
The Role of Killer Ig-like Receptors in Diseases from A to Z





Abstract
1. Introduction
2. Natural Killer Cells
3. Killer Ig-like Receptors
3.1. Biological Function
3.2. KIRs and HLA Interaction
4. KIR Genes in Autoimmune Diseases
4.1. Ankylosing Spondylitis
4.2. Atopic Dermatitis
4.3. Autoimmune Hepatitis
4.4. Behçet’s Disease
4.5. Celiac Disease
4.6. Graves’ Disease
4.7. Hashimoto’s Thyroiditis
4.8. Immune Thrombocytopenia
4.9. Inflammatory Bowel Disease: Crohn’s Disease and Ulcerative Colitis
4.10. Juvenile Idiopathic Arthritis
4.11. Myasthenia Gravis
4.12. Multiple Sclerosis
4.13. Non-Celiac Wheat Sensitivity
4.14. Psoriatic Arthritis
4.15. Psoriasis Vulgaris
4.16. Rheumatoid Arthritis
4.17. Sjögren’s Syndrome
4.18. Systemic Lupus Erythematosus
4.19. Systemic Sclerosis
4.20. Type 1 Diabetes Mellitus
4.21. Vogt–Koyanagi–Harada Syndrome

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Predisposing KIRs | Protective KIRs | |
|---|---|---|
| Ankylosing spondylitis | KIR3DS1 [26,27] KIR2DS1 [27] KIR2DS4 [26] KIR2DS5 [26,27] KIR2DL5 [26,27] KIR3DL1 [26] | KIR2DL2 [27] KIR2DS2 [27] |
| Atopic dermatitis | KIR2DS1 [31,32] KIR2DL5 [31,32] KIR2DS5 [31,32] KIR2DL4 [32] KIR2DS4 [32] | KIR2DS1 [30] |
| Autoimmune hepatitis | KIR2DS1 [37] KIR2DS4 [38] KIR3DL1 with HLA-B Bw4-80Ile [39] | KIR2DL3 [37] KIR3DL1 with HLA-B Bw4-80Thr [37] KIR2DL1 with HLA-C2 [39] KIR2DL2 [38] KIR3DL1 [39] |
| Behçet’s disease | KIR3DS1 [45] | KIR3DL1 [46] |
| Celiac disease | KIR2DL5B [50,52] KIR3DL1 [51] KIR2DL2 with HLA-C1 [52] KIR2DS2 [52] KIR2DS3 [52] KIR2DL3 with HLA-C1 [53] KIR2DS5 [54] KIR3DS1 [54] | \ |
| Graves’ Disease | KIR2DS2-, KIR2DL2-, KIR2DL3+, KIR2DL1+, KIR3DL1+, KIR3DS1-, KIR2DL5-, KIR2DS3-, KIR2DS5-, KIR2DS1-, KIR2DS4- [57] | KIR2DL1 with HLA-C2 [60] |
| Hashimoto’s thyroiditis | KIR2DS2 with HLA-C1 [63] | KIR2DS2-/KIR2DL2+/KIR2DL3+/HLA-C1 [63] |
| Immune thrombocytopenia | KIR2DS2 [65,66,67] KIR2DS3 [67] KIR2DL2 [65,66] | KIR2DS5 [67] KIR2DL3 [64] KIR3DL2 [64] KIR3DL1 [64] |
| Inflammatory bowel disease | KIR2DS2 [68] KIR2DL2 [68] KIR3DL1 with HLA-Bw4 [69] KIR2DL3 with HLA-C1 [70] KIR2DS3 [69] KIR2DL5 [71,73] KIR2DS1 [71,73] | KIR2DS3 [71] KIR2DL3 with HLA-Cw1 [68] KIR2DL2 [70] KIR2DS2 [70] |
| Juvenile Idiopathic Arthritis | KIR3DL1/KIR2DL2 [75] | KIR2DS4 [74] |
| Myasthenia gravis | \ | \ |
| Multiple Sclerosis | KIR3DS1 [80] KIR2DL5 [80] KIR2DL1 [80] KIR2DS5 [80] KIR2DL2/KIR2DS2 [84] | KIR2DS1 [86] KIR2DL3 [84] KIR3DL1 with HLA-Bw4 [85] KIR2DL2/KIR2DL3 with HLA-Bw4 [87] |
| Non-celiac wheat sensitivity | / | KIR2DL5 [89] KIR2DS4 [89] KIR2DS5 [87] |
| Psoriatic Arthritis | KIR2DL1 [9] KIR2DL2 [4,5,6,7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90] KIR2DS1 [90,91,92,93,94,95] KIR2DS2 [90,91,92,93,94,95] KIR2DS3 [90,91,92,93,94,95] | \ |
| Psoriasis Vulgaris | KIR2DS1 [102] KIR2DS5 [102] KIR3DS1 [102] | KIR2DS4 [102] KIR3DL1 [102] KIR2DL1 [102] |
| Rheumatoid arthritis | KIR2DS2 [103,104] KIR2DL2 [104,109] KIR2DS4 [104,109] KIR2DL1 [105] KIR2DS1 [105] | KIR2DL3 [104,105,106,109] KIR2DL5A [104,109] KIR2DL5 [106] KIR2DS5 [106] KIR3DL3 [106] KIR2DL2/KIR2DS2 (treatment) [107] |
| Sjögren’s syndrome | KIR2DS2+/KIR2DL2- [111] | \ |
| Systemic Lupus Erythematosus | KIR2DS1 [112,113,114,115,116] KIR2DS2 [112,113,118] with HLA-C1 [120] KIR2DL3 [119] KIR3DL1 [119] KIR2DL2 [115,116] KIR3DS1 [119] KIR2DL5B [120] | KIR2DL3 with HLA-C1 [119,120] KIR2DL5 [112] with HLA-Bw4 [119] KIR2DL5B with HLA-C1 [120] KIR3DL1 [119] KIR2DL2 with HLA-C1 [120] KIR2DS2 with HLA-C1 [120] |
| Systemic sclerosis | KIR2DS2+/KIR2DL2- [118,122,123] KIR2DS3 [118,122,123] KIR3DL1 [124] KIR2DS2 [122,123] KIR2DL2 with HLA-C1/C2 [125] KIR2DS4del [125] | KIR2DS4full [125] |
| Type 1 Diabetes Mellitus | KIR2DL2 [129,130,134] KIR3DL1 [134] KIR2DS4 [134] KIR2DL5 [131] KIR2DS2 [131] KIR2DL1 [131] KIR3DL1 [131] KIR2DS4 [131] | KIR2DL1 [133,134,135] KIR2DL5 [134] KIR3DL1 with HLA-C1C2 [135] KIR2DL3 with HLA-C1C2 [135] KIR2DS2 with HLA-C1C2 [135] KIR2DS1 with HLA-C1C2 [133] |
| Vogt–Koyanagi–Harada syndrome | KIR3DS1 [136] without KIR3DL1 [136] KIR2DS1 [136] without KIR3DL1 [136] KIR2DS2 [136,139] KIR2DS3 [136,138,139] KIR2DS5 without KIR3DL1 [137] KIR2DL2 [139] KIR2DL5B [139] | KIR3DL1 [137] KIR2DL2/KIR2DL3 with HLA-C1 [138] |
5. KIR Genes and Cancer
5.1. Biliary Tract Cancer
5.2. Bladder Cancer
5.3. Breast Cancer
5.4. Cervical Neoplasia
5.5. Colorectal Cancer and Metastatic Colorectal Cancer
5.6. Dermal Neurofibroma
5.7. Hepatocellular Carcinoma
5.8. Kidney Cancer
5.9. Leukemia
5.10. Lung Cancer
5.11. Melanoma
5.12. Multiple Myeloma
5.13. Mycosis Fungoides
5.14. Neuroblastoma
5.15. Non-Hodgkin Lymphoma
5.16. Non-Melanoma Skin Tumors
5.17. Ovarian Cancer
5.18. Thyroid Cancer
| Predisposing KIRs | Protective KIRs | |
|---|---|---|
| Biliary tract Cancers | KIR2DL2 [145] KIR3DS1 [145] | KIR2DL3 [145] |
| Bladder Cancer | KIR2DL1 [147,148] KIR2DS4 [147] KIR2DL5 [148] | KIR2DL2 [147] KIR2DS2 [147] |
| Breast Cancer | KIR-Bx genotype [150,154] KIR2DS1 [150,154,155] KIR3DS1 [150,154] KIR2DS2 [159] KIR2DL2 [153,154] KIR2DL5 [154] KIR2DS5 [154] | KIR2DS2 [151] KIR2DS3 [151] KIR2DL5A [151] KIR2DL2 without HLA-C1 [156] KIR2DL3 without HLA-C1 [151] KIR2DL1 [152,154] KIR2DS4 (alleles 2DS4 003/4/6/7) [152,154,155] |
| Cervical neoplasia | KIR2DL1 [157] KIR2DL2 [157] KIR2DL3 [157,158] KIR2DL4 [157] KIR2DS2 [158] KIR2DS3 [158] KIR2DS4 [158] KIR2DS5 [158] KIR3DS1 [158,161] | KIR2DL5 [157] KIR3DL1 [167] KIR2DL2 [159,160] KIR2DS2 [159,160] KIR2DL1 [158] |
| Colorectal cancer and Metastatic Colorectal Cancer | KIR2DS1 [164,168] KIR2DS5 [164,166,167,168] KIR3DS1 [164,168] KIR3DS5 [165] KIR2DS3 [169] KIR2DL5 [164,168] KIR2DS4 [168] | KIR2DS4 [164,165] KIR3DL1 [164,165] KIR3DS2 [165] |
| Dermal neurofibroma | KIR2DL5 (N173D) [171] | |
| Hepatocellular carcinoma | KIR2DS4/1D [173] KIR2DS1 [176] KIR3DS1 [176] KIR2DL5 [176] KIR2DS5 [176] | KIR2DS4/1D [174] KIR2DS1 [174] KIR2DS2 [174] KIR3DS1 [174] KIR2DL3 [175] KIR AA Haplotype [176] |
| Kidney cancer | KIR3DL1 [178] | KIR2DL2 [178] KIR2DL3 [178] |
| Leukemia | KIR AB genotype [179] KIR2DL2 [179] KIR2DS4 [181] KIR3DL1 with HLA-Bw4 [180] KIR AA genotype [185] KIR2DS3 [181] | KIR2DL2 with HLA-C1 [180,181,186] KIR2DS2 with HLA-C1 [180] KIR2DS2 [182,186] KIR2DL2 [184,186] KIR2DL5 [184] KIR2DS1 [184] KIR2DS2 [179,186] KIR2DS3 [179,181,187] |
| Lung Cancer | KIR2DL2 with HLA-C1 [195] KIR2DS2 with HLA-C1 [195] KIR2DS4del [194] | KIR2DL2 [192] KIR2DS2 [192] KIR3DL1 [195] KIR3DS1 [195] |
| Melanoma | KIR3DL1 with HLA-Bw4Ile80 [198] KIR2DL2 with HLA-C1 [199] KIR BB genotype [199] KIR2DS5 [199] | KIR2DL2 with HLA-C [198] KIR2DL3 with HLA-C [198] KIR2DS1 without HLA-C [198] KIR2DL3 [197] KIR2DL3 with HLAC1 [197] KIR AA genotype [199] |
| Multiple Myeloma | KIR2DS4 [203,204] KIR2DS5 [203] KIR2DL1 [204] KIR3DL1 [205] | KIR3DL1 [204] KIR3DS4 [204] |
| Mycosis Fungoides | KIR3DL1 [207] | KIR3DL2 [207] |
| Neuroblastoma | KIR2DL2 [211] KIR2DS2 [211] KIR2DL1 [208] KIR3DL1 [208] KIR2DL3 [208] | KIR2DS3 [208,212] |
| Non-Hodgkin Lymphoma | KIR2DS1 [214] KIR2DL5 [214] KIR3DS1 [214] KIR2DL5A [214] KIR2DL5B [214] | KIR2DL3 [215] |
| Non-melanoma skin tumors | KIR2DL1 [218] KIR2DS4 [218] KIR2DS3 with HLA-C1 [219] | KIR3DL1 with HLA-Bw4 [218] |
| Ovarian cancer | KIR3DS4 [221] | / |
| Thyroid cancer | KIR3DS1 [222] KIR3DL5 [222] KIR2DS1 [222] KIR2DS5 [222] | / |
6. KIR Genes and Infectious Diseases
6.1. Cytomegalovirus
6.2. Ebola Virus Disease
6.3. Epstein–Barr Virus
6.4. Hepatitis B Virus
6.5. Hepatitis C Virus
6.6. Herpes Simplex Virus
6.7. Human Herpesvirus 8
6.8. Human Immunodeficiency Virus
6.9. Human Papillomavirus
6.10. Leprosy
6.11. Leptospirosis
6.12. Malaria
6.13. Q Fever
6.14. Sepsis
6.15. Severe Acute Respiratory Syndrome Coronavirus 2
6.16. Syphilis
6.17. Tuberculosis
| Predisposing KIRs | Protective KIRs | |
|---|---|---|
| Cytomegalovirus | KIR2DL1 [225] KIR2DL3 [225,226] with HLA-C1 [235,244] KIR3DS1 [225,226,227,240] KIR3DL1 [226] with HLA-Bw4T [229] KIR2DL2 with HLA-C1 [246] KIR2DS2 [229,232,238,244] with HLA-C1 [237] KIR2DS4 [230,238] KIR2DS5 [231,240] KIR2DS2 [231] KIR2DS1 [240] KIR2DL5A [240] | KIR2DS1 [227,230,231,236,239,241,242,243] KIR2DS5 [239,243] KIR2DL5A [239] KIR2DS3 [243] KIR1D [247] KIR2DS2 [229,232,245] KIR2DS4 [231,232,245] KIR3DS1 [239,243] |
| Ebola Virus Disease | KIR2DS1 [248] KIR2DS3 [248] KIR haplotype without KIR2DL2, KIR2DL5 KIR2DS1, KIR2DS2, KIR2DS3, KIR2DS5, and KIR3DS1 [249] KIR2DS4-003 [249] KIR2DL5 [249] | / |
| Epstein–Barr Virus | KIR2DS1 [250,251] KIR2DS5 [250] KIR3DS1 [250,251] KIR2DL5 [250] KIR2DS2 [250] KIR2DS4 [250] KIR2DL2 [250] KIR2DS3 [251] | KIR2DS1 [243] KIR2DS3 [243] KIR3DS1 [243] |
| Hepatitis B Virus | KIR2DS2 [256,260] KIR2DS3 [256,266] KIR3DL1 [227,259] KIR2DL1 [259] KIR2DL2 [260] KIR2DL3 [260] KIR2DS4 [259,261] KIR2DS5 [261,262] KIR2DP1 [261,262] KIR3DS1 [263] | KIR2DS1 [256,260,264] KIR3DS1 [256,257,258,264,265,266] KIR2DL5 [256] KIR2DL3 [257,258,261,262,266] KIR2DL2 [259,261] KIR3DL1 [260] KIR3DL2 [260] KIR2DP1 [260] KIR2DL5A [264] KIR2DL5 [264] KIR3DP1 [264] |
| Hepatitis C Virus | KIR2DL2 [269,273] KIR2DS2 [269,273] KIR2DS3 [273,277] KIR2DS5 [274] KIR2DS4 [277] KIR2DL3/KIR2DL3 [278] KIR2DL3/KIR2DL3 with HLA-C1 [278] KIR2DS1 [276] KIR3DS1 [276] KIR2DS4/KIR2DS1/KIR2DL1 with HLA-C [279] KIR2DL4 with HLA-G [280] KIR3DL2 with HLA-A [281] | KIR2DL3 [268] with HLA-C1 [267] KIR2DS5 [269] KIR2DL2/KIR2DL3/KIR2DS4 [272] KIR2DS5 [273] KIR2DS3 [274,275,276] KIR2DL2 [275] KIR2DS4 [275] KIR3DL1 [277] KIR2DL5A-/KIR2DL5B+ [278] |
| Herpes Simplex Virus | KIR2DL2 with HLA-C [282,283] KIR2DS2 [282,283] | KIR2DS4del with HLA-Bw4 [284] |
| Human Herpesvirus 8 | KIR2DS1 [286] with HLA-C [286] KIR3DS1 [286] KIR2DL2/KIR2DS2 [287] KIR2DL3 with HLA-C1 [287] KIR2DL2 [277] with HLA-C1 [287] KIR3DS1 with HLA-Bw4-80I [289] KIR2DL3 [290] | KIR2DL3 [287,289] with HLA-C1 [287] KIR2DL2 [289] with HLA-C1 [287] KIR3DS1 with HLA-Bw4-80I [287] |
| Human Immunodeficiency Virus | KIR3DS1 [291,306,308] KIR2DS2 [300,306] KIR3DS1 [291] KIR2DL5 [304,305] KIR2DL2 [304,305,306] KIR2DS1 [304] KIR2DS4 [305,306] KIR3DL1 [305] KIR2DL3 [305] KIR2DS3 [306] KIR2DL3 with HLA-C1C2 [307] | KIR3DS1 [292,294,305] with HLA-B Bw480I [291,299] KIR3DL1 [306] with HLA-B [291,292,293,294,295,296] KIR3DS1/KIR3DL1 [298] KIR2DL3 with HLA-C1 [307] KIR2DL3 with HLA-C1C2 [307] KIR2DL3 [304,305,309] KIR2DL2 [307,309] KIR2DL5 [305,308] KIR2DS3 [305] KIR2DL1 [305,308] KIR2DL2/KIR2DL3 [307] KIR2DS1 [208] KIR2DS5 [208] |
| Human Papillomavirus | KIR2DS5 [140] KIR2DL2 with HLA-C1 [315] KIR2DL3 with HLA-C1 [315] | KIR3DS1 [310] KIR2DS1 [310] KIR3DL2 with HLA-A3/11 [310] KIR2DL1 [158] KIR2DL5B [158] KIR2DL3 [158] |
| Leprosy | KIR2DL2 [317,318] KIR2DL3 [318] KIR3DL2 with its ligand [318] | KIR2DS3 [317] KIR2DS2 [317,318] KIR2DS1 [318] KIR3DS1 [318] |
| Leptospirosis | KIR2DL3 [320] KIR2DL5B [320] KIR2DS1 [320] KIR2DS5 [320] | / |
| Malaria | KIR3DL1 [322,324] with HLA-Bw4 [329] KIR3DS1 [322,326] KIR2DS4 [322] KIR2DL3 [328] with HLA-C1 [325] KIR2DS2 [326,328] with HLA-C1 [326] KIR2DL2 with HLA-C1 [326] KIR2DS5 [326] KIR2DL1 [328] with HLA-C2 [329] | KIR2DS5 [327] KIR2DS3 [327] |
| Q fever | / | / |
| Sepsis | / | KIR2DS1 [335] KIR3DS1 [335] |
| Severe Acute Respiratory Syndrome Coronavirus 2 | KIR2DL1 [338] with HLA-C2 [338] KIR2DL3 [338,339] KIR2DS4 [338,341,342,343] KIR3DL1/KIR3DL2 [339] KIR3DL1 [342] KIR2DL2 [345] with HLA-C2 [346] | KIR2DS2 with HLA-C1 [338] KIR2DP1 [339] KIR3DL1 with HLA-Bw4 [344] KIR3DL2 with HLA-A3/11 [344] KIR2DS1 [344] KIR2DS5 [344] |
| Syphilis | KIR1D/KIR1D [347] KIR2DS3 [348] KIR3DS1 [348] | KIR2DS5 [348] KIR2DL3 with HLA-C1C1 [348] |
| Tuberculosis | KIR2DL5 [349] KIR2DL5B [349] KIR2DS2 [349] KIR2DL3 [349] KIR3DL1 [349] KIR2DS1 [349] KIR2DS4 [349] | KIR2DS3 [349] |
| Yersinia pestis | / | / |
| West Nile Virus | / | / |
| Zika Virus | / | / |
6.18. Yersinia Pestis
6.19. West Nile Virus
6.20. Zika Virus
7. KIR Genes and Neurological Diseases
7.1. Autism Spectrum Disorder
7.2. Parkinson’s Disease
7.3. Schizophrenia
8. KIR Genes in Other Diseases
8.1. Acute Ischemic Stroke
8.2. Birdshot Chorioretinopathy
8.3. Endometriosis
8.4. Familial Mediterranean Fever
8.5. Gaucher Disease
8.6. Paroxysmal Nocturnal Hemoglobinuria
8.7. Polycystic Ovary Syndrome
8.8. Pre-Eclampsia
8.9. Takayasu’s Arteritis
| Predisposing KIRs | Protective KIRs | |
|---|---|---|
| Acute ischemic stroke | KIR2DL3 [363] KIR2DL4 [363] KIR2DL5B [363] KIR2DS2 [363] KIR2DS4 [363] KIR3DP1 [363] | / |
| Birdshot chorioretinopathy | KIR2DS2 [365] KIR2DS3 [365] KIR2DS4 [365] | KIR3DL1 with HLA-Bw4I80 [365] KIR2DL1 with HLA-C2 [365] |
| Endometriosis | KIR3DS1 [367] KIR2DS2 [369] | KIR2DS5 [368] |
| Familial Mediterranean Fever | KIR3DP1*003 [371] KIR2DS2 [368] | / |
| Gaucher Disease | / | KIR2DS2 [180,376] KIR2DL2 with HLA-C1 [180,376] |
