
2-(3,4-Dichlorophenoxy)triethylamine (DCPTA) Sustains Root Activity Through the Enhancement of Ascorbate-Glutathione in Spring Maize (Zea mays L.) Under Post-Tasseling Waterlogging

 , ,
, ,
Abstract
1. Introduction
2. Results
2.1. Yield and Agronomic Characters
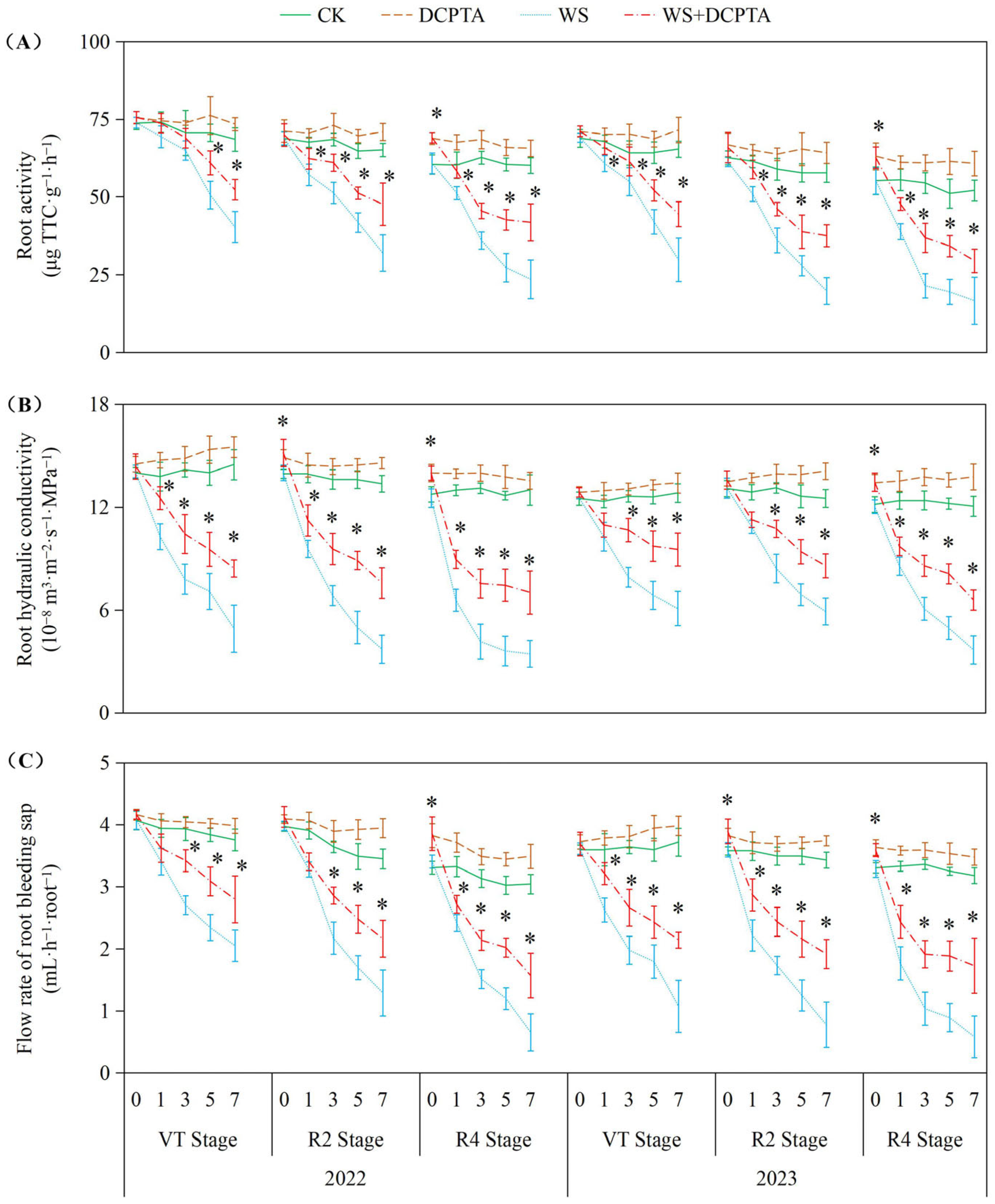
2.2. Water Transport Capacity to Overground Part
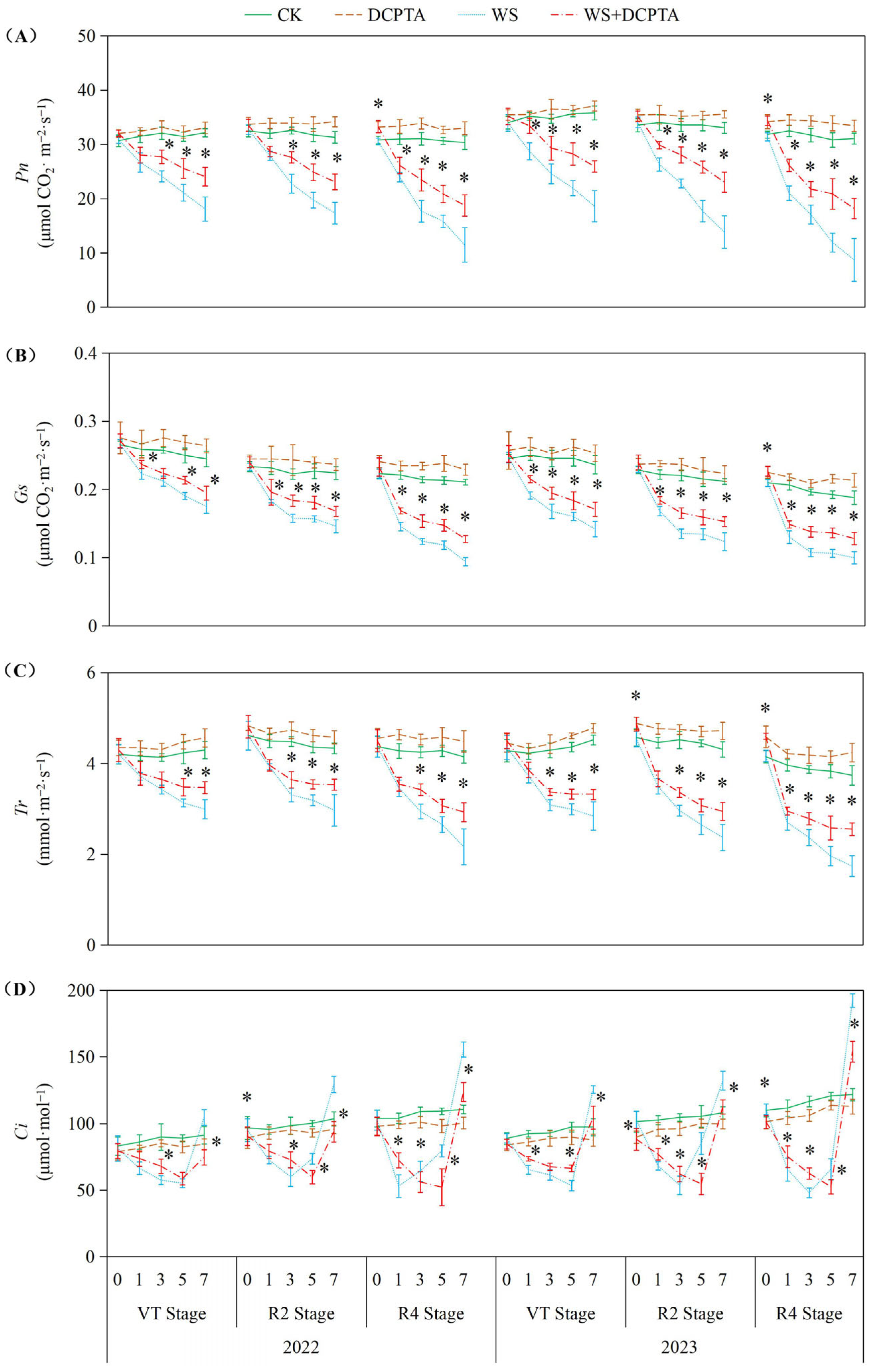
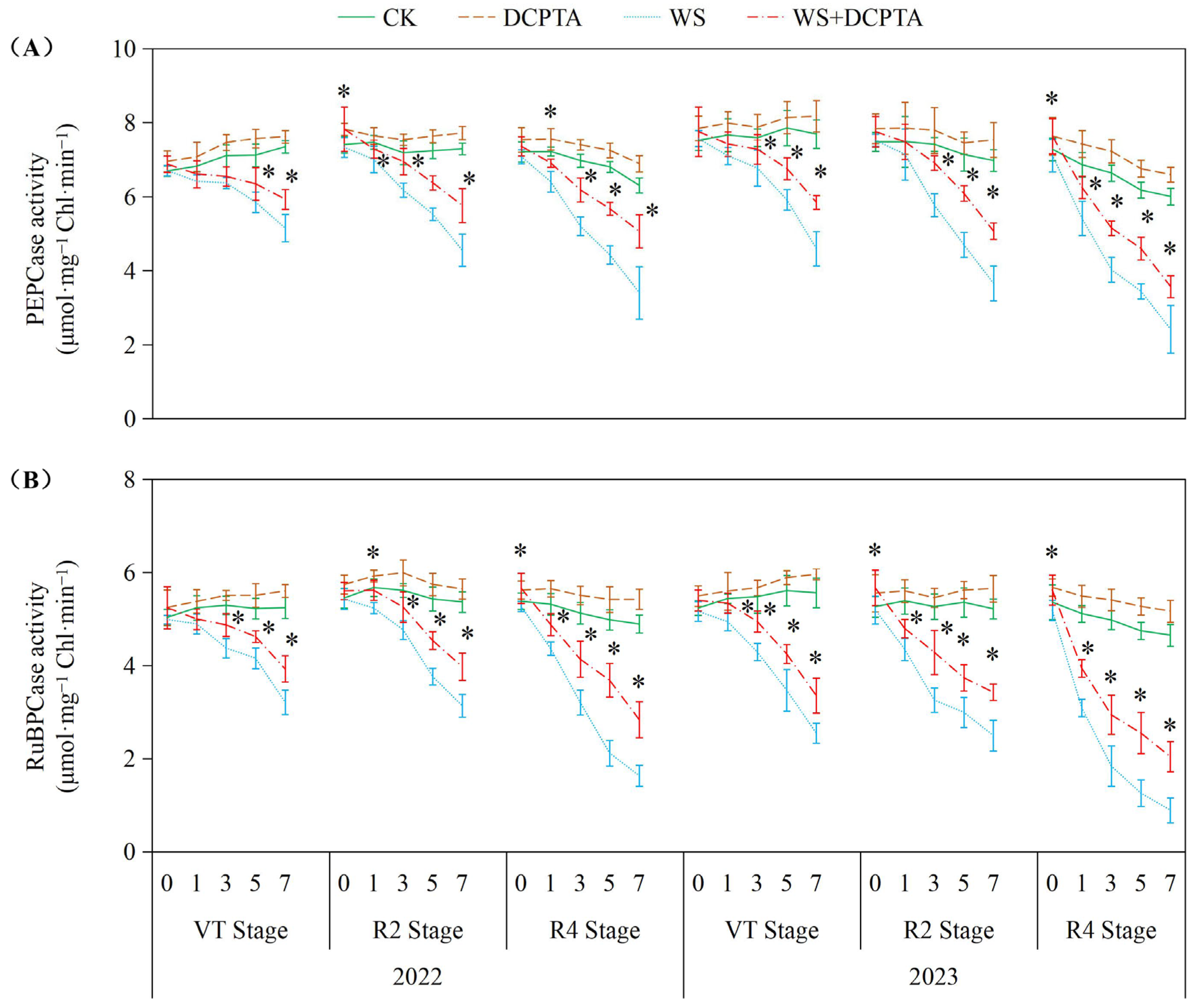
