as a key to blood-brain barrier penetration. Int. J. Pharm. 2025, 670, 125186. [Google Scholar] [CrossRef] [PubMed]
- McMoneagle, E.; Zhou, J.; Zhang, S.; Huang, W.; Josiah, S.S.; Ding, K.; Wang, Y.; Zhang, J. Neuronal K+-Cl− cotransporter KCC2 as a promising drug target for epilepsy treatment. Acta Pharmacol. Sin. 2024, 45, 1–22. [Google Scholar] [CrossRef]
- Trevelyan, A.J.; Marks, V.S.; Graham, R.T.; Denison, T.; Jackson, A.; Smith, E.H. On brain stimulation in epilepsy. Brain 2025, 148, 746–752. [Google Scholar] [CrossRef]
- Zhou, X.; Deng, X.; Liu, M.; He, M.; Long, W.; Xu, Z.; Zhang, K.; Liu, T.; So, K.-F.; Fu, Q.-L.; et al. Intranasal delivery of BDNF-loaded small extracellular vesicles for cerebral ischemia therapy. J. Control. Release 2023, 357, 1–19. [Google Scholar] [CrossRef]
- Saleem, S.; Banerjee, R.; Rajesh Kannan, R. Chrysin-Loaded Chitosan Nanoparticle-Mediated Neuroprotection in Aβ1-42-Induced Neurodegenerative Conditions in Zebrafish. ACS Chem. Neurosci. 2022, 13, 2017–2034. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Liu, S.-C.; Liu, Y.-Y.; Zhu, F.-Q.; Xiong, M.-J.; Hu, D.-X.; Zhang, W.-J. Therapeutic role of neural stem cells in neurological diseases. Front. Bioeng. Biotechnol. 2024, 12, 1329712. [Google Scholar] [CrossRef] [PubMed]
- Fatma, R.; Chauhan, W.; Shahi, M.H.; Afzal, M. Association of BDNF gene missense polymorphism rs6265 (Val66Met) with three quantitative traits, namely, intelligence quotient, body mass index, and blood pressure: A genetic association analysis from North India. Front. Neurol. 2023, 13, 1035885. [Google Scholar] [CrossRef]
- Kopanchuk, S.; Vavers, E.; Veiksina, S.; Ligi, K.; Zvejniece, L.; Dambrova, M.; Rinken, A. Intracellular dynamics of the Sigma-1 receptor observed with super-resolution imaging microscopy. PLoS ONE 2022, 17, e0268563. [Google Scholar] [CrossRef]
- Rao, B.; Li, S.; Yao, D.; Wang, Q.; Xia, Y.; Jia, Y.; Shen, Y.; Cao, Y. The cryo-EM structure of an ERAD protein channel formed by tetrameric human Derlin-1. Sci. Adv. 2021, 7, eabe8591. [Google Scholar] [CrossRef]
- Pré, D.; Wooten, A.T.; Biesmans, S.; Hinckley, S.; Zhou, H.; Sherman, S.P.; Kakad, P.; Gearhart, J.; Bang, A.G. Development of a platform to investigate long-term potentiation in human iPSC-derived neuronal networks. Stem Cell Rep. 2022, 17, 2141–2155. [Google Scholar] [CrossRef]
- Harrison, P.J.; Husain, S.M.; Lee, H.; Los Angeles, A.D.; Colbourne, L.; Mould, A.; Hall, N.A.L.; Haerty, W.; Tunbridge, E.M. CACNA1C (CaV1.2) and other L-type calcium channels in the pathophysiology and treatment of psychiatric disorders: Advances from functional genomics and pharmacoepidemiology. Neuropharmacology 2022, 220, 109262. [Google Scholar] [CrossRef] [PubMed]
- Wolf, D.; Ayon-Olivas, M.; Sendtner, M. BDNF-Regulated Modulation of Striatal Circuits and Implications for Parkinson’s Disease and Dystonia. Biomedicines 2024, 12, 1761. [Google Scholar] [CrossRef] [PubMed]
- Dubes, S.; Soula, A.; Benquet, S.; Tessier, B.; Poujol, C.; Favereaux, A.; Thoumine, O.; Letellier, M. miR-124-dependent tagging of synapses by synaptopodin enables input-specific homeostatic plasticity. EMBO J. 2022, 41, e109012. [Google Scholar] [CrossRef]
- Tian, T.; Li, J.; Zhang, G.; Wang, J.; Liu, D.; Wan, C.; Fang, J.; Wu, D.; Zhou, Y.; Zhu, W. Effects of childhood trauma experience and BDNF Val66Met polymorphism on brain plasticity relate to emotion regulation. Behav. Brain Res. 2021, 398, 112949. [Google Scholar] [CrossRef] [PubMed]
- Roddy, D.; Kelly, J.R.; Farrell, C.; Doolin, K.; Roman, E.; Nasa, A.; Frodl, T.; Harkin, A.; O’Mara, S.; O’Hanlon, E.; et al. Amygdala substructure volumes in Major Depressive Disorder. NeuroImage Clin. 2021, 31, 102781. [Google Scholar] [CrossRef]
- Kraguljac, N.V.; Guerreri, M.; Strickland, M.J.; Zhang, H. Neurite Orientation Dispersion and Density Imaging in Psychiatric Disorders: A Systematic Literature Review and a Technical Note. Biol. Psychiatry Glob. Open Sci. 2022, 3, 10–21. [Google Scholar] [CrossRef]
- Keeler, J.L.; Bahnsen, K.; Wronski, M.-L.; Bernardoni, F.; Tam, F.; Arold, D.; King, J.A.; Kolb, T.; Poitz, D.M.; Roessner, V.; et al. Longitudinal changes in brain-derived neurotrophic factor (BDNF) but not cytokines contribute to hippocampal recovery in anorexia nervosa above increases in body mass index. Psychol. Med. 2024, 54, 2242–2253. [Google Scholar] [CrossRef]
- Luna, F.G.; Lupiáñez, J.; Martín-Arévalo, E. Microstructural white matter connectivity underlying the attentional networks system. Behav. Brain Res. 2021, 401, 113079. [Google Scholar] [CrossRef]
- Jaehne, E.J.; Semaan, H.; Grosman, A.; Xu, X.; Schwarz, Q.; van den Buuse, M. Enhanced methamphetamine sensitisation in a rat model of the brain-derived neurotrophic factor Val66Met variant: Sex differences and dopamine receptor gene expression. Neuropharmacology 2023, 240, 109719. [Google Scholar] [CrossRef]
- Nicholson, E.L.; Garry, M.I.; Ney, L.J.; Hsu, C.-M.K.; Zuj, D.V.; Felmingham, K.L. The influence of the BDNF Val66Met genotype on emotional recognition memory in post-traumatic stress disorder. Sci. Rep. 2023, 13, 5033. [Google Scholar] [CrossRef]
- Fungaro Rissatti, L.; Wilson, D.; Palace-Berl, F.; de Mello Ponteciano, B.; Sardela de Miranda, F.; Alece Arantes Moreno, I.; dos Santos Vieira, T.; Pereira Sorroche, B.; Rebolho Batista Arantes, L.M.; Madeira Alvares da Silva, A.; et al. BDNF methylation associated with stress in women: Novel insights in epigenetics and inflammation. Brain Behav. Immun. Health 2024, 42, 100900. [Google Scholar] [CrossRef] [PubMed]
- Jeong, Y.; Cho, H.-Y.; Kim, M.; Oh, J.-P.; Kang, M.S.; Yoo, M.; Lee, H.-S.; Han, J.-H. Synaptic plasticity-dependent competition rule influences memory formation. Nat. Commun. 2021, 12, 3915. [Google Scholar] [CrossRef]
- Hou, W.; Qin, X.; Li, H.; Wang, Q.; Ding, Y.; Chen, X.; Wang, R.; Dong, F.; Bo, Q.; Li, X.; et al. Interaction between BDNF Val66Met polymorphism and mismatch negativity for working memory capacity in schizophrenia. Schizophr 2024, 10, 70. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, A.; Falkai, P.; Papiol, S. Neurodevelopmental disturbances in schizophrenia: Evidence from genetic and environmental factors. J. Neural Transm. 2023, 130, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Pisani, A.; Paciello, F.; Del Vecchio, V.; Malesci, R.; De Corso, E.; Cantone, E.; Fetoni, A.R. The Role of BDNF as a Biomarker in Cognitive and Sensory Neurodegeneration. J. Pers. Med. 2023, 13, 652. [Google Scholar] [CrossRef]
- Xavier, F.A.C.; Barbieri, S.S.; Popoli, M.; Ieraci, A. Short- and Long-Term Effects of Subchronic Stress Exposure in Male and Female Brain-Derived Neurotrophic Factor Knock-In Val66Met Mice. Biology 2024, 13, 303. [Google Scholar] [CrossRef]
- Shafik, A.M.; Allen, E.G.; Jin, P. Epitranscriptomic Dynamics in Brain Development and Disease. Mol. Psychiatry 2022, 27, 3633–3646. [Google Scholar] [CrossRef]
- Gutiérrez-Rodríguez, A.; Cruz-Fuentes, C.S.; Genis-Mendoza, A.D.; Nicolini, H. CRISPR/Cas9 genome editing approaches for psychiatric research. Braz. J. Psychiatry 2023, 45, 137–145. [Google Scholar] [CrossRef]
- Mielniczek, M.; Aune, T.K. The Effect of High-Intensity Interval Training (HIIT) on Brain-Derived Neurotrophic Factor Levels (BNDF): A Systematic Review. Brain Sci. 2025, 15, 34. [Google Scholar] [CrossRef]
- Kalogerakou, T.; Antoniadou, M. The Role of Dietary Antioxidants, Food Supplements and Functional Foods for Energy Enhancement in Healthcare Professionals. Antioxidants 2024, 13, 1508. [Google Scholar] [CrossRef]
- Vitaliano, G.D.; Kim, J.K.; Kaufman, M.J.; Adam, C.W.; Zeballos, G.; Shanmugavadivu, A.; Subburaju, S.; McLaughlin, J.P.; Lukas, S.E.; Vitaliano, F. Clathrin-nanoparticles deliver BDNF to hippocampus and enhance neurogenesis, synaptogenesis and cognition in HIV/neuroAIDS mouse model. Commun. Biol. 2022, 5, 236. [Google Scholar] [CrossRef] [PubMed]
- Narváez-Pérez, L.F.; Paz-Bermúdez, F.; Avalos-Fuentes, J.A.; Campos-Romo, A.; Florán-Garduño, B.; Segovia, J. CRISPR/sgRNA-directed synergistic activation mediator (SAM) as a therapeutic tool for Parkinson’s disease. Gene Ther. 2024, 31, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Barha, C.K.; Starkey, S.Y.; Hsiung, G.Y.R.; Tam, R.; Liu-Ambrose, T. Aerobic exercise improves executive functions in females, but not males, without the BDNF Val66Met polymorphism. Biol. Sex Differ. 2023, 14, 16. [Google Scholar] [CrossRef] [PubMed]
