
Acetylcholine Sustains LNCaP Prostate Cancer Cell Migration, Invasion and Proliferation Through Glyoxalase 1/MG-H1 Axis with the Involvement of Osteopontin


 ,
,  ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
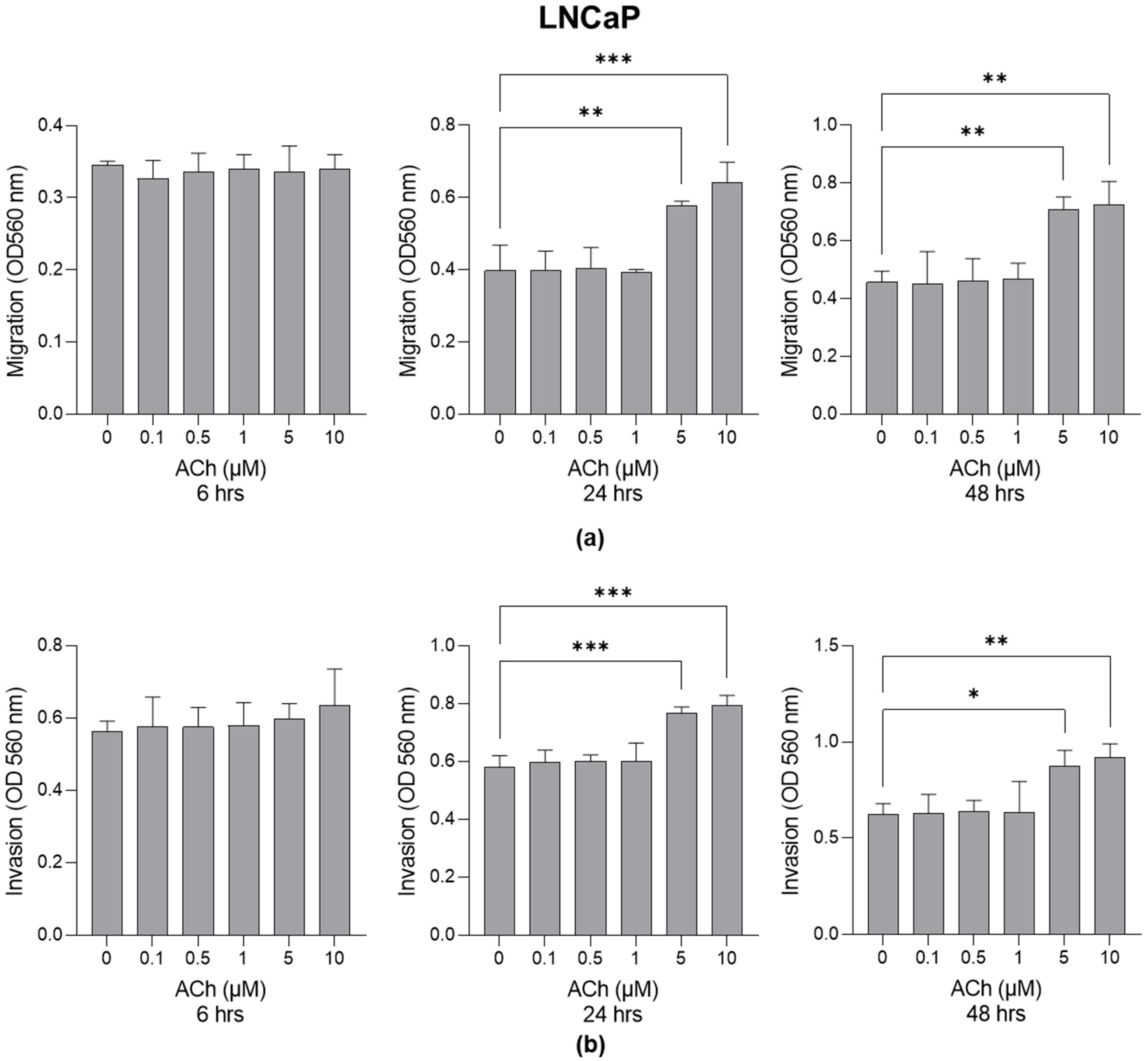
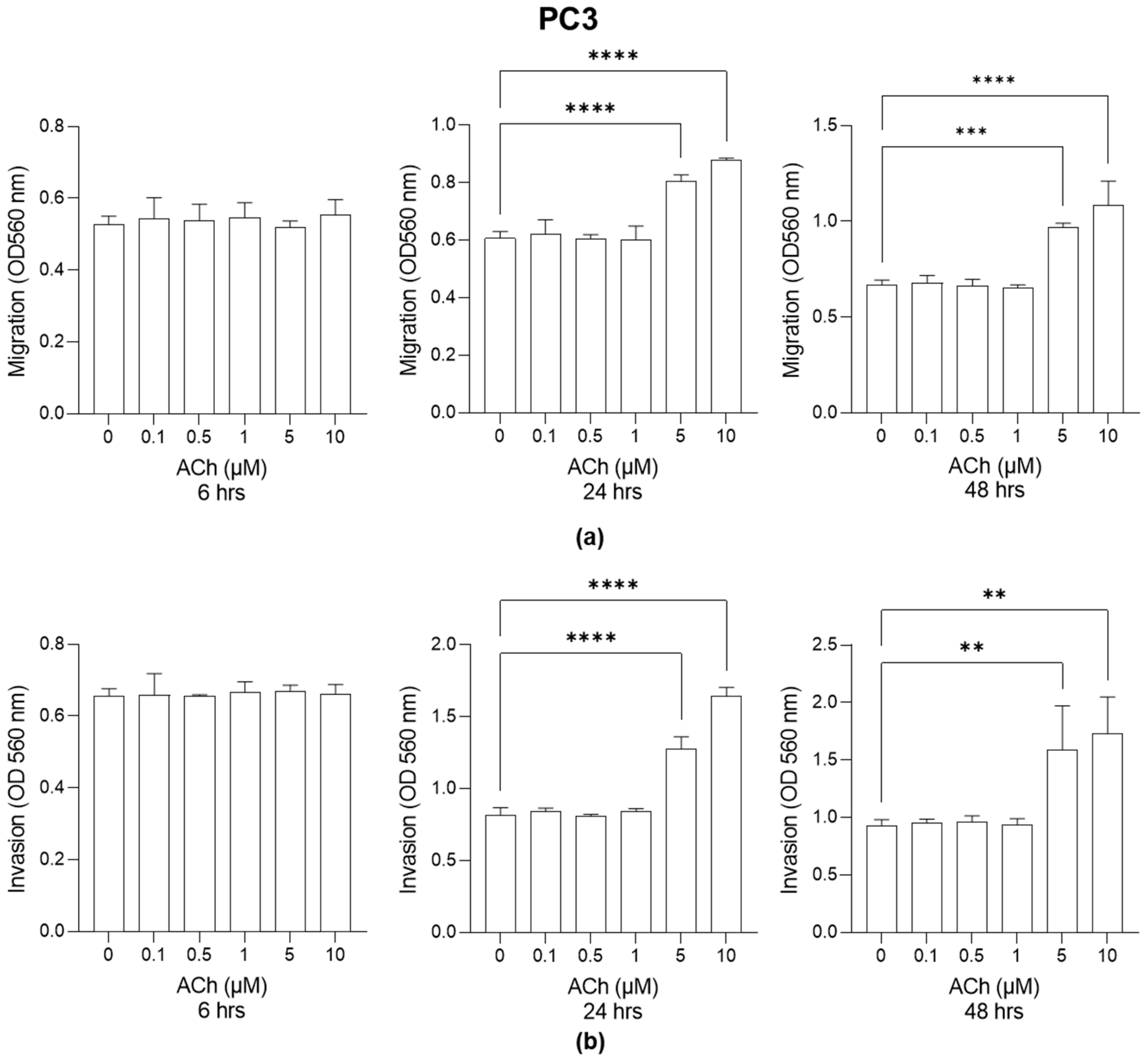
2.1. Effect of ACh on LNCaP and PC3 Cell Migration and Invasion
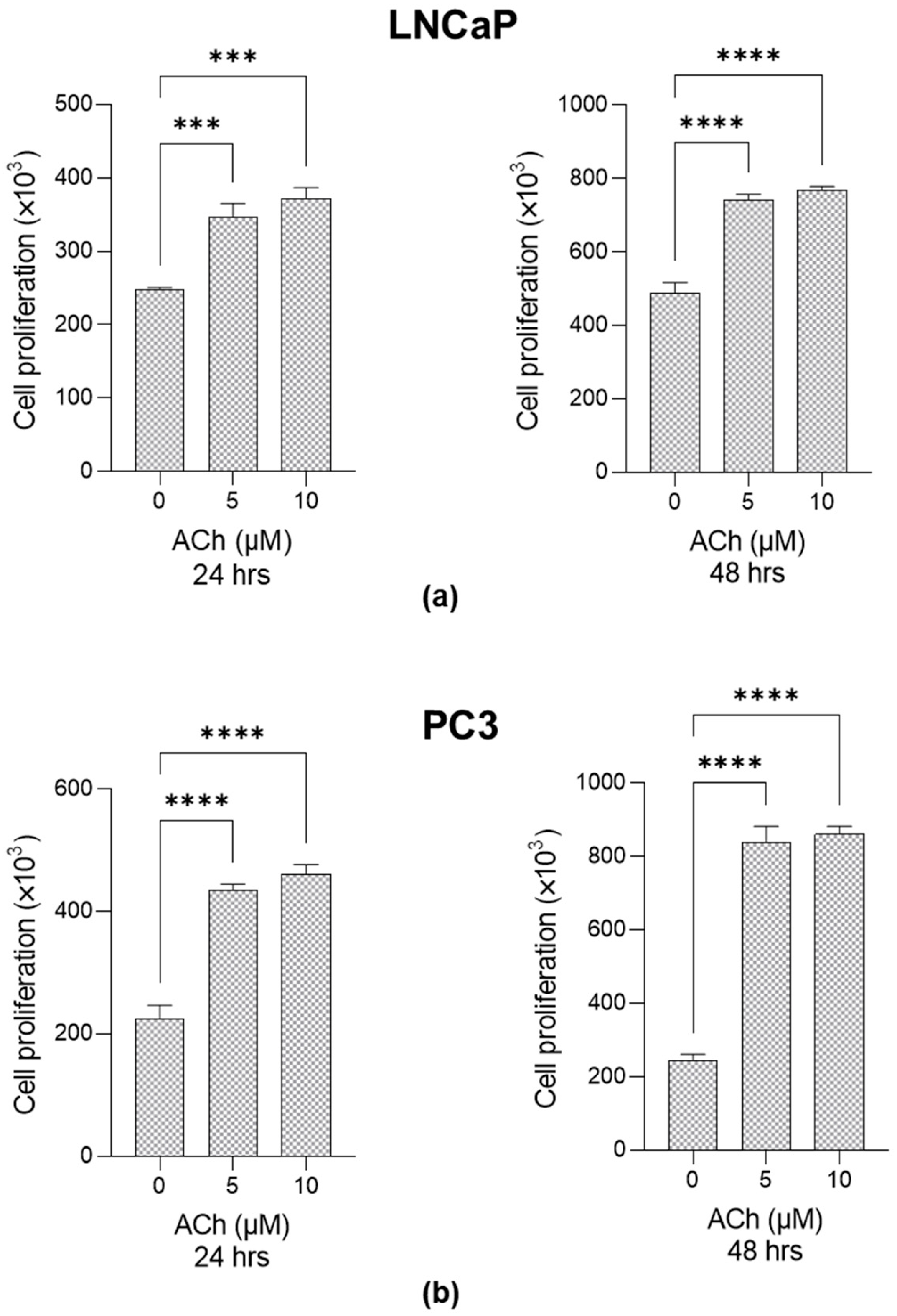
2.2. Effect of ACh on LNCaP and PC3 Cell Proliferation
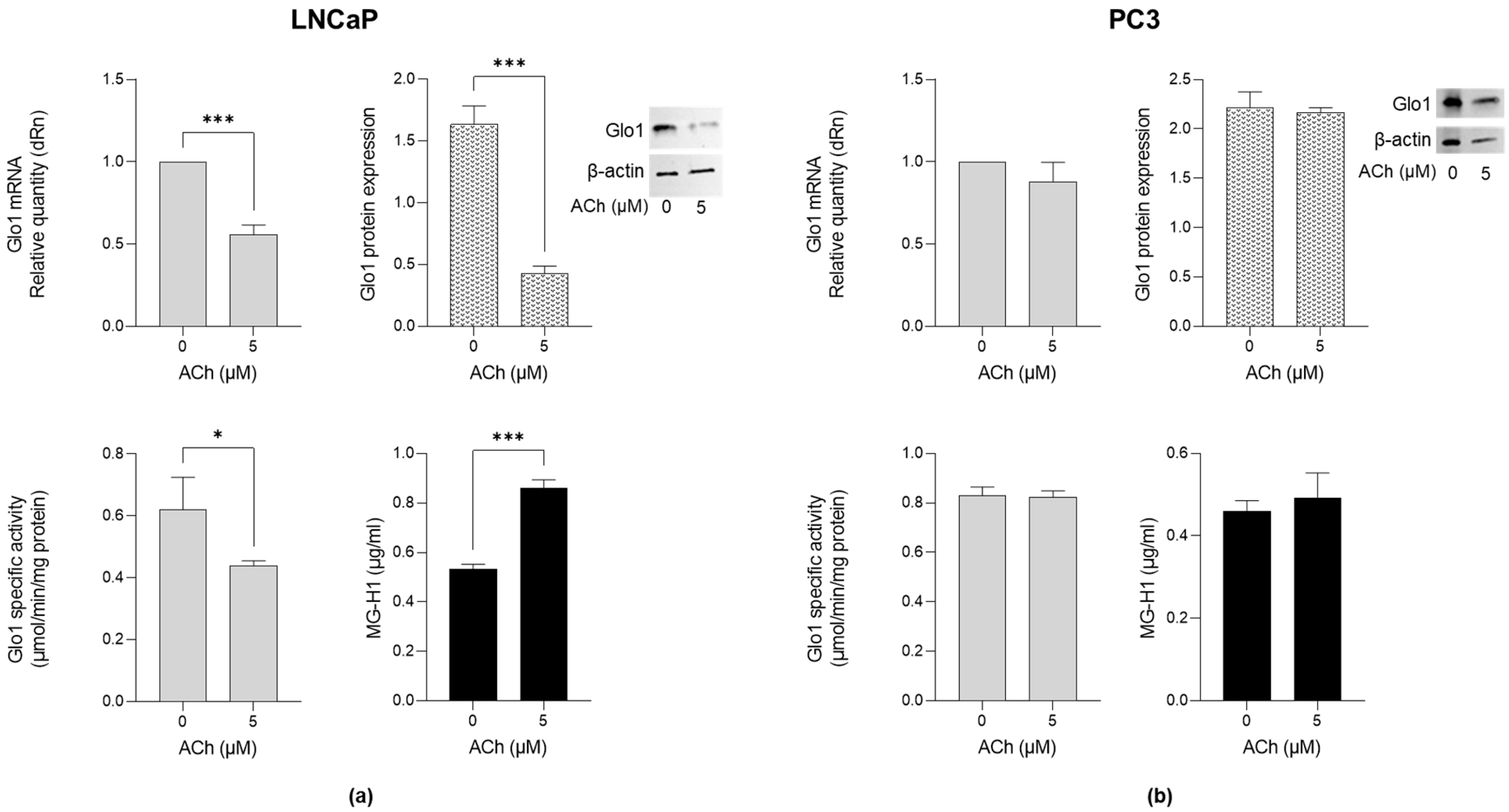
2.3. Effect of ACh on Glo1 Expression and MG-H1 Levels in LNCaP and PC3 Cells
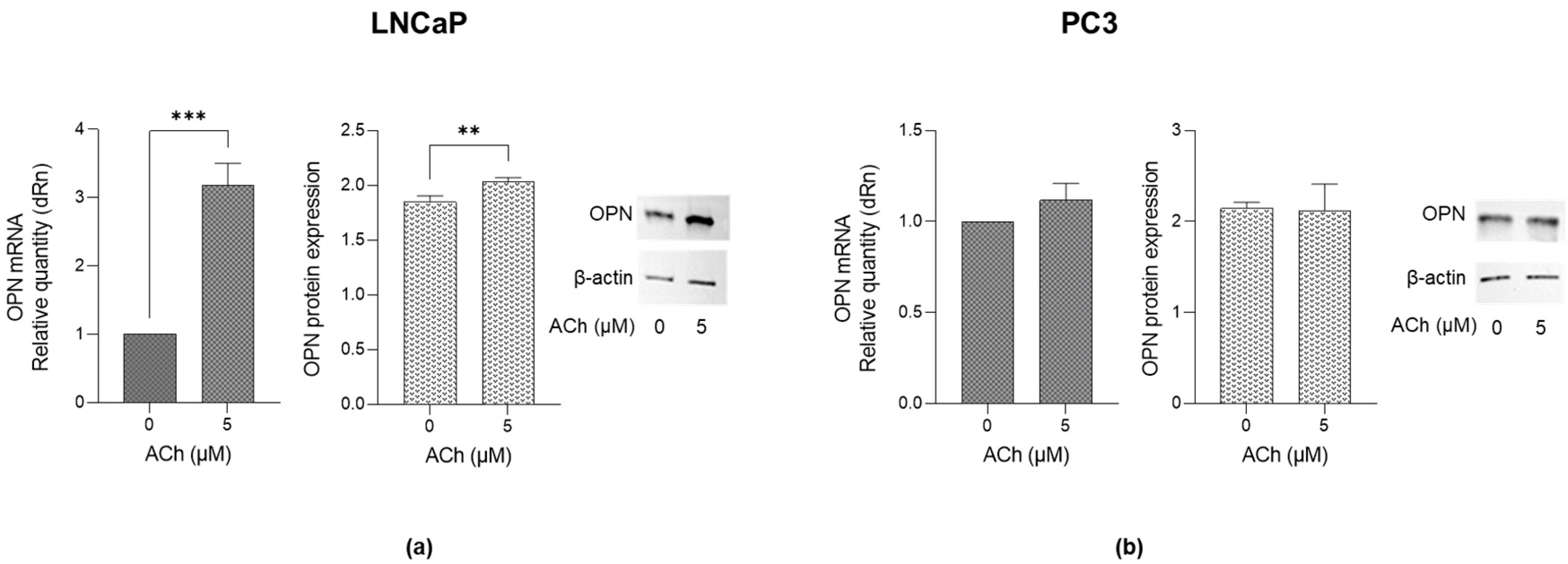
2.4. Effects of ACh on Osteopontin (OPN) Expression in LNCaP and PC3 Cells
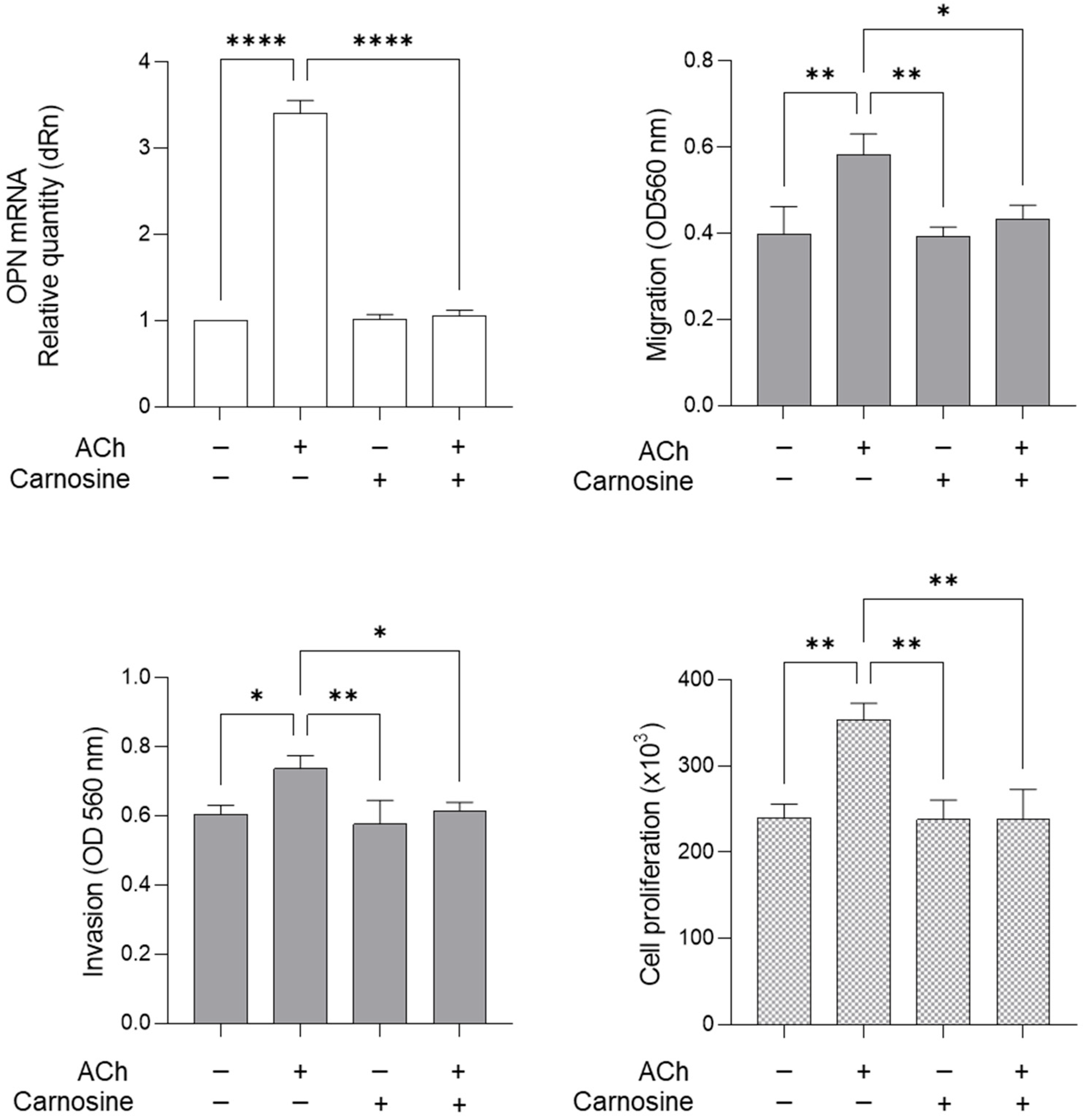
2.5. ACh Sustains LNCaP Migration, Invasion and Proliferation Through Glo1-Dependent MG-H1-Mediated Osteopontin Upregulation
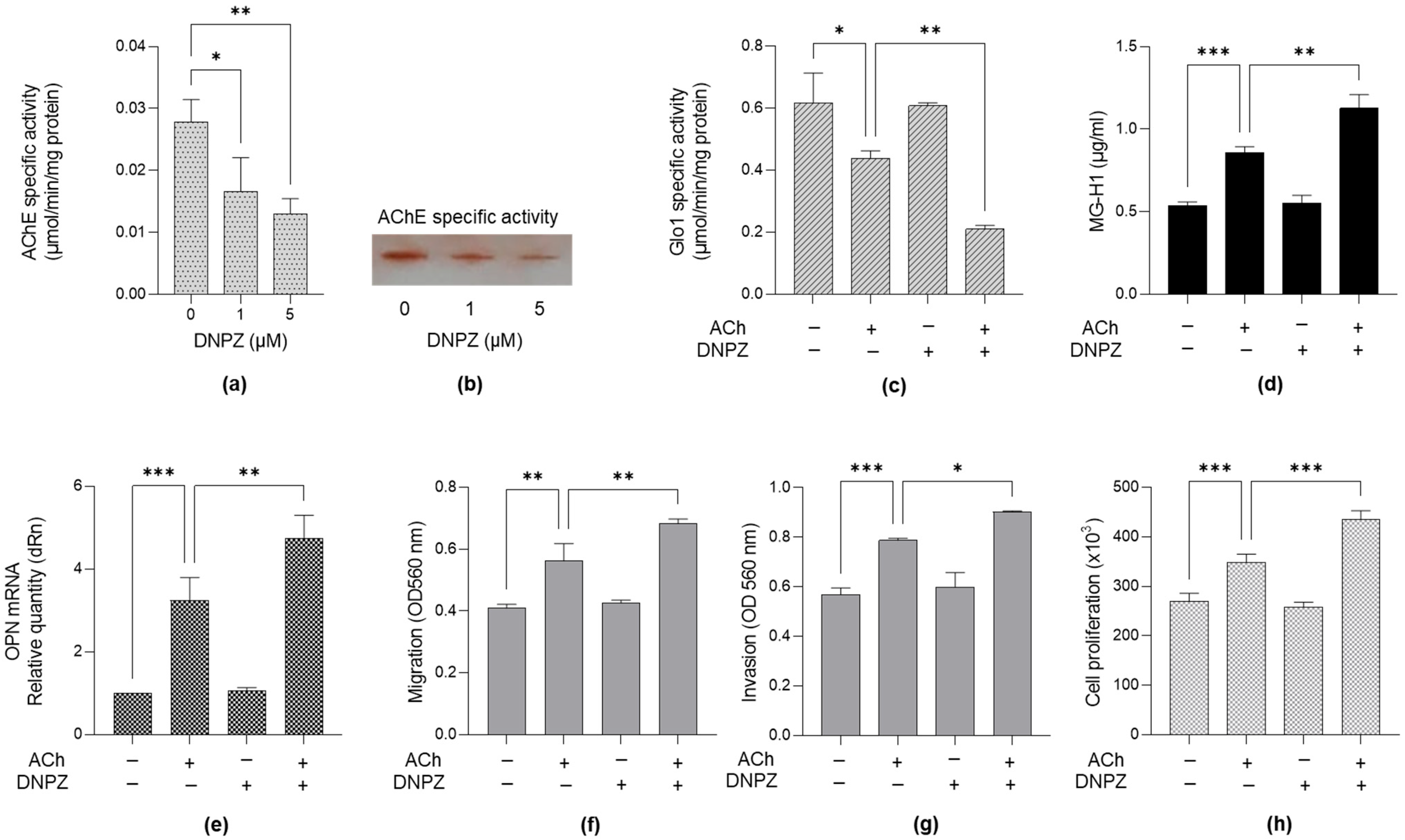
2.6. Pre-Treatment of LNCaP PCa Cells with Donepezil, an Acetylcholinesterase (AChE) Inhibitor, and ACh Further Supports the Involvement of the Glo1/MG-H1 Axis and Osteopontin in the Pro-Tumorigenic Effect Driven by ACh Through Cell Migration, Invasion and Proliferation Control
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Models and Culture Conditions
4.3. RNA Isolation, Reverse Transcription and Real-Time Reverse Transcriptase-Polymerase Chain Reaction (RT-PCR)
4.4. Cell Lysates
4.5. Glo1 Enzyme Activity
4.6. Western Blot
4.7. MG-H1 Detection
4.8. Migration and Invasion
4.9. Cell Proliferation
4.10. ACh Levels
4.11. AChE Enzyme Activity
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Park, H.; Lee, C.H. The Contribution of the Nervous System in the Cancer Progression. BMB Rep. 2024, 57, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Tibensky, M.; Mravec, B. Role of the Parasympathetic Nervous System in Cancer Initiation and Progression. Clin. Transl. Oncol. 2021, 23, 669–681. [Google Scholar] [CrossRef] [PubMed]