2.3. Leaf Photosynthetic Performance
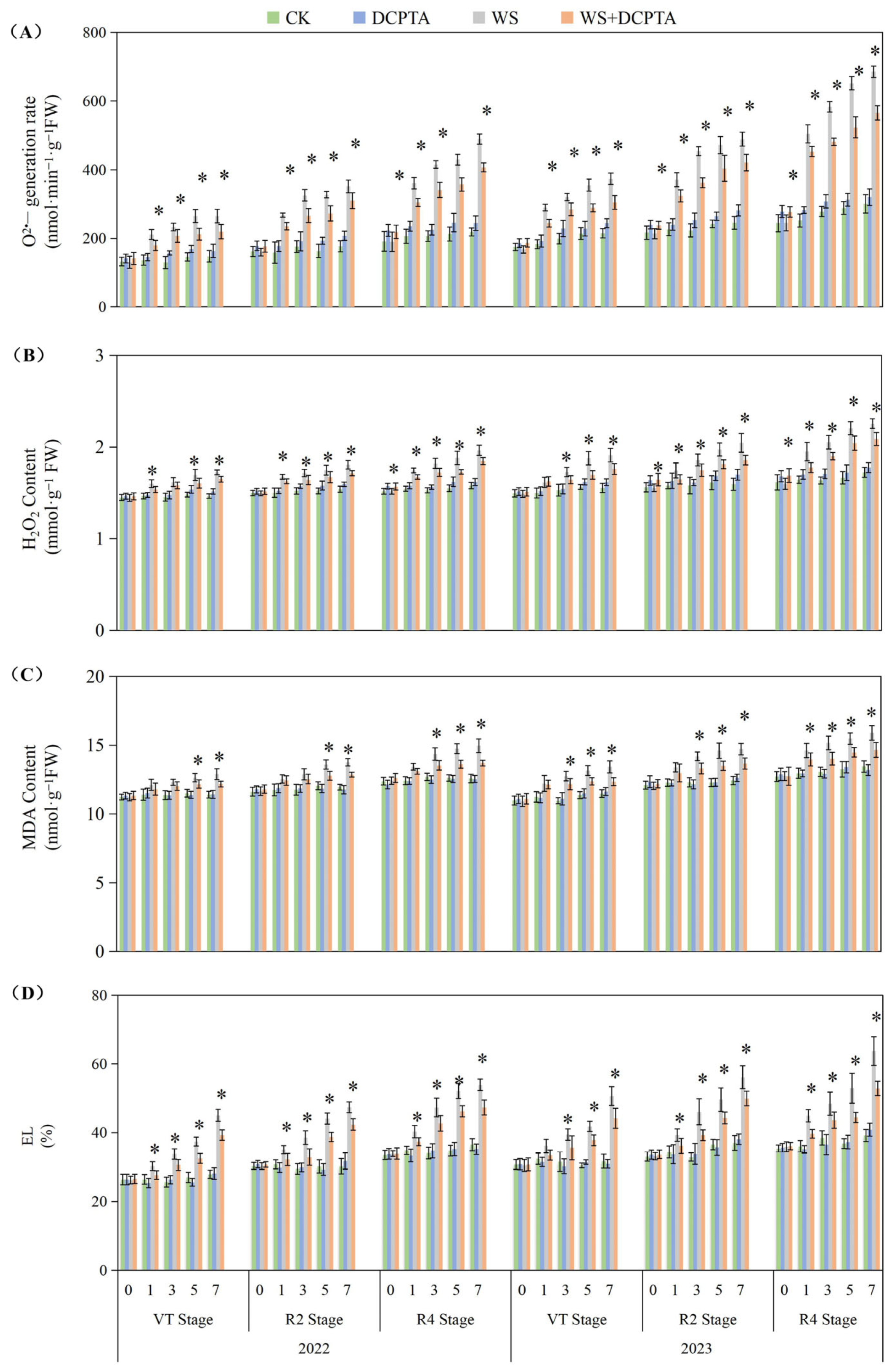
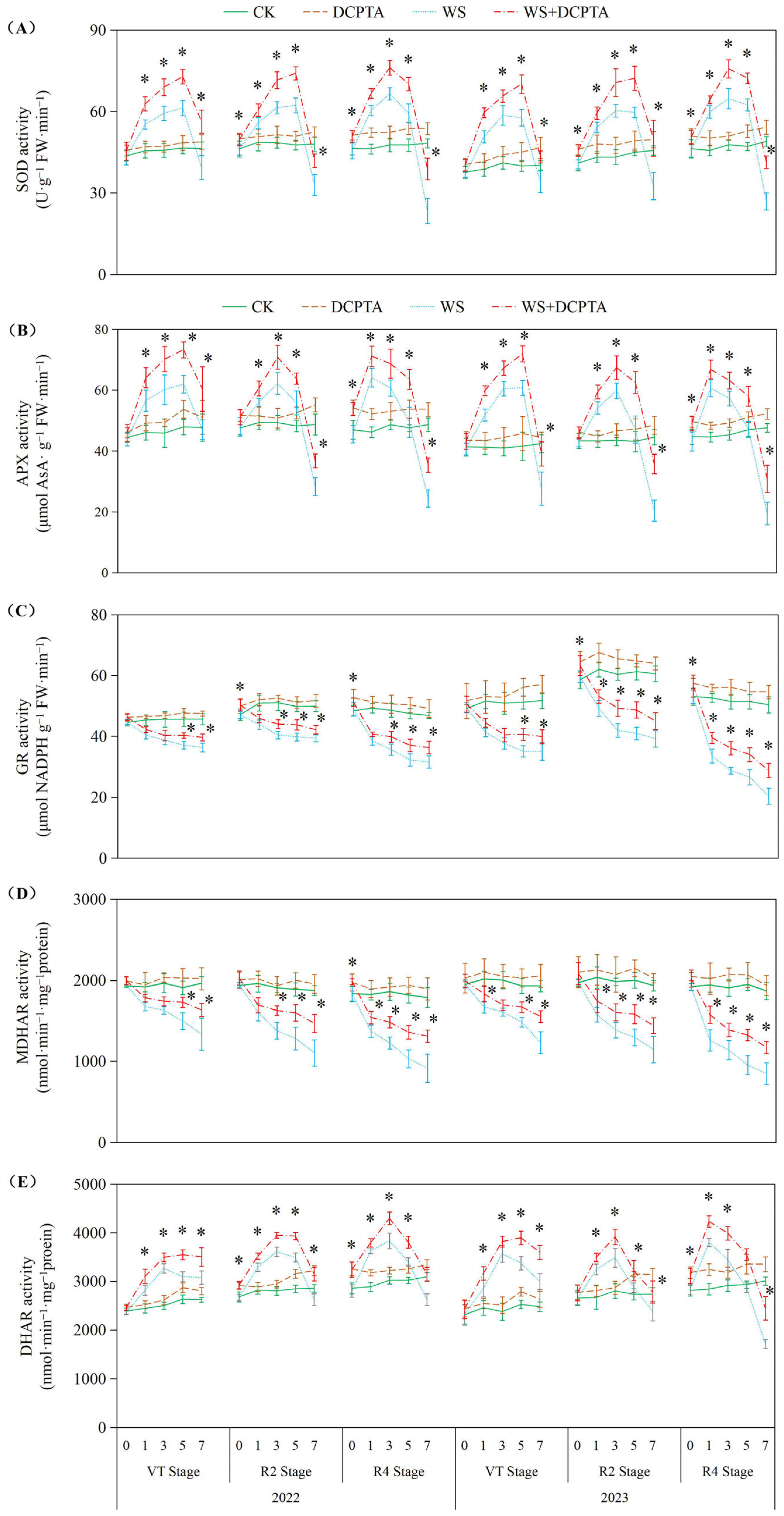
2.4. Root Antioxidant System
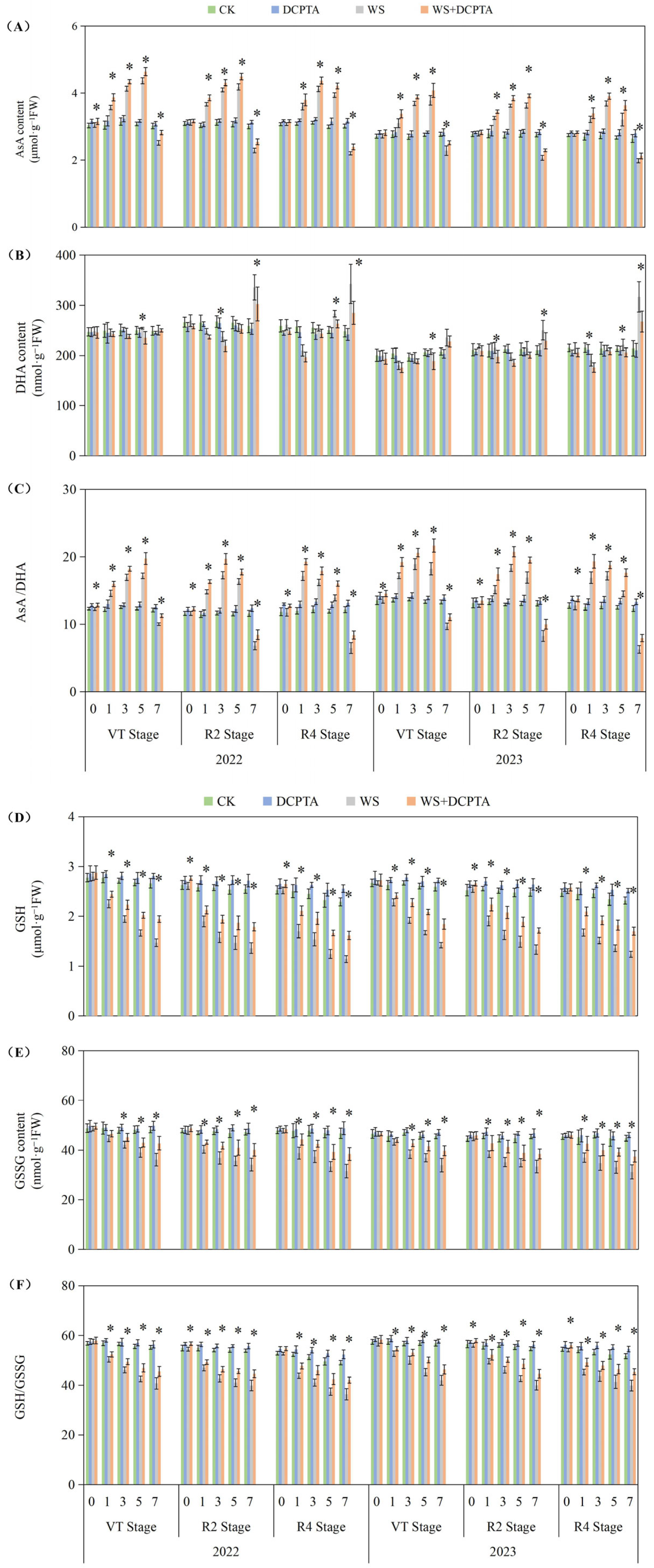
2.5. Ascorbate/Dehydroascorbate (ASA/DHA) and Reduced Glutathione/Oxidized Glutathione (GSH/GSSG) Levels
3. Discussion
3.1. Effects of Post-Tasseling Waterlogging on the Yield of Spring Maize and Exogenous DCPTA Regulating Effect
3.2. Effects of Post-Tasseling Waterlogging on the Leaf Photosynthesis of Spring Maize and Exogenous DCPTA Regulating Effect
3.3. Effects of Post-Tasseling Waterlogging on the Root Antioxidant System of Spring Maize and Exogenous DCPTA Regulating Effect
4. Materials and Methods
4.1. Plant Materials and Experimental Design
- (1)
- Control group (non-waterlogged): Plants received water (10 mL·plant−1) at the six-leaf stage, and were not subjected to waterlogging at any growth stage (VT, R2, R4).
- (2)
- DCPTA treatments: Plants were foliar sprayed with 10 mL·plant−1 DCPTA solution (35 mg L−1, determined based on previous concentration screening) at the six-leaf stage.
- (3)
- Waterlogged treatments: Separate groups of plants were exposed to waterlogging at VT/R2/R4 stages, maintained with flooded water to a level of 4~5 cm above the soil surface for 7 days continuously.