| Paroxysmal nocturnal hemoglobinuria | KIR3DL1 with HLA-Bw4 [379] | / |
| Polycystic ovary syndrome | KIR3DS1 with HLA-Bw4 [381] | / |
| Preeclampsia | KIR2DL5 [383] KIR2DL1 [390,391,392] KIR3DL2 [387] KIR2DS1 [388] | KIR2DL1 [387,389] KIR2DS5 [389,390] KIR2DL4 [384,385] KIR2DS2 [390] KIR2DS3 [390] |
| Takayasu Arteritis | / | KIR2DS4 [394] |
| Xeroderma pigmentosum | / | / |
8.10. Xeroderma Pigmentosum
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Campbell, K.S.; Purdy, A.K. Structure/function of human killer cell immunoglobulin-like receptors: Lessons from polymorphisms, evolution, crystal structures and mutations. Immunology 2011, 132, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Islam, R.; Pupovac, A.; Evtimov, V.; Boyd, N.; Shu, R.; Boyd, R.; Trounson, A. Enhancing a Natural Killer: Modification of NK Cells for Cancer Immunotherapy. Cells 2021, 10, 1058. [Google Scholar] [CrossRef]
- Vivier, E.; Raulet, D.H.; Moretta, A.; Caligiuri, M.A.; Zitvogel, L.; Lanier, L.L.; Yokoyama, W.M.; Ugolini, S. Innate or adaptive immunity? The example of natural killer cells. Science 2011, 331, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Pazina, T.; Shemesh, A.; Brusilovsky, M.; Porgador, A.; Campbell, K.S. Regulation of natural killer cell receptor functions by interactions with different ligands and alterations in the expression of splice variants. Front. Immunol. 2017, 8, 369. [Google Scholar] [CrossRef]
- Prager, I.; Watzl, C. Mechanisms of natural killer cell-mediated cellular cytotoxicity. J. Leukoc. Biol. 2019, 105, 1319–1329. [Google Scholar] [CrossRef]
- Sivori, S.; Vacca, P.; Del Zotto, G.; Munari, E.; Mingari, M.C.; Moretta, L. Human NK cells: Surface receptors, inhibitory checkpoints, and translational applications. Cell. Mol. Immunol. 2019, 16, 430–441. [Google Scholar] [CrossRef]
- Freud, A.G.; Mundy-Bosse, B.L.; Yu, J.; Caligiuri, M.A. The Broad Spectrum of Human Natural Killer Cell Diversity. Immunity 2017, 47, 820–833. [Google Scholar] [CrossRef]
- Lin, S.J.; Kuo, M.L.; Hsiao, H.S.; Lee, P.T.; Lee, W.I.; Chen, J.Y.; Huang, J.L. Cytotoxic Function and Cytokine Production of Natural Killer Cells and Natural Killer T-Like Cells in Systemic Lupus Erythematosis Regulation with Interleukin-15. Mediat. Inflamm. 2019, 2019, 4236562. [Google Scholar] [CrossRef]
- Gardiner, C.M.; Finlay, D.K. What fuels natural killers? Metabolism and NK cell responses. Front. Immunol. 2017, 8, 367. [Google Scholar] [CrossRef]
- Steiner, N.K.; Dakshanamurthy, S.; VandenBussche, C.J.; Hurley, C.K. Extracellular domain alterations impact surface expression of stimulatory natural killer cell receptor KIR2DS5. Immunogenetics 2008, 60, 655–667. [Google Scholar] [CrossRef]
- Roe, D.; Vierra-Green, C.; Pyo, C.W.; Geraghty, D.E.; Spellman, S.R.; Maiers, M.; Kuang, R. A Detailed View of KIR Haplotype Structures and Gene Families as Provided by a New Motif-Based Multiple Sequence Alignment. Front. Immunol. 2020, 11, 585731. [Google Scholar] [CrossRef] [PubMed]
- Downing, J.; D’Orsogna, L. High-resolution human KIR genotyping. Immunogenetics 2022, 74, 369–379. [Google Scholar] [CrossRef]
- Parham, P.; Norman, P.J.; Abi-Rached, L.; Guethlein, L.A. Human-specific evolution of killer cell immunoglobulin-like receptor recognition of major histocompatibility complex class I molecules. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2012, 367, 800–811. [Google Scholar] [CrossRef] [PubMed]
- Dizaji Asl, K.; Velaei, K.; Rafat, A.; Tayefi Nasrabadi, H.; Movassaghpour, A.A.; Mahdavi, M.; Nozad Charoudeh, H. The role of KIR positive NK cells in diseases and its importance in clinical intervention. Int. Immunopharmacol. 2021, 92, 107361. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.; Halliwell, J.A.; Hayhurst, J.D.; Flicek, P.; Parham, P.; Marsh, S.G. The IPD and IMGT/HLA database: Allele variant databases. Nucleic Acids Res. 2015, 43, D423–D431. [Google Scholar] [CrossRef]
- Ljunggren, H.G.; Karre, K. In search of the “missing self”. MHC molecules and NK cell recognition. Immunol. Today 1990, 11, 237–244. [Google Scholar] [CrossRef]
- Pollock, N.R.; Harrison, G.F.; Norman, P.J. Immunogenomics of Killer Cell Immunoglobulin-Like Receptor (KIR) and HLA Class I: Coevolution and Consequences for Human Health. J. Allergy Clin. Immunol. Pract. 2022, 10, 1763–1775. [Google Scholar] [CrossRef]
- Posch, P.E.; Hurley, C.K. Histocompatibility: HLA and other systems. In Blood and Bone Marrow Pathology, 2nd ed.; Porwit, A., McCullough, J., Erber, W.N., Eds.; Churchill Livingstone: London, UK, 2011. [Google Scholar]
- Pisetsky, D.S. Pathogenesis of autoimmune disease. Nat. Rev. Nephrol. 2023, 19, 509–524. [Google Scholar] [CrossRef]
- Agrawal, S.; Prakash, S. Significance of KIR like natural killer cell receptors in autoimmune disorders. Clin. Immunol. 2020, 216, 108449. [Google Scholar] [CrossRef]
- Chen, B.; Li, J.; He, C.; Li, D.; Tong, W.; Zou, Y.; Xu, W. Role of HLA-B27 in the pathogenesis of ankylosing spondylitis (Review). Mol. Med. Rep. 2017, 15, 1943–1951. [Google Scholar] [CrossRef]
- Parameswaran, P.; Lucke, M. HLA-B27 syndromes. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Lopez-Larrea, C.; Blanco-Gelaz, M.A.; Torre-Alonso, J.C.; Bruges Armas, J.; Suarez-Alvarez, B.; Pruneda, L.; Couto, A.R.; Gonzalez, S.; Lopez-Vázquez, A.; Martinez-Borra, J. Contribution of KIR3DL1/3DS1 to ankylosing spondylitis in human leukocyte antigen-B27 Caucasian populations. Arthritis Res. Ther. 2006, 8, R101. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Li, G.; Ge, R.; Duan, Z.; Zeng, Z.; Zhang, T.; Gao, J.; Yang, T.; Liu, S.; Wu, S.; et al. Association of KIR genotype with susceptibility to HLA-B27-positive ankylosing spondylitis. Mod. Rheumatol. 2013, 23, 538–541. [Google Scholar] [CrossRef] [PubMed]
- Kuijpers, T.W.; Vendelbosch, S.; van den Berg, M.; Baeten, D.L. Killer immunoglobulin receptor genes in spondyloarthritis. Curr. Opin. Rheumatol. 2016, 28, 368–375. [Google Scholar] [CrossRef]
- Fan, D.; Liu, S.; Yang, T.; Wu, S.; Wang, S.; Li, G.; Zeng, Z.; Duan, Z.; Xia, G.; Ye, D.; et al. Association between KIR polymorphisms and ankylosing spondylitis in populations: A meta-analysis. Mod. Rheumatol. 2014, 24, 985–991. [Google Scholar] [CrossRef]
- Rezaei, R.; Mostafaei, S.; Aslani, S.; Jamshidi, A.; Mahmoudi, M. Association study between killer immunoglobulin-like receptor polymorphisms and ankylosing spondylitis disease: An updated meta-analysis. Int. J. Rheum. Dis. 2018, 21, 1746–1755. [Google Scholar] [CrossRef]
- Mack, M.R.; Brestoff, J.R.; Berrien-Elliott, M.M.; Trier, A.M.; Yang, T.B.; McCullen, M.; Collins, P.L.; Niu, H.; Bodet, N.D.; Wagner, J.A.; et al. Blood natural killer cell deficiency reveals an immunotherapy strategy for atopic dermatitis. Sci. Transl. Med. 2020, 12, eaay1005. [Google Scholar] [CrossRef] [PubMed]
- Kabashima, K.; Matsumura, T.; Komazaki, H.; Kawashima, M.; Nemolizumab-JP01 Study Group. Trial of Nemolizumab and Topical Agents for Atopic Dermatitis with Pruritus. N. Engl. J. Med. 2020, 383, 141–150. [Google Scholar] [CrossRef]
- Niepiekło-Miniewska, W.; Majorczyk, E.; Matusiak, L.; Gendzekhadze, K.; Nowak, I.; Narbutt, J.; Lesiak, A.; Kuna, P.; Ponińska, J.; Pietkiewicz-Sworowska, A.; et al. Protective effect of the KIR2DS1 gene in atopic dermatitis. Gene 2013, 527, 594–600. [Google Scholar] [CrossRef]
- Margolis, D.J.; Mitra, N.; Hoffstad, O.J.; Kim, B.S.; Monos, D.S.; Phillips, E.J. Association of KIR Genes and MHC Class I Ligands with Atopic Dermatitis. J. Immunol. 2021, 207, 1522–1529. [Google Scholar] [CrossRef]
- Margolis, D.J.; Mitra, N.; Hoffstad, O.J.; Chopra, A.; Phillips, E.J. KIR Allelic Variation and the Remission of Atopic Dermatitis Over Time. Immunohorizons 2023, 7, 30–40. [Google Scholar] [CrossRef]
- Muratori, L.; Lohse, A.W.; Lenzi, M. Diagnosis and management of autoimmune hepatitis. BMJ 2023, 380, e070201. [Google Scholar] [CrossRef]
- Yoshizawa, K.; Ota, M.; Katsuyama, Y.; Ichijo, T.; Matsumoto, A.; Tanaka, E.; Kiyosawa, K. Genetic analysis of the HLA region of Japanese patients with type 1 autoimmune hepatitis. J. Hepatol. 2005, 42, 578–584. [Google Scholar] [CrossRef] [PubMed]
- van Gerven, N.M.; de Boer, Y.S.; Zwiers, A.; Verwer, B.J.; Drenth, J.P.; van Hoek, B.; van Erpecum, K.J.; Beuers, U.; van Buuren, H.R.; den Ouden, J.W.; et al. HLA-DRB103:01 and HLA-DRB104:01 modify the presentation and outcome in autoimmune hepatitis type-1. Genes Immun. 2015, 16, 247–252. [Google Scholar] [CrossRef]
- Mendoza-Carrera, F.; Gastélum-Meza, M.Á.; Ramírez-García, J.; Dávalos-Cobián, C.; Castro-Martínez, X.H.; Arellano-Olivera, M.I.C.; Hernández-Ramos, L.E.; Leal-Cortés, C. No association of HLA-DRB1 and TNF alleles in Mexican patients with autoimmune hepatitis. Genes Immun. 2019, 20, 678–683. [Google Scholar] [CrossRef]
- Littera, R.; Chessa, L.; Onali, S.; Figorilli, F.; Lai, S.; Secci, L.; La Nasa, G.; Caocci, G.; Arras, M.; Melis, M.; et al. Exploring the role of killer cell immunoglobulin-like receptors and their HLA class I ligands in autoimmune hepatitis. PLoS ONE 2016, 11, e0146086. [Google Scholar] [CrossRef]
- Podhorzer, A.; Paladino, N.; Cuarterolo, M.L.; Fainboim, H.A.; Paz, S.; Theiler, G.; Capucchio, M.; López, S.I.; Machicote, A.; Montal, S.; et al. The early onset of type 1 autoimmune hepatitis has a strong genetic influence: Role of HLA and KIR genes. Genes Immun. 2016, 17, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Umemura, T.; Joshita, S.; Saito, H.; Yoshizawa, K.; Norman, G.L.; Tanaka, E.; Ota, M. KIR/HLA genotypes confer susceptibility and progression in patients with autoimmune hepatitis. JHEP Rep. 2019, 1, 353–360. [Google Scholar] [CrossRef]
- Scherrer, M.A.R.; Rocha, V.B.; Garcia, L.C. Behçet’s disease: Review with emphasis on dermatological aspects. An. Bras. Dermatol. 2017, 92, 452–464. [Google Scholar] [CrossRef]
- van der Houwen, T.B.; van Hagen, P.M.; van Laar, J.A.M. Immunopathogenesis of Behçet’s disease and treatment modalities. Semin. Arthritis Rheum. 2022, 52, 151956. [Google Scholar] [CrossRef]
- Middleton, D.; Meenagh, A.; Sleator, C.; Gourraud, P.A.; Ayna, T.; Tozkir, H.; Köse, A.A.; Azizleri, G.; Diler, A.S. No association of KIR genes with Behçet’s disease. Tissue Antigens 2007, 70, 435–438. [Google Scholar] [CrossRef]
- Arayssi, T.K.; El Hajj, N.; Shamseddine, W.; Ibrahim, G.; Nasr, J.; Sabbagh, A.S.; Greige, L.; Zaatari, G.S.; Mahfouz, R.A. Killer cell immunoglobulin-like receptor genotypes in Behçet’s disease patients: Any role for the 3DP1*001/002 pseudogene? Genet. Test. Mol. Biomark. 2009, 13, 319–324. [Google Scholar] [CrossRef]
- Mohammad-Ebrahim, H.; Kamali-Sarvestani, E.; Mahmoudi, M.; Beigy, M.; Karami, J.; Ahmadzadeh, N.; Shahram, F. Association of killer cell immunoglobulin-like receptor (KIR) genes and their HLA ligands with susceptibility to Behçet’s disease. Scand. J. Rheumatol. 2018, 47, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Erer, B.; Takeuchi, M.; Ustek, D.; Tugal-Tutkun, I.; Seyahi, E.; Özyazgan, Y.; Duymaz-Tozkir, J.; Gül, A.; Kastner, D.L.; Remmers, E.F.; et al. Evaluation of KIR3DL1/KIR3DS1 polymorphism in Behçet’s disease. Genes Immun. 2016, 17, 396–399. [Google Scholar] [CrossRef] [PubMed]
- Castaño-Núñez, Á.; Montes-Cano, M.A.; García-Lozano, J.R.; Ortego-Centeno, N.; García-Hernández, F.J.; Espinosa, G.; Graña-Gil, G.; Sánchez-Bursón, J.; Juliá, M.R.; Solans, R.; et al. Association of Functional Polymorphisms of KIR3DL1/DS1 with Behçet’s Disease. Front. Immunol. 2019, 10, 2755. [Google Scholar] [CrossRef]
- Caio, G.; Volta, U.; Sapone, A.; Leffler, D.A.; De Giorgio, R.; Catassi, C.; Fasano, A. Celiac disease: A comprehensive current review. BMC Med. 2019, 17, 142. [Google Scholar] [CrossRef]
- Iversen, R.; Sollid, L.M. The immunobiology and pathogenesis of celiac disease. Annu. Rev. Pathol. 2023, 18, 47–70. [Google Scholar] [CrossRef]
- Moodie, S.J.; Norman, P.J.; King, A.L.; Fraser, J.S.; Curtis, D.; Ellis, H.J.; Vaughan, R.W.; Ciclitira, P.J. Analysis of candidate genes on chromosome 19 in coeliac disease: An association study of the KIR and LILR gene clusters. Eur. J. Immunogenet. 2002, 29, 287–291. [Google Scholar] [CrossRef]
- Santin, I.; Castellanos-Rubio, A.; Perez de Nanclares, G.; Vitoria, J.C.; Castaño, L.; Bilbao, J.R. Association of KIR2DL5B gene with celiac disease supports the susceptibility locus on 19q13.4. Genes Immun. 2007, 8, 171–176. [Google Scholar] [CrossRef]
- Fernandez-Jimenez, N.; Santín, I.; Irastorza, I.; Plaza-Izurieta, L.; Castellanos-Rubio, A.; Vitoria, J.C.; Bilbao, J.R. Upregulation of KIR3DL1 gene expression in intestinal mucosa in active celiac disease. Hum. Immunol. 2011, 72, 617–620. [Google Scholar] [CrossRef]
