
The Protective Role of DUSP4 in Retinal Pigment Epithelium Senescence and Degeneration

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
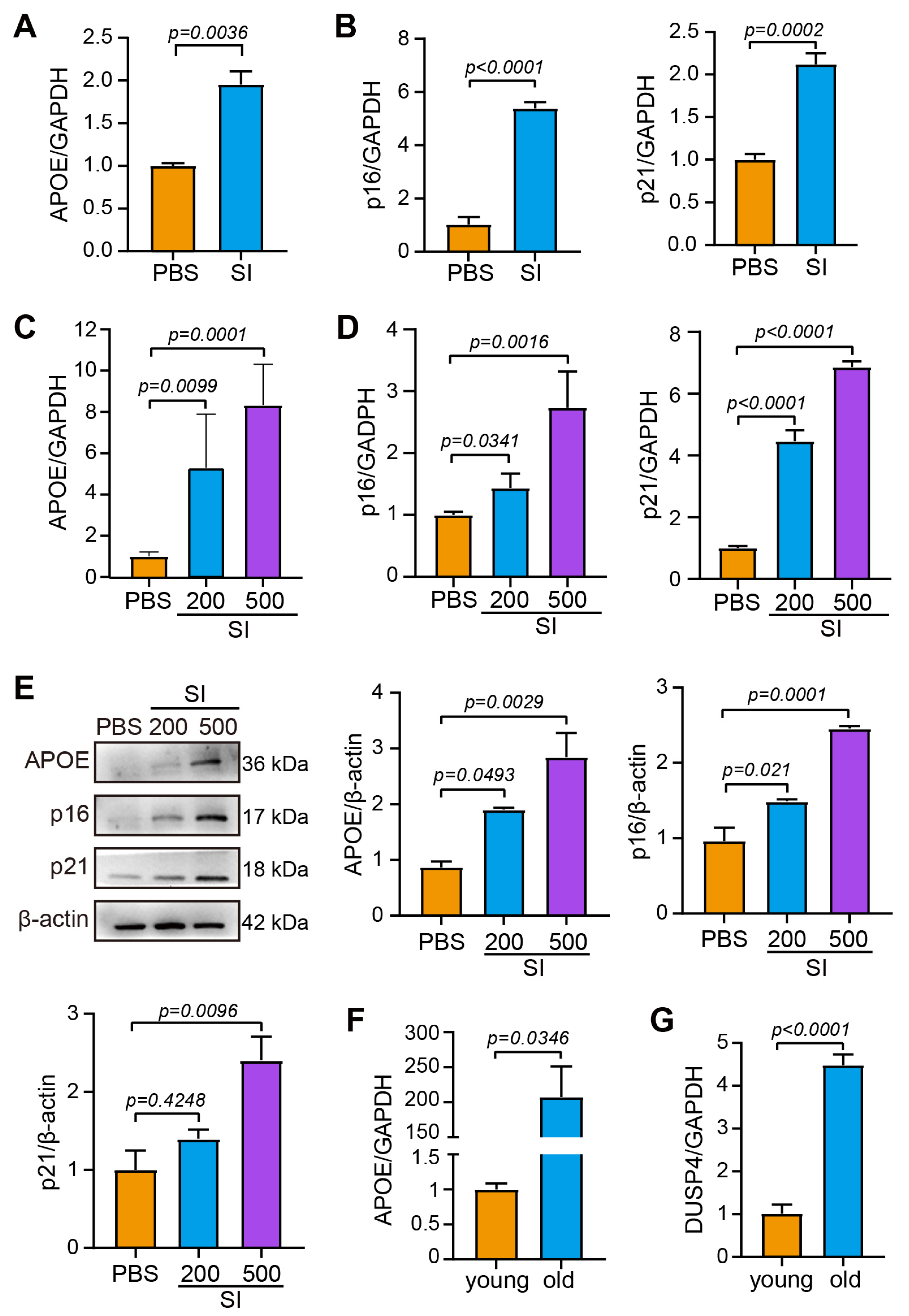
2.1. The Upregulation of DUSP4 in Dry AMD
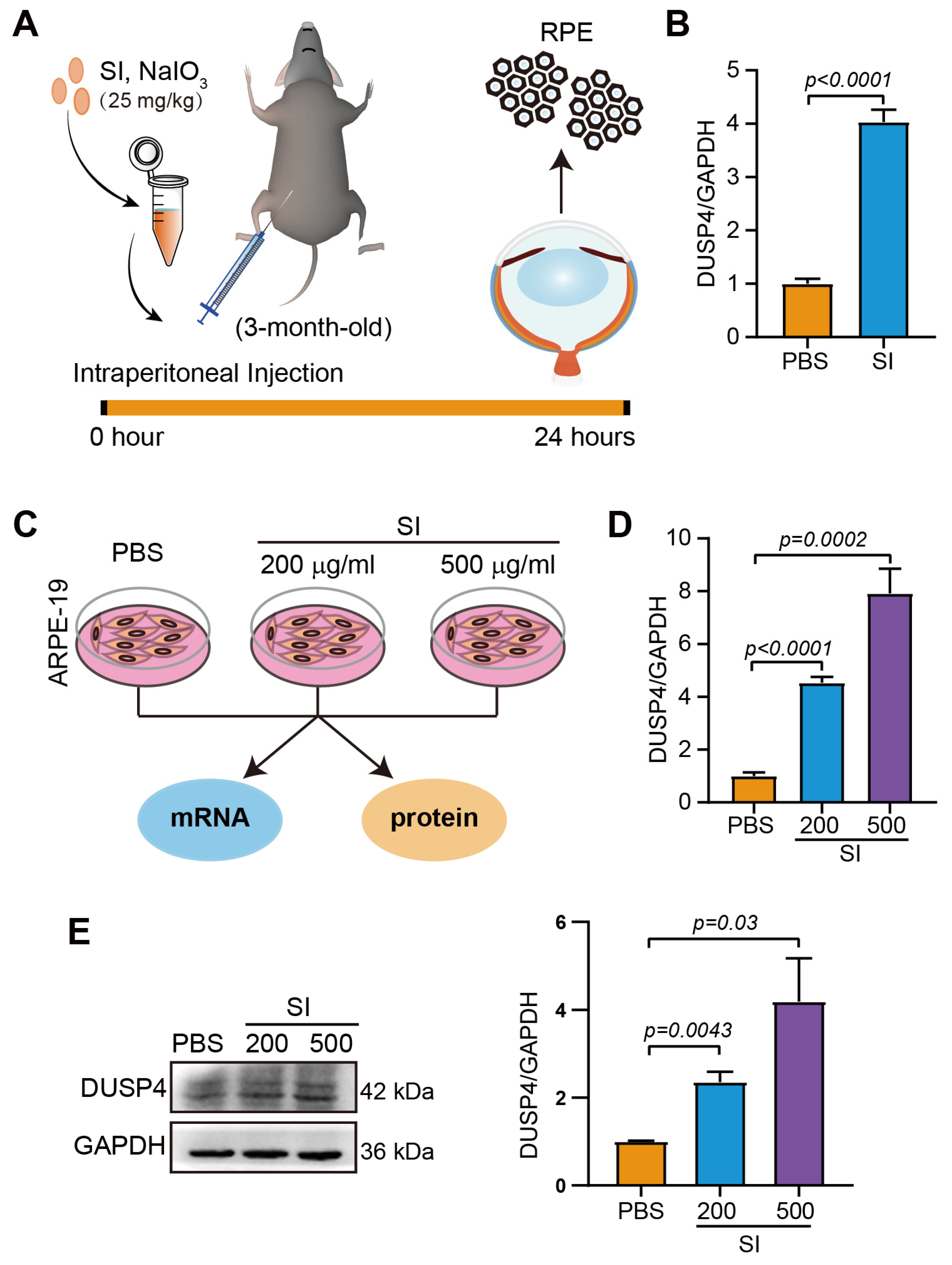
2.2. Senescence of the RPE Is Accompanied by High Expression of DUSP4
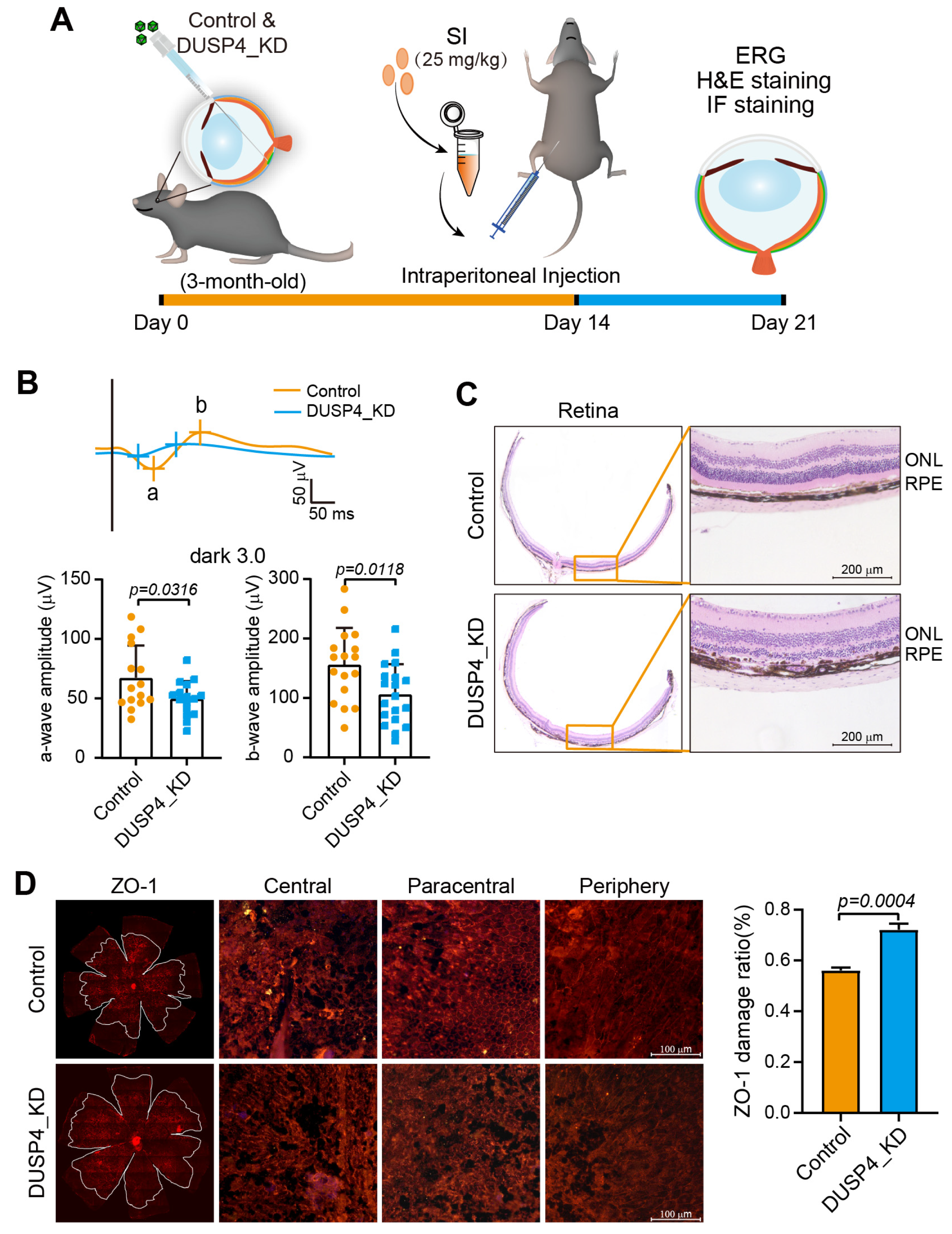
2.3. DUSP4 Deficiency in RPE Exacerbates AMD Progression
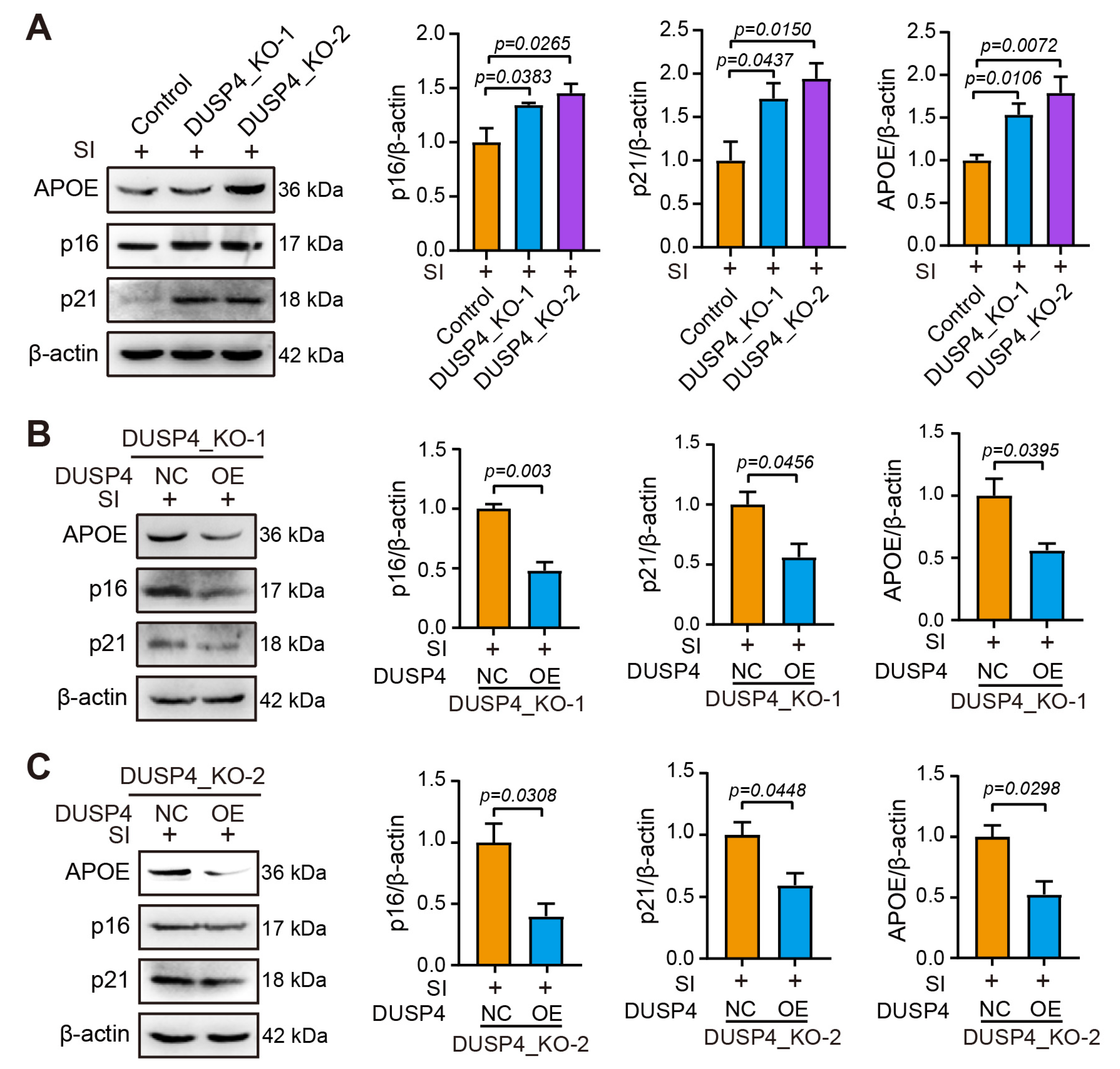
2.4. DUSP4 Dysfunction Exacerbates RPE Senescence
2.5. DUSP4 Slows RPE Senescence by Coordinately Regulating p38, p53, and NF-κB Pathways
3. Discussion
4. Materials and Methods
4.1. Antibodies, Plasmids and Chemicals
4.2. Animals
4.3. Cell Culture and Transfection
4.4. DUSP4_Knockout (KO) Stable Cell Lines Edited by CRISPR/Cas9 Technology
4.5. SI-Induced Dry AMD Model
4.6. Immunofluorescent (IF) Staining of ZO-1
4.7. DUSP4_KD in the RPE of Mice
4.8. Optical Coherence Tomography (OCT) and Fundus Imaging
4.9. Electroretinography (ERG)
4.10. Quantitative Real-Time Polymerase Chain Reaction (qPCR)
4.11. Western Blotting
4.12. Retinal Histology
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| DUSP4 | Dual-specificity phosphatase 4 |
| AMD | Age-related macular degeneration |
| SI | Sodium iodate |
| APOE | Apolipoprotein E |
| p16 | p16INK4a |
| p21 | p21Cip/WAF1 |
| ERG | Electroretinography |
| H&E | Hematoxylin and eosin |
