
Identification of Potential Prophylactic Medical Countermeasures Against Acute Radiation Syndrome (ARS)

 ,
,  , ,
, , _Guda.png) , , and
, , and
Abstract
1. Introduction
2. Results
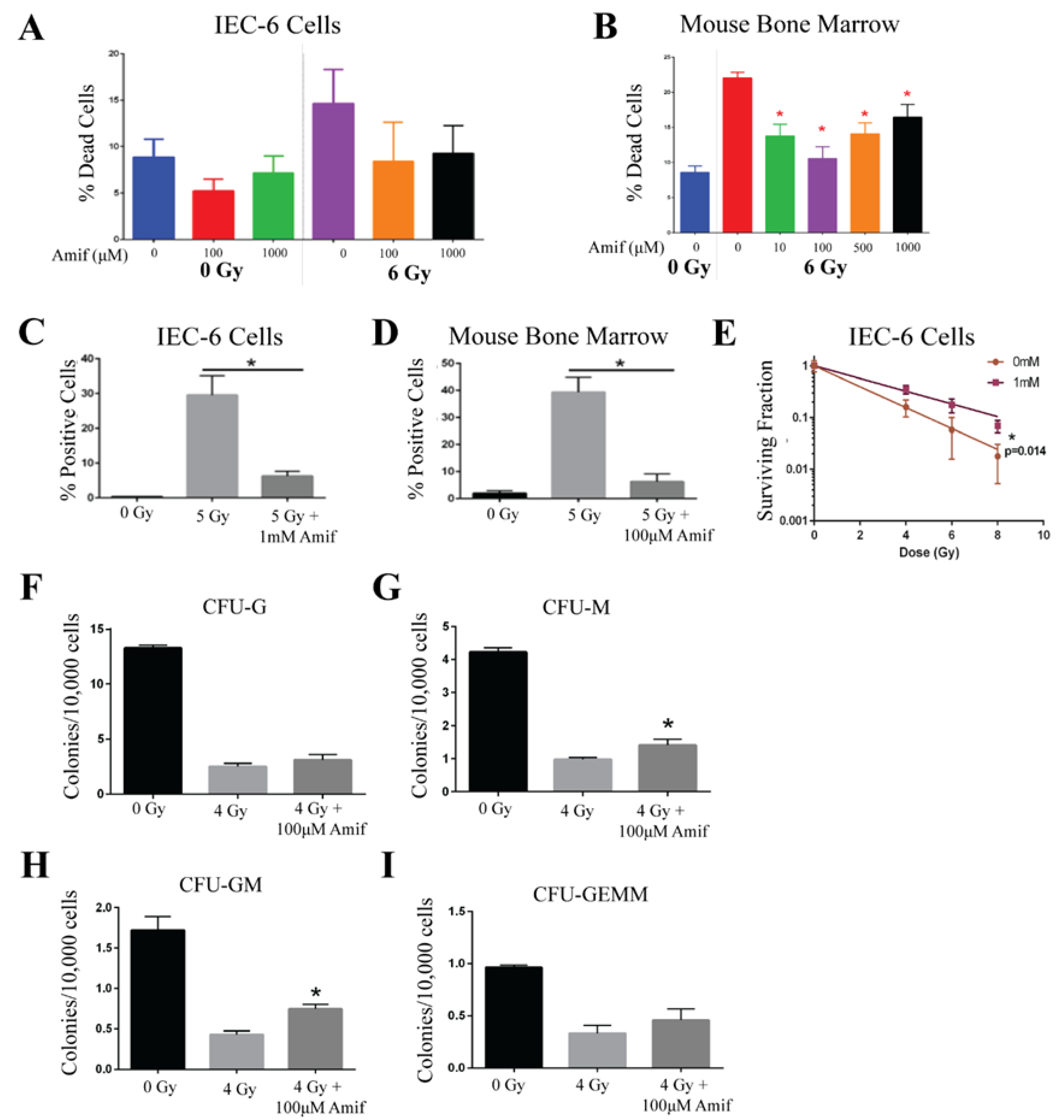
2.1. Amifostine Protects Against Radiation Damage
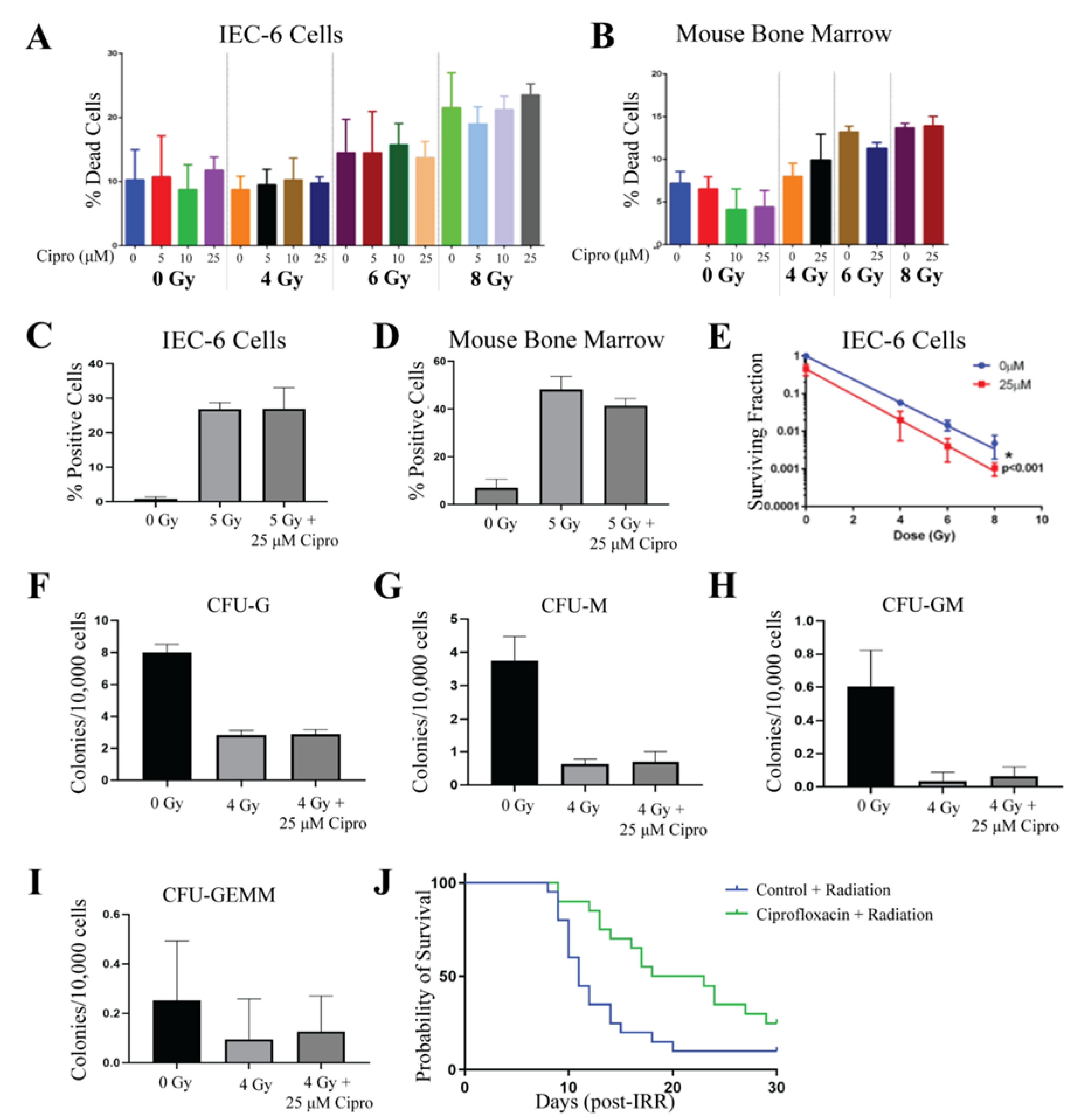
2.2. Ciprofloxacin Does Not Protect Against Radiation-Induced Toxicities
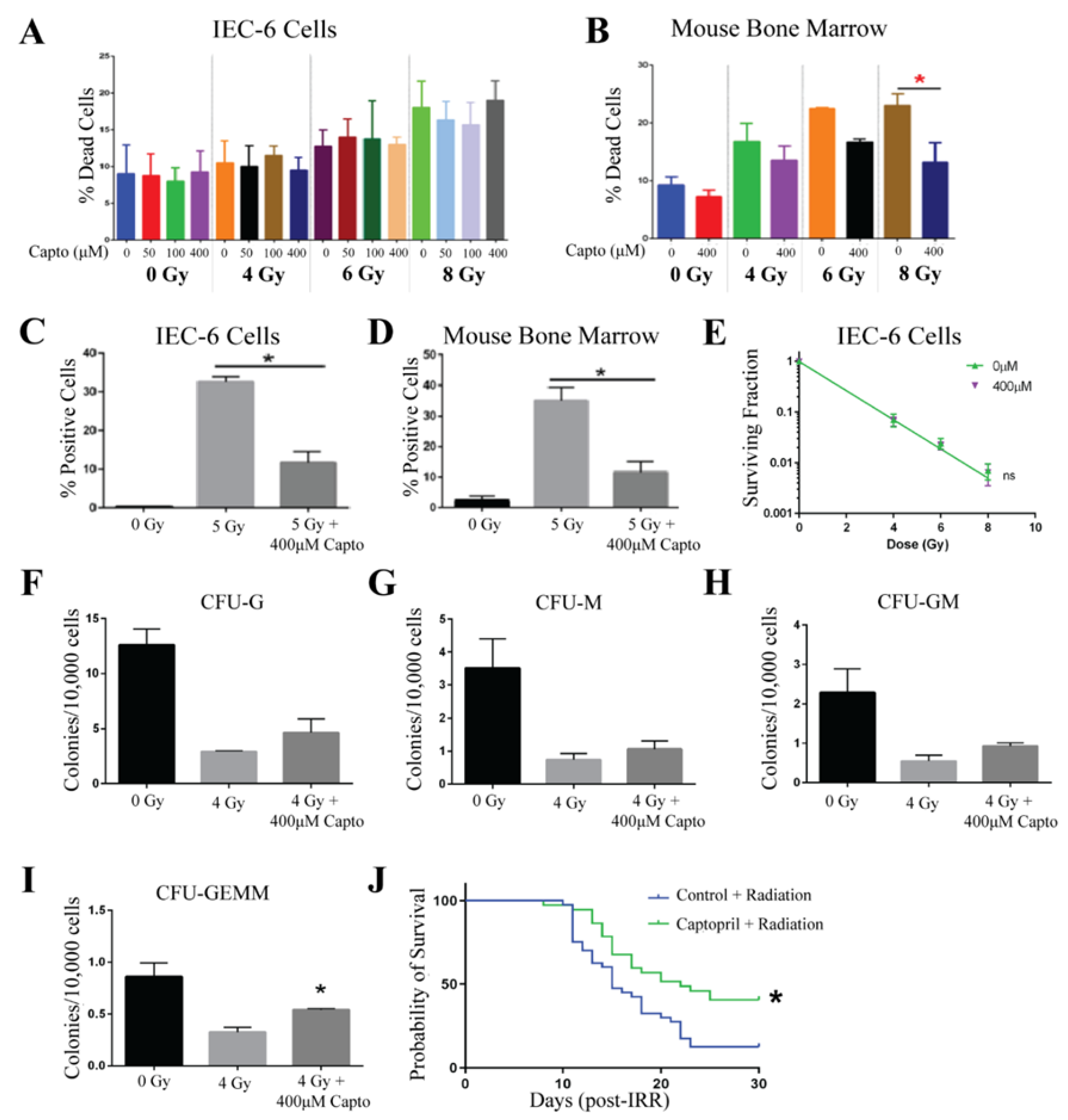
2.3. Captopril
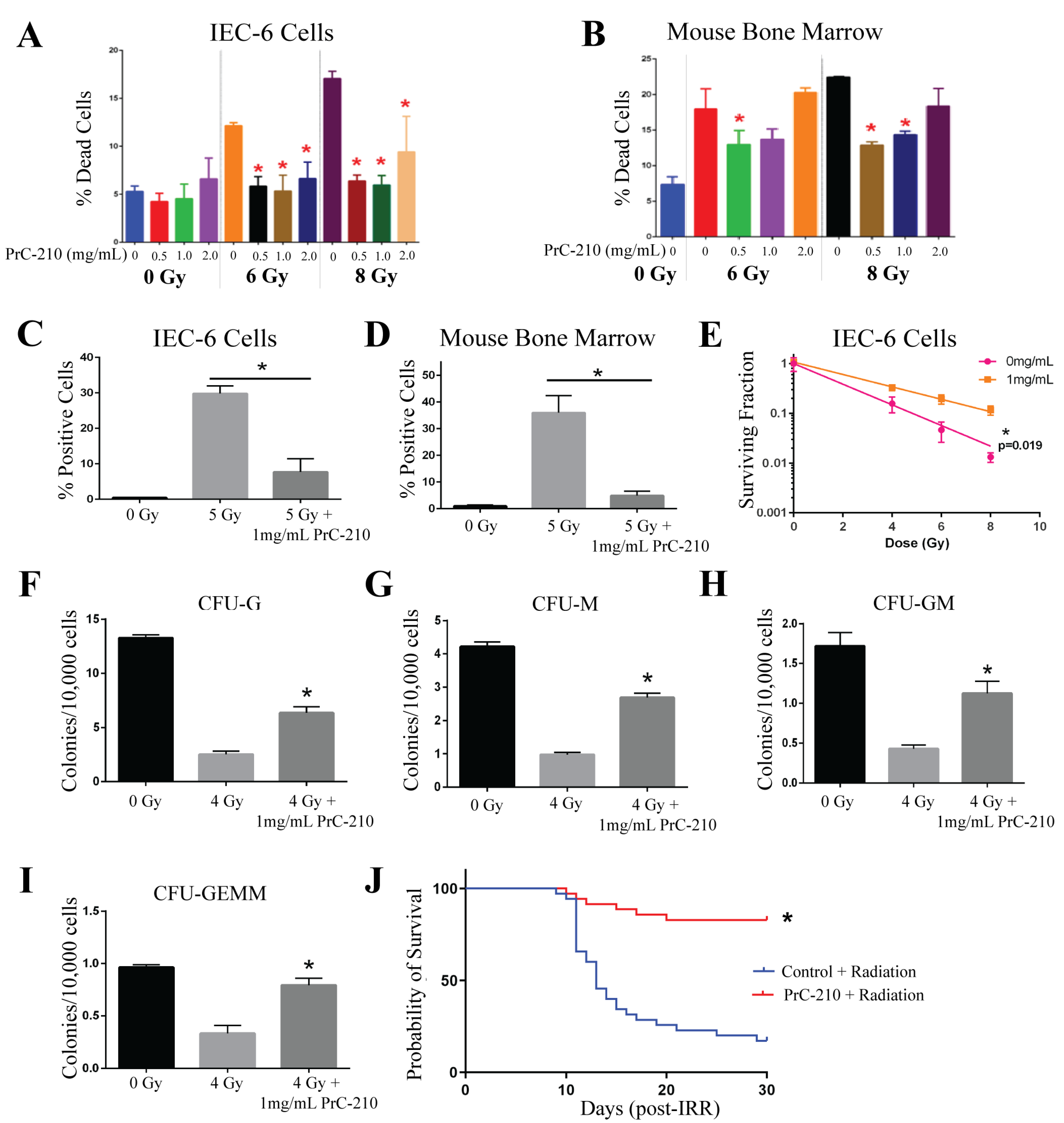
2.4. PrC-210
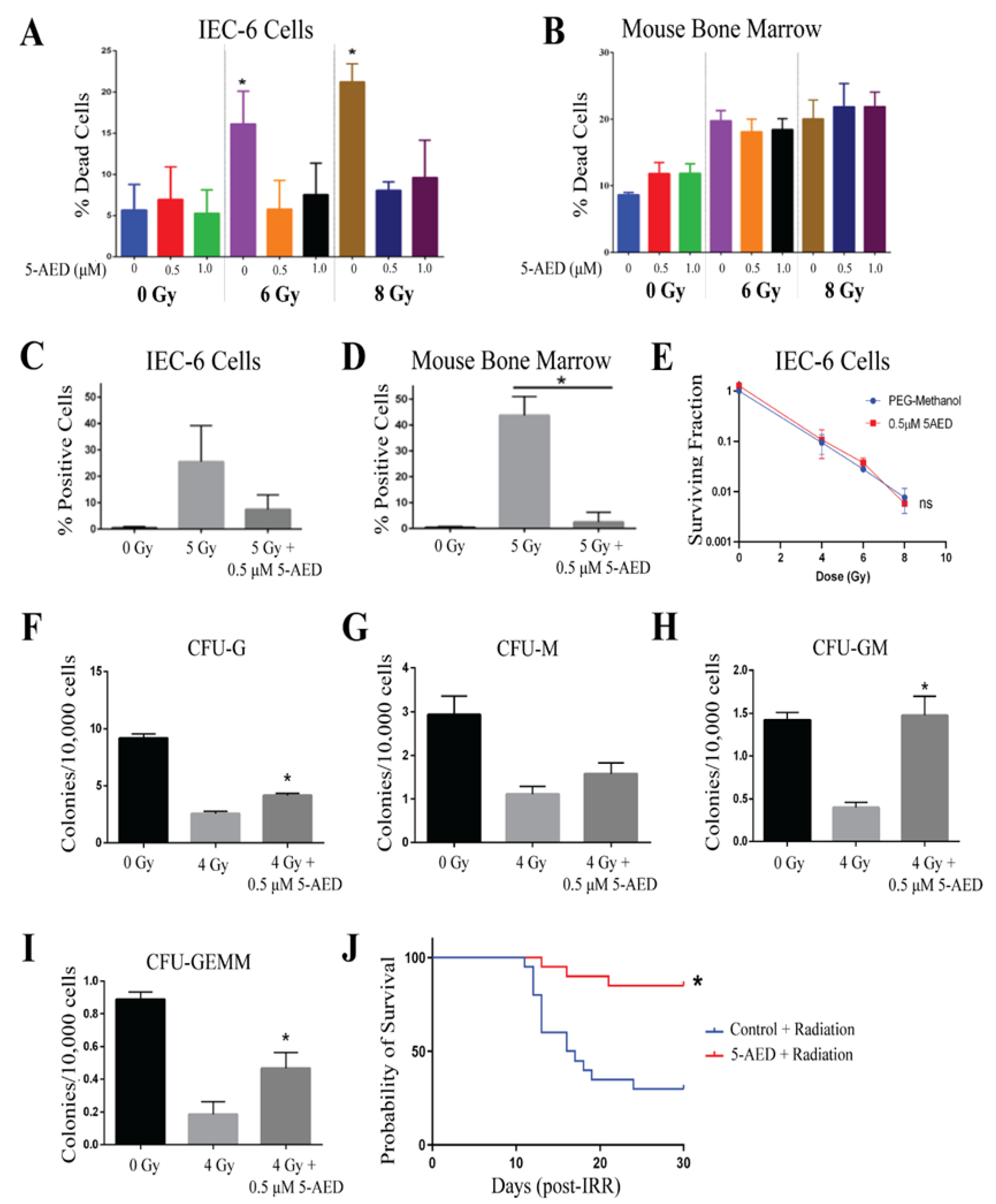
2.5. 5-AED
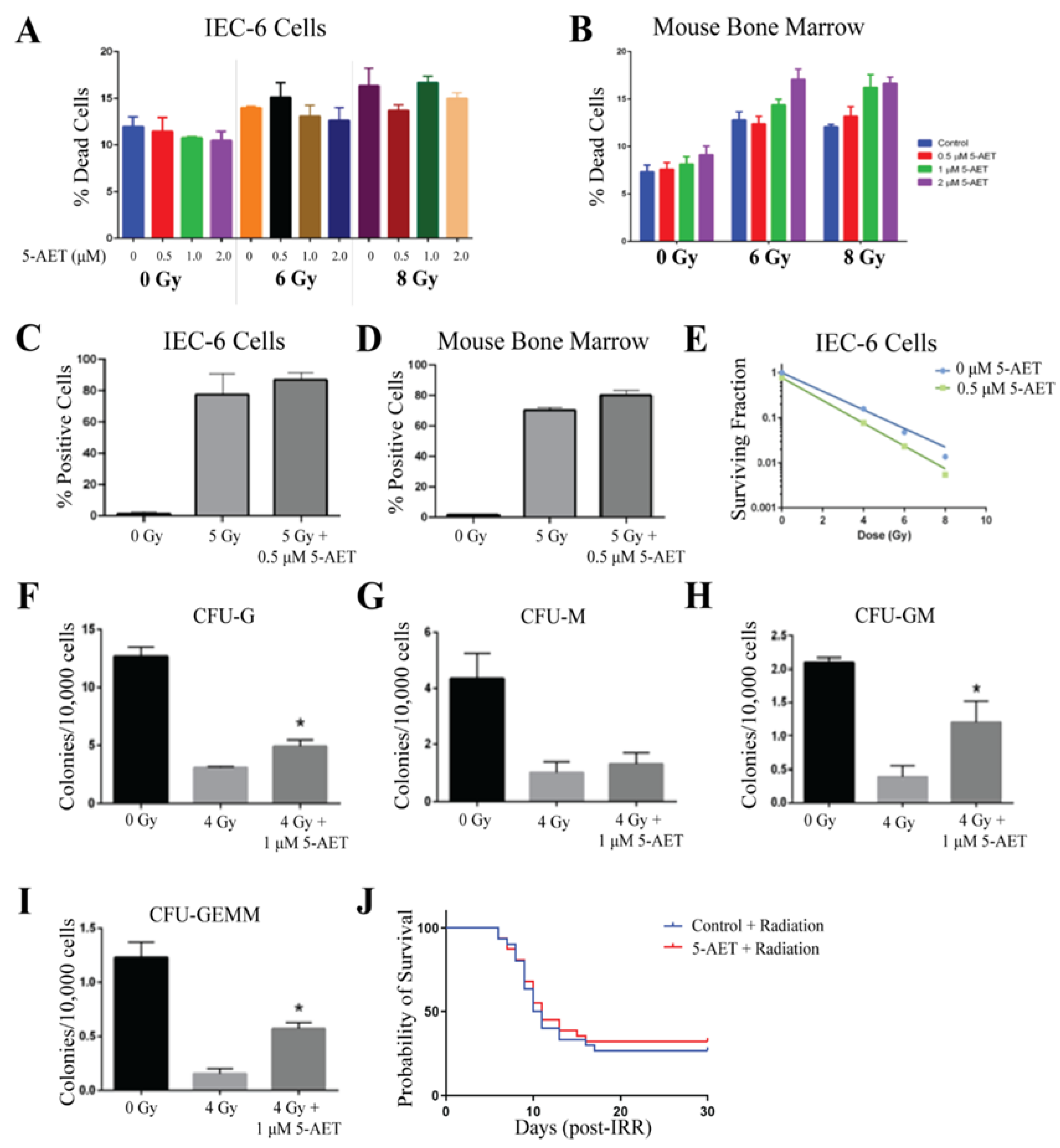
2.6. 5-AET
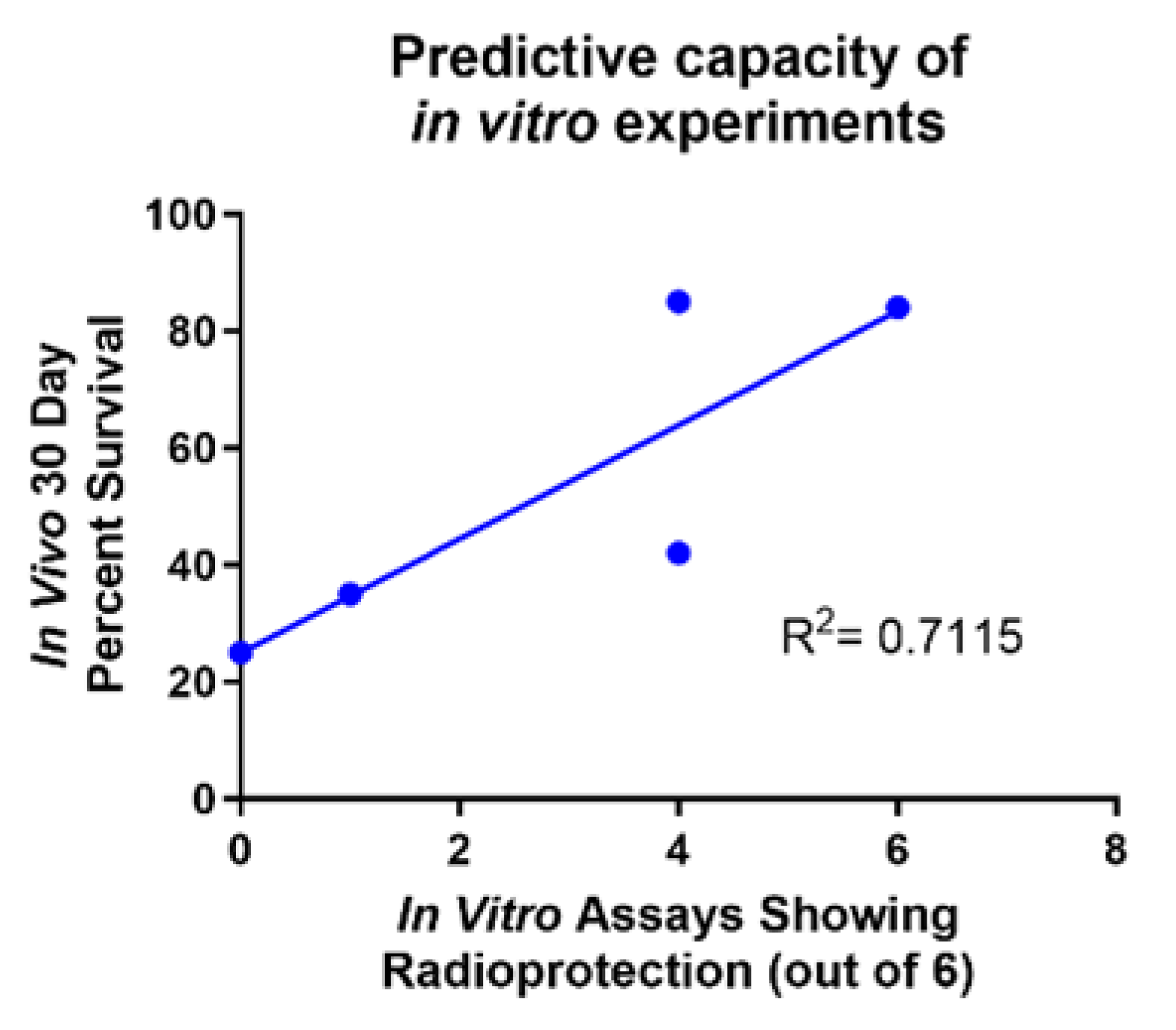
2.7. In Vitro Assays Predict In Vivo Survival
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Experimental Animals
4.3. Molecules Tested
4.4. Mouse Bone Marrow Isolation
4.5. Viability for IEC-6 Cells
4.6. DNA Damage (γ-H2AX Staining)
4.7. Clonogenic Survival
| Treatment | Number of Cells Seeded |
| 0 Gy | 500 |
| 4 Gy | 1000 |
| 6 Gy | 2000 |
| 8 Gy | 4000 |
4.8. Colony Formation
4.9. In Vivo Drug Treatment
4.10. Animal Radiation Treatments
4.11. Mean Survival Time
4.12. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hall, E.J.; Giaccia, A.J. Radiobiology for the Radiologist, 6th ed.; Lippincott Williams and Wilkins: Philadellphia, PA, USA, 2006. [Google Scholar]
- Du, C.; Gao, Z.; Venkatesha, V.A.; Kalen, A.L.; Chaudhuri, L.; Spitz, D.R.; Cullen, J.J.; Oberley, L.W.; Goswami, P.C. Mitochondrial ROS and radiation induced transformation in mouse embryonic fibroblasts. Cancer Biol. Ther. 2009, 8, 1962–1971. [Google Scholar] [CrossRef] [PubMed]