- El-Kadi, R.A.; AbdelKader, N.F.; Zaki, H.F.; Kamel, A.S. Vilazodone Alleviates Neurogenesis-Induced Anxiety in the Chronic Unpredictable Mild Stress Female Rat Model: Role of Wnt/β-Catenin Signaling. Mol. Neurobiol. 2024, 61, 9060–9077. [Google Scholar] [CrossRef]
- Latif-Hernandez, A.; Yang, T.; Butler, R.R.; Losada, P.M.; Minhas, P.S.; White, H.; Tran, K.C.; Liu, H.; Simmons, D.A.; Langness, V.; et al. A TrkB and TrkC partial agonist restores deficits in synaptic function and promotes activity-dependent synaptic and microglial transcriptomic changes in a late-stage Alzheimer’s mouse model. Alzheimer’s Dement. 2024, 20, 4434–4460. [Google Scholar] [CrossRef]
- Zhang, T.; Si, H.; Liao, J.; Ma, R. Association of plasma BDNF and MMP-9 levels with mild cognitive impairment: A matched case-control study. Sci. Rep. 2024, 14, 30911. [Google Scholar] [CrossRef]
- Yang, S.; Hu, J.; Chen, Y.; Zhang, Z.; Wang, J.; Zhu, G. DCC, a potential target for controlling fear memory extinction and hippocampal LTP in male mice receiving single prolonged stress. Neurobiol. Stress 2024, 32, 100666. [Google Scholar] [CrossRef]
- McCaughey-Chapman, A.; Burgers, A.L.; Combrinck, C.; Marriott, L.; Gordon, D.; Connor, B. Reprogrammed human lateral ganglionic eminence precursors generate striatal neurons and restore motor function in a rat model of Huntington’s disease. Stem Cell Res. Ther. 2024, 15, 448. [Google Scholar] [CrossRef]
- Shibata, A.C.E.; Ueda, H.H.; Eto, K.; Onda, M.; Sato, A.; Ohba, T.; Nabekura, J.; Murakoshi, H. Photoactivatable CaMKII induces synaptic plasticity in single synapses. Nat. Commun. 2021, 12, 751. [Google Scholar] [CrossRef]
- Gillies, N.A.; Wilson, B.C.; Miller, J.R.; Roy, N.C.; Scholey, A.; Braakhuis, A.J. Effects of a Flavonoid-Rich Blackcurrant Beverage on Markers of the Gut-Brain Axis in Healthy Females: Secondary Findings from a 4-Week Randomized Crossover Control Trial. Curr. Dev. Nutr. 2024, 8, 102158. [Google Scholar] [CrossRef]
- Song, J. BDNF Signaling in Vascular Dementia and Its Effects on Cerebrovascular Dysfunction, Synaptic Plasticity, and Cholinergic System Abnormality. J. Lipid Atheroscler. 2024, 13, 122–138. [Google Scholar] [CrossRef] [PubMed]
- Colom-Cadena, M.; Davies, C.; Sirisi, S.; Lee, J.-E.; Simzer, E.M.; Tzioras, M.; Querol-Vilaseca, M.; Sánchez-Aced, É.; Chang, Y.Y.; Holt, K.; et al. Synaptic oligomeric tau in Alzheimer’s disease—A potential culprit in the spread of tau pathology through the brain. Neuron 2023, 111, 2170–2183.e6. [Google Scholar] [CrossRef]
- Rinauro, D.J.; Chiti, F.; Vendruscolo, M.; Limbocker, R. Misfolded protein oligomers: Mechanisms of formation, cytotoxic effects, and pharmacological approaches against protein misfolding diseases. Mol. Neurodegener. 2024, 19, 20. [Google Scholar] [CrossRef] [PubMed]
- Lei, L.; Luo, Y.; Kang, D.; Yang, F.; Meng, D.; Wang, J.-Z.; Liu, R.; Wang, X.; Li, H.-L. Gypenoside IX restores Akt/GSK-3β pathway and alleviates Alzheimer’s disease-like neuropathology and cognitive deficits. Aging 2023, 15, 14172–14191. [Google Scholar] [CrossRef] [PubMed]
- Lyu, D.; Jia, J. Cryptotanshinone Attenuates Amyloid-β42-induced Tau Phosphorylation by Regulating PI3K/Akt/GSK3β Pathway in HT22 Cells. Mol. Neurobiol. 2022, 59, 4488–4500. [Google Scholar] [CrossRef]
- Cattaneo, M.; Baragetti, A.; Malovini, A.; Ciaglia, E.; Lopardo, V.; Olmastroni, E.; Casula, M.; Ciacci, C.; Catapano, A.L.; Puca, A.A. Longevity-associated BPIFB4 gene counteracts the inflammatory signaling. Immun. Ageing 2024, 21, 19. [Google Scholar] [CrossRef]
- El Ouaamari, Y.; Van den Bos, J.; Willekens, B.; Cools, N.; Wens, I. Neurotrophic Factors as Regenerative Therapy for Neurodegenerative Diseases: Current Status, Challenges and Future Perspectives. Int. J. Mol. Sci. 2023, 24, 3866. [Google Scholar] [CrossRef]
- Lu, S.; Ji, N.; Wang, W.; Lin, X.; Gao, D.; Geng, D. Salidroside improves cognitive function in Parkinson’s disease via Braf-mediated mitogen-activated protein kinase signaling pathway. Biomed. Pharmacother. 2024, 177, 116968. [Google Scholar] [CrossRef]
- Marchetti, B.; Giachino, C.; Tirolo, C.; Serapide, M.F. “Reframing” dopamine signaling at the intersection of glial networks in the aged Parkinsonian brain as innate Nrf2/Wnt driver: Therapeutical implications. Aging Cell 2022, 21, e13575. [Google Scholar] [CrossRef]
- Naren, P.; Samim, K.S.; Tryphena, K.P.; Vora, L.K.; Srivastava, S.; Singh, S.B.; Khatri, D.K. Microtubule acetylation dyshomeostasis in Parkinson’s disease. Transl. Neurodegener. 2023, 12, 20. [Google Scholar] [CrossRef]
- Kim, S.; Pajarillo, E.; Nyarko-Danquah, I.; Aschner, M.; Lee, E. Role of Astrocytes in Parkinson’s Disease Associated with Genetic Mutations and Neurotoxicants. Cells 2023, 12, 622. [Google Scholar] [CrossRef]
- van der Geest, A.T.; Jakobs, C.E.; Ljubikj, T.; Huffels, C.F.M.; Cañizares Luna, M.; Vieira de Sá, R.; Adolfs, Y.; de Wit, M.; Rutten, D.H.; Kaal, M.; et al. Molecular pathology, developmental changes and synaptic dysfunction in (pre-) symptomatic human C9ORF72-ALS/FTD cerebral organoids. Acta Neuropathol. Commun. 2024, 12, 152. [Google Scholar] [CrossRef]
- Kim, L.J.Y.; Kundu, B.; Moretti, P.; Lozano, A.M.; Rahimpour, S. Advancements in surgical treatments for Huntington disease: From pallidotomy to experimental therapies. Neurotherapeutics 2024, 21, e00452. [Google Scholar] [CrossRef] [PubMed]
- Piao, X.; Li, D.; Liu, H.; Guo, Q.; Yu, Y. Advances in gene and cellular therapeutic approaches for Huntington’s disease. Protein Cell 2024, pwae042. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, F.; Qin, D.; Chen, H.; Wang, J.; Wang, J.; Song, S.; Wang, C.; Wang, Y.; Liu, S.; et al. The role of brain derived neurotrophic factor in central nervous system. Front. Aging Neurosci. 2022, 14, 986443. [Google Scholar] [CrossRef] [PubMed]
- Salari, Z.; Ashabi, G.; Fartoosi, A.; Fartoosi, A.; Shariatpanahi, M.; Aghsami, M.; Montazeri, H.; Kheradmand, A. Sericin alleviates motor dysfunction by modulating inflammation and TrkB/BDNF signaling pathway in the rotenone-induced Parkinson’s disease model. BMC Pharmacol. Toxicol. 2023, 24, 60. [Google Scholar] [CrossRef]
- Benatar, M.; Robertson, J.; Andersen, P.M. Amyotrophic lateral sclerosis caused by SOD1 variants: From genetic discovery to disease prevention. Lancet Neurol. 2025, 24, 77–86. [Google Scholar] [CrossRef]
- Guan, T.; Zhou, T.; Zhang, X.; Guo, Y.; Yang, C.; Lin, J.; Zhang, J.V.; Cheng, Y.; Marzban, H.; Wang, Y.T.; et al. Selective removal of misfolded SOD1 delays disease onset in a mouse model of amyotrophic lateral sclerosis. Cell. Mol. Life Sci. 2023, 80, 304. [Google Scholar] [CrossRef]
- Zainullina, L.F.; Vakhitova, Y.V.; Lusta, A.Y.; Gudasheva, T.A.; Seredenin, S.B. Dimeric mimetic of BDNF loop 4 promotes survival of serum-deprived cell through TrkB-dependent apoptosis suppression. Sci. Rep. 2021, 11, 7781. [Google Scholar] [CrossRef]
- Forgham, H.; Liu, L.; Zhu, J.; Javed, I.; Cai, W.; Qiao, R.; Davis, T.P. Vector enabled CRISPR gene editing—A revolutionary strategy for targeting the diversity of brain pathologies. Coord. Chem. Rev. 2023, 487, 215172. [Google Scholar] [CrossRef]
- Washburn, H.R. Molecular Mechanisms of the EphB-NMD Receptor Extracellular Interaction. Ph.D. Thesis, Thomas Jefferson University, Philadelphia, PA, USA, 2021; pp. 1–299. [Google Scholar]
- Vidak, S.; Serebryannyy, L.A.; Pegoraro, G.; Misteli, T. Activation of endoplasmic reticulum stress in premature aging via the inner nuclear membrane protein SUN2. Cell Rep. 2023, 42, 112534. [Google Scholar] [CrossRef] [PubMed]
- Stykel, M.G.; Ryan, S.D. Network analysis of S-nitrosylated synaptic proteins demonstrates unique roles in health and disease. Biochim. Et Biophys. Acta (BBA) Mol. Cell Research 2024, 1871, 119720. [Google Scholar] [CrossRef]
- Luebke, M.; Parulekar, M.; Thomas, F.P. Fluid biomarkers for the diagnosis of neurodegenerative diseases. Biomark. Neuropsychiatry 2023, 8, 100062. [Google Scholar] [CrossRef]
- Toader, C.; Dobrin, N.; Brehar, F.-M.; Popa, C.; Covache-Busuioc, R.-A.; Glavan, L.A.; Costin, H.P.; Bratu, B.-G.; Corlatescu, A.D.; Popa, A.A.; et al. From Recognition to Remedy: The Significance of Biomarkers in Neurodegenerative Disease Pathology. Int. J. Mol. Sci. 2023, 24, 16119. [Google Scholar] [CrossRef]
- Dadkhah, M.; Saadat, M.; Ghorbanpour, A.M.; Moradikor, N. Experimental and clinical evidence of physical exercise on BDNF and cognitive function: A comprehensive review from molecular basis to therapy. Brain Behav. Immun. Integr. 2023, 3, 100017. [Google Scholar] [CrossRef]