- Caggiari, L.; Toffoli, G.; De Re, V.; Orzes, N.; Spina, M.; De Zorzi, M.; Maiero, S.; Cannizzaro, R.; Canzonieri, V. KIR/HLA combination associated with the risk of complications in celiac disease. Int. J. Biol. Markers 2011, 26, 221–228. [Google Scholar] [CrossRef]
- Smigoc Schweiger, D.; Mendez, A.; Kunilo Jamnik, S.; Bratanic, N.; Bratina, N.; Battelino, T.; Brecelj, J.; Vidan-Jeras, B. Genetic risk for co-occurrence of type 1 diabetes and celiac disease is modified by HLA-C and killer immunoglobulin-like receptors. Tissue Antigens 2014, 84, 471–478. [Google Scholar] [CrossRef]
- Akar, H.H.; Patiroglu, T.; Sevinc, E.; Aslan, D.; Okdemir, D.; Kurtoglu, S. Contribution of KIR genes, HLA class I ligands, and KIR/HLA class I ligand combinations on the genetic predisposition to celiac disease and coexisting celiac disease and type 1 diabetes mellitus. Rev. Esp. Enferm. Dig. 2015, 107, 547–553. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Uricoechea, H.; Nogueira, J.P.; Pinzón-Fernández, M.V.; Schwarzstein, D. The usefulness of thyroid antibodies in the diagnostic approach to autoimmune thyroid disease. Antibodies 2023, 12, 48. [Google Scholar] [CrossRef] [PubMed]
- Solerte, S.B.; Precerutti, S.; Gazzaruso, C.; Locatelli, E.; Zamboni, M.; Schifino, N.; Bonacasa, R.; Rondanelli, M.; Taccani, D.; Ferrari, E.; et al. Defect of a subpopulation of natural killer immune cells in Graves’ disease and Hashimoto’s thyroiditis: Normalizing effect of dehydroepiandrosterone sulfate. Eur. J. Endocrinol. 2005, 152, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.Q.; Zhao, J.J.; Zhao, Y.R.; Guan, Q.B.; Gao, L.; Song, H.D. Genotype analysis of killer cell immunoglobulin-like receptors in Graves’ disease patients. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi 2009, 25, 699–701. (In Chinese) [Google Scholar]
- Ashouri, E.; Dabbaghmanesh, M.H.; Hadaegh, A.; Rowhanirad, S.; Bakhshayashkaram, M.; Omrani, G.R. KIR gene content does not contribute to susceptibility to Graves’ disease. Iran. J. Immunol. 2013, 10, 150–157. [Google Scholar]
- Dastmalchi, R.; Farazmand, A.; Noshad, S.; Mozafari, M.; Mahmoudi, M.; Esteghamati, A.; Amirzargar, A. Polymorphism of killer cell immunoglobulin-like receptors (KIR) and their HLA ligands in Graves’ disease. Mol. Biol. Rep. 2014, 41, 5367–5374. [Google Scholar] [CrossRef]
- Li, S.Q.; Guo, C.; Wang, X.S.; Hou, Y.F.; Li, J.T.; Zhang, H.Q. Correlation between gene polymorphisms of killer cell immunoglobulin-like receptors and their ligands and Graves’ disease. Zhonghua Yi Xue Za Zhi 2023, 103, 344–349. (In Chinese) [Google Scholar] [CrossRef]
- Ralli, M.; Angeletti, D.; Fiore, M.; D’Aguanno, V.; Lambiase, A.; Artico, M.; de Vincentiis, M.; Greco, A. Hashimoto’s thyroiditis: An update on pathogenic mechanisms, diagnostic protocols, therapeutic strategies, and potential malignant transformation. Autoimmun. Rev. 2020, 19, 102649. [Google Scholar] [CrossRef]
- Ashouri, E.; Dabbaghmanesh, M.H.; Ranjbar Omrani, G. Presence of more activating KIR genes is associated with Hashimoto’s thyroiditis. Endocrine 2014, 46, 519–525. [Google Scholar] [CrossRef]
- Li, J.T.; Guo, C.; Li, M.L.; Wei, Y.Q.; Hou, Y.F.; Jiao, Y.L.; Zhao, Y.R.; Sun, H.; Xu, J.; Cao, M.F.; et al. Killer cell immunoglobulin-like receptor genes and their HLA-C ligands in Hashimoto thyroiditis in a Chinese population. Endocr. Pract. 2016, 22, 935–940. [Google Scholar] [CrossRef]
- Olsson, B.; Andersson, P.O.; Jernås, M.; Jacobsson, S.; Carlsson, B.; Carlsson, L.M.; Wadenvik, H. T-cell-mediated cytotoxicity toward platelets in chronic idiopathic thrombocytopenic purpura. Nat. Med. 2003, 9, 1123–1124. [Google Scholar] [CrossRef]
- Nourse, J.P.; Lea, R.; Crooks, P.; Wright, G.; Tran, H.; Catalano, J.; Brighton, T.; Grigg, A.; Marlton, P.; Gandhi, M.K. The KIR2DS2/DL2 genotype is associated with adult persistent/chronic and relapsed immune thrombocytopenia independently of FCGR3a-158 polymorphisms. Blood Coagul. Fibrinolysis 2012, 23, 45–50. [Google Scholar] [CrossRef] [PubMed]
- El-Beblawy, N.M.; Elbarbary, N.S.; Kamal, T.M.; Mahmoud, P.M. A study of human killer cell immunoglobulin-like receptor and multidrug resistance gene polymorphisms in children with immune thrombocytopenia. Clin. Appl. Thromb. Hemost. 2016, 22, 429–440. [Google Scholar] [CrossRef] [PubMed]
- Seymour, L.A.; Nourse, J.P.; Crooks, P.; Wockner, L.; Bird, R.; Tran, H.; Gandhi, M.K. The presence of KIR2DS5 confers protection against adult immune thrombocytopenia. Tissue Antigens 2014, 83, 154–160. [Google Scholar] [CrossRef]
- Jones, D.C.; Edgar, R.S.; Ahmad, T.; Cummings, J.R.; Jewell, D.P.; Trowsdale, J.; Young, N.T. Killer Ig-like receptor (KIR) genotype and HLA ligand combinations in ulcerative colitis susceptibility. Genes Immun. 2006, 7, 576–582. [Google Scholar] [CrossRef] [PubMed]
- Hollenbach, J.A.; Ladner, M.B.; Saeteurn, K.; Taylor, K.D.; Mei, L.; Haritunians, T.; McGovern, D.P.; Erlich, H.A.; Rotter, J.I.; Trachtenberg, E.A. Susceptibility to Crohn’s disease is mediated by KIR2DL2/KIR2DL3 heterozygosity and the HLA-C ligand. Immunogenetics 2009, 61, 663–671. [Google Scholar] [CrossRef]
- Díaz-Peña, R.; Vidal-Castiñeira, J.R.; Moro-García, M.A.; Alonso-Arias, R.; Castro-Santos, P. Significant association of the KIR2DL3/HLA-C1 genotype with susceptibility to Crohn’s disease. Hum. Immunol. 2016, 77, 104–109. [Google Scholar] [CrossRef]
- Fathollahi, A.; Aslani, S.; Mostafaei, S.; Rezaei, N.; Mahmoudi, M. The role of killer-cell immunoglobulin-like receptor (KIR) genes in susceptibility to inflammatory bowel disease: Systematic review and meta-analysis. Inflamm. Res. 2018, 67, 727–736. [Google Scholar] [CrossRef]
- Samarani, S.; Mack, D.R.; Bernstein, C.N.; Iannello, A.; Debbeche, O.; Jantchou, P.; Faure, C.; Deslandres, C.; Amre, D.K.; Ahmad, A. Activating Killer-cell Immunoglobulin-like Receptor genes confer risk for Crohn’s disease in children and adults of the Western European descent: Findings based on case-control studies. PLoS ONE 2019, 14, e0217767. [Google Scholar] [CrossRef]
- Beigmohammadi, F.; Mahmoudi, M.; Karami, J.; Ahmadzadeh, N.; Ebrahimi-Daryani, N.; Rezaei, N. Analysis of Killer Cell Immunoglobulin-Like Receptor Genes and Their HLA Ligands in Inflammatory Bowel Diseases. J. Immunol. Res. 2020, 2020, 4873648. [Google Scholar] [CrossRef]
- Zhou, J.; Tang, X.; Ding, Y.; An, Y.; Zhao, X. Natural killer cell activity and frequency of killer cell immunoglobulin-like receptors in children with different forms of juvenile idiopathic arthritis. Pediatr. Allergy Immunol. 2013, 24, 691–696. [Google Scholar] [CrossRef] [PubMed]
- Gaur, P.; Misra, R.; Aggarwal, A. Natural killer cell and gamma delta T cell alterations in enthesitis related arthritis category of juvenile idiopathic arthritis. Clin. Immunol. 2015, 161, 163–169. [Google Scholar] [CrossRef]
- Jalalvand, M.; Beigmohammadi, F.; Soltani, S.; Ehsan, S.; Rajabkhah, S.; Madreseh, E.; Akhtari, M.; Jamshidi, A.; Farhadi, E.; Mahmoudi, M.; et al. The investigation of killer-cell immunoglobulin-like receptors (KIRs) and their HLA ligands in Iranian patients with myasthenia gravis. Clin. Neurol. Neurosurg. 2024, 238, 108171. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Kiapour, N.; Kapoor, S.; Khan, T.; Thamilarasan, M.; Tao, Y.; Cohen, S.; Miller, R.; Sobel, R.A.; Markovic-Plese, S. IL-11 Induces Encephalitogenic Th17 Cells in Multiple Sclerosis and Experimental Autoimmune Encephalomyelitis. J. Immunol. 2019, 203, 1142–1150. [Google Scholar] [CrossRef]
- Seyedsadr, M.; Wang, Y.; Elzoheiry, M.; Shree Gopal, S.; Jang, S.; Duran, G.; Chervoneva, I.; Kasimoglou, E.; Wrobel, J.A.; Hwang, D.; et al. IL-11 induces NLRP3 inflammasome activation in monocytes and inflammatory cell migration to the central nervous system. Proc. Natl. Acad. Sci. USA 2023, 120, e2221007120. [Google Scholar] [CrossRef] [PubMed]
- Baecher-Allan, C.; Kaskow, B.J.; Weiner, H.L. Multiple Sclerosis: Mechanisms and Immunotherapy. Neuron 2018, 97, 742–768. [Google Scholar] [CrossRef]
- García-León, J.A.; Pinto-Medel, M.J.; García-Trujillo, L.; López-Gómez, C.; Oliver-Martos, B.; Prat-Arrojo, I.; Marín-Bañasco, C.; Suardíaz-García, M.; Maldonado-Sanchez, R.; Fernández-Fernández, O.; et al. Killer cell immunoglobulin-like receptor genes in Spanish multiple sclerosis patients. Mol. Immunol. 2011, 48, 1896–1902. [Google Scholar] [CrossRef]
- Bettencourt, A.; Silva, A.M.; Carvalho, C.; Leal, B.; Santos, E.; Costa, P.P.; Silva, B.M. The role of KIR2DS1 in multiple sclerosis—KIR in Portuguese MS patients. J. Neuroimmunol. 2014, 269, 52–55. [Google Scholar] [CrossRef]
- Lorentzen, A.R.; Karlsen, T.H.; Olsson, M.; Smestad, C.; Mero, I.L.; Woldseth, B.; Sun, J.Y.; Senitzer, D.; Celius, E.G.; Thorsby, E.; et al. Killer immunoglobulin-like receptor ligand HLA-Bw4 protects against multiple sclerosis. Ann. Neurol. 2009, 65, 658–666. [Google Scholar] [CrossRef]
- Fusco, C.; Guerini, F.R.; Nocera, G.; Ventrella, G.; Caputo, D.; Valentino, M.A.; Agliardi, C.; Gallotti, J.; Morra, V.B.; Florio, C.; et al. KIRs and their HLA ligands in remitting-relapsing multiple sclerosis. J. Neuroimmunol. 2010, 229, 232–237. [Google Scholar] [CrossRef]
- Jelcić, I.; Hsu, K.C.; Kakalacheva, K.; Breiden, P.; Dupont, B.; Uhrberg, M.; Martin, R.; Münz, C.; Lünemann, J.D. Killer immunoglobulin-like receptor locus polymorphisms in multiple sclerosis. Mult. Scler. 2012, 18, 951–958. [Google Scholar] [CrossRef] [PubMed]
- Hollenbach, J.A.; Pando, M.J.; Caillier, S.J.; Gourraud, P.A.; Oksenberg, J.R. The killer immunoglobulin-like receptor KIR3DL1 in combination with HLA-Bw4 is protective against multiple sclerosis in African Americans. Genes Immun. 2016, 17, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Shahsavar, F.; Mapar, S.; Ahmadi, S.A. Multiple sclerosis is accompanied by lack of KIR2DS1 gene: A meta-analysis. Genom. Data 2016, 10, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Mack, S.J.; Udell, J.; Cohen, F.; Osoegawa, K.; Hawbecker, S.K.; Noonan, D.A.; Ladner, M.B.; Goodridge, D.; Trachtenberg, E.A.; Oksenberg, J.R.; et al. High resolution HLA analysis reveals independent class I haplotypes and amino-acid motifs protective for multiple sclerosis. Genes Immun. 2019, 20, 308–326. [Google Scholar] [CrossRef]
- Roszkowska, A.; Pawlicka, M.; Mroczek, A.; Bałabuszek, K.; Nieradko-Iwanicka, B. Non-Celiac Gluten Sensitivity: A Review. Medicina 2019, 55, 222. [Google Scholar] [CrossRef]
- Gambino, C.M.; Agnello, L.; Vidali, M.; Lo Sasso, B.; Mansueto, P.; Seidita, A.; Giuliano, A.; Scazzone, C.; Massa, D.; Masucci, A.; et al. The role of killer immunoglobulin-like receptors (KIRs) in the genetic susceptibility to non-celiac wheat sensitivity (NCWS). Clin. Chem. Lab. Med. 2024, 62, 1814–1823. [Google Scholar] [CrossRef]
- Martin, M.P.; Nelson, G.; Lee, J.H.; Pellett, F.; Gao, X.; Wade, J.; Wilson, M.J.; Trowsdale, J.; Gladman, D.; Carrington, M. Cutting edge: Susceptibility to psoriatic arthritis: Influence of activating killer Ig-like receptor genes in the absence of specific HLA-C alleles. J. Immunol. 2002, 169, 2818–2822. [Google Scholar] [CrossRef]
- Suzuki, Y.; Hamamoto, Y.; Ogasawara, Y.; Ishikawa, K.; Yoshikawa, Y.; Sasazuki, T.; Muto, M. Genetic polymorphisms of killer cell immunoglobulin-like receptors are associated with susceptibility to psoriasis vulgaris. J. Investig. Dermatol. 2004, 122, 1133–1136. [Google Scholar] [CrossRef]
- Holm, S.J.; Sakuraba, K.; Mallbris, L.; Wolk, K.; Ståhle, M.; Sánchez, F.O. Distinct HLA-C/KIR genotype profile associates with guttate psoriasis. J. Investig. Dermatol. 2005, 125, 721–730. [Google Scholar] [CrossRef]
- Williams, F.; Meenagh, A.; Sleator, C.; Cook, D.; Fernandez-Vina, M.; Bowcock, A.M.; Middleton, D. Activating killer cell immunoglobulin-like receptor gene KIR2DS1 is associated with psoriatic arthritis. Hum. Immunol. 2005, 66, 836–841. [Google Scholar] [CrossRef] [PubMed]
- Chandran, V.; Bull, S.B.; Pellett, F.J.; Ayearst, R.; Pollock, R.A.; Gladman, D.D. Killer-cell immunoglobulin-like receptor gene polymorphisms and susceptibility to psoriatic arthritis. Rheumatology 2014, 53, 233–239. [Google Scholar] [CrossRef]
- Enciso-Vargas, M.; Alvarado-Ruíz, L.; Suárez-Villanueva, A.S.; Macías-Barragán, J.; Montoya-Buelna, M.; Oceguera-Contreras, E.; Alvarado-Navarro, A.; Graciano-Machuca, O. Association Study between Psoriatic Arthritis and Killer Immunoglobulin-Like Receptor (KIR) Genes: A Meta-Analysis. Immunol. Investig. 2021, 50, 152–163. [Google Scholar] [CrossRef]
- Lowes, M.A.; Suárez-Fariñas, M.; Krueger, J.G. Immunology of psoriasis. Annu. Rev. Immunol. 2014, 32, 227–255. [Google Scholar] [CrossRef] [PubMed]
- Łuszczek, W.; Mańczak, M.; Cisło, M.; Nockowski, P.; Wiśniewski, A.; Jasek, M.; Kuśnierczyk, P. Gene for the activating natural killer cell receptor, KIR2DS1, is associated with susceptibility to psoriasis vulgaris. Hum. Immunol. 2004, 65, 758–766. [Google Scholar] [CrossRef]