- Indo, H.P.; Inanami, O.; Koumura, T.; Suenaga, S.; Yen, H.-C.; Kakinuma, S.; Matsumoto, K.-I.; Nakanishi, I.; Clair, W.S.; Clair, D.K.S.; et al. Roles of mitochondria-generated reactive oxygen species on X-ray-induced apoptosis in a human hepatocellular carcinoma cell line, HLE. Free Radic. Res. 2012, 46, 1029–1043. [Google Scholar] [CrossRef] [PubMed]
- Riley, P.A. Free radicals in biology: Oxidative stress and the effects of ionizing radiation. Int. J. Radiat. Biol. 1994, 65, 27–33. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Kashiwakura, I. Role of reactive oxygen species in the radiation response of human hematopoietic stem/progenitor cells. PLoS ONE 2013, 8, e70503. [Google Scholar] [CrossRef]
- López, M.; Martín, M. Medical management of the acute radiation syndrome. Rep. Pract. Oncol. Radiother. 2011, 16, 138–146. [Google Scholar] [CrossRef]
- Vercellino, J.; Małachowska, B.; Kulkarni, S.; Bell, B.I.; Shajahan, S.; Shinoda, K.; Eichenbaum, G.; Verma, A.K.; Ghosh, S.P.; Yang, W.-L.; et al. Thrombopoietin mimetic stimulates bone marrow vascular and stromal niches to mitigate acute radiation syndrome. Stem Cell Res. Ther. 2024, 15, 123. [Google Scholar] [CrossRef]
- Singh, V.K.; Newman, V.L.; Berg, A.N.; MacVittie, T.J. Animal models for acute radiation syndrome drug discovery. Expert Opin. Drug Discov. 2015, 10, 497–517. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, I.; Nagai, T.; Miyaishi, K.; Maehara, Y.; Niibe, H. Clinical study of the radioprotective effects of Amifostine (YM-08310, WR-2721) on chronic radiation injury. Int. J. Radiat. Oncol. Biol. Phys. 1986, 12, 935–938. [Google Scholar] [CrossRef]