- Aronowitz, A.L.; Ali, S.R.; Glaun, M.D.E.; Amit, M. Acetylcholine in Carcinogenesis and Targeting Cholinergic Receptors in Oncology. Adv. Biol. 2022, 6, 2200053. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Yao, M.; Xu, J.; Quan, Y.; Zhang, K.; Yang, R.; Gao, W.-Q. Autocrine Activation of CHRM3 Promotes Prostate Cancer Growth and Castration Resistance via CaM/CaMKK-Mediated Phosphorylation of Akt. Clin. Cancer Res. 2015, 21, 4676–4685. [Google Scholar] [CrossRef]
- Yin, Q.-Q.; Xu, L.-H.; Zhang, M.; Xu, C. Muscarinic Acetylcholine Receptor M1 Mediates Prostate Cancer Cell Migration and Invasion through Hedgehog Signaling. Asian J. Androl. 2018, 20, 608. [Google Scholar] [CrossRef]
- Qi, J.; Xue, W.; Zhang, Y.; Qu, C.; Lu, B.; Yin, Y.; Liu, K.; Wang, D.; Li, W.; Zhao, Z. Cholinergic A5 Nicotinic Receptor Is Involved in the Proliferation and Invasion of Human Prostate Cancer Cells. Oncol. Rep. 2020, 43, 159–168. [Google Scholar] [CrossRef]
- Antognelli, C.; Talesa, V. Glyoxalases in Urological Malignancies. Int. J. Mol. Sci. 2018, 19, 415. [Google Scholar] [CrossRef]
- Antognelli, C.; Cecchetti, R.; Riuzzi, F.; Peirce, M.J.; Talesa, V.N. Glyoxalase 1 Sustains the Metastatic Phenotype of Prostate Cancer Cells via EMT Control. J. Cell. Mol. Med. 2018, 22, 2865–2883. [Google Scholar] [CrossRef]
- Antognelli, C.; Marinucci, L.; Frosini, R.; Macchioni, L.; Talesa, V.N. Metastatic Prostate Cancer Cells Secrete Methylglyoxal-Derived MG-H1 to Reprogram Human Osteoblasts into a Dedifferentiated, Malignant-like Phenotype: A Possible Novel Player in Prostate Cancer Bone Metastases. Int. J. Mol. Sci. 2021, 22, 10191. [Google Scholar] [CrossRef]
- Sullivan, J.; Blair, L.; Alnajar, A.; Aziz, T.; Chipitsyna, G.; Gong, Q.; Yeo, C.J.; Arafat, H.A. Expression and Regulation of Nicotine Receptor and Osteopontin Isoforms in Human Pancreatic Ductal Adenocarcinoma. Histol. Histopathol. 2011, 26, 893–904. [Google Scholar] [CrossRef]