- Chang, Y.T.; Chou, C.T.; Shiao, Y.M.; Lin, M.W.; Yu, C.W.; Chen, C.C.; Huang, C.H.; Lee, D.D.; Liu, H.N.; Wang, W.J.; et al. The killer cell immunoglobulin-like receptor genes do not confer susceptibility to psoriasis vulgaris independently in Chinese. J. Investig. Dermatol. 2006, 126, 2335–2338. [Google Scholar] [CrossRef]
- Jobim, M.; Jobim, L.F.; Salim, P.H.; Cestari, T.F.; Toresan, R.; Gil, B.C.; Jobim, M.R.; Wilson, T.J.; Kruger, M.; Schlottfeldt, J.; et al. A study of the killer cell immunoglobulin-like receptor gene KIR2DS1 in a Caucasoid Brazilian population with psoriasis vulgaris. Tissue Antigens 2008, 72, 392–396. [Google Scholar] [CrossRef]
- Solgi, G.; Ghafari, H.; Ashouri, E.; Alimoghdam, K.; Rajalingam, R.; Amirzargar, A. Comparison of KIR gene content profiles revealed a difference between northern and southern Persians in the distribution of KIR2DS5 and its linked loci. Hum. Immunol. 2011, 72, 1079–1083. [Google Scholar] [CrossRef] [PubMed]
- Graciano-Machuca, O.; Alvarado-Navarro, A.; Ramírez-Dueñas, M.G.; Villanueva-Quintero, D.G.; Velarde-de la Cruz, E.E.; Machado-Sulbarán, A.C.; Montoya-Buelna, M.; Sánchez-Hernández, P.E. Diversity of KIR/HLA Genotypes and Their Association with Psoriasis Vulgaris in the Western Mexican Population. Genes 2020, 11, 338. [Google Scholar] [CrossRef]
- Macías-Barragán, J.; Montoya-Buelna, M.; Enciso-Vargas, M.; Alvarado-Ruíz, L.; Oceguera-Contreras, E.; Guerra-Renteria, A.S.; Graciano-Machuca, O. Assessment of the Relationship between Clinical Variants of Psoriasis and Killer Immunoglobulin-like Receptor (KIR) Genes: A Systematic Review with Meta-analysis. Immunol. Investig. 2022, 51, 480–495. [Google Scholar] [CrossRef]
- Yen, J.H.; Moore, B.E.; Nakajima, T.; Scholl, D.; Schaid, D.J.; Weyand, C.M.; Goronzy, J.J. Major histocompatibility complex class I-recognizing receptors are disease risk genes in rheumatoid arthritis. J. Exp. Med. 2001, 193, 1159–1167. [Google Scholar] [CrossRef]
- Ramírez-De los Santos, S.; Sánchez-Hernández, P.E.; Muñoz-Valle, J.F.; Palafox-Sánchez, C.A.; Rosales-Rivera, L.Y.; García-Iglesias, T.; Daneri-Navarro, A.; Ramírez-Dueñas, M.G. Associations of killer cell immunoglobulin-like receptor genes with rheumatoid arthritis. Dis. Markers 2012, 33, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xia, Q.; Fan, D.; Cai, G.; Yang, X.; Wang, L.; Xin, L.; Ding, N.; Hu, Y.; Liu, L.; et al. Association between KIR gene polymorphisms and rheumatoid arthritis susceptibility: A meta-analysis. Hum. Immunol. 2015, 76, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Aghaei, H.; Mostafaei, S.; Aslani, S.; Jamshidi, A.; Mahmoudi, M. Association study between KIR polymorphisms and rheumatoid arthritis disease: An updated meta-analysis. BMC Med. Genet. 2019, 20, 24. [Google Scholar] [CrossRef]
- Velarde-de la Cruz, E.E.; Sánchez-Hernández, P.E.; Muñoz-Valle, J.F.; Palafox-Sánchez, C.A.; Ramírez-de Los Santos, S.; Graciano-Machuca, O.; García-Iglesias, T.; Montoya-Buelna, M.; Ramírez-Dueñas, M.G. KIR2DL2 and KIR2DS2 as genetic markers to the methotrexate response in rheumatoid arthritis patients. Immunopharmacol. Immunotoxicol. 2016, 38, 303–309. [Google Scholar] [CrossRef]
- Ramírez, S.; Ramírez, M.G.; Muñoz, J.F.; Martínez, G.E.; Velarde, E.E.; Sánchez, P.E. Association of KIR2DL2 gene with anti-cyclic citrullinated protein antibodies for serodiagnosis in rheumatoid arthritis. Medicina (B Aires) 2019, 79, 161–166. [Google Scholar]
- Ansari-Moghaddam, B.; Asghar Kiani, A.; Sheikhian, A.; Birjandi, M.; Ahmadi, S.A.Y.; Mousavi, N.; Torang, H.A.; Shahsavar, F. Rheumatoid arthritis susceptibility is associated with the KIR2DS4-full of killer-cell immunoglobulin-like receptor genes in the Lur population of Iran. Rep. Biochem. Mol. Biol. 2021, 10, 84–94. [Google Scholar] [CrossRef]
- Reksten, T.R.; Lessard, C.J.; Sivils, K.L. Genetics in Sjögren syndrome. Rheum. Dis. Clin. N. Am. 2016, 42, 435–447. [Google Scholar] [CrossRef]
- Lowe, D.P.; Cook, M.A.; Bowman, S.J.; Briggs, D.C.; UK Sjögren’s Interest Group. Association of killer cell immunoglobulin-like receptors with primary Sjögren’s syndrome. Rheumatology 2009, 48, 359–362. [Google Scholar] [CrossRef]
- Pellett, F.; Siannis, F.; Vukin, I.; Lee, P.; Urowitz, M.B.; Gladman, D.D. KIRs and autoimmune disease: Studies in systemic lupus erythematosus and scleroderma. Tissue Antigens 2007, 69 (Suppl. S1), 106–108. [Google Scholar] [CrossRef]
- Kimoto, Y.; Horiuchi, T.; Tsukamoto, H.; Kiyohara, C.; Mitoma, H.; Uchino, A.; Furugo, I.; Yoshizawa, S.; Ueda, A.; Harashima, S.; et al. Association of killer cell immunoglobulin-like receptor 2DL5 with systemic lupus erythematosus and accompanying infections. Rheumatology 2010, 49, 1346–1353. [Google Scholar] [CrossRef]
- Chen, A.M.; Liu, Q.P.; Cui, X.Y.; Cui, M.Y.; Yan, W.Y.; Xie, S.M.; Zhu, N.; Wang, X.D. Study on the polymorphism of killer cell immunoglobulin-like receptor (KIR) gene with systemic lupus erythematosus of North population in China. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi 2008, 24, 811–813. (In Chinese) [Google Scholar] [PubMed]
- Hou, Y.F.; Zhang, Y.C.; Jiao, Y.L.; Wang, L.C.; Li, J.F.; Pan, Z.L.; Yang, Q.R.; Sun, H.S.; Zhao, Y.R. Disparate distribution of activating and inhibitory killer cell immunoglobulin-like receptor genes in patients with systemic lupus erythematosus. Lupus 2010, 19, 20–26. [Google Scholar] [CrossRef]
- Pedroza, L.S.; Sauma, M.F.; Vasconcelos, J.M.; Takeshita, L.Y.; Ribeiro-Rodrigues, E.M.; Sastre, D.; Barbosa, C.M.; Chies, J.A.; Veit, T.D.; Lima, C.P.; et al. Systemic lupus erythematosus: Association with KIR and SLC11A1 polymorphisms, ethnic predisposition and influence in clinical manifestations at onset revealed by ancestry genetic markers in an urban Brazilian population. Lupus 2011, 20, 265–273. [Google Scholar] [CrossRef]
- Akhtari, M.; Farazmand, A.; Mahmoudi, M.; Akbarian, M.; Ahmadzadeh, N.; Mirkazemi, Z.; Mostafaei, S.; Jamshidi, A.R. Analysis of killer cell immunoglobulin-like receptors and their human leukocyte antigen-ligands gene polymorphisms in Iranian patients with systemic lupus erythematosus. Lupus 2016, 25, 1244–1253. [Google Scholar] [CrossRef] [PubMed]
- Tozkır, J.D.; Tozkır, H.; Gürkan, H.; Dönmez, S.; Eker, D.; Pamuk, G.E.; Pamuk, Ö.N. The investigation of killer cell immunoglobulin-like receptor genotyping in patients with systemic lupus erythematosus and systemic sclerosis. Clin. Rheumatol. 2016, 35, 919–925. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.L.; Ma, S.J.; Tan, H.Z. Association between killer cell immunoglobulin-like receptor (KIR) polymorphisms and systemic lupus erythematosus (SLE) in populations: A PRISMA-compliant meta-analysis. Medicine 2017, 96, e6166. [Google Scholar] [CrossRef]
- Gambino, C.M.; Di Bona, D.; Aiello, A.; Carru, C.; Duro, G.; Guggino, G.; Ferrante, A.; Zinellu, A.; Caruso, C.; Candore, G.; et al. HLA-C1 ligands are associated with increased susceptibility to systemic lupus erythematosus. Hum. Immunol. 2018, 79, 172–177. [Google Scholar] [CrossRef]
- Segerberg, F.; Lundtoft, C.; Reid, S.; Hjorton, K.; Leonard, D.; Nordmark, G.; Carlsten, M.; Hagberg, N. Autoantibodies to killer cell immunoglobulin-like receptors in patients with systemic lupus erythematosus induce natural killer cell hyporesponsiveness. Front. Immunol. 2019, 10, 2164. [Google Scholar] [CrossRef]
- Momot, T.; Koch, S.; Hunzelmann, N.; Krieg, T.; Ulbricht, K.; Schmidt, R.E.; Witte, T. Association of killer cell immunoglobulin-like receptors with scleroderma. Arthritis Rheum. 2004, 50, 1561–1565. [Google Scholar] [CrossRef]
- Salim, P.H.; Jobim, M.; Bredemeier, M.; Chies, J.A.; Schlottfeldt, J.; Brenol, J.C.; Jobim, L.F.; Xavier, R.M. Killer cell immunoglobulin-like receptor (KIR) genes in systemic sclerosis. Clin. Exp. Immunol. 2010, 160, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudi, M.; Fallahian, F.; Sobhani, S.; Ghoroghi, S.; Jamshidi, A.; Poursani, S.; Dolati, M.; Hosseinpour, Z.; Gharibdoost, F. Analysis of killer cell immunoglobulin-like receptors (KIRs) and their HLA ligand genes polymorphisms in Iranian patients with systemic sclerosis. Clin. Rheumatol. 2017, 36, 853–862. [Google Scholar] [CrossRef]
- Machado-Sulbaran, A.C.; Ramírez-Dueñas, M.G.; Navarro-Zarza, J.E.; Muñoz-Valle, J.F.; Mendoza-Carrera, F.; Baños-Hernández, C.J.; Parra-Rojas, I.; Montoya-Buelna, M.; Sánchez-Hernández, P.E. KIR/HLA gene profile implication in systemic sclerosis patients from Mexico. J. Immunol. Res. 2019, 2019, 6808061. [Google Scholar] [CrossRef] [PubMed]
- Karimizadeh, E.; Mostafaei, S.; Aslani, S.; Gharibdoost, F.; Xavier, R.M.; Salim, P.H.; Kavosi, H.; Farhadi, E.; Mahmoudi, M. Evaluation of the association between KIR polymorphisms and systemic sclerosis: A meta-analysis. Adv. Rheumatol. 2020, 60, 8. [Google Scholar] [CrossRef] [PubMed]
- Kieleväinen, V.; Turtinen, M.; Luopajärvi, K.; Härkönen, T.; Ilonen, J.; Knip, M.; Finnish Pediatric Diabetes Register. Increased HLA class II risk is associated with a more aggressive presentation of clinical type 1 diabetes. Acta Paediatr. 2023, 112, 522–528. [Google Scholar] [CrossRef]
- Luckett, A.M.; Weedon, M.N.; Hawkes, G.; Leslie, R.D.; Oram, R.A.; Grant, S.F.A. Utility of genetic risk scores in type 1 diabetes. Diabetologia 2023, 66, 1589–1600. [Google Scholar] [CrossRef]
- Van der Slik, A.R.; Koeleman, B.P.; Verduijn, W.; Bruining, G.J.; Roep, B.O.; Giphart, M.J. KIR in type 1 diabetes: Disparate distribution of activating and inhibitory natural killer cell receptors in patients versus HLA-matched control subjects. Diabetes 2003, 52, 2639–2642. [Google Scholar] [CrossRef]
- Santin, I.; de Nanclares, G.P.; Calvo, B.; Gaafar, A.; Castaño, L.; GEPV-N Group; Bilbao, J.R. Killer cell immunoglobulin-like receptor (KIR) genes in the Basque population: Association study of KIR gene contents with type 1 diabetes mellitus. Hum. Immunol. 2006, 67, 118–124. [Google Scholar] [CrossRef]
- Sanjeevi, S.; Sun, C.; Kanungo, A.; Sanjeevi, C.B. Killer immunoglobulin receptor genes and their HLA-C ligand are associated with type 1 diabetes in an Eastern Indian population. Diabet. Med. 2016, 33, 91–96. [Google Scholar] [CrossRef]
- Sun, C.; Sanjeevi, S.; Luo, F.; Zhi, D.; Sanjeevi, C.B. Interactions between maternal killer cell immunoglobulin receptor genes and fetal HLA ligand genes contribute to type 1 diabetes susceptibility in Han Chinese. Int. J. Immunogenet. 2016, 43, 125–130. [Google Scholar] [CrossRef]
- Liu, S.L.; Zheng, A.J.; Ding, L. Association between KIR gene polymorphisms and type 1 diabetes mellitus (T1DM) susceptibility: A PRISMA-compliant meta-analysis. Medicine 2017, 96, e9439. [Google Scholar] [CrossRef] [PubMed]
- Soltani, S.; Mostafaei, S.; Aslani, S.; Farhadi, E.; Mahmoudi, M. Association of KIR gene polymorphisms with type 1 diabetes: A meta-analysis. J. Diabetes Metab. Disord. 2020, 19, 1777–1786. [Google Scholar] [CrossRef]
- Gunavathy, N.; Asirvatham, A.; Chitra, A.; Jayalakshmi, M. Association of killer cell immunoglobulin-like receptors and their HLA-ligands with type 1 diabetes among South Indian population. Immunol. Investig. 2023, 52, 270–285. [Google Scholar] [CrossRef] [PubMed]
- Levinson, R.D.; Du, Z.; Luo, L.; Holland, G.N.; Rao, N.A.; Reed, E.F.; Rajalingam, R. KIR and HLA gene combinations in Vogt-Koyanagi-Harada disease. Hum. Immunol. 2008, 69, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Levinson, R.D.; Okada, A.A.; Ashouri, E.; Keino, H.; Rajalingam, R. Killer cell immunoglobulin-like receptor gene-cluster 3DS1-2DL5-2DS1-2DS5 predisposes susceptibility to Vogt-Koyanagi-Harada syndrome in Japanese individuals. Hum. Immunol. 2010, 71, 192–194. [Google Scholar] [CrossRef]
- Sheereen, A.; Gaafar, A.; Iqneibi, A.; Eldali, A.; Tabbara, K.F.; Adra, C.; Al-Hussein, K. A study of KIR genes and HLA-C in Vogt-Koyanagi-Harada disease in Saudi Arabia. Mol. Vis. 2011, 17, 3523–3528. [Google Scholar]
- Levinson, R.D.; Yung, M.; Meguro, A.; Ashouri, E.; Yu, F.; Mizuki, N.; Ohno, S.; Rajalingam, R. KIR and HLA genotypes implicated in reduced killer lymphocytes immunity are associated with Vogt-Koyanagi-Harada disease. PLoS ONE 2016, 11, e0160392. [Google Scholar] [CrossRef]
- Boudreau, J.E.; Hsu, K.C. Natural killer cell education and the response to infection and cancer therapy: Stay tuned. Trends Immunol. 2018, 39, 222–239. [Google Scholar] [CrossRef]
- Spear, P.; Wu, M.R.; Sentman, M.L. NKG2D ligands as therapeutic targets. Cancer Immunol. Immunother. 2019, 68, 1353–1365. [Google Scholar] [CrossRef]
- Saunders, P.M.; MacLachlan, B.J.; Pymm, P.; Illing, P.T.; Deng, Y.; Wong, S.C.; Oates, C.V.L.; Purcell, A.W.; Rossjohn, J.; Vivian, J.P.; et al. The molecular basis of how buried human leukocyte antigen polymorphism modulates natural killer cell function. Proc. Natl. Acad. Sci. USA 2020, 117, 11636–11647. [Google Scholar] [CrossRef]