- Singh, V.K.; Seed, T.M. The efficacy and safety of amifostine for the acute radiation syndrome. Expert Opin. Drug Saf. 2019, 18, 1077–1090. [Google Scholar] [CrossRef]
- Vance-Bryan, K.; Guay, D.R.P.; Rotschafer, J.C. Clinical pharmacokinetics of ciprofloxacin. Clin. Pharmacokinet. 1990, 19, 434–461. [Google Scholar] [CrossRef]
- Fukumoto, R.; Cary, L.H.; Gorbunov, N.V.; Lombardini, E.D.; Elliott, T.B.; Kiang, J.G. Ciprofloxacin modulates cytokine/chemokine profile in serum, improves bone marrow repopulation, and limits apoptosis and autophagy in ileum after whole body ionizing irradiation combined with skin-wound trauma. PLoS ONE 2013, 8, e58389. [Google Scholar] [CrossRef] [PubMed]
- Kiang, J.G.; Fukumoto, R. Ciprofloxacin increases survival after ionizing irradiation combined injury: Gamma-H2AX formation, cytokine/chemokine, and red blood cells. Health Phys. 2014, 106, 720–726. [Google Scholar] [CrossRef]
- Kiang, J.G.; Garrison, B.R.; Smith, J.T.; Fukumoto, R. Ciprofloxacin as a potential radio-sensitizer to tumor cells and a radio-protectant for normal cells: Differential effects on gamma-H2AX formation, p53 phosphorylation, Bcl-2 production, and cell death. Mol. Cell. Biochem. 2014, 393, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Salvetti, A.; Pedrinelli, R.; Arzilli, F.; Abdel-Haq, B.; Magagna, A.; Lucarini, A.; Graziadei, L.; Nuccorini, A.; Taddei, S. Angiotensin-converting enzyme inhibitors in hypertension: A review. Int. J. Clin. Pharmacol. Res. 1985, 5, 429–438. [Google Scholar] [PubMed]
- Chisi, J.E.; Wdzieczak-Bakala, J.; Riches, A.C. Inhibitory action of the peptide AcSDKP on the proliferative state of hematopoietic stem cells in the presence of captopril but not lisinopril. Stem Cells 1997, 15, 455–460. [Google Scholar] [CrossRef]