- Weerasinghe-Mudiyanselage, P.D.E.; Ang, M.J.; Kang, S.; Kim, J.-S.; Moon, C. Structural Plasticity of the Hippocampus in Neurodegenerative Diseases. Int. J. Mol. Sci. 2022, 23, 3349. [Google Scholar] [CrossRef] [PubMed]
- Dell’Oste, V.; Palego, L.; Betti, L.; Fantasia, S.; Gravina, D.; Bordacchini, A.; Pedrinelli, V.; Giannaccini, G.; Carmassi, C. Plasma and Platelet Brain-Derived Neurotrophic Factor (BDNF) Levels in Bipolar Disorder Patients with Post-Traumatic Stress Disorder (PTSD) or in a Major Depressive Episode Compared to Healthy Controls. Int. J. Mol. Sci. 2024, 25, 3529. [Google Scholar] [CrossRef] [PubMed]
- Maiworm, M.; Koerbel, K.; Anschütz, V.; Jakob, J.; Schaller-Paule, M.A.; Schäfer, J.H.; Friedauer, L.; Wenger, K.J.; Hoelter, M.C.; Steffen, F.; et al. BDNF levels in serum and CSF are associated with clinicoradiological characteristics of aggressive disease in MS patients. J. Neurol. 2025, 272, 147. [Google Scholar] [CrossRef]
- Colvee-Martin, H.; Parra, J.R.; Gonzalez, G.A.; Barker, W.; Duara, R. Neuropathology, Neuroimaging, and Fluid Biomarkers in Alzheimer’s Disease. Diagnostics 2024, 14, 704. [Google Scholar] [CrossRef]
- Kaagman, D.G.M.; van Wegen, E.E.H.; Cignetti, N.; Rothermel, E.; Vanbellingen, T.; Hirsch, M.A. Effects and Mechanisms of Exercise on Brain-Derived Neurotrophic Factor (BDNF) Levels and Clinical Outcomes in People with Parkinson’s Disease: A Systematic Review and Meta-Analysis. Brain Sci. 2024, 14, 194. [Google Scholar] [CrossRef]
- Di Lazzaro, G.; Picca, A.; Boldrini, S.; Bove, F.; Marzetti, E.; Petracca, M.; Piano, C.; Bentivoglio, A.R.; Calabresi, P. Differential profiles of serum cytokines in Parkinson’s disease according to disease duration. Neurobiol. Dis. 2024, 190, 106371. [Google Scholar] [CrossRef] [PubMed]
- Harro, C.C.; Shoemaker, M.J.; Coatney, C.M.; Lentine, V.E.; Lieffers, L.R.; Quigley, J.J.; Rollins, S.G.; Stewart, J.D.; Hall, J.; Khoo, S.K. Effects of nordic walking exercise on gait, motor/non-motor symptoms, and serum brain-derived neurotrophic factor in individuals with Parkinson’s disease. Front. Rehabil. Sci. 2022, 3, 1010097. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Ge, M.; Liu, Y.; Tan, W.; Zhang, Y.; Zou, M.; Xiang, L.; Song, X.; Guo, H. From inflammation to depression: Key biomarkers for IBD-related major depressive disorder. J. Transl. Med. 2024, 22, 997. [Google Scholar] [CrossRef]
- Gaitán, J.M.; Moon, H.Y.; Stremlau, M.; Dubal, D.B.; Cook, D.B.; Okonkwo, O.C.; van Praag, H. Effects of Aerobic Exercise Training on Systemic Biomarkers and Cognition in Late Middle-Aged Adults at Risk for Alzheimer’s Disease. Front. Endocrinol. 2021, 12, 660181. [Google Scholar] [CrossRef]
- Ruiz-González, D.; Hernández-Martínez, A.; Valenzuela, P.L.; Morales, J.S.; Soriano-Maldonado, A. Effects of physical exercise on plasma brain-derived neurotrophic factor in neurodegenerative disorders: A systematic review and meta-analysis of randomized controlled trials. Neurosci. Biobehav. Rev. 2021, 128, 394–405. [Google Scholar] [CrossRef] [PubMed]
- Boi, L.; Johansson, Y.; Tonini, R.; Moratalla, R.; Fisone, G.; Silberberg, G. Serotonergic and dopaminergic neurons in the dorsal raphe are differentially altered in a mouse model for parkinsonism. eLife 2024, 12, RP90278. [Google Scholar] [CrossRef] [PubMed]
- Cavaleri, D.; Moretti, F.; Bartoccetti, A.; Mauro, S.; Crocamo, C.; Carrà, G.; Bartoli, F. The role of BDNF in major depressive disorder, related clinical features, and antidepressant treatment: Insight from meta-analyses. Neurosci. Biobehav. Rev. 2023, 149, 105159. [Google Scholar] [CrossRef]
- Targum, S.D.; Schappi, J.; Koutsouris, A.; Bhaumik, R.; Rapaport, M.H.; Rasgon, N.; Rasenick, M.M. A novel peripheral biomarker for depression and antidepressant response. Mol. Psychiatry 2022, 27, 1640–1646. [Google Scholar] [CrossRef]
- Sathyanarayanan, A.; Mueller, T.T.; Ali Moni, M.; Schueler, K.; Baune, B.T.; Lio, P.; Mehta, D.; Baune, B.T.; Dierssen, M.; Ebert, B.; et al. Multi-omics data integration methods and their applications in psychiatric disorders. Eur. Neuropsychopharmacol. 2023, 69, 26–46. [Google Scholar] [CrossRef]
- Rajan, K.B.; McAninch, E.A.; Aggarwal, N.T.; Barnes, L.L.; Wilson, R.S.; Weuve, J.; DeCarli, C.S.; Evans, D.A. Longitudinal Changes in Blood Biomarkers of Clinical Alzheimer Disease in a Biracial Population Sample. Neurology 2023, 100, e874–e883. [Google Scholar] [CrossRef]
- Li, Y.; Chen, J.; Yu, H.; Ye, J.; Wang, C.; Kong, L. Serum brain-derived neurotrophic factor as diagnosis clue for Alzheimer’s disease: A cross-sectional observational study in the elderly. Front. Psychiatry 2023, 14, 1127658. [Google Scholar] [CrossRef]
- Chatziefstathiou, A.; Canaslan, S.; Kanata, E.; Vekrellis, K.; Constantinides, V.C.; Paraskevas, G.P.; Kapaki, E.; Schmitz, M.; Zerr, I.; Xanthopoulos, K.; et al. SIMOA Diagnostics on Alzheimer’s Disease and Frontotemporal Dementia. Biomedicines 2024, 12, 1253. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Hu, Y.-Z.; Gao, S.; Wang, P.-F.; Hu, Z.-L.; Dai, R.-P. ProBDNF and its receptors in immune-mediated inflammatory diseases: Novel insights into the regulation of metabolism and mitochondria. Front. Immunol. 2023, 14, 1155333. [Google Scholar] [CrossRef]
- Nilsson, J.; Ekblom, M.; Moberg, M.; Lövdén, M. The role of acute changes in mBDNF, cortisol and pro-BDNF in predicting cognitive performance in old age. Sci. Rep. 2023, 13, 9418. [Google Scholar] [CrossRef]
- Shen, Y.; Timsina, J.; Heo, G.; Beric, A.; Ali, M.; Wang, C.; Yang, C.; Wang, Y.; Western, D.; Liu, M.; et al. CSF proteomics identifies early changes in autosomal dominant Alzheimer’s disease. Cell 2024, 187, 6309–6326.e15. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, X.; Yang, Z.; Wang, B.; Gong, H.; Zhang, K.; Lin, Y.; Sun, M. Extracellular vesicles: Biological mechanisms and emerging therapeutic opportunities in neurodegenerative diseases. Transl. Neurodegener. 2024, 13, 60. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Xiong, W.; Chen, Z.; Li, Y.; Xu, F.; Yang, X.; Long, M.; Guo, W.; Wu, S.; Sun, L.; et al. Exercise-conditioned plasma ameliorates postoperative cognitive dysfunction by activating hippocampal cholinergic circuit and enhancing BDNF/TrkB signaling. Cell Commun. Signal. 2024, 22, 551. [Google Scholar] [CrossRef]
- Jemni, M.; Zaman, R.; Carrick, F.R.; Clarke, N.D.; Marina, M.; Bottoms, L.; Matharoo, J.S.; Ramsbottom, R.; Hoffman, N.; Groves, S.J.; et al. Exercise improves depression through positive modulation of brain-derived neurotrophic factor (BDNF). A review based on 100 manuscripts over 20 years. Front. Physiol. 2023, 14, 1102526. [Google Scholar] [CrossRef]
- Colniță, A.; Toma, V.-A.; Brezeștean, I.A.; Tahir, M.A.; Dina, N.E. A Review on Integrated ZnO-Based SERS Biosensors and Their Potential in Detecting Biomarkers of Neurodegenerative Diseases. Biosensors 2023, 13, 499. [Google Scholar] [CrossRef]
- Guo, Z.-H.; Khattak, S.; Rauf, M.A.; Ansari, M.A.; Alomary, M.N.; Razak, S.; Yang, C.-Y.; Wu, D.-D.; Ji, X.-Y. Role of Nanomedicine-Based Therapeutics in the Treatment of CNS Disorders. Molecules 2023, 28, 1283. [Google Scholar] [CrossRef]
- Haberstroh, C.; Weider, S.; Flemmen, G.; Loe, H.; Andersson, H.W.; Hallgren, M.; Mosti, M.P. The effect of high-intensity interval training on cognitive function in patients with substance use disorder: Study protocol for a two-armed randomized controlled trial. Front. Sports Act. Living 2022, 4, 954561. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Liu, Y.; Li, S.; Chen, P.; Zhang, J.; Feng, L. Depletion of placental brain-derived neurotrophic factor (BDNF) is attributed to premature ovarian insufficiency (POI) in mice offspring. J. Ovarian Res. 2024, 17, 141. [Google Scholar] [CrossRef] [PubMed]
- Green, Z.D.; John, C.S.; Kueck, P.J.; Blankenship, A.E.; Kemna, R.E.; Johnson, C.N.; Yoksh, L.E.; Best, S.R.; Donald, J.S.; Mahnken, J.D.; et al. Acute exercise alters brain glucose metabolism in aging and Alzheimer’s disease. J. Physiol. 2024, early view. [CrossRef]
- Han, W.; Jiang, L.; Song, X.; Li, T.; Chen, H.; Cheng, L. VEGF Modulates Neurogenesis and Microvascular Remodeling in Epileptogenesis After Status Epilepticus in Immature Rats. Front. Neurol. 2021, 12, 808568. [Google Scholar] [CrossRef] [PubMed]