- Augusto, D.G. The impact of KIR polymorphism on the risk of developing cancer: Not as strong as imagined? Front. Genet. 2016, 7, 121. [Google Scholar] [CrossRef]
- Tariq, N.U.; McNamara, M.G.; Valle, J.W. Biliary tract cancers: Current knowledge, clinical candidates and future challenges. Cancer Manag. Res. 2019, 11, 2623–2642. [Google Scholar] [CrossRef]
- Cornillet, M.; Jansson, H.; Schaffer, M.; Hertwig, L.; Berglin, L.; Zimmer, C.L.; Johansson, H.; Ellis, E.; Isaksson, B.; Gonzalez-Galarza, F.F.; et al. Imbalance of genes encoding natural killer immunoglobulin-like receptors and human leukocyte antigen in patients with biliary cancer. Gastroenterology 2019, 157, 1067–1080.e9. [Google Scholar] [CrossRef]
- Lenis, A.T.; Lec, P.M.; Chamie, K.; Mshs, M.D. Bladder cancer: A review. JAMA 2020, 324, 1980–1991. [Google Scholar] [CrossRef]
- Jamali, E.; Barani, S.; Yousefinejad, F.; Ariafar, A.; Talei, G.R.; Ghaderi, A. KIRs gene content diversity in Iranians with urothelial bladder cancer. Mol. Biol. Rep. 2018, 45, 713–719. [Google Scholar] [CrossRef] [PubMed]
- Guillamón, C.F.; Gimeno, L.; Server, G.; Martínez-Sánchez, M.V.; Escudero, J.F.; López-Cubillana, P.; Cabezas-Herrera, J.; Campillo, J.A.; Abellan, D.J.; Martínez-García, J.; et al. Immunological risk stratification of bladder cancer based on peripheral blood natural killer cell biomarkers. Eur. Urol. Oncol. 2021, 4, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Spezia, M.; Huang, S.; Yuan, C.; Zeng, Z.; Zhang, L.; Ji, X.; Liu, W.; Huang, B.; Luo, W.; et al. Breast cancer development and progression: Risk factors, cancer stem cells, signaling pathways, genomics, and molecular pathogenesis. Genes Dis. 2018, 5, 77–106. [Google Scholar] [CrossRef]
- Ozturk, O.G.; Gun, F.D.; Polat, G. Killer cell immunoglobulin-like receptor genes in patients with breast cancer. Med. Oncol. 2012, 29, 511–515. [Google Scholar] [CrossRef]
- Alomar, S.Y.; Alkhuriji, A.; Trayhyrn, P.; Alhetheel, A.; Al-Jurayyan, A.; Mansour, L. Association of the genetic diversity of killer cell immunoglobulin-like receptor genes and HLA-C ligand in Saudi women with breast cancer. Immunogenetics 2017, 69, 69–76. [Google Scholar] [CrossRef]
- Ashouri, E.; Rajalingam, K.; Barani, S.; Farjadian, S.; Ghaderi, A.; Rajalingam, R. Coexistence of inhibitory and activating killer-cell immunoglobulin-like receptors to the same cognate HLA-C2 and Bw4 ligands confer breast cancer risk. Sci. Rep. 2021, 11, 7932. [Google Scholar] [CrossRef]
- Jobim, M.R.; Jobim, M.; Salim, P.H.; Portela, P.; Jobim, L.F.; Leistner-Segal, S.; Bittelbrunn, A.C.; Menke, C.H.; Biazús, J.V.; Roesler, R.; et al. Analysis of KIR gene frequencies and HLA class I genotypes in breast cancer and control group. Hum. Immunol. 2013, 74, 1130–1133. [Google Scholar] [CrossRef] [PubMed]
- Hematian Larki, M.; Barani, S.; Talei, A.R.; Ghaderi, A. Diversity of KIRs in invasive breast cancer patients and healthy controls along with the clinical significance in ER/PR/HER2+ patients. Genes Immun. 2020, 21, 380–389. [Google Scholar] [CrossRef]
- Canossi, A.; Aureli, A.; Del Beato, T.; Novelli, G.; Buonomo, O.; Rossi, P.; Venditti, A.; Papola, F.; Sconocchia, G. Impact of HLA Class I Antigen, Killer Inhibitory Receptor, and FCGR3A Genotypes on Breast Cancer Susceptibility and Tumor Stage. Curr. Mol. Med. 2024, 24, 920–930. [Google Scholar] [CrossRef] [PubMed]
- Okunade, K.S. Human papillomavirus and cervical cancer. J. Obstet. Gynaecol. 2020, 40, 602–608. [Google Scholar] [CrossRef]
- Arnheim, L.; Dillner, J.; Sanjeevi, C.B. A population-based cohort study of KIR genes and genotypes in relation to cervical intraepithelial neoplasia. Tissue Antigens 2005, 65, 252–259. [Google Scholar] [CrossRef]
- Tembhurne, A.K.; Maheshwari, A.; Warke, H.; Chaudhari, H.; Kerkar, S.C.; Deodhar, K.; Rekhi, B.; Mania-Pramanik, J. Killer cell immunoglobulin-like receptor (KIR) gene contents: Are they associated with cervical cancer? J. Med. Virol. 2023, 95, e27873. [Google Scholar] [CrossRef]
- Bao, X.; Hanson, A.L.; Madeleine, M.M.; Wang, S.S.; Schwartz, S.M.; Newell, F.; Pettersson-Kymmer, U.; Hemminki, K.; Tiews, S.; Steinberg, W.; et al. HLA and KIR Associations of Cervical Neoplasia. J. Infect. Dis. 2018, 218, 2006–2015. [Google Scholar] [CrossRef] [PubMed]
- Brestovac, B.; Wong, M.E.; Tjendera, R.; Costantino, P.J.; Mamotte, C.; Witt, C.S. Human papillomavirus, high-grade intraepithelial neoplasia and killer immunoglobulin-like receptors: A Western Australian cohort study. Infect. Agent Cancer 2013, 8, 33. [Google Scholar] [CrossRef]
- Carrington, M.; Wang, S.; Martin, M.P.; Gao, X.; Schiffman, M.; Cheng, J.; Herrero, R.; Rodriguez, A.C.; Kurman, R.; Mortel, R.; et al. Hierarchy of resistance to cervical neoplasia mediated by combinations of killer immunoglobulin-like receptor and human leukocyte antigen loci. J. Exp. Med. 2005, 201, 1069–1075. [Google Scholar] [CrossRef]
- Zhen, J.; Zhu, L.; Li, W.; Hu, H.; Deng, Z.; Xiong, L. A protective effect conferred by KIR3DL1 and its cognate ligand against cervical cancer among ethnic Han Chinese population and its potential mechanism. Zhonghua Yi Xue Yi Chuan Xue Za Zhi 2019, 36, 1035–1038. [Google Scholar] [CrossRef]
- Duan, B.; Zhao, Y.; Bai, J.; Wang, J.; Duan, X.; Luo, X.; Zhang, R.; Pu, Y.; Kou, M.; Lei, J.; et al. Colorectal Cancer: An Overview. In Gastrointestinal Cancers; Morgado-Diaz, J.A., Ed.; Exon Publications: Brisbane, Australia, 2022; Chapter 1. [Google Scholar]
- De Re, V.; Caggiari, L.; De Zorzi, M.; Talamini, R.; Racanelli, V.; D’Andrea, M.; Buonadonna, A.; Zagonel, V.; Cecchin, E.; Innocenti, F.; et al. Genetic diversity of the KIR/HLA system and outcome of patients with metastatic colorectal cancer treated with chemotherapy. PLoS ONE 2014, 9, e84940. [Google Scholar] [CrossRef]
- Kim, H.J.; Choi, H.B.; Jang, J.P.; Baek, I.C.; Choi, E.J.; Park, M.; Kim, T.G.; Oh, S.T. HLA-Cw polymorphism and killer cell immunoglobulin-like receptor (KIR) gene analysis in Korean colorectal cancer patients. Int. J. Surg. 2014, 12, 815–820. [Google Scholar] [CrossRef] [PubMed]
- Ghanadi, K.; Shayanrad, B.; Ahmadi, S.A.; Shahsavar, F.; Eliasy, H. Colorectal cancer and the KIR genes in the human genome: A meta-analysis. Genom. Data 2016, 10, 118–126. [Google Scholar] [CrossRef]
- Portela, P.; Merzoni, J.; Lindenau, J.D.; Damin, D.C.; Wilson, T.J.; Roesler, R.; Schwartsmann, G.; Jobim, L.F.; Jobim, M. KIR genes and HLA class I ligands in a Caucasian Brazilian population with colorectal cancer. Hum. Immunol. 2017, 78, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Barani, S.; Hosseini, S.V.; Ghaderi, A. Activating and inhibitory killer cell immunoglobulin-like receptors (KIR) genes are involved in an increased susceptibility to colorectal adenocarcinoma and protection against invasion and metastasis. Immunobiology 2019, 224, 681–686. [Google Scholar] [CrossRef]
- Diaz-Peña, R.; Mondelo-Macía, P.; Molina de la Torre, A.J.; Sanz-Pamplona, R.; Moreno, V.; Martín, V. Analysis of Killer Immunoglobulin-Like Receptor Genes in Colorectal Cancer. Cells 2020, 9, 514. [Google Scholar] [CrossRef]
- Ge, L.L.; Xing, M.Y.; Zhang, H.B.; Wang, Z.C. Neurofibroma Development in Neurofibromatosis Type 1: Insights from Cellular Origin and Schwann Cell Lineage Development. Cancers 2022, 14, 4513. [Google Scholar] [CrossRef]
- Anastasaki, C.; Dahiya, S.; Gutmann, D.H. KIR2DL5 mutation and loss underlies sporadic dermal neurofibroma pathogenesis and growth. Oncotarget 2017, 8, 47574–47585. [Google Scholar] [CrossRef]
- Shen, C.; Jiang, X.; Li, M.; Luo, Y. Hepatitis Virus and Hepatocellular Carcinoma: Recent Advances. Cancers 2023, 15, 533. [Google Scholar] [CrossRef]
- Pan, N.; Jiang, W.; Sun, H.; Miao, F.; Qiu, J.; Jin, H.; Xu, J.; Shi, Q.; Xie, W.; Zhang, J. KIR and HLA loci are associated with hepatocellular carcinoma development in patients with hepatitis B virus infection: A case-control study. PLoS ONE 2011, 6, e25682. [Google Scholar] [CrossRef]
- Littera, R.; Zamboni, F.; Tondolo, V.; Fantola, G.; Chessa, L.; Orrù, N.; Sanna, M.; Valentini, D.; Cappai, L.; Mulargia, M.; et al. Absence of activating killer immunoglobulin-like receptor genes combined with hepatitis C viral genotype is predictive of hepatocellular carcinoma. Hum. Immunol. 2013, 74, 1288–1294. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Jiao, B.; Hong, W.; Cai, C.; Zhong, Y.; Quan, Z.; Chen, H.; Xu, Y. Distribution of KIR/HLA alleles among ethnic Han Chinese patients with hepatocellular carcinoma from southern China. Zhonghua Yi Xue Yi Chuan Xue Za Zhi 2019, 36, 439–442. [Google Scholar] [CrossRef]
- Abdelmaguid, W.; Maher, D.; Kohla, M.A.S.; Ezzat, S.; Moaz, I.; Abdel-Mageed, W.S.; El-Halfawy, K.A.; Abdel-Rahman, M.H. KIR Genotypes Impact Progression to Hepatocellular Carcinoma in Patients with Chronic Hepatitis C Infection. Livers 2023, 3, 354–368. [Google Scholar] [CrossRef]
- Linehan, W.M.; Schmidt, L.S.; Crooks, D.R.; Wei, D.; Srinivasan, R.; Lang, M.; Ricketts, C.J. The Metabolic Basis of Kidney Cancer. Cancer Discov. 2019, 9, 1006–1021. [Google Scholar] [CrossRef]
- Al Omar, S.; Middleton, D.; Marshall, E.; Porter, D.; Xinarianos, G.; Raji, O.; Field, J.K.; Christmas, S.E. Associations between genes for killer immunoglobulin-like receptors and their ligands in patients with solid tumors. Hum. Immunol. 2010, 71, 976–981. [Google Scholar] [CrossRef] [PubMed]
- Verheyden, S.; Bernier, M.; Demanet, C. Identification of natural killer cell receptor phenotypes associated with leukemia. Leukemia 2004, 18, 2002–2007. [Google Scholar] [CrossRef] [PubMed]
- Middleton, D.; Diler, A.S.; Meenagh, A.; Sleator, C.; Gourraud, P.A. Killer immunoglobulin-like receptors (KIR2DL2 and/or KIR2DS2) in presence of their ligand (HLA-C1 group) protect against chronic myeloid leukaemia. Tissue Antigens 2009, 73, 553–560. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, B.; Ye, S.; Liu, S.; Liu, M.; Shen, C.; Teng, Y.; Qi, J. Killer cell immunoglobulin-like receptor gene polymorphisms in patients with leukemia: Possible association with susceptibility to the disease. Leuk. Res. 2010, 34, 55–58. [Google Scholar] [CrossRef]
- Almalte, Z.; Samarani, S.; Iannello, A.; Debbeche, O.; Duval, M.; Infante-Rivard, C.; Amre, D.K.; Sinnett, D.; Ahmad, A. Novel associations between activating killer-cell immunoglobulin-like receptor genes and childhood leukemia. Blood 2011, 118, 1323–1328. [Google Scholar] [CrossRef]
- Babor, F.; Manser, A.; Schönberg, K.; Enczmann, J.; Borkhardt, A.; Meisel, R.; Uhrberg, M. Lack of association between KIR genes and acute lymphoblastic leukemia in children. Blood 2012, 120, 2770–2772. [Google Scholar] [CrossRef]
- Sugioka, D.K.; Gonçalves, C.E.; Bicalho, M.D. KIR repertory in patients with hematopoietic diseases and healthy family members. BMC Hematol. 2016, 16, 25. [Google Scholar] [CrossRef]
- Al-Tamimi, J.; Alomar, S.; Aljuaimlani, A.; Mansour, L. Association of killer immunoglobulin-like receptor genotypes and haplotypes with acute lymphoblastic leukemia risk. Innate Immun. 2025, 31, 17534259251314774. [Google Scholar] [CrossRef] [PubMed]
- de Smith, A.J.; Walsh, K.M.; Ladner, M.B.; Zhang, S.; Xiao, C.; Cohen, F.; Moore, T.B.; Chokkalingam, A.P.; Metayer, C.; Buffler, P.A.; et al. The role of KIR genes and their cognate HLA class I ligands in childhood acute lymphoblastic leukemia. Blood 2014, 123, 2497–2503. [Google Scholar] [CrossRef]
- Varbanova, V.P.; Mihailova, S.; Naumova, E.; Mihaylova, A.P. Certain Killer Immunoglobulin-Like Receptor (KIR)/KIR HLA Class I Ligand Genotypes Influence Natural Killer Antitumor Activity in Myelogenous Leukemia but Not in Acute Lymphoblastic Leukemia: A Case Control Leukemia Association Study. Turk. J. Haematol. 2019, 36, 238–246. [Google Scholar] [CrossRef]
- Cianga, V.A.; Rusu, C.; Pavel-Tanasa, M.; Dascalescu, A.; Danaila, C.; Harnau, S.; Aanei, C.M.; Cianga, P. Combined flow cytometry natural killer immunophenotyping and KIR/HLA-C genotyping reveal remarkable differences in acute myeloid leukemia patients, but suggest an overall impairment of the natural killer response. Front. Med. 2023, 10, 1148748. [Google Scholar] [CrossRef] [PubMed]
- Heidenreich, F.; Falk, B.; Baldauf, H.; Massalski, C.; Schäfer, G.; Rücker-Braun, E.; Altmann, H.; Sauter, J.; Solloch, U.V.; Lange, V.; et al. Large case-control study indicates no association of KIR genotype and risk of developing acute myeloid leukemia. Blood Adv. 2023, 7, 2994–3004. [Google Scholar] [CrossRef]
- Siddiqui, F.; Vaqar, S.; Siddiqui, A.H. Lung Cancer. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2025; Available online: https://www.statpearls.com (accessed on 8 May 2023).