| IF | Immunofluorescence |
| ZO-1 | Zonula occludens-1 |
| KO | Knockout |
| KD | Knockdown |
| GFP | Green fluorescent protein |
References
- Guymer, R.H.; Campbell, T.G. Age-related macular degeneration. Lancet 2023, 401, 1459–1472. [Google Scholar] [CrossRef] [PubMed]
- Flaxel, C.J.; Adelman, R.A.; Bailey, S.T.; Fawzi, A.; Lim, J.I.; Vemulakonda, G.A.; Ying, G.-S. Age-Related Macular Degeneration Preferred Practice Pattern(R). Ophthalmology 2020, 127, 1–65. [Google Scholar] [CrossRef] [PubMed]
- Rudnicka, A.R.; Jarrar, Z.; Wormald, R.; Cook, D.G.; Fletcher, A.; Owen, C.G. Age and gender variations in age-related macular degeneration prevalence in populations of European ancestry: A meta-analysis. Ophthalmology 2012, 119, 571–580. [Google Scholar] [CrossRef] [PubMed]
- Rim, T.H.; Kawasaki, R.; Tham, Y.C.; Kang, S.W.; Ruamviboonsuk, P.; Bikbov, M.M.; Miyake, M.; Hao, J.; Fletcher, A.; Sasaki, M.; et al. Prevalence and pattern of geographic atrophy in asia: The asian eye epidemiology consortium. Ophthalmology 2020, 127, 1371–1381. [Google Scholar] [CrossRef]
- Wiley, C.D.; Campisi, J. The metabolic roots of senescence: Mechanisms and opportunities for intervention. Nat. Metab. 2021, 3, 1290–1301. [Google Scholar] [CrossRef]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. Hallmarks of aging: An expanding universe. Cell 2023, 186, 243–278. [Google Scholar] [CrossRef]