- Jaberi, S.; Fahnestock, M. Mechanisms of the Beneficial Effects of Exercise on Brain-Derived Neurotrophic Factor Expression in Alzheimer’s Disease. Biomolecules 2023, 13, 1577. [Google Scholar] [CrossRef]
- Schirò, G.; Iacono, S.; Ragonese, P.; Aridon, P.; Salemi, G.; Balistreri, C.R. A Brief Overview on BDNF-Trk Pathway in the Nervous System: A Potential Biomarker or Possible Target in Treatment of Multiple Sclerosis? Front. Neurol. 2022, 13, 917527. [Google Scholar] [CrossRef]
- Aghjayan, S.L.; Bournias, T.; Kang, C.; Zhou, X.; Stillman, C.M.; Donofry, S.D.; Kamarck, T.W.; Marsland, A.L.; Voss, M.W.; Fraundorf, S.H.; et al. Aerobic exercise improves episodic memory in late adulthood: A systematic review and meta-analysis. Commun. Med. 2022, 2, 15. [Google Scholar] [CrossRef]
- Latomme, J.; Calders, P.; Van Waelvelde, H.; Mariën, T.; De Craemer, M. The Role of Brain-Derived Neurotrophic Factor (BDNF) in the Relation between Physical Activity and Executive Functioning in Children. Children 2022, 9, 596. [Google Scholar] [CrossRef]
- Kim, S.; Park, D.; Kim, J.; Kim, D.; Kim, H.; Mori, T.; Jung, H.; Lee, D.; Hong, S.; Jeon, J.; et al. Npas4 regulates IQSEC3 expression in hippocampal somatostatin interneurons to mediate anxiety-like behavior. Cell Rep. 2021, 36, 109417. [Google Scholar] [CrossRef]
- Brown, D.T.; Vickers, J.C.; Stuart, K.E.; Cechova, K.; Ward, D.D. The BDNF Val66Met Polymorphism Modulates Resilience of Neurological Functioning to Brain Ageing and Dementia: A Narrative Review. Brain Sci. 2020, 10, 195. [Google Scholar] [CrossRef]
- Martella, G. Molecular Mechanisms of Synaptic Plasticity: Dynamic Changes in Neuron Functions. Int. J. Mol. Sci. 2023, 24, 12567. [Google Scholar] [CrossRef]
- Chang, N.; Li, J.; Lin, S.; Zhang, J.; Zeng, W.; Ma, G.; Wang, Y. Emerging roles of SIRT1 activator, SRT2104, in disease treatment. Sci. Rep. 2024, 14, 5521. [Google Scholar] [CrossRef] [PubMed]
- Panunggal, B.; Yeh, T.-H.; Tsao, S.-P.; Pan, C.-H.; Shih, W.-T.; Lin, Y.-T.; Faradina, A.; Fang, C.-L.; Huang, H.-Y.; Huang, S.-Y. Treadmill intervention attenuates motor deficit with 6-OHDA-induced Parkinson’s disease rat via changes in lipid profiles in brain and muscle. Aging 2025, 17, 232–250. [Google Scholar] [CrossRef] [PubMed]
- Mackay, C. The Effect of Aerobic Exercise on Brain Derived Neurotrophic Factor, Motor and Cognitive Function in People with Stroke. 2024. Ph.D. Thesis, School of Health and Rehabilitation Sciences, The University of Queensland, St Lucia, QLD, Australia. [CrossRef]
- Hijikata, N.; Kawakami, M.; Ishii, R.; Tsuzuki, K.; Nakamura, T.; Okuyama, K.; Liu, M. Item Difficulty of Fugl-Meyer Assessment for Upper Extremity in Persons with Chronic Stroke with Moderate-to-Severe Upper Limb Impairment. Front. Neurol. 2020, 11, 577855. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Gutiérrez, E.; Torres-Costoso, A.; Saz-Lara, A.; Bizzozero-Peroni, B.; Guzmán-Pavón, M.J.; Sánchez-López, M.; Martínez-Vizcaíno, V. Effectiveness of high-intensity interval training on peripheral brain-derived neurotrophic factor in adults: A systematic review and network meta-analysis. Scand. J. Med. Sci. Sports 2024, 34, e14496. [Google Scholar] [CrossRef]
- Paterno, A.; Polsinelli, G.; Federico, B. Changes of brain-derived neurotrophic factor (BDNF) levels after different exercise protocols: A systematic review of clinical studies in Parkinson’s disease. Front. Physiol. 2024, 15, 1352305. [Google Scholar] [CrossRef]
- Ross, R.E.; Saladin, M.E.; George, M.S.; Gregory, C.M. High-Intensity Aerobic Exercise Acutely Increases Brain-derived Neurotrophic Factor. Med. Sci. Sports Exerc. 2019, 51, 1698–1709. [Google Scholar] [CrossRef]
- Moriarty, T.A.; Mermier, C.; Kravitz, L.; Gibson, A.; Beltz, N.; Zuhl, M. Acute Aerobic Exercise Based Cognitive and Motor Priming: Practical Applications and Mechanisms. Front. Psychol. 2019, 10, 2790. [Google Scholar] [CrossRef]
- Tsai, C.-L.; Pan, C.-Y.; Tseng, Y.-T.; Chen, F.-C.; Chang, Y.-C.; Wang, T.-C. Acute effects of high-intensity interval training and moderate-intensity continuous exercise on BDNF and irisin levels and neurocognitive performance in late middle-aged and older adults. Behav. Brain Res. 2021, 413, 113472. [Google Scholar] [CrossRef]
- Cefis, M.; Chaney, R.; Wirtz, J.; Méloux, A.; Quirié, A.; Leger, C.; Prigent-Tessier, A.; Garnier, P. Molecular mechanisms underlying physical exercise-induced brain BDNF overproduction. Front. Mol. Neurosci. 2023, 16, 1275924. [Google Scholar] [CrossRef]
- Marzola, P.; Melzer, T.; Pavesi, E.; Gil-Mohapel, J.; Brocardo, P.S. Exploring the Role of Neuroplasticity in Development, Aging, and Neurodegeneration. Brain Sci. 2023, 13, 1610. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, L.; Zhang, Z.; Wang, Y.; Zuo, C.; Bo, S. A Shorter-Bout of HIIT Is More Effective to Promote Serum BDNF and VEGF-A Levels and Improve Cognitive Function in Healthy Young Men. Front. Physiol. 2022, 13, 898603. [Google Scholar] [CrossRef] [PubMed]
- Soga, K.; Higuchi, A.; Tomita, N.; Kobayashi, K.; Kataoka, H.; Imankulova, A.; Salazar, C.; Thyreau, B.; Nakamura, S.; Tsushita, Y.; et al. Beneficial Effects of a 26-Week Exercise Intervention Using IoT Devices on Cognitive Function and Health Indicators. J. Sci. Sport Exerc. 2025. [Google Scholar] [CrossRef]
- Pahlavani, H.A. Exercise therapy to prevent and treat Alzheimer’s disease. Front. Aging Neurosci. 2023, 15, 1243869. [Google Scholar] [CrossRef]
- Pisano, F.; Mellace, D.; Fugatti, A.; Aiello, E.N.; Diotti, S.; Curti, B.; Giust, A.; Marfoli, A.; Perin, C.; De Sandi, A.; et al. Cerebellar tDCS combined with augmented reality treadmill for freezing of gait in Parkinson’s disease: A randomized controlled trial. J. Neuroeng. Rehabil. 2024, 21, 173. [Google Scholar] [CrossRef]
- Jalouli, M.; Rahman, M.A.; Biswas, P.; Rahman, H.; Harrath, A.H.; Lee, I.-S.; Kang, S.; Choi, J.; Park, M.N.; Kim, B. Targeting natural antioxidant polyphenols to protect neuroinflammation and neurodegenerative diseases: A comprehensive review. Front. Pharmacol. 2025, 16, 1492517. [Google Scholar] [CrossRef] [PubMed]
- Paduchová, Z.; Katrenčíková, B.; Vaváková, M.; Laubertová, L.; Nagyová, Z.; Garaiova, I.; Ďuračková, Z.; Trebatická, J. The Effect of Omega-3 Fatty Acids on Thromboxane, Brain-Derived Neurotrophic Factor, Homocysteine, and Vitamin D in Depressive Children and Adolescents: Randomized Controlled Trial. Nutrients 2021, 13, 1095. [Google Scholar] [CrossRef]
- Butler, M.J.; Deems, N.P.; Muscat, S.; Butt, C.M.; Belury, M.A.; Barrientos, R.M. Dietary DHA prevents cognitive impairment and inflammatory gene expression in aged male rats fed a diet enriched with refined carbohydrates. Brain Behav. Immun. 2021, 98, 198–209. [Google Scholar] [CrossRef]
- Mathew, A.R.; Di Matteo, G.; La Rosa, P.; Barbati, S.A.; Mannina, L.; Moreno, S.; Tata, A.M.; Cavallucci, V.; Fidaleo, M. Vitamin B12 Deficiency and the Nervous System: Beyond Metabolic Decompensation—Comparing Biological Models and Gaining New Insights into Molecular and Cellular Mechanisms. Int. J. Mol. Sci. 2024, 25, 590. [Google Scholar] [CrossRef]
- Dong, H.; Sun, Y.; Nie, L.; Cui, A.; Zhao, P.; Leung, W.K.; Wang, Q. Metabolic memory: Mechanisms and diseases. Signal Transduct. Target. Ther. 2024, 9, 38. [Google Scholar] [CrossRef]
- Pannu, A.; Sharma, P.C.; Thakur, V.K.; Goyal, R.K. Emerging Role of Flavonoids as the Treatment of Depression. Biomolecules 2021, 11, 1825. [Google Scholar] [CrossRef]
- Tanase, D.M.; Apostol, A.G.; Costea, C.F.; Tarniceriu, C.C.; Tudorancea, I.; Maranduca, M.A.; Floria, M.; Serban, I.L. Oxidative Stress in Arterial Hypertension (HTN): The Nuclear Factor Erythroid Factor 2-Related Factor 2 (Nrf2) Pathway, Implications and Future Perspectives. Pharmaceutics 2022, 14, 534. [Google Scholar] [CrossRef] [PubMed]
- Çelik, M.; Ceylan, M.R.; Koyuncu, İ.; Dinçer, N.G.; Alkan, S. Nuclear Factor Erythroid 2-Related Factor (NRF2), Heme Oxygenase 1 (HO-1) and Total Oxidant-Antioxidant Status in Patients with COVID-19. Iberoam. J. Med. 2024, 6, 3–9. [Google Scholar] [CrossRef]
- Szulc, A.; Wiśniewska, K.; Żabińska, M.; Gaffke, L.; Szota, M.; Olendzka, Z.; Węgrzyn, G.; Pierzynowska, K. Effectiveness of Flavonoid-Rich Diet in Alleviating Symptoms of Neurodegenerative Diseases. Foods 2024, 13, 1931. [Google Scholar] [CrossRef] [PubMed]
- RGarcía-Villalba, R.; Giménez-Bastida, J.A.; Cortés-Martín, A.; Ávila-Gálvez, M.Á.; Tomás-Barberán, F.A.; Selma, M.V.; Espín, J.C.; González-Sarrías, A. Urolithins: A Comprehensive Update on their Metabolism, Bioactivity, and Associated Gut Microbiota. Mol. Nutr. Food Res. 2022, 66, 2101019. [Google Scholar] [CrossRef]