- Chipitsyna, G.; Gong, Q.; Anandanadesan, R.; Alnajar, A.; Batra, S.K.; Wittel, U.A.; Cullen, D.M.; Akhter, M.P.; Denhardt, D.T.; Yeo, C.J.; et al. Induction of Osteopontin Expression by Nicotine and Cigarette Smoke in the Pancreas and Pancreatic Ductal Adenocarcinoma Cells. Int. J. Cancer 2009, 125, 276–285. [Google Scholar] [CrossRef] [PubMed]
- Silver, S.V.; Popovics, P. The Multifaceted Role of Osteopontin in Prostate Pathologies. Biomedicines 2023, 11, 2895. [Google Scholar] [CrossRef] [PubMed]
- Lukianova, N.; Zadvornyi, T.; Borikun, T.; Mushii, O.; Pavlova, A.; Tymoshenko, A.; Stakhovskyi, E.; Vitruk, I.; Chekhun, V. Significance of osteopontin for predicting aggressiveness of prostate cancer. Exp. Oncol. 2023, 45, 312–321. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Huan, X.; Xiao, G.-H.; Yu, W.-H.; Li, T.-F.; Gao, X.-D.; Zhang, Y.-C. Osteopontin and Its Downstream Carcinogenic Molecules: Regulatory Mechanisms and Prognostic Value in Cancer Progression. Neoplasma 2022, 69, 1253–1269. [Google Scholar] [CrossRef]
- Butti, R.; Kumar, T.V.S.; Nimma, R.; Banerjee, P.; Kundu, I.G.; Kundu, G.C. Osteopontin Signaling in Shaping Tumor Microenvironment Conducive to Malignant Progression. In Tumor Microenvironment; Advances in Experimental Medicine and Biology; Birbrair, A., Ed.; Springer International Publishing: Cham, Switzerland, 2021; Volume 1329, pp. 419–441. ISBN 978-3-030-73118-2. [Google Scholar]
- Mourkioti, I.; Polyzou, A.; Veroutis, D.; Theocharous, G.; Lagopati, N.; Gentile, E.; Stravokefalou, V.; Thanos, D.-F.; Havaki, S.; Kletsas, D.; et al. A GATA2-CDC6 Axis Modulates Androgen Receptor Blockade-Induced Senescence in Prostate Cancer. J. Exp. Clin. Cancer Res. 2023, 42, 187. [Google Scholar] [CrossRef]