- Milanovic, M.; Fan, D.N.Y.; Belenki, D.; Däbritz, J.H.M.; Zhao, Z.; Yu, Y.; Dörr, J.R.; Dimitrova, L.; Lenze, D.; Monteiro Barbosa, I.A.; et al. Senescence-associated reprogramming promotes cancer stemness. Nature 2018, 553, 96–100. [Google Scholar] [CrossRef]
- Janic, A.; Valente, L.J.; Wakefield, M.J.; Di Stefano, L.; Milla, L.; Wilcox, S.; Yang, H.; Tai, L.; Vandenberg, C.J.; Kueh, A.J.; et al. DNA repair processes are critical mediators of p53-dependent tumor suppression. Nat. Med. 2018, 24, 947–953. [Google Scholar] [CrossRef]
- Li, X.; Wang, J.; Wang, L.; Gao, Y.; Feng, G.; Li, G.; Zou, J.; Yu, M.; Li, Y.F.; Liu, C.; et al. Lipid metabolism dysfunction induced by age-dependent DNA methylation accelerates aging. Signal Transduct. Target. Ther. 2022, 7, 162. [Google Scholar] [CrossRef]
- Senabouth, A.; Daniszewski, M.; Lidgerwood, G.E.; Liang, H.H.; Hernández, D.; Mirzaei, M.; Keenan, S.N.; Zhang, R.; Han, X.; Neavin, D.; et al. Transcriptomic and proteomic retinal pigment epithelium signatures of age-related macular degeneration. Nat. Commun. 2022, 13, 4233. [Google Scholar] [CrossRef]
- Do, K.V.; Kautzmann, M.-A.I.; Jun, B.; Gordon, W.C.; Nshimiyimana, R.; Yang, R.; Petasis, N.A.; Bazan, N.G. Elovanoids counteract oligomeric beta-amyloid-induced gene expression and protect photoreceptors. Proc. Natl. Acad. Sci. USA 2019, 116, 24317–24325. [Google Scholar] [CrossRef] [PubMed]