4.2. Yeild and Yield Components
4.3. Agronomic Characters
4.4. Root Activity, Root Hydraulic Conductivity and Flow Rate of the Root-Bleeding Sap
4.5. ROS Damage Characters
4.6. Antioxidant Enzyme Activities
4.7. AsA/DHA and GSH/GSSG in Roots
4.8. Chlorophyll Content
4.9. Chlorophyll Fluorescence Parameters and Gas Exchange
4.10. Gas Exchange
4.11. Carbon Metabolism-Related Enzyme Activity
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ren, B.; Zhang, J.; Dong, S.; Liu, P.; Zhao, B. Responses of carbon metabolism and antioxidant system of summer maize to waterlogging at different stages. J. Agron. Crop Sci. 2018, 204, 505–514. [Google Scholar] [CrossRef]
- Yu, X.; Ma, Y. Spatial and temporal analysis of extreme climate events over northeast China. Atmosphere 2022, 13, 1197. [Google Scholar] [CrossRef]
- Yan, D.; Gao, Y.; Zhang, Y.; Li, D.; Dirk, L.M.A.; Bruce, D.A.; Zhao, T. Raffinose catabolism enhances maize waterlogging tolerance by stimulating adventitious root growth and development. J. Exp. Bot. 2024, 75, 5955–5970. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Cheng, H.; Qiao, C.; Feng, J.; Yan, P.; Yang, R.; Song, J.; Sun, J.; Zhao, Y.; Zhang, Z. Root-zone oxygen supply mitigates waterlogging stress in tomato by enhancing root growth, photosynthetic performance, and antioxidant capacity. Plant Physiol. Biochem. 2025, 222, 109744. [Google Scholar] [CrossRef]
- Yamane, K.; Mariyama, M.; Hirooka, Y.; Iijima, M. Root pruning is effective in alleviating the inhibition of soybean growth caused by anaerobic stress for a short period. J. Integr. Agric. 2023, 22, 1035–1044. [Google Scholar] [CrossRef]
- Zeng, R.; Chen, L.; Wang, X.; Cao, J.; Li, X.; Xu, X.; Xia, Q.; Chen, T.; Zhang, L. Effect of waterlogging stress on dry matter accumulation, photosynthesis characteristics, yield, and yield components in three different ecotypes of peanut (Arachis hypogaea L.). Agronomy 2020, 10, 1244. [Google Scholar] [CrossRef]
- Zhang, R.; Yue, Z.; Chen, X.; Huang, R.; Zhou, Y.; Cao, X. Effects of waterlogging at different growth stages on the photosynthetic characteristics and grain yield of sorghum (Sorghum bicolor L.). Sci. Rep. 2023, 13, 7212. [Google Scholar] [CrossRef]
- Sharmin, R.A.; Karikari, B.; Bhuiyan, M.R.; Kong, K.; Yu, Z.; Zhang, C.; Zhao, T. Comparative morpho–physiological, biochemical, and gene expressional analyses uncover mechanisms of waterlogging tolerance in two soybean introgression lines. Plants 2024, 13, 1011. [Google Scholar] [CrossRef]
- Kaur, G.; Vikal, Y.; Kaur, L.; Kalia, A.; Mittal, A.; Kaur, D.; Yadav, I. Elucidating the morpho–physiological adaptation and molecular responses under long–term waterlogging stress in maize through gene expression analysis. Plant Sci. 2021, 304, 110823. [Google Scholar] [CrossRef]
- Liang, K.; Tang, K.; Fang, T.; Qiu, F. Waterlogging tolerance in maize: Genetic and molecular basis. Mol. Breed. 2020, 40, 111. [Google Scholar] [CrossRef]
- Yang, H.; Cai, X.; Lu, D. Effects of Waterlogging at flowering stage on the grain yield and starch quality of waxy maize. Plants 2023, 13, 108. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Yu, W.; Liu, P.; Zhao, B.; Zhang, J.; Ren, B. Responses of canopy functionality, crop growth and grain yield of summer maize to shading, waterlogging, and their combination stress at different crop stages. Eur. J. Agron. 2023, 144, 126761. [Google Scholar] [CrossRef]
- Huang, C.; Gao, Y.; Qin, A.; Liu, Z.; Zhao, B.; Ning, D.; Ma, S.; Duan, A.; Liu, Z. Effects of waterlogging at different stages and durations on maize growth and grain yields. Agric. Water Manag. 2022, 261, 107334. [Google Scholar] [CrossRef]
- Tian, L.; Li, J.; Bi, W.; Zuo, S.; Li, L.; Li, W.; Sun, L. Effects of waterlogging stress at different growth stages on the photosynthetic characteristics and grain yield of spring maize (Zea mays L.) under field conditions. Agric. Water Manag. 2019, 218, 250–258. [Google Scholar] [CrossRef]
- Hu, J.; Ren, B.; Dong, S.; Liu, P.; Zhao, B.; Zhang, J. 6-Benzyladenine increasing subsequent waterlogging-induced waterlogging tolerance of summer maize by increasing hormone signal transduction. Ann. N. Y. Acad. Sci. 2021, 1509, 89–112. [Google Scholar] [CrossRef]
- Wang, X.; Wang, Q.; Zhang, M.; Zhao, Y.; Dong, P.; Zhao, Y.; Li, H.; Jia, X.; An, P.; Tang, Y.; et al. Foliar application of spermidine alleviates waterlogging–induced damages to maize seedlings by enhancing antioxidative capacity, modulating polyamines and ethylene biosynthesis. Life 2022, 12, 1921. [Google Scholar] [CrossRef]
- Keithly, J.H.; Yokoyama, H.; Gausman, H.W. Effect of 2–(3,4–dichlorophenoxy) triethylamine (DCPTA) on the growth and development of sugarbeet. Plant Sci. 1990, 68, 57–64. [Google Scholar] [CrossRef]