- de Cavanagh, E.M.; Fraga, C.G.; Ferder, L.; Inserra, F. Enalapril and captopril enhance antioxidant defenses in mouse tissues. Am. J. Physiol. 1997, 272 Pt 2, R514–R518. [Google Scholar] [CrossRef]
- Davis, T.A.; Landauer, M.R.; Mog, S.R.; Barshishat-Kupper, M.; Zins, S.R.; Amare, M.F.; Day, R.M. Timing of captopril administration determines radiation protection or radiation sensitization in a murine model of total body irradiation. Exp. Hematol. 2010, 38, 270–281. [Google Scholar] [CrossRef]
- Islam, A.; Bolduc, D.L.; Zhai, M.; Kiang, J.G.; Swift, J.M. Captopril Increases Survival after Whole-Body Ionizing Irradiation but Decreases Survival when Combined with Skin-Burn Trauma in Mice. Radiat. Res. 2015, 184, 273–279. [Google Scholar] [CrossRef]
- Rittase, W.B.; McCart, E.A.; Muir, J.M.; Bouten, R.M.; Slaven, J.E.; Mungunsukh, O.; Bylicky, M.A.; Wilkins, W.L.; Lee, S.-H.; Gudmundsson, K.O.; et al. Effects of captopril against radiation injuries in the Gottingen minipig model of hematopoietic-acute radiation syndrome. PLoS ONE 2021, 16, e0256208. [Google Scholar] [CrossRef]
- Copp, R.R.; Peebles, D.D.; Fahl, W.E. Synthesis and growth regulatory activity of a prototype member of a new family of aminothiol radioprotectors. Bioorg. Med. Chem. Lett. 2011, 21, 7426–7430. [Google Scholar] [CrossRef]
- Copp, R.R.; Peebles, D.D.; Soref, C.M.; Fahl, W.E. Radioprotective efficacy and toxicity of a new family of aminothiol analogs. Int. J. Radiat. Biol. 2013, 89, 485–492. [Google Scholar] [CrossRef] [PubMed]