- Carrillo, J.Á.; Arcusa, R.; Xandri-Martínez, R.; Cerdá, B.; Zafrilla, P.; Marhuenda, J. Impact of Polyphenol-Rich Nutraceuticals on Cognitive Function and Neuroprotective Biomarkers: A Randomized, Double-Blind, Placebo-Controlled Clinical Trial. Nutrients 2025, 17, 601. [Google Scholar] [CrossRef] [PubMed]
- Clemente-Suárez, V.J.; Martín-Rodríguez, A.; Curiel-Regueros, A.; Rubio-Zarapuz, A.; Tornero-Aguilera, J.F. Neuro-Nutrition and Exercise Synergy: Exploring the Bioengineering of Cognitive Enhancement and Mental Health Optimization. Bioengineering 2025, 12, 208. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Rojo, I.C.; Cuesta, P.; López, M.E.; de Frutos-Lucas, J.; Bruña, R.; Pereda, E.; Barabash, A.; Montejo, P.; Montenegro-Peña, M.; Marcos, A.; et al. BDNF Val66Met Polymorphism and Gamma Band Disruption in Resting State Brain Functional Connectivity: A Magnetoencephalography Study in Cognitively Intact Older Females. Front. Neurosci. 2018, 12, 684. [Google Scholar] [CrossRef]
- Mallick, R.; Duttaroy, A.K. Epigenetic modification impacting brain functions: Effects of physical activity, micronutrients, caffeine, toxins, and addictive substances. Neurochem. Int. 2023, 171, 105627. [Google Scholar] [CrossRef]
- Arjunan, A.; Song, J. Pharmacological and physiological roles of adipokines and myokines in metabolic-related dementia. Biomed. Pharmacother. 2023, 163, 114847. [Google Scholar] [CrossRef]
- Ichimura-Shimizu, M.; Kurrey, K.; Miyata, M.; Dezawa, T.; Tsuneyama, K.; Kojima, M. Emerging Insights into the Role of BDNF on Health and Disease in Periphery. Biomolecules 2024, 14, 444. [Google Scholar] [CrossRef]
- Cano, R.; Bermúdez, V.; Galban, N.; Garrido, B.; Santeliz, R.; Gotera, M.P.; Duran, P.; Boscan, A.; Carbonell-Zabaleta, A.-K.; Durán-Agüero, S.; et al. Dietary Polyphenols and Gut Microbiota Cross-Talk: Molecular and Therapeutic Perspectives for Cardiometabolic Disease: A Narrative Review. Int. J. Mol. Sci. 2024, 25, 9118. [Google Scholar] [CrossRef] [PubMed]
- Rahmatkar, S.N.; Singh, D. Decoding the Role of Neurotrophins in Glycogen Synthase Kinase 3-Beta Regulation in Alzheimer’s Disease. Mol. Neurobiol. 2025. [Google Scholar] [CrossRef] [PubMed]
- Basnet, J.; Eissa, M.A.; Yanes Cardozo, L.L.; Romero, D.G.; Rezq, S. Impact of Probiotics and Prebiotics on Gut Microbiome and Hormonal Regulation. Gastrointest. Disord. 2024, 6, 801–815. [Google Scholar] [CrossRef] [PubMed]
- Numakawa, T.; Kajihara, R. An Interaction between Brain-Derived Neurotrophic Factor and Stress-Related Glucocorticoids in the Pathophysiology of Alzheimer’s Disease. Int. J. Mol. Sci. 2024, 25, 1596. [Google Scholar] [CrossRef]
- Pickersgill, J.W.; Turco, C.V.; Ramdeo, K.; Rehsi, R.S.; Foglia, S.D.; Nelson, A.J. The Combined Influences of Exercise, Diet and Sleep on Neuroplasticity. Front. Psychol. 2022, 13, 831819. [Google Scholar] [CrossRef]
- Johne, M.; Maculewicz, E.; Mastalerz, A.; Białek, M.; Wojtak, W.; Osuch, B.; Majewska, M.; Czauderna, M.; Białek, A. Dietary Patterns, Serum BDNF and Fatty Acid Profiles in Physically Active Male Young Adults: A Cluster Analysis Study. Nutrients 2024, 16, 4326. [Google Scholar] [CrossRef]
- Rathor, P.; Ch, R. The Impacts of Dietary Intervention on Brain Metabolism and Neurological Disorders: A Narrative Review. Dietetics 2024, 3, 289–307. [Google Scholar] [CrossRef]
- Armeli, F.; Bonucci, A.; Maggi, E.; Pinto, A.; Businaro, R. Mediterranean Diet and Neurodegenerative Diseases: The Neglected Role of Nutrition in the Modulation of the Endocannabinoid System. Biomolecules 2021, 11, 790. [Google Scholar] [CrossRef]
- Coelho-Júnior, H.J.; Trichopoulou, A.; Panza, F. Cross-sectional and longitudinal associations between adherence to Mediterranean diet with physical performance and cognitive function in older adults: A systematic review and meta-analysis. Ageing Res. Rev. 2021, 70, 101395. [Google Scholar] [CrossRef]
- Whyte, A.R.; Cheng, N.; Butler, L.T.; Lamport, D.J.; Williams, C.M. Flavonoid-Rich Mixed Berries Maintain and Improve Cognitive Function Over a 6 h Period in Young Healthy Adults. Nutrients 2019, 11, 2685. [Google Scholar] [CrossRef]
- Sittiprapaporn, P.; Bumrungpert, A.; Suyajai, P.; Stough, C. Effectiveness of Fish Oil-DHA Supplementation for Cognitive Function in Thai Children: A Randomized, Doubled-Blind, Two-Dose, Placebo-Controlled Clinical Trial. Foods 2022, 11, 2595. [Google Scholar] [CrossRef]
- Mahalakshmi, A.M.; Ray, B.; Tuladhar, S.; Hediyal, T.A.; Raj, P.; Rathipriya, A.G.; Qoronfleh, M.W.; Essa, M.M.; Chidambaram, S.B. Impact of Pharmacological and Non-Pharmacological Modulators on Dendritic Spines Structure and Functions in Brain. Cells 2021, 10, 3405. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Pang, J.; Zhao, J.; Xiao, X.; Li, J.; Li, J.; Wang, W.; Zhou, S.; Zhao, Y.; Zhang, Z.; et al. Unveiling the neuroprotective potential of dietary polysaccharides: A systematic review. Front. Nutr. 2023, 10, 1299117. [Google Scholar] [CrossRef] [PubMed]
- Albadrani, H.M.; Chauhan, P.; Ashique, S.; Babu, M.A.; Iqbal, D.; Almutary, A.G.; Abomughaid, M.M.; Kamal, M.; Paiva-Santos, A.C.; Alsaweed, M.; et al. Mechanistic insights into the potential role of dietary polyphenols and their nanoformulation in the management of Alzheimer’s disease. Biomed. Pharmacother. 2024, 174, 116376. [Google Scholar] [CrossRef]
- Grabska-Kobyłecka, I.; Szpakowski, P.; Król, A.; Książek-Winiarek, D.; Kobyłecki, A.; Głąbiński, A.; Nowak, D. Polyphenols and Their Impact on the Prevention of Neurodegenerative Diseases and Development. Nutrients 2023, 15, 3454. [Google Scholar] [CrossRef]
- Chitre, N.M.; Wood, B.J.; Ray, A.; Moniri, N.H.; Murnane, K.S. Docosahexaenoic acid protects motor function and increases dopamine synthesis in a rat model of Parkinson’s disease via mechanisms associated with increased protein kinase activity in the striatum. Neuropharmacology 2020, 167, 107976. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Xiong, J.-Y.; Chai, Y.-Q.; Huang, L.; Tang, Z.-Y.; Zhang, X.-F.; Liu, B.; Zhang, J.-T. Possible antidepressant mechanisms of omega-3 polyunsaturated fatty acids acting on the central nervous system. Front. Psychiatry 2022, 13, 933704. [Google Scholar] [CrossRef]
- Correia, A.S.; Cardoso, A.; Vale, N. BDNF Unveiled: Exploring Its Role in Major Depression Disorder Serotonergic Imbalance and Associated Stress Conditions. Pharmaceutics 2023, 15, 2081. [Google Scholar] [CrossRef]
- Dyńka, D.; Kowalcze, K.; Paziewska, A. The Role of Ketogenic Diet in the Treatment of Neurological Diseases. Nutrients 2022, 14, 5003. [Google Scholar] [CrossRef]
- Mansour, S.; Alkhaaldi, S.M.I.; Sammanasunathan, A.F.; Ibrahim, S.; Farhat, J.; Al-Omari, B. Precision Nutrition Unveiled: Gene–Nutrient Interactions, Microbiota Dynamics, and Lifestyle Factors in Obesity Management. Nutrients 2024, 16, 581. [Google Scholar] [CrossRef]
- Lin, P.-H.; Kuo, L.-T.; Luh, H.-T. The Roles of Neurotrophins in Traumatic Brain Injury. Life 2021, 12, 26. [Google Scholar] [CrossRef] [PubMed]
- Qiu, S.; Cai, Y.; Yao, H.; Lin, C.; Xie, Y.; Tang, S.; Zhang, A. Small molecule metabolites: Discovery of biomarkers and therapeutic targets. Signal Transduct. Target. Ther. 2023, 8, 132. [Google Scholar] [CrossRef]
- Gudden, J.; Arias Vasquez, A.; Bloemendaal, M. The Effects of Intermittent Fasting on Brain and Cognitive Function. Nutrients 2021, 13, 3166. [Google Scholar] [CrossRef]
- Seidler, K.; Barrow, M. Intermittent fasting and cognitive performance—Targeting BDNF as potential strategy to optimise brain health. Front. Neuroendocrinol. 2022, 65, 100971. [Google Scholar] [CrossRef]
- Kassem, H.; Beevi, A.A.; Basheer, S.; Lutfi, G.; Cheikh Ismail, L.; Papandreou, D. Investigation and Assessment of AI’s Role in Nutrition—An Updated Narrative Review of the Evidence. Nutrients 2025, 17, 190. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, G.V.; Niaz, H.; Bai, R.; Kim, D.H.; Kim, J.W.; Asghar, J.; Ramzan, T.; Maqbool, M.; Abushalha, N.B.; Arif, S.; et al. The Role of Omega-3 Fatty Acid Supplementation in Slowing Cognitive Decline Among Elderly Patients With Alzheimer’s Disease: A Systematic Review of Randomized Controlled Trials. Cureus 2024, 16, e73390. [Google Scholar] [CrossRef]