- Maiese, K.; Chong, Z.Z.; Hou, J.; Shang, Y.C. The vitamin nicotinamide: Translating nutrition into clinical care. Molecules 2009, 14, 3446–3485. [Google Scholar] [CrossRef] [PubMed]
- Saini, J.S.; Corneo, B.; Miller, J.D.; Kiehl, T.R.; Wang, Q.; Boles, N.C.; Blenkinsop, T.A.; Stern, J.H.; Temple, S. Nicotinamide ameliorates disease phenotypes in a human iPSC model of age-related macular degeneration. Cell Stem Cell 2017, 20, 635–647. [Google Scholar] [CrossRef] [PubMed]
- Chae, J.-B.; Jang, H.; Son, C.; Park, C.-W.; Choi, H.; Jin, S.; Lee, H.-Y.; Lee, H.; Ryu, J.-H.; Kim, N.; et al. Targeting senescent retinal pigment epithelial cells facilitates retinal regeneration in mouse models of age-related macular degeneration. Geroscience 2022, 44(3), 1885. [Google Scholar] [CrossRef]
- Al-Mutairi, M.S.; Cadalbert, L.C.; McGachy, H.A.; Shweash, M.; Schroeder, J.; Kurnik, M.; Sloss, C.M.; Bryant, C.E.; Alexander, J.; Plevin, R. MAP kinase phosphatase-2 plays a critical role in response to infection by Leishmania mexicana. PLoS Pathog. 2010, 6, e1001192. [Google Scholar] [CrossRef]
- Checker, R.; Gambhir, L.; Sharma, D.; Kumar, M.; Sandur, S.K. Plumbagin induces apoptosis in lymphoma cells via oxidative stress mediated glutathionylation and inhibition of mitogen-activated protein kinase phosphatases (MKP1/2). Cancer Lett. 2015, 357, 265–278. [Google Scholar] [CrossRef]