- Yamashita, S.-N.; Tanaka, Y.; Kitagawa, T.; Baron, B.; Tokuda, K.; Paudel, D.; Nakagawa, K.; Ohta, T.; Hamada, J.-I.; Kobayashi, M.; et al. Down-Regulating Effect of a Standardized Extract of Cultured Lentinula Edodes Mycelia on Cortactin in Prostate Cancer Cells Is Dependent on Malignant Potential. Anticancer Res. 2023, 43, 1159–1166. [Google Scholar] [CrossRef]
- Panda, V.K.; Mishra, B.; Nath, A.N.; Butti, R.; Yadav, A.S.; Malhotra, D.; Khanra, S.; Mahapatra, S.; Mishra, P.; Swain, B.; et al. Osteopontin: A Key Multifaceted Regulator in Tumor Progression and Immunomodulation. Biomedicines 2024, 12, 1527. [Google Scholar] [CrossRef]
- Wetzel, C.; Gallenstein, N.; Peters, V.; Fleming, T.; Marinovic, I.; Bodenschatz, A.; Du, Z.; Küper, K.; Dallanoce, C.; Aldini, G.; et al. Histidine Containing Dipeptides Protect Epithelial and Endothelial Cell Barriers from Methylglyoxal Induced Injury. Sci. Rep. 2024, 14, 26640. [Google Scholar] [CrossRef]
- Matthews, J.J.; Turner, M.D.; Santos, L.; Elliott-Sale, K.J.; Sale, C. Carnosine Increases Insulin-Stimulated Glucose Uptake and Reduces Methylglyoxal-Modified Proteins in Type-2 Diabetic Human Skeletal Muscle Cells. Amino Acids 2023, 55, 413–420. [Google Scholar] [CrossRef]
- Zheng, J.; Zhao, G.; Hu, Z.; Jia, C.; Li, W.; Peng, Y.; Zheng, J. Metabolic Activation and Cytotoxicity of Donepezil Induced by CYP3A4. Chem. Res. Toxicol. 2024, 37, 2003–2012. [Google Scholar] [CrossRef]
- Bagrowska, W.; Karasewicz, A.; Góra, A. Comprehensive Analysis of Acetylcholinesterase Inhibitor and Reactivator Complexes: Implications for Drug Design and Antidote Development. Drug Discov. Today 2024, 29, 104217. [Google Scholar] [CrossRef]
- Muñoz, J.P.; Calaf, G.M. Acetylcholine, Another Factor in Breast Cancer. Biology 2023, 12, 1418. [Google Scholar] [CrossRef]