- Soref, C.M.; Hacker, T.A.; Fahl, W.E. A new orally active, aminothiol radioprotector-free of nausea and hypotension side effects at its highest radioprotective doses. Int. J. Radiat. Oncol. Biol. Phys. 2012, 82, e701–e707. [Google Scholar] [CrossRef]
- Grace, M.B.; Singh, V.K.; Rhee, J.G.; Jackson, W.E.; Kao, T.-C.; Whitnall, M.H. 5-AED enhances survival of irradiated mice in a G-CSF-dependent manner, stimulates innate immune cell function, reduces radiation-induced DNA damage and induces genes that modulate cell cycle progression and apoptosis. J. Radiat. Res. 2012, 53, 840–853. [Google Scholar] [CrossRef] [PubMed]
- Loria, R.M.; Conrad, D.H.; Huff, T.; Carter, H.; Ben-Nathan, D. Androstenetriol and androstenediol. Protection against lethal radiation and restoration of immunity after radiation injury. Ann. N. Y. Acad. Sci. 2000, 917, 860–867. [Google Scholar] [CrossRef]
- Whitnall, M.H.; Villa, V.; Seed, T.M.; Benjack, J.; Miner, V.; Lewbart, M.L.; Dowding, C.A.; Jackson, W.E., III. Molecular specificity of 5-androstenediol as a systemic radioprotectant in mice. Immunopharmacol. Immunotoxicol. 2005, 27, 15–32. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Liu, W.; Fan, T.; Zhong, H.; Zhou, H.; Guo, W.; Zhu, X. 5-Androstenediol prevents radiation injury in mice by promoting NF-kappaB signaling and inhibiting AIM2 inflammasome activation. Biomed. Pharmacother. 2020, 121, 109597. [Google Scholar] [CrossRef]
- Aerts-Kaya, F.S.; Visser, T.P.; Arshad, S.; Frincke, J.; Stickney, D.R.; Reading, C.L.; Wagemaker, G. 5-Androstene-3beta,17beta-diol promotes recovery of immature hematopoietic cells following myelosuppressive radiation and synergizes with thrombopoietin. Int. J. Radiat. Oncol. Biol. Phys. 2012, 84, e401–e407. [Google Scholar] [CrossRef]