- Polański, J.; Chabowski, M.; Jankowska-Polańska, B.; Janczak, D.; Rosińczuk, J. Histological subtype of lung cancer affects acceptance of illness, severity of pain, and quality of life. J. Pain Res. 2018, 11, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Wiśniewski, A.; Jankowska, R.; Passowicz-Muszyńska, E.; Wiśniewska, E.; Majorczyk, E.; Nowak, I.; Frydecka, I.; Kuśnierczyk, P. KIR2DL2/S2 and HLA-C C1C1 genotype is associated with better response to treatment and prolonged survival of patients with non-small cell lung cancer in a Polish Caucasian population. Hum. Immunol. 2012, 73, 927–931. [Google Scholar] [CrossRef]
- Li, Y.; Liu, S.; Hong, C.; Ma, Q.; Tan, F.; Liu, C.; Kuśnierczyk, P.; Li, C.; Shi, L.; Yao, Y. The association of HLA/KIR genes with non-small cell lung cancer (adenocarcinoma) in a Han Chinese population. J. Cancer 2019, 10, 4731–4738. [Google Scholar] [CrossRef]
- Yu, H.; Liu, F.; Sansas, B.; Kang, B.; Preville, X.; Wu, X.; Chang, J.; Micol, R.; Wang, J.; Meng, X. Typing of killer-cell immunoglobulin-like receptors and their cognate human leukocyte antigen class I ligands predicts survival of Chinese Han patients with metastatic non-small-cell lung cancer. Mol. Clin. Oncol. 2017, 6, 279–285. [Google Scholar] [CrossRef]
- Hematian Larki, M.; Ashouri, E.; Barani, S.; Ghayumi, S.M.A.; Ghaderi, A.; Rajalingam, R. KIR-HLA gene diversities and susceptibility to lung cancer. Sci. Rep. 2022, 12, 17237. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, B.; Qadir, M.I.; Ghafoor, S. Malignant Melanoma: Skin Cancer-Diagnosis, Prevention, and Treatment. Crit. Rev. Eukaryot. Gene Expr. 2020, 30, 291–297. [Google Scholar] [CrossRef]
- Campillo, J.A.; Legaz, I.; López-Álvarez, M.R.; Bolarín, J.M.; Las Heras, B.; Muro, M.; Minguela, A.; Moya-Quiles, M.R.; Blanco-García, R.; Martínez-Banaclocha, H.; et al. KIR gene variability in cutaneous malignant melanoma: Influence of KIR2D/HLA-C pairings on disease susceptibility and prognosis. Immunogenetics 2013, 65, 333–343. [Google Scholar] [CrossRef]
- Naumova, E.; Mihaylova, A.; Ivanova, M.; Mihailova, S. Impact of KIR/HLA ligand combinations on immune responses in malignant melanoma. Cancer Immunol. Immunother. 2007, 56, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Kandilarova, S.M.; Paschen, A.; Mihaylova, A.; Ivanova, M.; Schadendorf, D.; Naumova, E. The influence of HLA and KIR genes on malignant melanoma development and progression. Arch. Immunol. Ther. Exp. 2016, 64 (Suppl. S1), 73–81. [Google Scholar] [CrossRef] [PubMed]
- Fauriat, C.; Mallet, F.; Olive, D.; Costello, R.T. Impaired activating receptor expression pattern in natural killer cells from patients with multiple myeloma. Leukemia 2006, 20, 732–733. [Google Scholar] [CrossRef]
- Frohn, C.; Höppner, M.; Schlenke, P.; Kirchner, H.; Koritke, P.; Luhm, J. Anti-myeloma activity of natural killer lymphocytes. Br. J. Haematol. 2002, 119, 660–664. [Google Scholar] [CrossRef]
- Jurisic, V.; Srdic, T.; Konjevic, G.; Markovic, O.; Colovic, M. Clinical stage-depending decrease of NK cell activity in multiple myeloma patients. Med. Oncol. 2007, 24, 312–317. [Google Scholar] [CrossRef]
- Hoteit, R.; Bazarbachi, A.; Antar, A.; Salem, Z.; Shammaa, D.; Mahfouz, R. KIR genotype distribution among patients with multiple myeloma: Higher prevalence of KIR 2DS4 and KIR 2DS5 genes. Meta Gene 2014, 2, 730–736. [Google Scholar] [CrossRef]
- Theeranawakam, A.; Vejbaesya, S.; Khuhapinant, A.; Sae-Tam, P. Killer cell immunoglobulin-like receptors in Thai patients with multiple myeloma. Asian Pac. J. Allergy Immunol. 2024, 42, 77–80. [Google Scholar] [CrossRef]
- Beelen, N.A.; Molenbroeck, S.J.J.; Groeneveld, L.; Voorter, C.E.; Bos, G.M.J.; Wieten, L. HLA class I NK-epitopes and KIR diversities in patients with multiple myeloma. Immunogenetics 2024, 76, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.J.; Hess, S.; Richardson, S.K.; Newton, S.; Showe, L.C.; Benoit, B.M.; Ubriani, R.; Vittorio, C.C.; Junkins-Hopkins, J.M.; Wysocka, M.; et al. Immunopathogenesis and therapy of cutaneous T cell lymphoma. J. Clin. Investig. 2005, 115, 798–812. [Google Scholar] [CrossRef] [PubMed]
- Brazzelli, V.; Rivetti, N.; Badulli, C.; Carugno, A.; Cananzi, R.; De Silvestri, A.; Martinetti, M.; Borroni, G. Mycosis fungoides: Association of KIR ligands and HLA-DQB1*05 with bad prognosis of the disease. J. Eur. Acad. Dermatol. Venereol. 2016, 30, 266–269. [Google Scholar] [CrossRef]
- Sezgin, G.; Görüroğlu Öztürk, Ö.; Özkan, A.; Küpeli, S.; Bayram, İ. Clinical Impact of KIR2DS3 and KIR2DL3 Genes in Neuroblastoma Patients. Med. Princ. Pract. 2022, 31, 532–539. [Google Scholar] [CrossRef]
- Tarek, N.; Le Luduec, J.B.; Gallagher, M.M.; Zheng, J.; Venstrom, J.M.; Chamberlain, E.; Modak, S.; Heller, G.; Dupont, B.; Cheung, N.K.; et al. Unlicensed NK cells target neuroblastoma following anti-GD2 antibody treatment. J. Clin. Investig. 2012, 122, 3260–3270. [Google Scholar] [CrossRef]
- Cheung, N.K.; Cheung, I.Y.; Kushner, B.H.; Ostrovnaya, I.; Chamberlain, E.; Kramer, K.; Modak, S. Murine anti-GD2 monoclonal antibody 3F8 combined with granulocyte-macrophage colony-stimulating factor and 13-cis-retinoic acid in high-risk patients with stage 4 neuroblastoma in first remission. J. Clin. Oncol. 2012, 30, 3264–3270. [Google Scholar] [CrossRef] [PubMed]
- Keating, S.E.; Ní Chorcora, C.; Dring, M.M.; Stallings, R.L.; O’Meara, A.; Gardiner, C.M. Increased frequencies of the killer immunoglobulin-like receptor genes KIR2DL2 and KIR2DS2 are associated with neuroblastoma. Tissue Antigens 2015, 86, 172–177. [Google Scholar] [CrossRef]
- Siebert, N.; Jensen, C.; Troschke-Meurer, S.; Zumpe, M.; Jüttner, M.; Ehlert, K.; Kietz, S.; Müller, I.; Lode, H.N. Neuroblastoma patients with high-affinity FCGR2A, -3A and stimulatory KIR 2DS2 treated by long-term infusion of anti-GD2 antibody ch14.18/CHO show higher ADCC levels and improved event-free survival. Oncoimmunology 2016, 5, e1235108. [Google Scholar] [CrossRef] [PubMed]
- Sapkota, S.; Shaikh, H. Non-Hodgkin Lymphoma. Available online: https://www.ncbi.nlm.nih.gov/books/NBK559328/ (accessed on 10 January 2025).
- Pamuk, G.E.; Tozkir, H.; Uyanik, M.S.; Gurkan, H.; Duymaz, J.; Pamuk, O.N. Natural killer cell killer immunoglobulin-like gene receptor polymorphisms in non-Hodgkin lymphoma: Possible association with clinical course. Leuk Lymphoma 2015, 56, 2902–2907. [Google Scholar] [CrossRef]
- Daniela, M.C.; Amanda, V.M.; Fernando, G.; Silvia, M.; Sofia, L.; Márcia, D.; Francisco José Penteado, A.; Jeane Eliete Laguila, V.; Cármino Antonio, S. Association of KIR Genes Polymorphism and its HLA Ligands in Diffuse Large B-cell Lymphoma. Clin. Lymphoma Myeloma Leuk. 2023, 23, 438–445. [Google Scholar] [CrossRef]
- Cives, M.; Mannavola, F.; Lospalluti, L.; Sergi, M.C.; Cazzato, G.; Filoni, E.; Cavallo, F.; Giudice, G.; Stucci, L.S.; Porta, C.; et al. Non-Melanoma Skin Cancers: Biological and Clinical Features. Int. J. Mol. Sci. 2020, 21, 5394. [Google Scholar] [CrossRef] [PubMed]
- de Souza Fernandez Pereira, M.; Carr, D.R.; Gatti-Mays, M.E.; Olsen, M.R.; Setty, B.A.; Shahwan, K.T.; Lee, D.A. Natural Killer Cell Recognition and Control of Epithelial Cancers. Cancer J. 2022, 28, 263–269. [Google Scholar] [CrossRef]
- Yousefinejad, F.; Jowkar, F.; Barani, S.; Jamali, E.; Mahmoudi, E.; Ramezani, A.; Mahmoudi Maymand, E.; Ghaderi, A. Killer Cell Immunoglobulin-Like Receptors (KIRs) Genotype and Haplotype Analysis in Iranians with Non-Melanoma Skin Cancers. Iran. Biomed. J. 2019, 23, 330–337. [Google Scholar] [CrossRef]
- Vineretsky, K.A.; Karagas, M.R.; Christensen, B.C.; Kuriger-Laber, J.K.; Perry, A.E.; Storm, C.A.; Nelson, H.H. Skin Cancer Risk Is Modified by KIR/HLA Interactions That Influence the Activation of Natural Killer Immune Cells. Cancer Res. 2016, 76, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Kurman, R.J.; Shih, I.e.M. The origin and pathogenesis of epithelial ovarian cancer: A proposed unifying theory. Am. J. Surg. Pathol. 2010, 34, 433–443. [Google Scholar] [CrossRef]
- Giebel, S.; Boratyn-Nowicka, A.; Karabon, L.; Jedynak, A.; Pamula-Pilat, J.; Tecza, K.; Kula, D.; Kowal, M.; Frydecka, I.; Grzybowska, E. Associations between genes for killer immunoglobulin-like receptors and their ligands in patients with epithelial ovarian cancer. Hum. Immunol. 2014, 75, 508–513. [Google Scholar] [CrossRef] [PubMed]
- Ashouri, E.; Dabbaghmanesh, M.H.; Rowhanirad, S.; Bakhshayeshkaram, M.; Ranjbar Omrani, G.; Ghaderi, A. Activating KIR2DS5 receptor is a risk for thyroid cancer. Hum. Immunol. 2012, 73, 1017–1022. [Google Scholar] [CrossRef]
- Ligotti, M.E.; Aiello, A.; Accardi, G.; Calabrò, A.; Ciaccio, M.; Colomba, C.; Di Bona, D.; Lo Sasso, B.; Pojero, F.; Tuttolomondo, A.; et al. Distribution of KIR Genes and Their HLA Ligands in Different Viral Infectious Diseases: Frequency Study in Sicilian Population. Int. J. Mol. Sci. 2022, 23, 15466. [Google Scholar] [CrossRef]
- Aiello, A.; Candore, G.; Accardi, G.; Caruso, C.; Colomba, C.; Duro, G.; Gambino, C.M.; Ligotti, M.E.; Di Bona, D. Translation of Basic Research into Clinics: Killer Immunoglobulin-like Receptors Genes in Autoimmune and Infectious Diseases. Curr. Pharm. Des. 2018, 24, 3113–3122. [Google Scholar] [CrossRef]
- Charoudeh, H.N.; Terszowski, G.; Czaja, K.; Gonzalez, A.; Schmitter, K.; Stern, M. Modulation of the natural killer cell KIR repertoire by cytomegalovirus infection. Eur. J. Immunol. 2013, 43, 480–487. [Google Scholar] [CrossRef]
- Djaoud, Z.; David, G.; Bressollette, C.; Willem, C.; Rettman, P.; Gagne, K.; Legrand, N.; Mehlal, S.; Cesbron, A.; Imbert-Marcille, B.M.; et al. Amplified NKG2C+ NK cells in cytomegalovirus (CMV) infection preferentially express killer cell Ig-like receptor 2DL: Functional impact in controlling CMV-infected dendritic cells. J. Immunol. 2013, 191, 2708–2716. [Google Scholar] [CrossRef] [PubMed]
- Béziat, V.; Liu, L.L.; Malmberg, J.A.; Ivarsson, M.A.; Sohlberg, E.; Björklund, A.T.; Retière, C.; Sverremark-Ekström, E.; Traherne, J.; Ljungman, P.; et al. NK cell responses to cytomegalovirus infection lead to stable imprints in the human KIR repertoire and involve activating KIRs. Blood 2013, 121, 2678–2688. [Google Scholar] [CrossRef] [PubMed]
- Di Bona, D.; Scafidi, V.; Plaia, A.; Colomba, C.; Nuzzo, D.; Occhino, C.; Tuttolomondo, A.; Giammanco, G.; De Grazia, S.; Montalto, G.; et al. HLA and killer cell immunoglobulin-like receptors influence the natural course of CMV infection. J. Infect. Dis. 2014, 210, 1083–1089. [Google Scholar] [CrossRef] [PubMed]
- Di Bona, D.; Accardi, G.; Aiello, A.; Bilancia, M.; Candore, G.; Colomba, C.; Caruso, C.; Duro, G.; Gambino, C.M.; Macchia, L.; et al. Association between γ marker, human leucocyte antigens and killer immunoglobulin-like receptors and the natural course of human cytomegalovirus infection: A pilot study performed in a Sicilian population. Immunology 2018, 153, 523–531. [Google Scholar] [CrossRef]
- Crespo, Â.C.; Strominger, J.L.; Tilburgs, T. Expression of KIR2DS1 by decidual natural killer cells increases their ability to control placental HCMV infection. Proc. Natl. Acad. Sci. USA 2016, 113, 15072–15077. [Google Scholar] [CrossRef]
- Cook, M.; Briggs, D.; Craddock, C.; Mahendra, P.; Milligan, D.; Fegan, C.; Darbyshire, P.; Lawson, S.; Boxall, E.; Moss, P. Donor KIR genotype has a major influence on the rate of cytomegalovirus reactivation following T-cell replete stem cell transplantation. Blood 2006, 107, 1230–1232. [Google Scholar] [CrossRef]
- Zaia, J.A.; Sun, J.Y.; Gallez-Hawkins, G.M.; Thao, L.; Oki, A.; Lacey, S.F.; Dagis, A.; Palmer, J.; Diamond, D.J.; Forman, S.J.; et al. The effect of single and combined activating killer immunoglobulin-like receptor genotypes on cytomegalovirus infection and immunity after hematopoietic cell transplantation. Biol. Blood Marrow Transplant. 2009, 15, 315–325. [Google Scholar] [CrossRef]
- Wang, S.; He, B.; Liu, H.; Muhammad, I.; Cai, J.; Wang, F. Immunoglobulin-like receptor genotype-associated protection from cytomegalovirus infection after liver transplantation. Transpl. Immunol. 2025, 88, 102171. [Google Scholar] [CrossRef]
- Moroso, V.; van der Meer, A.; Tilanus, H.W.; Kazemier, G.; van der Laan, L.J.; Metselaar, H.J.; Joosten, I.; Kwekkeboom, J. Donor and recipient HLA/KIR genotypes do not predict liver transplantation outcome. Transpl. Int. 2011, 24, 932–942. [Google Scholar]