- Oz, M.; King, J.R.; Yang, K.-H.S.; Khushaish, S.; Tchugunova, Y.; Khajah, M.A.; Luqmani, Y.A.; Kabbani, N. A7 Nicotinic Acetylcholine Receptor Interaction with G Proteins in Breast Cancer Cell Proliferation, Motility, and Calcium Signaling. PLoS ONE 2023, 18, e0289098. [Google Scholar] [CrossRef]
- Sampaio Moura, N.; Schledwitz, A.; Alizadeh, M.; Kodan, A.; Njei, L.-P.; Raufman, J.-P. Cholinergic Mechanisms in Gastrointestinal Neoplasia. Int. J. Mol. Sci. 2024, 25, 5316. [Google Scholar] [CrossRef]
- Pan, J.; Zhang, L.; Wang, X.; Li, L.; Yang, C.; Wang, Z.; Su, K.; Hu, X.; Zhang, Y.; Ren, G.; et al. Chronic Stress Induces Pulmonary Epithelial Cells to Produce Acetylcholine That Remodels Lung Pre-Metastatic Niche of Breast Cancer by Enhancing NETosis. J. Exp. Clin. Cancer Res. 2023, 42, 255. [Google Scholar] [CrossRef]
- Zou, H.; Chen, Y.; Zhu, X.; Zhao, X.; Cao, J.; Chen, Y.; Zhang, Z.; Zhu, Y.; Li, Q.; Li, M. Spinosad Blocks CHRNA5 Mediated EGFR Signaling Pathway Activation to Inhibit Lung Adenocarcinoma Proliferation. Biomed. Pharmacother. 2024, 177, 117105. [Google Scholar] [CrossRef]
- Kooshan, Z.; Srinivasan, S.; Janjua, T.I.; Popat, A.; Batra, J. Lactoferrin Conjugated Radicicol Nanoparticles Enhanced Drug Delivery and Cytotoxicity in Prostate Cancer Cells. Eur. J. Pharmacol. 2025, 991, 177300. [Google Scholar] [CrossRef]
- Cardoso, H.J.; Carvalho, T.M.A.; Fonseca, L.R.S.; Figueira, M.I.; Vaz, C.V.; Socorro, S. Revisiting Prostate Cancer Metabolism: From Metabolites to Disease and Therapy. Med. Res. Rev. 2021, 41, 1499–1538. [Google Scholar] [CrossRef]
- Higgins, L.H.; Withers, H.G.; Garbens, A.; Love, H.D.; Magnoni, L.; Hayward, S.W.; Moyes, C.D. Hypoxia and the Metabolic Phenotype of Prostate Cancer Cells. Biochim. Biophys. Acta—Bioenerg. 2009, 1787, 1433–1443. [Google Scholar] [CrossRef]
- Antognelli, C.; Mandarano, M.; Prosperi, E.; Sidoni, A.; Talesa, V.N. Glyoxalase-1-Dependent Methylglyoxal Depletion Sustains PD-L1 Expression in Metastatic Prostate Cancer Cells: A Novel Mechanism in Cancer Immunosurveillance Escape and a Potential Novel Target to Overcome PD-L1 Blockade Resistance. Cancers 2021, 13, 2965. [Google Scholar] [CrossRef]