- Stickney, D.R.; Ahlem, C.N.; Morgan, E.; Reading, C.L.; Onizuka, N.; Frincke, J.M. Phase I and Phase II clinical trials of androst-5-ene-3beta,7beta,17beta-triol. Am. J. Transl. Res. 2011, 3, 275–283. [Google Scholar]
- Zhou, S.; Li, Y.; He, L.; Chen, M.; Li, W.; Xiao, T.; Guan, J.; Qi, Z.; Wang, Q.; Li, S.; et al. Haptoglobin is an early indicator of survival after radiation-induced severe injury and bone marrow transplantation in mice. Stem Cell Res. Ther. 2022, 13, 461. [Google Scholar] [CrossRef]
- Wu, T.; Pelus, L.M.; Plett, P.A.; Sampson, C.H.; Chua, H.L.; Fisher, A.; Feng, H.; Liu, L.; Li, H.; Ortiz, M.; et al. Further Characterization of Multi-Organ DEARE and Protection by 16,16 Dimethyl Prostaglandin E2 in a Mouse Model of the Hematopoietic Acute Radiation Syndrome. Radiat. Res. 2023, 199, 468–489. [Google Scholar] [CrossRef]
- Marwah, P.; Marwah, A.; Lardy, H.A.; Miyamoto, H.; Chang, C. C19-steroids as androgen receptor modulators: Design, discovery, and structure-activity relationship of new steroidal androgen receptor antagonists. Bioorg. Med. Chem. 2006, 14, 5933–5947. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drug Name | Experimental Endpoint | Concentration | Cell Types Used | Radiation Protection? |
|---|---|---|---|---|
| Amifostine | Viability | 100, 1000 µM | IEC-6 | NO |
| 10, 100, 500, 1000 µM | Mouse Bone Marrow | YES | ||
| γ-H2AX | 1 mM | IEC-6 | YES | |
| 100 µM | Mouse bone marrow | YES | ||
| Clonogenicity/Colony Formation | 1 mM | IEC-6 | YES | |
| 100 µM | Mouse bone marrow | YES | ||