- Beck, J.C.; Wagner, J.E.; DeFor, T.E.; Brunstein, C.G.; Schleiss, M.R.; Young, J.A.; Weisdorf, D.H.; Cooley, S.; Miller, J.S.; Verneris, M.R. Impact of cytomegalovirus (CMV) reactivation after umbilical cord blood transplantation. Biol. Blood Marrow Transplant. 2010, 16, 215–222. [Google Scholar] [CrossRef]
- Hadaya, K.; de Rham, C.; Bandelier, C.; Ferrari-Lacraz, S.; Jendly, S.; Berney, T.; Buhler, L.; Kaiser, L.; Seebach, J.D.; Tiercy, J.M.; et al. Natural killer cell receptor repertoire and their ligands, and the risk of CMV infection after kidney transplantation. Am. J. Transplant. 2008, 8, 2674–2683. [Google Scholar] [CrossRef] [PubMed]
- van Duin, D.; Avery, R.K.; Hemachandra, S.; Yen-Lieberman, B.; Zhang, A.; Jain, A.; Butler, R.S.; Barnard, J.; Schold, J.D.; Fung, J.; et al. KIR and HLA interactions are associated with control of primary CMV infection in solid organ transplant recipients. Am. J. Transplant. 2014, 14, 156–162. [Google Scholar] [CrossRef]
- Stern, M.; Elsässer, H.; Hönger, G.; Steiger, J.; Schaub, S.; Hess, C. The number of activating KIR genes inversely correlates with the rate of CMV infection/reactivation in kidney transplant recipients. Am. J. Transplant. 2008, 8, 1312–1317. [Google Scholar] [CrossRef] [PubMed]
- Stern, M.; Hadaya, K.; Hönger, G.; Martin, P.Y.; Steiger, J.; Hess, C.; Villard, J. Telomeric rather than centromeric activating KIR genes protect from cytomegalovirus infection after kidney transplantation. Am. J. Transplant. 2011, 11, 1302–1307. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.C.; Peacock, S.; Hughes, D.; Traherne, J.A.; Allen, R.L.; Barnardo, M.C.; Friend, P.; Taylor, C.J.; Fuggle, S.; Trowsdale, J.; et al. Killer immunoglobulin-like receptor gene repertoire influences viral load of primary human cytomegalovirus infection in renal transplant patients. Genes Immun. 2014, 15, 562–568. [Google Scholar] [CrossRef]
- Nakamura, R.; Gendzekhadze, K.; Palmer, J.; Tsai, N.C.; Mokhtari, S.; Forman, S.J.; Zaia, J.A.; Senitzer, D.; Marcucci, G.; Stein, A. Influence of donor KIR genotypes on reduced relapse risk in acute myelogenous leukemia after hematopoietic stem cell transplantation in patients with CMV reactivation. Leuk. Res. 2019, 87, 106230. [Google Scholar] [CrossRef]
- Farzamikia, N.; Hejazian, S.M.; Haghi, M.; Hejazian, S.S.; Zununi Vahed, S.; Ardalan, M. Evaluation of telomeric KIR genes and their association with CMV infection in kidney transplant recipients. Immunogenetics 2022, 74, 207–212. [Google Scholar] [CrossRef]
- Gao, F.; Shi, Z.; Shi, J.; Luo, Y.; Yu, J.; Fu, H.; Lai, X.; Liu, L.; Yuan, Z.; Zheng, Z.; et al. Donor aKIR genes influence the risk of EBV and CMV reactivation after anti-thymocyte globulin-based haploidentical hematopoietic stem cell transplantation. HLA 2024, 103, e15320. [Google Scholar] [CrossRef]
- Chen, C.; Busson, M.; Rocha, V.; Appert, M.L.; Lepage, V.; Dulphy, N.; Haas, P.; Socié, G.; Toubert, A.; Charron, D.; et al. Activating KIR genes are associated with CMV reactivation and survival after non-T-cell depleted HLA-identical sibling bone marrow transplantation for malignant disorders. Bone Marrow Transplant. 2006, 38, 437–444. [Google Scholar] [CrossRef]
- Gallez-Hawkins, G.M.; Franck, A.E.; Li, X.; Thao, L.; Oki, A.; Gendzekhadze, K.; Dagis, A.; Palmer, J.; Nakamura, R.; Forman, S.J.; et al. Expression of activating KIR2DS2 and KIR2DS4 genes after hematopoietic cell transplantation: Relevance to cytomegalovirus infection. Biol. Blood Marrow Transplant. 2011, 17, 1662–1672. [Google Scholar] [CrossRef]
- Deborska-Materkowska, D.; Perkowska-Ptasinska, A.; Sadowska-Jakubowicz, A.; Gozdowska, J.; Ciszek, M.; Pazik, J.; Ostaszewska, A.; Kosieradzki, M.; Nowak, J.; Durlik, M. Killer immunoglobulin-like receptor 2DS2 (KIR2DS2), KIR2DL2-HLA-C1, and KIR2DL3 as genetic markers for stratifying the risk of cytomegalovirus infection in kidney transplant recipients. Int. J. Mol. Sci. 2019, 20, 546. [Google Scholar] [CrossRef]
- Wu, X.; Yao, Y.; Bao, X.; Zhou, H.; Tang, X.; Han, Y.; Ma, X.; Liu, Y.; Chen, J.; Zhou, H.; et al. KIR2DS4 and its variant KIR1D are associated with acute graft-versus-host disease, cytomegalovirus, and overall survival after sibling-related HLA-matched transplantation in patients with donors with KIR gene haplotype A. Biol. Blood Marrow Transplant. 2016, 22, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Wauquier, N.; Padilla, C.; Becquart, P.; Leroy, E.; Vieillard, V. Association of KIR2DS1 and KIR2DS3 with fatal outcome in Ebola virus infection. Immunogenetics 2010, 62, 767–771. [Google Scholar] [CrossRef]
- Wawina-Bokalanga, T.; Vanmechelen, B.; Lhermitte, V.; Martí-Carreras, J.; Vergote, V.; Koundouno, F.R.; Akoi-Boré, J.; Thom, R.; Tipton, T.; Steeds, K.; et al. Human diversity of killer cell immunoglobulin-like receptors and human leukocyte antigen class I alleles and Ebola virus disease outcomes. Emerg. Infect. Dis. 2021, 27, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Huo, L.; Jiang, M.Y.; Li, Q.; Zhu, Y.P. Novel association of killer cell immunoglobulin-like receptor genes with EBV-infectious diseases in children. Biomed. Environ. Sci. 2015, 28, 303–307. [Google Scholar] [CrossRef]
- Wang, X.; Liu, X.F.; Shang, Q.N.; Yu, X.X.; Fan, Z.Y.; Cao, X.H.; Huo, M.R.; Chang, Y.J.; Zhao, X.S.; Wang, Y.; et al. Donor activating killer cell immunoglobulin-like receptors genes correlated with Epstein-Barr virus reactivation after haploidentical haematopoietic stem cell transplantation. Br. J. Haematol. 2022, 196, 1007–1017. [Google Scholar] [CrossRef] [PubMed]
- Muriuki, B.M.; Forconi, C.S.; Oluoch, P.O.; Bailey, J.A.; Ghansah, A.; Moormann, A.M.; Ong’echa, J.M. Association of killer cell immunoglobulin-like receptors with endemic Burkitt lymphoma in Kenyan children. Sci. Rep. 2021, 11, 11343. [Google Scholar] [CrossRef]
- Jiang, P.; Nolte, I.M.; Hepkema, B.G.; Stulp, M.; van den Berg, A.; Diepstra, A. Killer Cell Immunoglobulin-Like Receptor Haplotype B Modulates Susceptibility to EBV-Associated Classic Hodgkin Lymphoma. Front. Immunol. 2022, 13, 829943. [Google Scholar] [CrossRef]
- Hsu, Y.C.; Huang, D.Q.; Nguyen, M.H. Global burden of hepatitis B virus: Current status, missed opportunities and a call for action. Nat. Rev. Gastroenterol. Hepatol. 2023, 20, 524–537. [Google Scholar] [CrossRef]
- Zhi-ming, L.; Yu-lian, J.; Zhao-lei, F.; Chun-xiao, W.; Zhen-fang, D.; Bing-chang, Z.; Yue-ran, Z. Polymorphisms of killer cell immunoglobulin-like receptor gene: Possible association with susceptibility to or clearance of hepatitis B virus infection in Chinese Han population. Croat. Med. J. 2007, 48, 800–806. [Google Scholar] [CrossRef]
- Kibar, F.; Goruroglu Ozturk, O.; Ulu, A.; Erken, E.; Inal, S.; Dinkci, S.; Kurtaran, B.; Tasova, Y.; Aksu, H.S.; Yaman, A. Role of KIR genes and genotypes in susceptibility to or protection against hepatitis B virus infection in a Turkish cohort. Med. Sci. Monit. 2014, 20, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Di Bona, D.; Aiello, A.; Colomba, C.; Bilancia, M.; Accardi, G.; Rubino, R.; Giannitrapani, L.; Tuttolomondo, A.; Cascio, A.; Caiaffa, M.F.; et al. KIR2DL3 and the KIR ligand groups HLA-A-Bw4 and HLA-C2 predict the outcome of hepatitis B virus infection. J. Viral Hepat. 2017, 24, 768–775. [Google Scholar] [CrossRef] [PubMed]
- Torimiro, J.; Yengo, C.K.; Bimela, J.S.; Tiedeu, A.B.; Lebon, P.A.; Sake, C.S.; Kouanfack, C.; Nchinda, G.; Rowland-Jones, S.; Yindom, L.M. Killer cell immunoglobulin-like receptor genotypes and haplotypes contribute to susceptibility to hepatitis B virus and hepatitis C virus infection in Cameroon. OMICS 2020, 24, 110–115. [Google Scholar] [CrossRef]
- Sorgho, P.A.; Martinson, J.J.; Djigma, F.W.; Yonli, A.T.; Nagalo, B.M.; Compaore, T.R.; Obiri-Yeboah, D.; Diarra, B.; Sombie, H.K.; Zongo, A.W.; et al. Insights into the interplay between KIR gene frequencies and chronic HBV infection in Burkina Faso. Mediterr. J. Hematol. Infect. Dis. 2018, 10, e2018060. [Google Scholar] [CrossRef]
- Auer, E.D.; Tong, H.V.; Amorim, L.M.; Malheiros, D.; Hoan, N.X.; Issler, H.C.; Petzl-Erler, M.L.; Beltrame, M.H.; Boldt, A.B.W.; Toan, N.L.; et al. Natural killer cell receptor variants and chronic hepatitis B virus infection in the Vietnamese population. Int. J. Infect. Dis. 2020, 96, 541–547. [Google Scholar] [CrossRef]
- Bazie, M.M.; Djigma, F.W.; Sanou, M.; Sorgho, P.A.; Ouattara, A.K.; Obiri-Yeboah, D.; Kapieko, N.; Sombie, H.K.; Bado, P.; Yelemkoure, E.T.; et al. Killer cell immunoglobulin-like receptor alleles influence susceptibility to occult hepatitis B infection in West African population. J. Public Health Afr. 2023, 14, 2586. [Google Scholar] [CrossRef] [PubMed]
- Yindom, L.M.; Mendy, M.; Bodimeade, C.; Chambion, C.; Aka, P.; Whittle, H.C.; Rowland-Jones, S.L.; Walton, R. KIR content genotypes associate with carriage of hepatitis B surface antigen, e antigen and HBV viral load in Gambians. PLoS ONE 2017, 12, e0188307. [Google Scholar] [CrossRef]
- Shah-Hosseini, A.; Jafari, M.; Mohammadi, A.; Sanaei, R.; Alavian, S.M.; Doosti-Irani, A.; Nooradeh Keykavousi, M.; Tajik, N. The impact of KIR-HLA genotype on hepatitis B virus clearance in Iranian infected individuals. Med. Microbiol. Immunol. 2017, 206, 463–470. [Google Scholar] [CrossRef]
- Li, W.; Shen, X.; Fu, B.; Guo, C.; Liu, Y.; Ye, Y.; Sun, R.; Li, J.; Tian, Z.; Wei, H. KIR3DS1/HLA-B Bw4-80Ile genotype is correlated with the IFN-α therapy response in hepatitis B e antigen-positive chronic hepatitis B. Front. Immunol. 2017, 8, 1285. [Google Scholar] [CrossRef]
- Zhuang, Y.L.; Li, X.X.; Xu, H.C.; Ye, H.; Sun, D.; Liu, X.Z.; Ren, G.J. Association between KIR genes and efficacy of treatment of HBeAg-positive chronic hepatitis B patients with entecavir. Iran. J. Immunol. 2018, 15, 112–121. [Google Scholar]
- Khakoo, S.I.; Thio, C.L.; Martin, M.P.; Brooks, C.R.; Gao, X.; Astemborski, J.; Cheng, J.; Goedert, J.J.; Vlahov, D.; Hilgartner, M.; et al. HLA and NK cell inhibitory receptor genes in resolving hepatitis C virus infection. Science 2004, 305, 872–874. [Google Scholar] [CrossRef] [PubMed]
- Knapp, S.; Warshow, U.; Hegazy, D.; Brackenbury, L.; Guha, I.N.; Fowell, A.; Little, A.M.; Alexander, G.J.; Rosenberg, W.M.; Cramp, M.E.; et al. Consistent beneficial effects of killer cell immunoglobulin-like receptor 2DL3 and group 1 human leukocyte antigen-C following exposure to hepatitis C virus. Hepatology 2010, 51, 1168–1175. [Google Scholar] [CrossRef]
- Paladino, N.; Flores, A.C.; Marcos, C.Y.; Fainboim, H.; Theiler, G.; Arruvito, L.; Williams, F.; Middleton, D.; Fainboim, L. Increased frequencies of activating natural killer receptors are associated with liver injury in individuals who do not eliminate hepatitis C virus. Tissue Antigens 2007, 69 (Suppl. S1), 109–111. [Google Scholar] [CrossRef] [PubMed]
- Rauch, A.; Laird, R.; McKinnon, E.; Telenti, A.; Furrer, H.; Weber, R.; Smillie, D.; Gaudieri, S.; Swiss HIV Cohort Study. Influence of inhibitory killer immunoglobulin-like receptors and their HLA-C ligands on resolving hepatitis C virus infection. Tissue Antigens 2007, 69 (Suppl. S1), 237–240. [Google Scholar] [CrossRef]
- Marangon, A.V.; Silva, G.F.; de Moraes, C.F.; Grotto, R.M.; Pardini, M.I.; de Pauli, D.S.; Sell, A.M.; Visentainer, J.E.; Moliterno, R.A. KIR genes and their human leukocyte antigen ligands in the progression to cirrhosis in patients with chronic hepatitis C. Hum. Immunol. 2011, 72, 1074–1078. [Google Scholar] [CrossRef]
- Zúñiga, J.; Romero, V.; Azocar, J.; Terreros, D.; Vargas-Rojas, M.I.; Torres-García, D.; Jiménez-Alvarez, L.; Vargas-Alarcón, G.; Granados-Montiel, J.; Husain, Z.; et al. Protective KIR-HLA interactions for HCV infection in intravenous drug users. Mol. Immunol. 2009, 46, 2723–2727. [Google Scholar] [CrossRef] [PubMed]
- de Vasconcelos, J.M.; de Jesus Maués Pereira Móia, L.; Amaral, I.S.; Miranda, E.C.; Cicalisetakeshita, L.Y.; de Oliveira, L.F.; de Araújo Melo Mendes, L.; Sastre, D.; Tamegão-Lopes, B.P.; de Aquino Pedroza, L.S.; et al. Association of killer cell immunoglobulin-like receptor polymorphisms with chronic hepatitis C and responses to therapy in Brazil. Genet. Mol. Biol. 2013, 36, 22–27. [Google Scholar] [CrossRef]