- Clemente-Suárez, V.J.; Redondo-Flórez, L.; Beltrán-Velasco, A.I.; Belinchón-deMiguel, P.; Ramos-Campo, D.J.; Curiel-Regueros, A.; Martín-Rodríguez, A.; Tornero-Aguilera, J.F. The Interplay of Sports and Nutrition in Neurological Health and Recovery. J. Clin. Med. 2024, 13, 2065. [Google Scholar] [CrossRef]
- Lugenbühl, J.F.; Viho, E.M.G.; Binder, E.B.; Daskalakis, N.P. Stress Molecular Signaling in Interaction With Cognition. Biol. Psychiatry 2025, 97, 349–358. [Google Scholar] [CrossRef]
- Campbell, T.S.; Donoghue, K.M.; Ghosh, U.; Nelson, C.M.; Roth, T.L. Early Life Stress Affects Bdnf Regulation: A Role for Exercise Interventions. Int. J. Mol. Sci. 2022, 23, 11729. [Google Scholar] [CrossRef]
- Song, M.; Bai, Y.; Song, F. High-fat diet and neuroinflammation: The role of mitochondria. Pharmacol. Res. 2025, 212, 107615. [Google Scholar] [CrossRef]
- Fang, S.; Wu, Z.; Guo, Y.; Zhu, W.; Wan, C.; Yuan, N.; Chen, J.; Hao, W.; Mo, X.; Guo, X.; et al. Roles of microglia in adult hippocampal neurogenesis in depression and their therapeutics. Front. Immunol. 2023, 14, 1193053. [Google Scholar] [CrossRef] [PubMed]
- Ruggiero, R.N.; Rossignoli, M.T.; Marques, D.B.; de Sousa, B.M.; Romcy-Pereira, R.N.; Lopes-Aguiar, C.; Leite, J.P. Neuromodulation of Hippocampal-Prefrontal Cortical Synaptic Plasticity and Functional Connectivity: Implications for Neuropsychiatric Disorders. Front. Cell. Neurosci. 2021, 15, 732360. [Google Scholar] [CrossRef] [PubMed]
- Herhaus, B.; Heni, M.; Bloch, W.; Petrowski, K. Dynamic interplay of cortisol and BDNF in males under acute and chronic psychosocial stress—A randomized controlled study. Psychoneuroendocrinology 2024, 170, 107192. [Google Scholar] [CrossRef]
- Hernández-del Caño, C.; Varela-Andrés, N.; Cebrián-León, A.; Deogracias, R. Neurotrophins and Their Receptors: BDNF’s Role in GABAergic Neurodevelopment and Disease. Int. J. Mol. Sci. 2024, 25, 8312. [Google Scholar] [CrossRef] [PubMed]
- Shan, M.A.; Khan, M.U.; Ishtiaq, W.; Rehman, R.; Khan, S.; Javed, M.A.; Ali, Q. In silico analysis of the Val66Met mutation in BDNF protein: Implications for psychological stress. AMB Express 2024, 14, 11. [Google Scholar] [CrossRef]
- Pardossi, S.; Fagiolini, A.; Cuomo, A. Variations in BDNF and Their Role in the Neurotrophic Antidepressant Mechanisms of Ketamine and Esketamine: A Review. Int. J. Mol. Sci. 2024, 25, 13098. [Google Scholar] [CrossRef]
- Madsen, C.A.; Navarro, M.L.; Elfving, B.; Kessing, L.V.; Castrén, E.; Mikkelsen, J.D.; Knudsen, G.M. The effect of antidepressant treatment on blood BDNF levels in depressed patients: A review and methodological recommendations for assessment of BDNF in blood. Eur. Neuropsychopharmacol. 2024, 87, 35–55. [Google Scholar] [CrossRef]
- Antolasic, E.; Jaehne, E.; Buuse, M. van den Interaction of Brain-derived Neurotrophic Factor, Exercise, and Fear Extinction: Implications for Post-traumatic Stress Disorder. Curr. Neuropharmacol. 2024, 22, 543–556. [Google Scholar] [CrossRef]
- Liu, J.; Meng, F.; Dai, J.; Wu, M.; Wang, W.; Liu, C.; Zhao, D.; Wang, H.; Zhang, J.; Li, C. The BDNF-FoxO1 Axis in the medial prefrontal cortex modulates depressive-like behaviors induced by chronic unpredictable stress in postpartum female mice. Mol. Brain 2020, 13, 91. [Google Scholar] [CrossRef]
- Rodrigues, R.S.; Moreira, J.B.; Mateus, J.M.; Barateiro, A.; Paulo, S.L.; Vaz, S.H.; Lourenço, D.M.; Ribeiro, F.F.; Soares, R.; Loureiro-Campos, E.; et al. Cannabinoid type 2 receptor inhibition enhances the antidepressant and proneurogenic effects of physical exercise after chronic stress. Transl. Psychiatry 2024, 14, 170. [Google Scholar] [CrossRef]
- Linz, R.; Puhlmann, L.M.C.; Apostolakou, F.; Mantzou, E.; Papassotiriou, I.; Chrousos, G.P.; Engert, V.; Singer, T. Acute psychosocial stress increases serum BDNF levels: An antagonistic relation to cortisol but no group differences after mental training. Neuropsychopharmacology 2019, 44, 1797–1804. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, H.T.H.; Wood, R.J.; Prawdiuk, A.R.; Furness, S.G.B.; Xiao, J.; Murray, S.S.; Fletcher, J.L. TrkB Agonist LM22A-4 Increases Oligodendroglial Populations During Myelin Repair in the Corpus Callosum. Front. Mol. Neurosci. 2019, 12, 205. [Google Scholar] [CrossRef]
- Borsellino, P.; Krider, R.I.; Chea, D.; Grinnell, R.; Vida, T.A. Ketamine and the Disinhibition Hypothesis: Neurotrophic Factor-Mediated Treatment of Depression. Pharmaceuticals 2023, 16, 742. [Google Scholar] [CrossRef] [PubMed]
- Sanilevici, M.; Reuveni, O.; Lev-Ari, S.; Golland, Y.; Levit-Binnun, N. Mindfulness-Based Stress Reduction Increases Mental Wellbeing and Emotion Regulation During the First Wave of the COVID-19 Pandemic: A Synchronous Online Intervention Study. Front. Psychol. 2021, 12, 720965. [Google Scholar] [CrossRef]
- Hang, P.-Z.; Zhu, H.; Li, P.-F.; Liu, J.; Ge, F.-Q.; Zhao, J.; Du, Z.-M. The Emerging Role of BDNF/TrkB Signaling in Cardiovascular Diseases. Life 2021, 11, 70. [Google Scholar] [CrossRef]
- Medeiros, D.; Ayala-Baylon, K.; Egido-Betancourt, H.; Miller, E.; Chapleau, C.; Robinson, H.; Phillips, M.L.; Yang, T.; Longo, F.M.; Li, W.; et al. A small-molecule TrkB ligand improves dendritic spine phenotypes and atypical behaviors in female Rett syndrome mice. Dis. Model. Mech. 2024, 17, dmm050612. [Google Scholar] [CrossRef]
- Mead, R.J.; Shan, N.; Reiser, H.J.; Marshall, F.; Shaw, P.J. Amyotrophic lateral sclerosis: A neurodegenerative disorder poised for successful therapeutic translation. Nat. Rev. Drug Discov. 2023, 22, 185–212. [Google Scholar] [CrossRef]
- Kishimoto, Y.; Johnson, J.; Fang, W.; Halpern, J.; Marosi, K.; Liu, D.; Geisler, J.G.; Mattson, M.P. A mitochondrial uncoupler prodrug protects dopaminergic neurons and improves functional outcome in a mouse model of Parkinson’s disease. Neurobiol. Aging 2020, 85, 123–130. [Google Scholar] [CrossRef]
- Cubillos, S.; Engmann, O.; Brancato, A. BDNF as a Mediator of Antidepressant Response: Recent Advances and Lifestyle Interactions. Int. J. Mol. Sci. 2022, 23, 14445. [Google Scholar] [CrossRef]
- Kurita, M.; Nishino, S.; Kato, M.; Numata, Y.; Sato, T. Plasma brain-derived neurotrophic factor levels predict the clinical outcome of depression treatment in a naturalistic study. PLoS ONE 2012, 7, e39212. [Google Scholar] [CrossRef]
- Riggs, L.M.; Gould, T.D. Ketamine and the Future of Rapid-Acting Antidepressants. Annu. Rev. Clin. Psychol. 2021, 17, 207–231. [Google Scholar] [CrossRef] [PubMed]
- Vecera, C.M.; Courtes, A.A.; Jones, G.; Soares, J.C.; Machado-Vieira, R. Pharmacotherapies Targeting GABA-Glutamate Neurotransmission for Treatment-Resistant Depression. Pharmaceuticals 2023, 16, 1572. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Zhang, Y.; Sterling, K.; Song, W. Brain-derived neurotrophic factor in Alzheimer’s disease and its pharmaceutical potential. Transl. Neurodegener. 2022, 11, 4. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, H.; Li, R.; Sterling, K.; Song, W. Amyloid β-based therapy for Alzheimer’s disease: Challenges, successes and future. Signal Transduct. Target. Ther. 2023, 8, 248. [Google Scholar] [CrossRef]
- Ali, N.H.; Al-Kuraishy, H.M.; Al-Gareeb, A.I.; Alexiou, A.; Papadakis, M.; AlAseeri, A.A.; Alruwaili, M.; Saad, H.M.; Batiha, G.E. BDNF/TrkB activators in Parkinson’s disease: A new therapeutic strategy. J. Cell. Mol. Med. 2024, 28, e18368. [Google Scholar] [CrossRef] [PubMed]
- Maloney, M.T.; Wang, W.; Bhowmick, S.; Millan, I.; Kapur, M.; Herrera, N.; Frost, E.; Zhang, E.Y.; Song, S.; Wang, M.; et al. Failure to Thrive: Impaired BDNF Transport along the Cortical–Striatal Axis in Mouse Q140 Neurons of Huntington’s Disease. Biology 2023, 12, 157. [Google Scholar] [CrossRef]
- Meyer, R.A.; Neshat, S.Y.; Green, J.J.; Santos, J.L.; Tuesca, A.D. Targeting strategies for mRNA delivery. Mater. Today Adv. 2022, 14, 100240. [Google Scholar] [CrossRef]
- Rahman, M.; Bilal, M.; Shah, J.A.; Kaushik, A.; Teissedre, P.-L.; Kujawska, M. CRISPR-Cas9-Based Technology and Its Relevance to Gene Editing in Parkinson’s Disease. Pharmaceutics 2022, 14, 1252. [Google Scholar] [CrossRef]
- Hamilton, P.J.; Lim, C.J.; Nestler, E.J.; Heller, E.A. Neuroepigenetic Editing. Methods Mol. Biol. 2024, 2842, 129–152. [Google Scholar] [CrossRef]
- Koijam, A.S.; Singh, K.D.; Nameirakpam, B.S.; Haobam, R.; Rajashekar, Y. Drug addiction and treatment: An epigenetic perspective. Biomed. Pharmacother. 2024, 170, 115951. [Google Scholar] [CrossRef]