| Ciprofloxacin | Viability | 5, 10, 25 µM | IEC-6 | NO |
| 25 µM | Mouse Bone Marrow | NO | ||
| γ-H2AX | 25 µM | IEC-6 | NO | |
| 25 µM | Mouse bone marrow | NO | ||
| Clonogenicity/Colony Formation | 5, 25 µM | IEC-6 | NO | |
| 25 µM | Mouse bone marrow | NO | ||
| In Vivo Survival | 0.67 mg/mL (dw) | - | NO | |
| Captopril | Viability | 50, 100, 400 µM | IEC-6 | NO |
| 400 µM | Mouse bone marrow | YES | ||
| γ-H2AX | 400 µM | IEC-6 | YES | |
| 400 µM | Mouse bone marrow | YES | ||
| Clonogenicity/Colony Formation | 400 µM | IEC-6 | NO | |
| 400 µM | Mouse bone marrow | YES | ||
| In Vivo Survival | 110 mg/kg (gav) | - | YES | |
| PrC-210 | Viability | 0.5, 1, 2 mg/mL | IEC-6 | YES |
| 0.5, 1, 2 mg/mL | Mouse bone marrow | YES | ||
| γ-H2AX | 1 mg/mL | IEC-6 | YES | |
| 1 mg/mL | Mouse bone marrow | YES | ||
| Clonogenicity/Colony Formation | 1 mg/mL | IEC-6 | YES | |
| 1 mg/mL | Mouse bone marrow | YES | ||
| In Vivo Survival | 900 mg/kg (ip) | - | YES | |
| 5-AED | Viability | 0.5, 1 µM | IEC-6 | YES |
| 0.5, 1 µM | Mouse bone marrow | NO | ||
| γ-H2AX | 0.5 µM | IEC-6 | NO | |
| 0.5 µM | Mouse bone marrow | YES | ||
| Clonogenicity/Colony Formation | 0.5 µM | IEC-6 | NO | |
| 0.5 µM | Mouse bone marrow | YES | ||
| In Vivo Survival | 30 mg/kg (sc) | - | YES | |
| 5-AET | Viability | 0.5, 1, 2 µM | IEC-6 | NO |
| 0.5, 1, 2 µM | Mouse bone marrow | NO | ||
| γ-H2AX | 0.5 µM | IEC-6 | NO | |
| 0.5 µM | Mouse bone marrow | NO | ||
| Clonogenicity/Colony Formation | 0.5 µM | IEC-6 | NO | |
| 0.5 µM | Mouse bone marrow | YES | ||