- Gausman, H.W.; Burd, J.D.; Quisenberry, J.; Yokoyama, H.; Dilbeck, R.; Benedict, C.R. Effect of 2–diethylaminoethyl–3,4–dichlorophenylether (DCPTA) on cotton plant (Gossypium Hirsutum) growth and phenology. Nat. Biotechnol. 1985, 3, 255–257. [Google Scholar] [CrossRef]
- Li, L.; Gu, W.; Li, C.; Li, W.; Wei, S. Dual application of ethephon and DCPTA increases maize yield and stalk strength. Agron. J. 2019, 111, 612–627. [Google Scholar] [CrossRef]
- Gao, Y.; Zhang, X.; Wang, X.; Zhang, Q.; Tang, H.; Qiu, T.; Zhang, H.; Zhao, B.; Wang, H.; Liang, X.; et al. Exogenous DCPTA treatment increases mung bean yield by improving carbon metabolism pathway and up–regulating photosynthetic capacity and antioxidants. Front. Plant Sci. 2022, 13, 796694. [Google Scholar] [CrossRef]
- Xie, T.; Gu, W.; Meng, Y.; Li, J.; Li, L.; Wang, Y.; Qu, D.; Wei, S. Exogenous DCPTA ameliorates simulated drought conditions by improving the growth and photosynthetic capacity of maize seedlings. Sci. Rep. 2017, 7, 12684. [Google Scholar] [CrossRef] [PubMed]
- Xie, T.; Gu, W.; Zhang, L.; Li, L.; Qu, D.; Li, C.; Meng, Y.; Li, J.; Wei, S.; Li, W. Modulating the antioxidant system by exogenous 2–(3,4–dichlorophenoxy) triethylamine in maize seedlings exposed to polyethylene glycol–simulated drought stress. PLoS ONE 2018, 13, e0203626. [Google Scholar] [CrossRef] [PubMed]
- Xie, T.; Gu, W.; Li, C.; Li, J.; Wei, S. Exogenous DCPTA increases the tolerance of maize seedlings to PEG–simulated drought by regulating nitrogen metabolism–related enzymes. Agronomy 2019, 9, 676. [Google Scholar] [CrossRef]
- Li, L.; Gu, W.; Zhang, L.; Li, C.; Chen, X.; Qian, C.; Wang, Z.; Li, W.; Zuo, S.; Wei, S. Exogenous 2–(3,4–Dichlorophenoxy) triethylamine alleviates salinity stress in maize by enhancing photosynthetic capacity, improving water status and maintaining K+/Na+ homeostasis. BMC Plant Biol. 2020, 20, 348. [Google Scholar] [CrossRef]
- Xie, T.; Gu, W.; Wang, M.; Zhang, L.; Li, C.; Li, C.; Li, W.; Li, L.; Wei, S. Exogenous 2–(3,4–Dichlorophenoxy) triethylamine ameliorates the soil drought effect on nitrogen metabolism in maize during the pre–female inflorescence emergence stage. BMC Plant Biol. 2019, 19, 107. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, P.; Wang, R.; Wang, P.; Huang, S. Effects of variety and chemical regulators on cold tolerance during maize germination. J. Integr. Agric. 2018, 17, 66–73. [Google Scholar] [CrossRef]
- Asha, S.; Sultana, N.; Hassan, L.; Akhter, S.; Robin, A.H.K. Response of morphological and biochemical traits of maize genotypes under waterlogging stress. J. Phytol. 2021, 13, 108–121. [Google Scholar] [CrossRef]
- Ren, B.; Zhang, J.; Dong, S.; Liu, P.; Zhao, B. Root and shoot responses of summer maize to waterlogging at different stages. Agron. J. 2016, 108, 1060–1069. [Google Scholar] [CrossRef]
- Qiu, T.; Tang, H.; Zhang, H.; Wang, X.; Yang, J.; Li, H.; Zhang, X.; Zhang, Y.; Zheng, S.; Gao, Y. Effects of Different Plant Growth Regulators on Agronomic Traits and Photosynthetic Performance of Mung Bean. Am. J. Plant Biol. 2021, 6, 39–43. [Google Scholar] [CrossRef]
- Keithly, J.H.; Yokoyama, H.; Gausman, H. Enhanced yield of tomato in response to 2–(3,4–dichlorophenoxy) triethylamine (DCPTA). Plant Growth Regul. 1990, 9, 127–136. [Google Scholar] [CrossRef]
- Hsu, W.J.; Mendoza, K. Effect of 2–(3,4–dichlorophenoxy) triethylamine on guayule (Parthenium argentatum) cell suspension cultures. J. Agric. Food Chem. 1992, 40, 924–926. [Google Scholar] [CrossRef]
- Benedict, C.R.; Rosenfield, C.L.; Mahan, J.R.; Madhavan, S.; Yokoyama, H. The chemical regulation of carotenoid biosynthesis in citrus. Plant Sci. 1985, 41, 169–173. [Google Scholar] [CrossRef]
- Hu, J.; Ren, B.; Dong, S.; Liu, P.; Zhao, B.; Zhang, J. Poor development of spike differentiation triggered by lower photosynthesis and carbon partitioning reduces summer maize yield after waterlogging. Crop J. 2021, 2, 478–489. [Google Scholar] [CrossRef]
- Yang, H.; Wen, Z.; Huang, T.; Lu, W.; Lu, D. Effects of waterlogging at grain formation stage on starch structure and functionality of waxy maize. Food Chem. 2019, 294, 187–193. [Google Scholar] [CrossRef]
- Shen, S.; Liang, X.; Zhang, L.; Zhao, X.; Liu, Y.; Lin, S.; Gao, Z.; Wang, P.; Wang, Z.; Zhou, S. Intervening in sibling competition for assimilates by controlled pollination prevents seed abortion under postpollination drought in maize. Plant Cell Environ. 2020, 43, 903–919. [Google Scholar] [CrossRef]
- Ren, B.; Yu, W.; Liu, P.; Zhao, B.; Zhang, J. Responses of photosynthetic characteristics and leaf senescence in summer maize to simultaneous stresses of waterlogging and shading. Crop J. 2023, 1, 269–277. [Google Scholar] [CrossRef]