- Shafique, H.; Demers, J.C.; Biesiada, J.; Golani, L.K.; Cerne, R.; Smith, J.L.; Szostak, M.; Witkin, J.M. (R)-(-)-Ketamine: The Promise of a Novel Treatment for Psychiatric and Neurological Disorders. Int. J. Mol. Sci. 2024, 25, 6804. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.S.; Wu, Z.; Liu, X.; Edgington-Mitchell, L.; Ye, K. Treating Parkinson’s Disease via Activation of BDNF/TrkB Signaling Pathways and Inhibition of Delta-Secretase. Neurotherapeutics 2022, 19, 1283–1297. [Google Scholar] [CrossRef] [PubMed]
- Sharbafshaaer, M.; Cirillo, G.; Esposito, F.; Tedeschi, G.; Trojsi, F. Harnessing Brain Plasticity: The Therapeutic Power of Repetitive Transcranial Magnetic Stimulation (rTMS) and Theta Burst Stimulation (TBS) in Neurotransmitter Modulation, Receptor Dynamics, and Neuroimaging for Neurological Innovations. Biomedicines 2024, 12, 2506. [Google Scholar] [CrossRef] [PubMed]
- Zoladz, P.R.; Riggenbach, M.R.; Weiser, J.N.; Hipskind, J.J.; Wireman, L.E.; Hess, K.L.; Duffy, T.J.; Handel, J.K.; Kaschalk, M.G.; Reneau, K.E.; et al. Female Carriers of the Met Allele of the BDNF Val66Met Polymorphism Develop Weaker Fear Memories in a Fear-Potentiated Startle Paradigm. J. Behav. Brain Sci. 2022, 12, 211–235. [Google Scholar] [CrossRef]
- Chaldakov, G.N.; Aloe, L.; Yanev, S.G.; Fiore, M.; Tonchev, A.B.; Vinciguerra, M.; Evtimov, N.T.; Ghenev, P.; Dikranian, K. Trackins (Trk-Targeting Drugs): A Novel Therapy for Different Diseases. Pharmaceuticals 2024, 17, 961. [Google Scholar] [CrossRef]
- Farcas, A.; Hindmarch, C.; Iftene, F. BDNF gene Val66Met polymorphisms as a predictor for clinical presentation in schizophrenia—Recent findings. Front. Psychiatry 2023, 14, 1234220. [Google Scholar] [CrossRef]
- Dee, G.; Ryznar, R.; Dee, C. Epigenetic Changes Associated with Different Types of Stressors and Suicide. Cells 2023, 12, 1258. [Google Scholar] [CrossRef]
- Martínez-Dubarbie, F.; Guerra-Ruiz, A.; López-García, S.; Lage, C.; Fernández-Matarrubia, M.; Infante, J.; Pozueta-Cantudo, A.; García-Martínez, M.; Corrales-Pardo, A.; Bravo, M.; et al. Accuracy of plasma Aβ40, Aβ42, and p-tau181 to detect CSF Alzheimer’s pathological changes in cognitively unimpaired subjects using the Lumipulse automated platform. Alzheimer’s Res. Ther. 2023, 15, 163. [Google Scholar] [CrossRef]
- Hemdan, M.; Ali, M.A.; Doghish, A.S.; Mageed, S.S.A.; Elazab, I.M.; Khalil, M.M.; Mabrouk, M.; Das, D.B.; Amin, A.S. Innovations in Biosensor Technologies for Healthcare Diagnostics and Therapeutic Drug Monitoring: Applications, Recent Progress, and Future Research Challenges. Sensors 2024, 24, 5143. [Google Scholar] [CrossRef]
- Russo, L.; Giacomelli, C.; Fortino, M.; Marzo, T.; Ferri, G.; Calvello, M.; Viegi, A.; Magrì, A.; Pratesi, A.; Pietropaolo, A.; et al. Neurotrophic Activity and Its Modulation by Zinc Ion of a Dimeric Peptide Mimicking the Brain-Derived Neurotrophic Factor N-Terminal Region. ACS Chem. Neurosci. 2022, 13, 3453–3463. [Google Scholar] [CrossRef]
- Li, T.; Li, S.; Kang, Y.; Zhou, J.; Yi, M. Harnessing the evolving CRISPR/Cas9 for precision oncology. J. Transl. Med. 2024, 22, 749. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.; Breuillaud, L.; Yao, W.; Yin, T.; Norris, K.A.; Zehntner, S.P.; D’Adamio, L. TNF-α–mediated reduction in inhibitory neurotransmission precedes sporadic Alzheimer’s disease pathology in young Trem2R47H rats. J. Biol. Chem. 2021, 296, 100089. [Google Scholar] [CrossRef]
- Khalil, M.H. The BDNF-Interactive Model for Sustainable Hippocampal Neurogenesis in Humans: Synergistic Effects of Environmentally-Mediated Physical Activity, Cognitive Stimulation, and Mindfulness. Int. J. Mol. Sci. 2024, 25, 12924. [Google Scholar] [CrossRef]
- Arafah, A.; Khatoon, S.; Rasool, I.; Khan, A.; Rather, M.A.; Abujabal, K.A.; Faqih, Y.A.H.; Rashid, H.; Rashid, S.M.; Bilal Ahmad, S.; et al. The Future of Precision Medicine in the Cure of Alzheimer’s Disease. Biomedicines 2023, 11, 335. [Google Scholar] [CrossRef]
- Jovičić, S.M. Analysis of total RNA as a potential biomarker of developmental neurotoxicity in silico. Health Inform. J. 2024, 30, 14604582241285832. [Google Scholar] [CrossRef] [PubMed]
- Capelletti, S.; García Soto, S.C.; Gonçalves, M.A.F.V. On RNA-programmable gene modulation as a versatile set of principles targeting muscular dystrophies. Mol. Ther. 2024, 32, 3793–3807. [Google Scholar] [CrossRef]
- Cousins, K.A.Q.; Phillips, J.S.; Das, S.R.; O’Brien, K.; Tropea, T.F.; Chen-Plotkin, A.; Shaw, L.M.; Nasrallah, I.M.; Mechanic-Hamilton, D.; McMillan, C.T.; et al. Pathologic and cognitive correlates of plasma biomarkers in neurodegenerative disease. Alzheimer’s Dement. 2024, 20, 3889–3905. [Google Scholar] [CrossRef]
- Mohammadian Farsani, A.; Mokhtari, N.; Nooraei, S.; Bahrulolum, H.; Akbari, A.; Farsani, Z.M.; Khatami, S.; sadat Ebadi, M.; Ahmadian, G. Lipid nanoparticles: The game-changer in CRISPR-Cas9 genome editing. Heliyon 2024, 10, e24606. [Google Scholar] [CrossRef]
- Liu, J.J.; Borsari, B.; Li, Y.; Liu, S.X.; Gao, Y.; Xin, X.; Lou, S.; Jensen, M.; Garrido-Martín, D.; Verplaetse, T.L.; et al. Digital phenotyping from wearables using AI characterizes psychiatric disorders and identifies genetic associations. Cell 2025, 188, 515–529.e15. [Google Scholar] [CrossRef]
- Vo, D.-K.; Trinh, K.T.L. Advances in Wearable Biosensors for Healthcare: Current Trends, Applications, and Future Perspectives. Biosensors 2024, 14, 560. [Google Scholar] [CrossRef]
- Faraji, N.; Ebadpour, N.; Abavisani, M.; Gorji, A. Unlocking Hope: Therapeutic Advances and Approaches in Modulating the Wnt Pathway for Neurodegenerative Diseases. Mol. Neurobiol. 2025, 62, 3630–3652. [Google Scholar] [CrossRef] [PubMed]
- Keller, L.-A.; Merkel, O.; Popp, A. Intranasal drug delivery: Opportunities and toxicologic challenges during drug development. Drug Deliv. Transl. Res. 2022, 12, 735–757. [Google Scholar] [CrossRef]
- Greco, R.; Francavilla, M.; Facchetti, S.; Demartini, C.; Zanaboni, A.M.; Antonangeli, M.I.; Maffei, M.; Cattani, F.; Aramini, A.; Allegretti, M.; et al. Intranasal administration of recombinant human BDNF as a potential therapy for some primary headaches. J. Headache Pain 2024, 25, 184. [Google Scholar] [CrossRef] [PubMed]
- Benatti, B.M.; Adiletta, A.; Sgadò, P.; Malgaroli, A.; Ferro, M.; Lamanna, J. Epigenetic Modifications and Neuroplasticity in the Pathogenesis of Depression: A Focus on Early Life Stress. Behav. Sci. 2024, 14, 882. [Google Scholar] [CrossRef]
- Chen, T.-S.; Huang, T.-H.; Lai, M.-C.; Huang, C.-W. The Role of Glutamate Receptors in Epilepsy. Biomedicines 2023, 11, 783. [Google Scholar] [CrossRef]
- Pettorruso, M.; Miuli, A.; Clemente, K.; Mancusi, G.; Migliara, G.; Di Carlo, F.; Pernaci, G.; Di Crosta, T.; Santorelli, M.; d’Andrea, G.; et al. Enhanced peripheral levels of BDNF and proBDNF: Elucidating neurotrophin dynamics in cocaine use disorder. Mol. Psychiatry 2024, 29, 760–766. [Google Scholar] [CrossRef] [PubMed]
- Gliwińska, A.; Czubilińska-Łada, J.; Więckiewicz, G.; Świętochowska, E.; Badeński, A.; Dworak, M.; Szczepańska, M. The Role of Brain-Derived Neurotrophic Factor (BDNF) in Diagnosis and Treatment of Epilepsy, Depression, Schizophrenia, Anorexia Nervosa and Alzheimer’s Disease as Highly Drug-Resistant Diseases: A Narrative Review. Brain Sci. 2023, 13, 163. [Google Scholar] [CrossRef]
- Zakowicz, P.; Narozna, B.; Kozlowski, T.; Bargiel, W.; Grabarczyk, M.; Terczynska, M.; Pilecka, J.; Wasicka-Przewozna, K.; Pawlak, J.; Skibinska, M. Peripheral Brain-Derived Neurotrophic Factor (BDNF) and Its Regulatory miRNAs as Biological Correlates of Impulsivity in Young Adults. Metabolites 2024, 14, 529. [Google Scholar] [CrossRef]
- Cui, X.; Li, X.; Zheng, H.; Su, Y.; Zhang, S.; Li, M.; Hao, X.; Zhang, S.; Hu, Z.; Xia, Z.; et al. Human midbrain organoids: A powerful tool for advanced Parkinson’s disease modeling and therapy exploration. npj Park. Dis. 2024, 10, 189. [Google Scholar] [CrossRef]
- Haghighi, E.; Abolmaali, S.S.; Dehshahri, A.; Mousavi Shaegh, S.A.; Azarpira, N.; Tamaddon, A.M. Navigating the intricate in-vivo journey of lipid nanoparticles tailored for the targeted delivery of RNA therapeutics: A quality-by-design approach. J. Nanobiotechnol. 2024, 22, 710. [Google Scholar] [CrossRef]
- Wang, J.-H.; Gessler, D.J.; Zhan, W.; Gallagher, T.L.; Gao, G. Adeno-associated virus as a delivery vector for gene therapy of human diseases. Signal Transduct. Target. Ther. 2024, 9, 78. [Google Scholar] [CrossRef] [PubMed]