- Angelucci, A.; Festuccia, C.; Gravina, G.L.; Muzi, P.; Bonghi, L.; Vicentini, C.; Bologna, M. Osteopontin Enhances the Cell Proliferation Induced by the Epidermal Growth Factor in Human Prostate Cancer Cells. Prostate 2004, 59, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Kundu, G.; Elangovan, S. Investigating the Role of Osteopontin (OPN) in the Progression of Breast, Prostate, Renal and Skin Cancers. Biomedicines 2025, 13, 173. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.; Yoo, H.J.; Yoo, M.K.; Choi, J.H.; Lee, K.-W. Methylglyoxal-Derived Hydroimidazolone-1/RAGE Axis Induces Renal Oxidative Stress and Renal Fibrosis In Vitro and In Vivo. Toxicology 2024, 507, 153887. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.L.; de Oliveira, M.G.; Mónica, F.Z.; Antunes, E. Methylglyoxal and Advanced Glycation End Products (AGEs): Targets for the Prevention and Treatment of Diabetes-Associated Bladder Dysfunction? Biomedicines 2024, 12, 939. [Google Scholar] [CrossRef]
- Lyle, A.N.; Remus, E.W.; Fan, A.E.; Lassègue, B.; Walter, G.A.; Kiyosue, A.; Griendling, K.K.; Taylor, W.R. Hydrogen Peroxide Regulates Osteopontin Expression through Activation of Transcriptional and Translational Pathways. J. Biol. Chem. 2014, 289, 275–285. [Google Scholar] [CrossRef]
- Wu, Q.; Li, L.; Miao, C.; Hasnat, M.; Sun, L.; Jiang, Z.; Zhang, L. Osteopontin Promotes Hepatocellular Carcinoma Progression through Inducing JAK2/STAT3/NOX1-Mediated ROS Production. Cell Death Dis. 2022, 13, 341. [Google Scholar] [CrossRef]
- Nokin, M.-J.; Bellier, J.; Durieux, F.; Peulen, O.; Rademaker, G.; Gabriel, M.; Monseur, C.; Charloteaux, B.; Verbeke, L.; Van Laere, S.; et al. Methylglyoxal, a Glycolysis Metabolite, Triggers Metastasis through MEK/ERK/SMAD1 Pathway Activation in Breast Cancer. Breast Cancer Res. 2019, 21, 11. [Google Scholar] [CrossRef]