- Li, W.; Mo, W.; Ashraf, U.; Li, G.; Wen, T.; Abrar, M.; Gao, L.; Liu, J.; Hu, J. Evaluation of physiological indices of waterlogging tolerance of different maize varieties in South China. Appl. Ecol. Environ. Res. 2018, 16, 2059–2072. [Google Scholar] [CrossRef]
- Zhang, X.; Huang, C.; Meng, Y.; Liu, X.; Gao, Y.; Liu, Z.; Ma, S. Physiological mechanism of waterlogging stress on yield of waxy maize at the jointing stage. Plants 2023, 12, 3034. [Google Scholar] [CrossRef]
- Drew, B.M.C. Stomatal and nonstomatal components to inhibition of photosynthesis in leaves of capsicum annuum during progressive exposure to NaCl salinity. Plant Physiol. 1992, 99, 219–226. [Google Scholar]
- Kromdijk, J.; Glowacka, K.; Leonelli, L.; Gabilly, S.T.; Iwai, M.; Niyogi, K.K.; Long, S.P. Improving photosynthesis and crop productivity by accelerating recovery from photoprotection. Science 2016, 354, 857. [Google Scholar] [CrossRef]
- Shabala, S. Physiological and cellular aspects of phytotoxicity tolerance in plants: The role of membrane transporters and implications for crop breeding for waterlogging tolerance. New Phytol. 2011, 190, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Bian, X.; Wei, T.; Yan, J.; Sun, X.; Han, T.; Dong, B.; Zhang, G.; Li, J.; Zhang, A. ZmMPK5 phosphorylates ZmNAC49 to enhance oxidative stress tolerance in maize. New Phytol. 2021, 232, 2400–2417. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Xu, S.; Zou, X.; Zheng, Y.; Qiu, F. Changes of antioxidative enzymes and lipid peroxidation in leaves and roots of waterlogging–tolerant and waterlogging–sensitive maize genotypes at seedling stage. Agric. Sci. China 2010, 9, 651–661. [Google Scholar] [CrossRef]
- Li, S. Novel insight into functions of ascorbate peroxidase in plants: More than a simple antioxidant enzyme. Redox Biol. 2023, 64, 102789. [Google Scholar] [CrossRef]
- Soares, C.; Carvalho, M.E.A.; Azevedo, R.A.; Fidalgo, F. Plants facing oxidative challenges–A little help from the antioxidant networks. Environ. Exp. Bot. 2018, 161, 4–25. [Google Scholar] [CrossRef]
- Ushimaru, T.; Shibasaka, M.; Tsuji, H. Development of the O2·− detoxification system during adaptation to air of submerged rice seedlings. Plant Cell Biol. 1992, 33, 1065–1071. [Google Scholar]
- Biemelt, S.; Keetman, U.; Albrecht, G. Re–aeration following hypoxia or anoxia leads to activation of the antioxidative defense system in roots of wheat seedlings. Plant Physiol. 1998, 116, 651–658. [Google Scholar] [CrossRef]
- Skutnik, M.; Rychter, A.M. Differential response of antioxidant systems in leaves and roots of barley subjected to anoxia and post–anoxia. J. Plant Physiol. 2009, 166, 926–937. [Google Scholar] [CrossRef]
- Wu, Q.; Yang, J.; Cheng, N.; Hirschi, K.D.; White, F.F.; Park, S. Glutaredoxins in plant development, abiotic stress response, and iron homeostasis: From model organisms to crops. Environ. Exp. Bot. 2017, 139, 91–98. [Google Scholar] [CrossRef]
- Wang, J.; Yu, Y.; Zhang, Z.; Quan, R.; Zhang, H.; Ma, L.; Deng, X.; Huang, R. Arabidopsis CSN5B interacts with VTC1 and modulates ascorbic acid synthesis. Plant Cell 2013, 25, 625–636. [Google Scholar] [CrossRef]
- Hatano–Iwasaki, A.; Ogawa, K. Overexpression of GSH1 gene mimics transcriptional response to low temperature during seed vernalization treatment of Arabidopsis. Plant Cell Physiol. 2012, 53, 1195–1203. [Google Scholar] [CrossRef] [PubMed]
- Cottenie, A.; Verloo, M.; Kiekens, L.; Velghe, G.; Camerlynck, R. Chemical analysis of plant and soil. Chem. Anal. Plants Soils 1982, 63, 44–45. [Google Scholar]
- Niu, L.; Yan, Y.; Hou, P.; Bai, W.; Zhao, R.; Wang, Y.; Li, S.; Du, T.; Zhao, M.; Song, J.; et al. Influence of plastic film mulching and planting density on yield, leaf anatomy, and root characteristics of maize on the Loess Plateau. Crop J. 2020, 8, 548–564. [Google Scholar] [CrossRef]
- Elstner, E.F.; Heupel, A. Inhibition of nitrite formation from hydroxylammonium chloride: A simple assay for superoxide dismutase. Anal. Biochem. 1976, 70, 616–620. [Google Scholar] [CrossRef]
- Jana, S.; Choudhuri, M.A. Glycolate metabolism of three submersed aquatic angiosperms during ageing. Aquat. Bot. 1982, 12, 345–354. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Lutts, S.; Kinet, J.M.; Bouharmont, J. Changes in plant response to nacl during development of rice (Oryza sativa L.) varieties differing in salinity resistance. J. Exp. Bot. 1995, 46, 1843–1852. [Google Scholar] [CrossRef]
- Ni, L.; Fu, X.; Zhang, H.; Li, X.; Cai, X.; Zhang, P.; Liu, L.; Wang, Q.; Sun, M.; Wang, Q. Abscisic acid inhibits rice protein phosphatase PP45 via H2O2 and relieves repression of the Ca2+/CaM–dependent protein kinase DMI3. Plant Cell 2019, 31, 128–152. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Spinach chloroplasts scavenge hydrogen peroxide on illumination. Plant Cell Physiol. 1980, 21, 1295–1307. [Google Scholar] [CrossRef]