- Parambi, D.G.T.; Alharbi, K.S.; Kumar, R.; Harilal, S.; Batiha, G.E.-S.; Cruz-Martins, N.; Magdy, O.; Musa, A.; Panda, D.S.; Mathew, B. Gene Therapy Approach with an Emphasis on Growth Factors: Theoretical and Clinical Outcomes in Neurodegenerative Diseases. Mol. Neurobiol. 2022, 59, 191–233. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Pacheco, H.; Zepeda, R.C.; Picazo, O.; Quirarte, G.L.; Roldán-Roldán, G. Class I histone deacetylases inhibition reverses memory impairment induced by acute stress in mice. PLoS ONE 2024, 19, e0302374. [Google Scholar] [CrossRef]
- Mobed, A.; Charsouei, S.; Yazdani, Y.; Gargari, M.K.; Ahmadalipour, A.; Sadremousavi, S.R.; Farrahizadeh, M.; Shahbazi, A.; Haghani, M. Biosensors, Recent Advances in Determination of BDNF and NfL. Cell. Mol. Neurobiol. 2023, 43, 3801–3814. [Google Scholar] [CrossRef] [PubMed]
- Horschitz, S.; Frank, J.; Koch, P. Generation of three induced pluripotent stem cell lines carrying different variants of the BDNF (Val66Met) polymorphism. Stem Cell Res. 2023, 66, 102989. [Google Scholar] [CrossRef]
- Liberona, A.; Jones, N.; Zúñiga, K.; Garrido, V.; Zelada, M.I.; Silva, H.; Nieto, R.R. Brain-Derived Neurotrophic Factor (BDNF) as a Predictor of Treatment Response in Schizophrenia and Bipolar Disorder: A Systematic Review. Int. J. Mol. Sci. 2024, 25, 11204. [Google Scholar] [CrossRef]
- Saylam, B.; İncel, Ö.D. Quantifying Digital Biomarkers for Well-Being: Stress, Anxiety, Positive and Negative Affect via Wearable Devices and Their Time-Based Predictions. Sensors 2023, 23, 8987. [Google Scholar] [CrossRef]

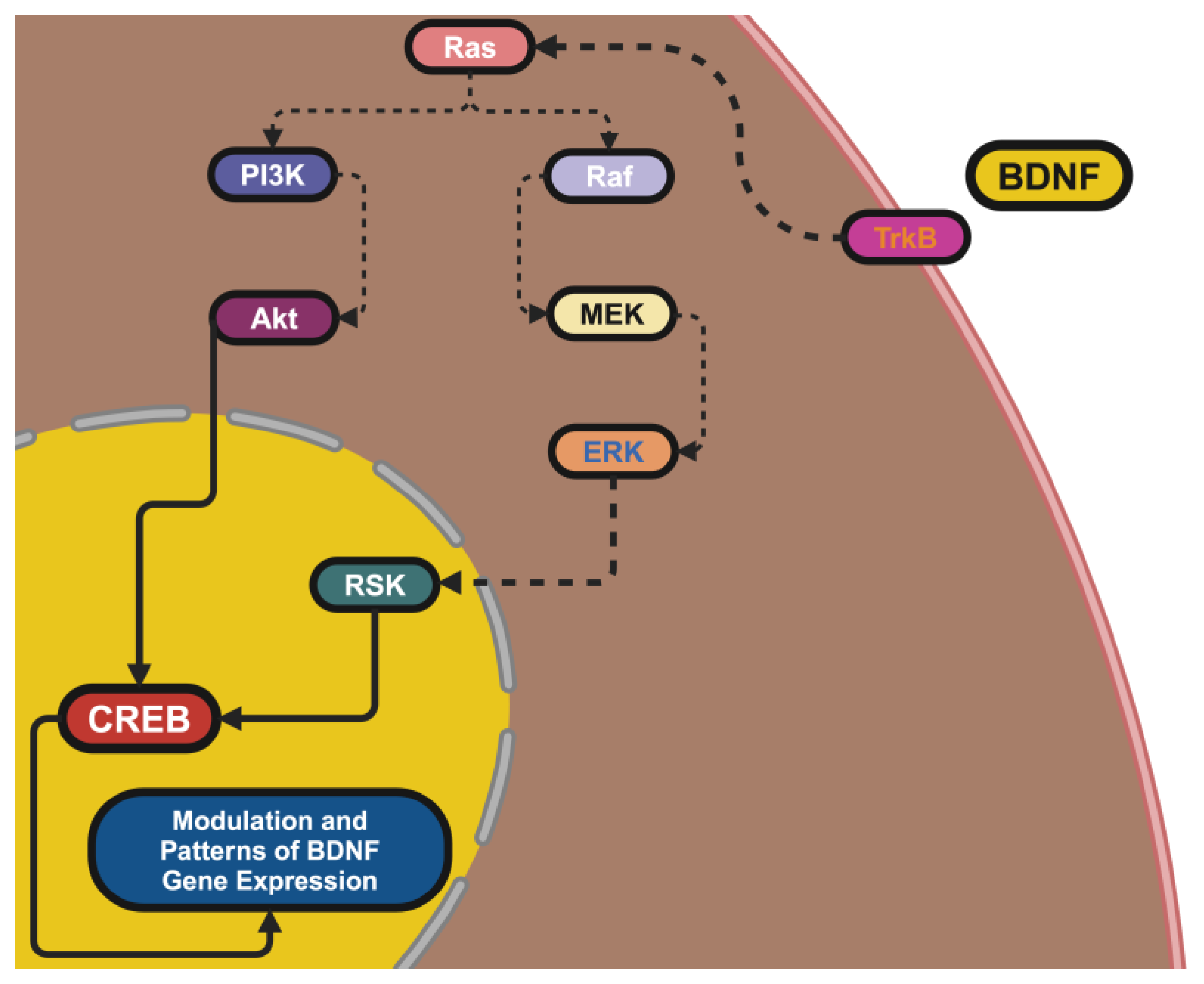
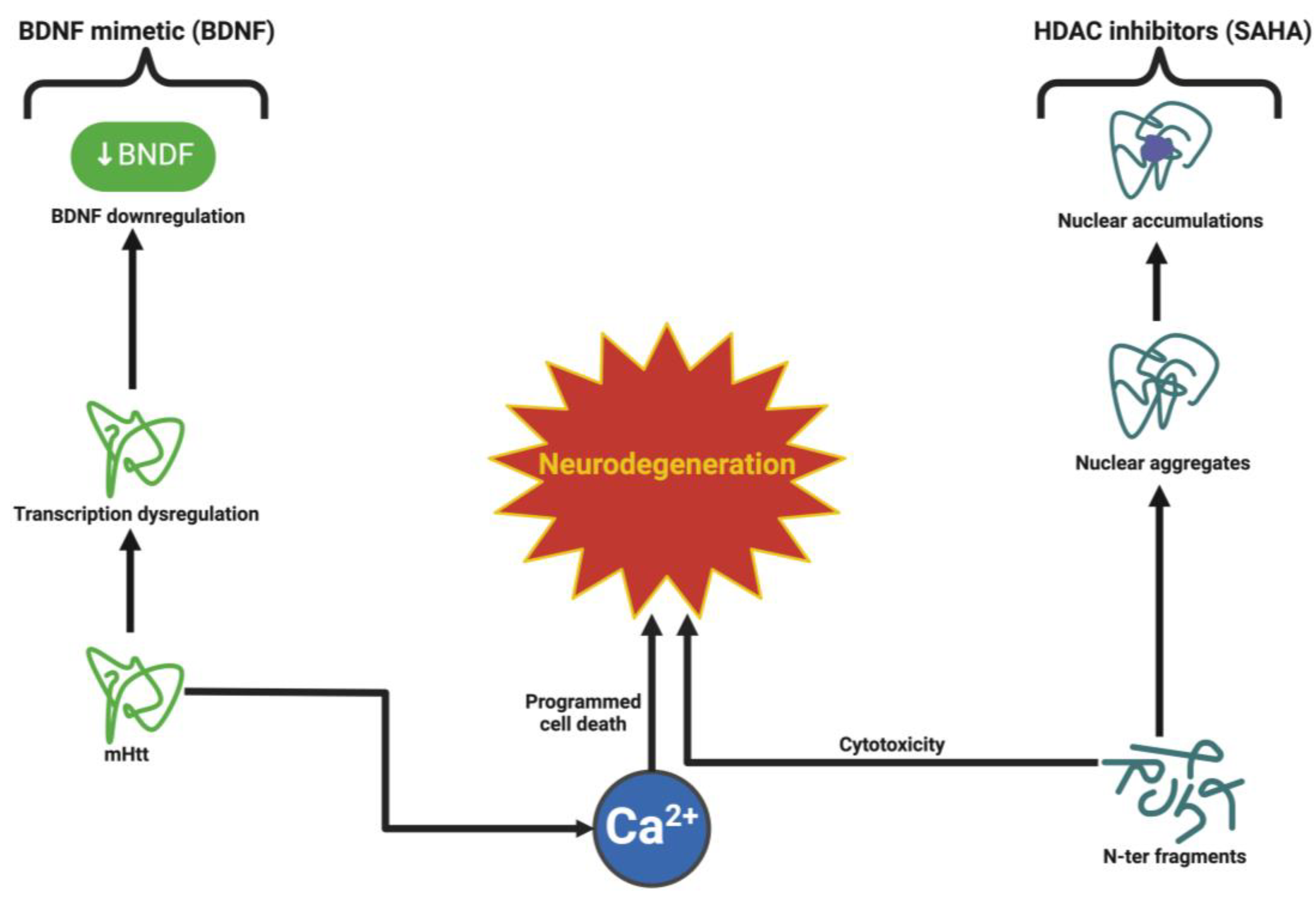

{kind=link}
{kind=link}
{kind=link}
| Research Area | Study Goal | Methodology | Major Findings | Impact and Implications | Citation |
|---|---|---|---|---|---|
| BDNF as an Early Alzheimer’s Biomarker | Evaluate plasma vs. CSF BDNF levels for early detection. | Longitudinal human study with biomarker tracking. | Plasma BDNF declines 10 years before clinical AD onset, correlating with hippocampal atrophy and CSF levels (R2 = 0.82). | Supports plasma BDNF as a non-invasive predictor, improving early diagnosis. | [21] |
| Exercise-Induced BDNF Transport Across the BBB | Determine how peripheral BDNF reaches the brain post-exercise. | Rodent treadmill study with BBB permeability assays. | LRP-1 receptor mediates BDNF transport, enhancing hippocampal plasticity and memory by 40%. | Establishes exercise as a mechanism to enhance central BDNF levels. | [22] |
| Stress, Epigenetics, and BDNF Regulation | Investigate the effects of chronic stress on BDNF gene expression. | Human cohort with DNA methylation analysis. | Hypermethylation of BDNF promoter IV correlates with hippocampal shrinkage; demethylation reverses resilience deficits. | Highlights epigenetic interventions as potential psychiatric treatments. | [23] |
| Gut–Brain Axis and BDNF in Inflammation | Assess how gut inflammation affects neuroplasticity. | Germ-free mouse model with induced colitis. | Colonic inflammation reduces systemic BDNF, impairing neurogenesis and cognition; probiotics restore function. | Suggests gut microbiota modulation as a neuroprotective strategy. | [24] |
| BDNF and Cardiomyocyte Survival Under Stress | Explore BDNF’s role in cardiac adaptation to hypoxia. | Cardiomyocyte cultures and ischemic rodent models. | Hypoxia triggers BDNF upregulation, promoting mitochondrial protection and cell survival via TrkB-PI3K activation. | Expands BDNF’s role beyond the CNS, identifying cardioprotective mechanisms. | [25] |
| BDNF in Vascular Repair During Systemic Inflammation | Examine its function in endothelial regeneration. | Preclinical sepsis model with vascular integrity assays. | BDNF enhances VEGF expression, improving endothelial stability and reducing inflammation-induced leakage. | Positions BDNF as a vascular repair mediator in inflammatory conditions. | [26] |
| Sex Differences in BDNF and Neuroprotection | Identify gender-based variations in BDNF dynamics. | Human cohort. | Women maintain higher BDNF levels, linked to lower neurodegeneration risk; estrogen enhances TrkB signaling. | Emphasizes sex-specific approaches in neurodegenerative disease prevention. | [27] |
| Differential Roles of proBDNF and mBDNF in Neurodegeneration | Analyze distinct effects of BDNF isoforms on disease progression. | Mass spectrometry of CSF samples in AD patients. | ProBDNF elevation signals early neurodegeneration, driving synaptic loss and inflammation. | Establishes proBDNF as a biomarker for early-stage neurodegenerative decline. | [28] |
| BDNF and Sensory Neuron Function in Diabetes | Assess BDNF’s role in diabetic neuropathy. | Patient cohort study with sensory function assessments. | Low systemic BDNF correlates with increased sensory deficits and impaired pain response. | Highlights BDNF’s therapeutic potential for neuropathy management. | [29] |
| BDNF and Tumor-Associated Neural Plasticity | Investigate its role in neural adaptations in cancer. | Tumor-derived nerve culture models and clinical biopsy analysis. | Cancer-secreted factors upregulate BDNF, promoting tumor-related axonal sprouting. | Identifies BDNF as a potential therapeutic target in neuro-oncology. | [30] |
| Therapeutic Approach | Target Condition | Key Outcome | Novel Contributions | Reference |
|---|---|---|---|---|
| LNP Delivery of BDNF mRNA | Alzheimer’s Disease | ↑ Synaptic density (200%), reversal of memory deficits in 4 weeks | First non-invasive BDNF mRNA therapy, overcoming BBB and degradation issues | [131] |
| CRISPR-Based BDNF Gene Activation | Parkinson’s Disease | 70% dopaminergic neuron survival, 50% motor improvement | First CRISPR-dCas9 approach for long-term neuroprotection without direct BDNF protein delivery | [132] |
| Aerobic Exercise and Serum BDNF | Cognitive Aging | ↑ Serum BDNF (30%), ↑ Executive function (35%) | Direct evidence of exercise-driven BDNF elevation improving cognitive resilience | [133] |
| Epigenetic Modulation via HDAC Inhibitors | Depression | ↑ Hippocampal BDNF (60%), reversal of behavioral deficits | Established epigenetic repression of BDNF as a drug target for depression | [134] |
| Cyclic TrkB Agonists | ALS | ↓ Motor neuron loss (50%), ↑ Motor function (40%), ↑ Survival (25%) | Developed BBB-penetrant TrkB agonists for motor neuron preservation | [135] |
| Plasma BDNF as a Biomarker | Preclinical AD. | BDNF decline 10 years pre-symptom onset, correlated with hippocampal atrophy (R2 = 0.82) | Identified plasma BDNF as a non-invasive early biomarker for AD | [136] |
| Viral BDNF Delivery in the Amygdala | PTSD | ↑ Fear extinction (45%), ↓ Hypervigilance | Demonstrated region-specific BDNF upregulation for fear extinction therapy | [137] |
| BDNF mRNA Therapy | Huntington’s Disease | ↑ Spine density (50%), ↑ Motor function | First mRNA-based BDNF restoration for corticostriatal connectivity | [138] |
| Optogenetic Stimulation of BDNF | Cortical Plasticity | ↑ Activity-dependent BDNF, ↑ Decision-making accuracy (30%) | First optogenetic tool for precise BDNF modulation in brain networks | [139] |
| Flavonoid-Induced BDNF Enhancement | Cognitive Aging | ↑ Plasma BDNF (25%), ↓ Cognitive decline (35%) | Established dietary polyphenols as natural BDNF modulators | [140] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toader, C.; Serban, M.; Munteanu, O.; Covache-Busuioc, R.-A.; Enyedi, M.; Ciurea, A.V.; Tataru, C.P. From Synaptic Plasticity to Neurodegeneration: BDNF as a Transformative Target in Medicine. Int. J. Mol. Sci. 2025, 26, 4271. https://doi.org/10.3390/ijms26094271
Toader C, Serban M, Munteanu O, Covache-Busuioc R-A, Enyedi M, Ciurea AV, Tataru CP. From Synaptic Plasticity to Neurodegeneration: BDNF as a Transformative Target in Medicine. International Journal of Molecular Sciences. 2025; 26(9):4271. https://doi.org/10.3390/ijms26094271
Chicago/Turabian StyleToader, Corneliu, Matei Serban, Octavian Munteanu, Razvan-Adrian Covache-Busuioc, Mihaly Enyedi, Alexandru Vlad Ciurea, and Calin Petru Tataru. 2025. "From Synaptic Plasticity to Neurodegeneration: BDNF as a Transformative Target in Medicine" International Journal of Molecular Sciences 26, no. 9: 4271. https://doi.org/10.3390/ijms26094271
APA StyleToader, C., Serban, M., Munteanu, O., Covache-Busuioc, R.-A., Enyedi, M., Ciurea, A. V., & Tataru, C. P. (2025). From Synaptic Plasticity to Neurodegeneration: BDNF as a Transformative Target in Medicine. International Journal of Molecular Sciences, 26(9), 4271. https://doi.org/10.3390/ijms26094271