| In Vivo Survival | 30 mg/kg (sc) | - | NO |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liermann-Wooldrik, K.T.; Chatterjee, A.; Kosmacek, E.A.; Myers, M.S.; Adebisi, O.; Monga-Wells, L.; Mei, L.; Takacs, M.P.; Dussault, P.H.; Draney, D.R.; et al. Identification of Potential Prophylactic Medical Countermeasures Against Acute Radiation Syndrome (ARS). Int. J. Mol. Sci. 2025, 26, 4055. https://doi.org/10.3390/ijms26094055
Liermann-Wooldrik KT, Chatterjee A, Kosmacek EA, Myers MS, Adebisi O, Monga-Wells L, Mei L, Takacs MP, Dussault PH, Draney DR, et al. Identification of Potential Prophylactic Medical Countermeasures Against Acute Radiation Syndrome (ARS). International Journal of Molecular Sciences. 2025; 26(9):4055. https://doi.org/10.3390/ijms26094055
Chicago/Turabian StyleLiermann-Wooldrik, Kia T., Arpita Chatterjee, Elizabeth A. Kosmacek, Molly S. Myers, Oluwaseun Adebisi, Louise Monga-Wells, Liu Mei, Michelle P. Takacs, Patrick H. Dussault, Daniel R. Draney, and et al. 2025. "Identification of Potential Prophylactic Medical Countermeasures Against Acute Radiation Syndrome (ARS)" International Journal of Molecular Sciences 26, no. 9: 4055. https://doi.org/10.3390/ijms26094055
APA StyleLiermann-Wooldrik, K. T., Chatterjee, A., Kosmacek, E. A., Myers, M. S., Adebisi, O., Monga-Wells, L., Mei, L., Takacs, M. P., Dussault, P. H., Draney, D. R., Powers, R., Checco, J. W., Guda, C., Helikar, T., Berkowitz, D. B., Bayles, K. W., Epstein, A. H., Cary, L., Murry, D. J., & Oberley-Deegan, R. E. (2025). Identification of Potential Prophylactic Medical Countermeasures Against Acute Radiation Syndrome (ARS). International Journal of Molecular Sciences, 26(9), 4055. https://doi.org/10.3390/ijms26094055