- Foyer, C.H.; Halliwell, B. Purification and properties of dehydroascorbate reductase from spinach leaves. Phytochemistry 1977, 16, 1347–1350. [Google Scholar] [CrossRef]
- Pyngrope, S.; Bhoomika, K.; Dubey, R.S. Reactive oxygen species, ascorbate–glutathione pool, and enzymes of their metabolism in drought–sensitive and tolerant indica rice (Oryza sativa L.) seedlings subjected to progressing levels of water deficit. Protoplasma 2013, 250, 585–600. [Google Scholar] [CrossRef] [PubMed]
- Tai, F.; Wang, S.; Liang, B.; Li, Y.; Wu, J.; Fan, C.; Hu, X.; Wang, H.; He, R.; Wang, W. Quaternary ammonium iminofullerenes improve root growth of oxidative–stress maize through ASA–GSH cycle modulating redox homeostasis of roots and ROS–mediated root–hair elongation. J. Nanobiotechnol. 2022, 20, 15. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence–A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Stage | Treatment | Number of Ears (ears·mu–1) | Number of Kernels (kernels·ear–1) | 1000-Kernel Weight (g) | Yield (kg·mu–1) |
|---|---|---|---|---|---|---|
| 2022 | / | CK | 4661 ± 84 | 574 ± 17 | 316.33 ± 7.41 | 846.27 ± 22.80 |
| CK + DCPTA | 4691 ± 108 | 587 ± 10 | 325.16 ± 4.50 | 896.41 ± 35.47 | ||
| VT | Waterlogging | 4309 ± 47 | 499 ± 15 | 300.70 ± 3.51 | 646.50 ± 16.49 | |
| Waterlogging + DCPTA | 4436 ± 61 * | 545 ± 16 * | 312.80 ± 6.01 * | 756.53 ± 29.47 * | ||
| R2 | Waterlogging | 4411 ± 62 | 539 ± 18 | 288.47 ± 5.90 | 685.28 ± 19.81 | |
| Waterlogging + DCPTA | 4559 ± 84 * | 558 ± 18 | 307.53 ± 8.29 * | 782.11 ± 34.94 * | ||
| R4 | Waterlogging | 4494 ± 57 | 518 ± 20 | 262.24 ± 9.22 | 610.91 ± 33.23 | |
| Waterlogging + DCPTA | 4595 ± 49 | 533 ± 21 | 282.42 ± 4.96 * | 691.70 ± 39.88 * | ||
| 2023 | / | CK | 4554 ± 81 | 558 ± 17 | 309.73 ± 7.72 | 786.01 ± 16.82 |
| CK + DCPTA | 4611 ± 88 | 572 ± 8 | 319.22 ± 3.98 | 842.85 ± 35.29 | ||
| VT | Waterlogging | 4180 ± 58 | 475 ± 25 | 292.74 ± 4.60 | 581.36 ± 30.76 | |
| Waterlogging + DCPTA | 4314 ± 57 * | 537 ± 17 * | 305.40 ± 6.82 * | 707.55 ± 36.48 * | ||
| R2 | Waterlogging | 4273 ± 50 | 515 ± 12 | 277.92 ± 8.28 | 612.08 ± 23.75 | |
| Waterlogging + DCPTA | 4437 ± 104 * | 544 ± 18 * | 299.73 ± 10.28 * | 723.54 ± 46.36 * | ||
| R4 | Waterlogging | 4425 ± 69 | 500 ± 17 | 251.84 ± 8.33 | 557.25 ± 34.74 | |
| Waterlogging + DCPTA | 4473 ± 44 | 524 ± 19 * | 280.62 ± 7.08 * | 657.84 ± 31.19 * |
| Year | pH | HCO3− + CO32− | Cl− | SO42− | Ca2+ | Mg2+ | Na+ | K+ | N | P |
|---|---|---|---|---|---|---|---|---|---|---|
| 2022 | 7.2 | 205.3 | 284.6 | 472.9 | 86.9 | 38.5 | 4.2 | 29.7 | 16.7 | 3.6 |
| 2023 | 7.1 | 202.7 | 302.5 | 445.7 | 91.2 | 41.3 | 4.1 | 31.2 | 14.6 | 3.9 |
| Treatment | Foliar Spray (Six-Leaf Stage) | Waterlogging Stage | Waterlogging Duration |
|---|---|---|---|
| Control | Water | None | 0 days |
| DCPTA | DCPTA (35 mg·L−1) | None | 0 days |
| Waterlogging + Water | Water | VT, R2, or R4 | 7 days |
| Waterlogging + DCPTA | DCPTA (35 mg·L−1) | VT, R2, or R4 | 7 days |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, T.; Mei, L.; Yang, X.-G.; Wang, M.; Zhang, Q.; Li, W.; Zhang, H.; Zhang, M.; Yang, D.; Dou, J.; et al. 2-(3,4-Dichlorophenoxy)triethylamine (DCPTA) Sustains Root Activity Through the Enhancement of Ascorbate-Glutathione in Spring Maize (Zea mays L.) Under Post-Tasseling Waterlogging. Int. J. Mol. Sci. 2025, 26, 3698. https://doi.org/10.3390/ijms26083698
Xie T, Mei L, Yang X-G, Wang M, Zhang Q, Li W, Zhang H, Zhang M, Yang D, Dou J, et al. 2-(3,4-Dichlorophenoxy)triethylamine (DCPTA) Sustains Root Activity Through the Enhancement of Ascorbate-Glutathione in Spring Maize (Zea mays L.) Under Post-Tasseling Waterlogging. International Journal of Molecular Sciences. 2025; 26(8):3698. https://doi.org/10.3390/ijms26083698
Chicago/Turabian StyleXie, Tenglong, Linlin Mei, Xiao-Ge Yang, Meiyu Wang, Qian Zhang, Wei Li, He Zhang, Meng Zhang, Deguang Yang, Jingjie Dou, and et al. 2025. "2-(3,4-Dichlorophenoxy)triethylamine (DCPTA) Sustains Root Activity Through the Enhancement of Ascorbate-Glutathione in Spring Maize (Zea mays L.) Under Post-Tasseling Waterlogging" International Journal of Molecular Sciences 26, no. 8: 3698. https://doi.org/10.3390/ijms26083698
APA StyleXie, T., Mei, L., Yang, X.-G., Wang, M., Zhang, Q., Li, W., Zhang, H., Zhang, M., Yang, D., Dou, J., & Yang, X. (2025). 2-(3,4-Dichlorophenoxy)triethylamine (DCPTA) Sustains Root Activity Through the Enhancement of Ascorbate-Glutathione in Spring Maize (Zea mays L.) Under Post-Tasseling Waterlogging. International Journal of Molecular Sciences, 26(8), 3698. https://doi.org/10.3390/ijms26083698