- Antognelli, C.; Moretti, S.; Frosini, R.; Puxeddu, E.; Sidoni, A.; Talesa, V.N. Methylglyoxal Acts as a Tumor-Promoting Factor in Anaplastic Thyroid Cancer. Cells 2019, 8, 547. [Google Scholar] [CrossRef]
- Fuenzalida, M.; Pérez, M.Á.; Arias, H.R. Role of Nicotinic and Muscarinic Receptors on Synaptic Plasticity and Neurological Diseases. Curr. Pharm. Des. 2016, 22, 2004–2014. [Google Scholar] [CrossRef]
- Antognelli, C.; Mancuso, F.; Frosini, R.; Arato, I.; Calvitti, M.; Calafiore, R.; Talesa, V.N.; Luca, G. Testosterone and Follicle Stimulating Hormone–Dependent Glyoxalase 1 Up-Regulation Sustains the Viability of Porcine Sertoli Cells through the Control of Hydroimidazolone– and Argpyrimidine-Mediated NF-κB Pathway. Am. J. Pathol. 2018, 188, 2553–2563. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Ueda, K.; Zhao, J.; Kim, H.J.; Sparrow, J.R. Photodegradation of Retinal Bisretinoids in Mouse Models and Implications for Macular Degeneration. Proc. Natl. Acad. Sci. USA 2016, 113, 6904–6909. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Ueda, K.; Zhao, J.; Sparrow, J.R. Correlations between Photodegradation of Bisretinoid Constituents of Retina and Dicarbonyl Adduct Deposition. J. Biol. Chem. 2015, 290, 27215–27227. [Google Scholar] [CrossRef] [PubMed]
- Khalil, H.F.; Metwalli, N.E.E.D.; Magdy, S.; Shamel, M. Tissue-Friendly Dentin Treatments as a Potential Element in Revascularization Protocol (Ex-Vivo Study). BMC Oral Health 2025, 25, 184. [Google Scholar] [CrossRef]
- Elgamal, M.A.; Khodeer, D.M.; Abdel-Wahab, B.A.; Ibrahim, I.A.A.; Alzahrani, A.R.; Moustafa, Y.M.; Ali, A.A.; El-Sayed, N.M. Canagliflozin Alleviates Valproic Acid-Induced Autism in Rat Pups: Role of PTEN/PDK/PPAR-γ Signaling Pathways. Front. Pharmacol. 2023, 14, 1113966. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Feather-Stone, R.M. A New and Rapid Colorimetric Determination of Acetylcholinesterase Activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Talesa, V.; Romani, R.; Antognelli, C.; Giovannini, E.; Rosi, G. Soluble and Membrane-Bound Acetylcholinesterases in Mytilus galloprovincialis (Pelecypoda: Filibranchia) from the Northern Adriatic Sea. Chem.-Biol. Interact. 2001, 134, 151–166. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manfredelli, D.; Armeni, T.; de Bari, L.; Scirè, A.; Talesa, V.N.; Antognelli, C.; Pariano, M. Acetylcholine Sustains LNCaP Prostate Cancer Cell Migration, Invasion and Proliferation Through Glyoxalase 1/MG-H1 Axis with the Involvement of Osteopontin. Int. J. Mol. Sci. 2025, 26, 4107. https://doi.org/10.3390/ijms26094107
Manfredelli D, Armeni T, de Bari L, Scirè A, Talesa VN, Antognelli C, Pariano M. Acetylcholine Sustains LNCaP Prostate Cancer Cell Migration, Invasion and Proliferation Through Glyoxalase 1/MG-H1 Axis with the Involvement of Osteopontin. International Journal of Molecular Sciences. 2025; 26(9):4107. https://doi.org/10.3390/ijms26094107
Chicago/Turabian StyleManfredelli, Dominga, Tatiana Armeni, Lidia de Bari, Andrea Scirè, Vincenzo Nicola Talesa, Cinzia Antognelli, and Marilena Pariano. 2025. "Acetylcholine Sustains LNCaP Prostate Cancer Cell Migration, Invasion and Proliferation Through Glyoxalase 1/MG-H1 Axis with the Involvement of Osteopontin" International Journal of Molecular Sciences 26, no. 9: 4107. https://doi.org/10.3390/ijms26094107
APA StyleManfredelli, D., Armeni, T., de Bari, L., Scirè, A., Talesa, V. N., Antognelli, C., & Pariano, M. (2025). Acetylcholine Sustains LNCaP Prostate Cancer Cell Migration, Invasion and Proliferation Through Glyoxalase 1/MG-H1 Axis with the Involvement of Osteopontin. International Journal of Molecular Sciences, 26(9), 4107. https://doi.org/10.3390/ijms26094107