- Barajas-Espinosa, A.; Basye, A.; Angelos, M.G.; Chen, C.-A. Modulation of p38 kinase by DUSP4 is important in regulating cardiovascular function under oxidative stress. Free Radic. Biol. Med. 2015, 89, 170–181. [Google Scholar] [CrossRef]
- Dougherty, J.A.; Myers, J.K.; Khan, M.; Angelos, M.G.; Chen, C.-A. Dual-Specificity Phosphatase 4 Overexpression in Cells Prevents Hypoxia/Reoxygenation-Induced Apoptosis via the Upregulation of eNOS. Front. Cardiovasc. Med. 2017, 4, 22. [Google Scholar] [CrossRef]
- Hao, S.-H.; Ma, X.-D.; Xu, L.; Xie, J.-D.; Feng, Z.-H.; Chen, J.-W.; Chen, R.-X.; Wang, F.-W.; Tang, Y.-H.; Xie, D.; et al. Dual specific phosphatase 4 suppresses ferroptosis and enhances sorafenib resistance in hepatocellular carcinoma. Drug Resist. Updat. 2024, 73, 101052. [Google Scholar] [CrossRef]
- Jiao, H.; James, S.J.; Png, C.W.; Cui, C.; Li, H.; Li, L.; Ni Chia, W.; Min, N.; Li, W.; Claser, C.; et al. DUSP4 modulates RIG-I- and STING-mediated IRF3-type I IFN response. Cell Death Differ. 2024, 31, 280–291. [Google Scholar] [CrossRef]
- Hamnett, R.; Crosby, P.; Chesham, J.E.; Hastings, M.H. Vasoactive intestinal peptide controls the suprachiasmatic circadian clock network via ERK1/2 and DUSP4 signalling. Nat. Commun. 2019, 10, 542. [Google Scholar] [CrossRef] [PubMed]
- Newman, A.M.; Gallo, N.B.; Hancox, L.S.; Miller, N.J.; Radeke, C.M.; Maloney, M.A.; Cooper, J.B.; Hageman, G.S.; Anderson, D.H.; Johnson, L.V.; et al. Systems-level analysis of age-related macular degeneration reveals global biomarkers and phenotype-specific functional networks. Genome Med. 2012, 4, 16. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Li, D.; Cao, G.; Shi, Q.; Zhu, J.; Zhang, M.; Cheng, H.; Wen, Q.; Xu, H.; Zhu, L.; et al. IL-27 signalling promotes adipocyte thermogenesis and energy expenditure. Nature 2021, 600, 314–318. [Google Scholar] [CrossRef] [PubMed]
- Schinzel, R.T.; Higuchi-Sanabria, R.; Shalem, O.; Moehle, E.A.; Webster, B.M.; Joe, L.; Bar-Ziv, R.; Frankino, P.A.; Durieux, J.; Pender, C.; et al. The hyaluronidase, TMEM2, promotes ER homeostasis and longevity independent of the UPRER. Cell 2019, 179, 1306–1318. [Google Scholar] [CrossRef]
- Lukjanenko, L.; Jung, M.J.; Hegde, N.; Perruisseau-Carrier, C.; Migliavacca, E.; Rozo, M.; Karaz, S.; Jacot, G.; Schmidt, M.; Li, L.; et al. Loss of fibronectin from the aged stem cell niche affects the regenerative capacity of skeletal muscle in mice. Nat. Med. 2016, 22, 897–905. [Google Scholar] [CrossRef]
- Zeng, J.; Hills, S.A.; Ozono, E.; Diffley, J.F. Cyclin E-induced replicative stress drives p53-dependent whole-genome duplication. Cell 2023, 186, 528–542. [Google Scholar] [CrossRef]
- Garg, C.; Sharma, H.; Garg, M. Skin photo-protection with phytochemicals against photo-oxidative stress, photo-carcinogenesis, signal transduction pathways and extracellular matrix remodeling-An overview. Ageing Res. Rev. 2020, 62, 101127. [Google Scholar] [CrossRef]
- Strunz, M.; Simon, L.M.; Ansari, M.; Kathiriya, J.J.; Angelidis, I.; Mayr, C.H.; Tsidiridis, G.; Lange, M.; Mattner, L.F.; Yee, M.; et al. Alveolar regeneration through a Krt8+transitional stem cell state that persists in human lung fibrosis. Nat. Commun. 2020, 11, 3559. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Ni, Z.; Zhang, J.; Lin, X.; Wu, C.; Wu, Y.; Dong, L.; Zhang, Z.; Chi, Z.-L. The Protective Role of DUSP4 in Retinal Pigment Epithelium Senescence and Degeneration. Int. J. Mol. Sci. 2025, 26, 3735. https://doi.org/10.3390/ijms26083735
Liu X, Ni Z, Zhang J, Lin X, Wu C, Wu Y, Dong L, Zhang Z, Chi Z-L. The Protective Role of DUSP4 in Retinal Pigment Epithelium Senescence and Degeneration. International Journal of Molecular Sciences. 2025; 26(8):3735. https://doi.org/10.3390/ijms26083735
Chicago/Turabian StyleLiu, Xiyuan, Zhaoze Ni, Jing Zhang, Xiaoyan Lin, Chenxin Wu, Yuyang Wu, Lingqin Dong, Zongduan Zhang, and Zai-Long Chi. 2025. "The Protective Role of DUSP4 in Retinal Pigment Epithelium Senescence and Degeneration" International Journal of Molecular Sciences 26, no. 8: 3735. https://doi.org/10.3390/ijms26083735
APA StyleLiu, X., Ni, Z., Zhang, J., Lin, X., Wu, C., Wu, Y., Dong, L., Zhang, Z., & Chi, Z.-L. (2025). The Protective Role of DUSP4 in Retinal Pigment Epithelium Senescence and Degeneration. International Journal of Molecular Sciences, 26(8), 3735. https://doi.org/10.3390/ijms26083735