- Kuśnierczyk, P.; Mozer-Lisewska, I.; Zwolińska, K.; Kowala-Piaskowska, A.E.; Bura, M.; Bereszyńska, I.; Pauli, A.; Żeromski, J. Contribution of genes for killer cell immunoglobulin-like receptors (KIR) to the susceptibility to chronic hepatitis C virus infection and to viremia. Hum. Immunol. 2015, 76, 102–108. [Google Scholar] [CrossRef]
- Gauthiez, E.; Habfast-Robertson, I.; Rüeger, S.; Kutalik, Z.; Aubert, V.; Berg, T.; Cerny, A.; Gorgievski, M.; George, J.; Heim, M.H.; et al. A systematic review and meta-analysis of HCV clearance. Liver Int. 2017, 37, 1431–1445. [Google Scholar] [CrossRef]
- Ruiz-Extremera, A.; Pavón-Castillero, E.J.; Florido, M.; Muñoz de Rueda, P.; Muñoz-Gámez, J.A.; Casado, J.; Carazo, A.; Quiles, R.; Jiménez-Ruiz, S.M.; Gila, A.; et al. Influence of HLA class I, HLA class II and KIRs on vertical transmission and chronicity of hepatitis C virus in children. PLoS ONE 2017, 12, e0172527. [Google Scholar] [CrossRef]
- Podhorzer, A.; Dirchwolf, M.; Machicote, A.; Belen, S.; Montal, S.; Paz, S.; Fainboim, H.; Podestá, L.G.; Fainboim, L. The clinical features of patients with chronic hepatitis C virus infections are associated with killer cell immunoglobulin-like receptor genes and their expression on the surface of natural killer cells. Front. Immunol. 2018, 8, 1912. [Google Scholar] [CrossRef]
- Shan, Z.; Huang, J.; Liao, Q.; Huang, K.; Wang, M.; Xu, R.; Tang, X.; Zhang, W.; Nelson, K.; Fu, Y.; et al. Association of killer cell immunoglobulin-like receptors with spontaneous clearance of hepatitis C virus in the Chinese population. Transfusion 2018, 58, 1028–1035. [Google Scholar] [CrossRef]
- Shen, C.; Ge, Z.; Dong, C.; Wang, C.; Shao, J.; Cai, W.; Huang, P.; Fan, H.; Li, J.; Zhang, Y.; et al. Genetic variants in KIR/HLA-C genes are associated with the susceptibility to HCV infection in a high-risk Chinese population. Front. Immunol. 2021, 12, 632353. [Google Scholar] [CrossRef]
- Feng, Z.; Huang, P.; Zhang, J.; Xia, X.; Zhang, A.M.; Zeng, T.; Chen, Q.; Zhu, C.; Tan, W.; Zhang, Y.; et al. KIR2DL4/HLA-G polymorphisms were associated with HCV infection susceptibility among Chinese high-risk population. J. Med. Virol. 2023, 95, e28645. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zeng, T.; Huang, P.; Tan, W.; Feng, Y.; Xia, X.; Feng, Z.; Shen, C.; Fan, H.; Zhu, C.; et al. Investigating the role of killer cell immunoglobulin-like receptors and human leukocyte antigen genetic variants in hepatitis C virus infection. J. Med. Virol. 2024, 96, e29776. [Google Scholar] [CrossRef]
- Estefanía, E.; Gómez-Lozano, N.; Portero, F.; de Pablo, R.; Solís, R.; Sepúlveda, S.; Vaquero, M.; González, M.A.; Suárez, E.; Roustán, G.; et al. Influence of KIR gene diversity on the course of HSV-1 infection: Resistance to the disease is associated with the absence of KIR2DL2 and KIR2DS2. Tissue Antigens 2007, 70, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Moraru, M.; Cisneros, E.; Gómez-Lozano, N.; de Pablo, R.; Portero, F.; Cañizares, M.; Vaquero, M.; Roustán, G.; Millán, I.; López-Botet, M.; et al. Host genetic factors in susceptibility to herpes simplex type 1 virus infection: Contribution of polymorphic genes at the interface of innate and adaptive immunity. J. Immunol. 2012, 188, 4412–4420. [Google Scholar] [CrossRef] [PubMed]
- Magaret, A.; Dong, L.; John, M.; Mallal, S.A.; James, I.; Warren, T.; Gaudieri, S.; Koelle, D.M.; Wald, A. HLA Class I and II alleles, heterozygosity and HLA-KIR interactions are associated with rates of genital HSV shedding and lesions. Genes Immun. 2016, 17, 412–418. [Google Scholar] [CrossRef]
- Tuttolomondo, A.; Di Raimondo, D.; Vasto, S.; Casuccio, A.; Colomba, C.; Norrito, R.L.; Di Bona, D.; Arnao, V.; Siciliano, L.; Cascio, A.; et al. Protective and causative killer Ig-like receptor (KIR) and metalloproteinase genetic patterns associated with Herpes simplex virus 1 (HSV-1) encephalitis occurrence. J. Neuroimmunol. 2020, 344, 577241. [Google Scholar] [CrossRef]
- Guerini, F.R.; Mancuso, R.; Agostini, S.; Agliardi, C.; Zanzottera, M.; Hernis, A.; Tourlaki, A.; Calvo, M.G.; Bellinvia, M.; Brambilla, L.; et al. Activating KIR/HLA complexes in classic Kaposi’s Sarcoma. Infect. Agent Cancer 2012, 7, 9. [Google Scholar] [CrossRef]
- Borghi, A.; D’Accolti, M.; Rizzo, R.; Virgili, A.; Di Luca, D.; Corazza, M.; Caselli, E. High prevalence of specific KIR types in patients with HHV-8 positive cutaneous vascular lesions: A possible predisposing factor? Arch. Dermatol. Res. 2016, 308, 373–377. [Google Scholar] [CrossRef]
- Bortolotti, D.; Corazza, M.; Rotola, A.; Bencivelli, D.; Schiuma, G.; Danese, E.; Rizzo, S.; Beltrami, S.; Rizzo, R.; Borghi, A. Inhibitory KIR2DL2 receptor and HHV-8 in classic or endemic Kaposi sarcoma. Clin. Exp. Med. 2023, 23, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Goedert, J.J.; Martin, M.P.; Vitale, F.; Lauria, C.; Whitby, D.; Qi, Y.; Gao, X.; Carrington, M. Risk of classic Kaposi sarcoma with combinations of killer immunoglobulin-like receptor and human leukocyte antigen loci: A population-based case-control study. J. Infect. Dis. 2016, 213, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Li, Y.; Han, X.; Xu, Y.; Wang, H.; Wang, T.; Zhang, T. Role of the killer immunoglobulin-like receptor and human leukocyte antigen I complex polymorphisms in Kaposi sarcoma-associated herpesvirus infection. Open Forum Infect. Dis. 2023, 10, ofad435. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.P.; Gao, X.; Lee, J.H.; Nelson, G.W.; Detels, R.; Goedert, J.J.; Buchbinder, S.; Hoots, K.; Vlahov, D.; Trowsdale, J.; et al. Epistatic interaction between KIR3DS1 and HLA-B delays the progression to AIDS. Nat. Genet. 2002, 31, 429–434. [Google Scholar] [CrossRef]
- Boulet, S.; Sharafi, S.; Simic, N.; Bruneau, J.; Routy, J.P.; Tsoukas, C.M.; Bernard, N.F. Increased proportion of KIR3DS1 homozygotes in HIV-exposed uninfected individuals. AIDS 2008, 22, 595–599. [Google Scholar] [CrossRef]
- Martin, M.P.; Qi, Y.; Gao, X.; Yamada, E.; Martin, J.N.; Pereyra, F.; Colombo, S.; Brown, E.E.; Shupert, W.L.; Phair, J.; et al. Innate partnership of HLA-B and KIR3DL1 subtypes against HIV-1. Nat. Genet. 2007, 39, 733–740. [Google Scholar] [CrossRef]
- Boulet, S.; Song, R.; Kamya, P.; Bruneau, J.; Shoukry, N.H.; Tsoukas, C.M.; Bernard, N.F. HIV protective KIR3DL1 and HLA-B genotypes influence NK cell function following stimulation with HLA-devoid cells. J. Immunol. 2010, 184, 2057–2064. [Google Scholar] [CrossRef]
- Maruthamuthu, S.; Rajalingam, R.; Pandian, K.; Madasamy, S.; Manoharan, M.; Pitchai, L.; Murugesan, A.; Mariakuttikan, J. Inhibitory natural killer cell receptor KIR3DL1 with its ligand Bw4 constraints HIV-1 disease among South Indians. AIDS 2018, 32, 2679–2688. [Google Scholar] [CrossRef]
- López-Vázquez, A.; Miña-Blanco, A.; Martínez-Borra, J.; Njobvu, P.D.; Suárez-Alvarez, B.; Blanco-Gelaz, M.A.; González, S.; Rodrigo, L.; López-Larrea, C. Interaction between KIR3DL1 and HLA-B*57 supertype alleles influences the progression of HIV-1 infection in a Zambian population. Hum. Immunol. 2005, 66, 285–289. [Google Scholar] [CrossRef]
- Boulet, S.; Kleyman, M.; Kim, J.Y.; Kamya, P.; Sharafi, S.; Simic, N.; Bruneau, J.; Routy, J.P.; Tsoukas, C.M.; Bernard, N.F. A combined genotype of KIR3DL1 high expressing alleles and HLA-B*57 is associated with a reduced risk of HIV infection. AIDS 2008, 22, 1487–1491. [Google Scholar] [CrossRef] [PubMed]
- Pelak, K.; Need, A.C.; Fellay, J.; Shianna, K.V.; Feng, S.; Urban, T.J.; Ge, D.; De Luca, A.; Martinez-Picado, J.; Wolinsky, S.M.; et al. Copy number variation of KIR genes influences HIV-1 control. PLoS Biol. 2011, 9, e1001208. [Google Scholar] [CrossRef]
- Alter, G.; Altfeld, M. NK cells in HIV-1 infection: Evidence for their role in the control of HIV-1 infection. J. Intern. Med. 2009, 265, 29–42. [Google Scholar] [CrossRef] [PubMed]
- Gaudieri, S.; DeSantis, D.; McKinnon, E.; Moore, C.; Nolan, D.; Witt, C.S.; Mallal, S.A.; Christiansen, F.T. Killer immunoglobulin-like receptors and HLA act both independently and synergistically to modify HIV disease progression. Genes Immun. 2005, 6, 683–690. [Google Scholar] [CrossRef]
- Jennes, W.; Verheyden, S.; Demanet, C.; Adjé-Touré, C.A.; Vuylsteke, B.; Nkengasong, J.N.; Kestens, L. Cutting edge: Resistance to HIV-1 infection among African female sex workers is associated with inhibitory KIR in the absence of their HLA ligands. J. Immunol. 2006, 177, 6588–6592. [Google Scholar] [CrossRef]
- Jennes, W.; Verheyden, S.; Demanet, C.; Menten, J.; Vuylsteke, B.; Nkengasong, J.N.; Kestens, L. Low CD4+ T cell counts among African HIV-1 infected subjects with group B KIR haplotypes in the absence of specific inhibitory KIR ligands. PLoS ONE 2011, 6, e17043. [Google Scholar] [CrossRef]
- Jennes, W.; Verheyden, S.; Mertens, J.W.; Camara, M.; Seydi, M.; Dieye, T.N.; Mboup, S.; Demanet, C.; Kestens, L. Inhibitory KIR/HLA incompatibility between sexual partners confers protection against HIV-1 transmission. Blood 2013, 121, 1157–1164. [Google Scholar] [CrossRef]
- Zwolińska, K.; Błachowicz, O.; Tomczyk, T.; Knysz, B.; Gąsiorowski, J.; Zalewska, M.; Orzechowska, B.U.; Sochocka, M.; Piasecki, E. The effects of killer cell immunoglobulin-like receptor (KIR) genes on susceptibility to HIV-1 infection in the Polish population. Immunogenetics 2016, 68, 327–337. [Google Scholar] [CrossRef]
- Zhao, J.; Tang, W.; Yao, J.; Chen, Q.; Xu, Q.; Wu, S. The role of killer immunoglobulin-like receptor genes in susceptibility to HIV-1 infection and disease progression: A meta-analysis. AIDS Res. Hum. Retroviruses 2019, 35, 948–959. [Google Scholar] [CrossRef]
- Sorgho, P.A.; Djigma, F.W.; Martinson, J.J.; Yonli, A.T.; Nagalo, B.M.; Compaore, T.R.; Diarra, B.; Sombie, H.K.; Simpore, A.; Zongo, A.W.; et al. Role of Killer cell immunoglobulin-like receptors (KIR) genes in stages of HIV-1 infection among patients from Burkina Faso. Biomol. Concepts 2019, 10, 226–236. [Google Scholar] [CrossRef]
- Paximadis, M.; Minevich, G.; Winchester, R.; Schramm, D.B.; Gray, G.E.; Sherman, G.G.; Coovadia, A.H.; Kuhn, L.; Tiemessen, C.T. KIR-HLA and maternal-infant HIV-1 transmission in sub-Saharan Africa. PLoS ONE 2011, 6, e16541. [Google Scholar] [CrossRef] [PubMed]
- Luc-Aimé, K.S.; Louis-Marie, Y.; Gabriel, L.E.; Yengo, C.K.; Esemu Livo, F.; Assob, N.J.C. Killer-cell immunoglobulin-like receptors (KIR) in HIV-exposed infants in Cameroon. J. Immunol. Res. 2021, 2021, 9053280. [Google Scholar] [CrossRef]
- Soria, A.; Guerini, F.R.; Bandera, A.; Bolognesi, E.; Uglietti, A.; Fusco, C.; Zucchi, P.; Maserati, R.; Rizzardini, G.; Clerici, M.; et al. KIR-HLA genotypes in HIV-infected patients lacking immunological recovery despite effective antiretroviral therapy. PLoS ONE 2011, 6, e27349. [Google Scholar] [CrossRef] [PubMed]
- Bonagura, V.R.; Du, Z.; Ashouri, E.; Luo, L.; Hatam, L.J.; DeVoti, J.A.; Rosenthal, D.W.; Steinberg, B.M.; Abramson, A.L.; Gjertson, D.W.; et al. Activating killer cell immunoglobulin-like receptors 3DS1 and 2DS1 protect against developing the severe form of recurrent respiratory papillomatosis. Hum. Immunol. 2010, 71, 212–219. [Google Scholar] [CrossRef]
- Song, M.J.; Lee, C.W.; Kim, J.H.; Lee, S.J.; Kim, C.J.; Hur, S.Y.; Park, T.C.; Kim, T.G.; Park, J.S. Association of KIR genes and HLA-C alleles with HPV-related uterine cervical disease in Korean women. Tissue Antigens 2013, 81, 164–170. [Google Scholar] [CrossRef]
- Marangon, A.V.; Guelsin, G.A.; Visentainer, J.E.; Borelli, S.D.; Watanabe, M.A.; Consolaro, M.E.; Caleffi-Ferracioli, K.R.; Rudnick, C.C.; Sell, A.M. The association of the immune response genes to human papillomavirus-related cervical disease in a Brazilian population. BioMed Res. Int. 2013, 2013, 146079. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, R.; Gentili, V.; Rotola, A.; Bortolotti, D.; Cassai, E.; Di Luca, D. Implication of HLA-C and KIR alleles in human papillomavirus infection and associated cervical lesions. Viral Immunol. 2014, 27, 468–470. [Google Scholar] [CrossRef]
- Martin, M.P.; Borecki, I.B.; Zhang, Z.; Nguyen, L.; Ma, D.; Gao, X.; Qi, Y.; Carrington, M.; Rader, J.S. HLA-Cw group 1 ligands for KIR increase susceptibility to invasive cervical cancer. Immunogenetics 2013, 6, 761–765. [Google Scholar] [CrossRef]
- Ivancic, R.; Iqbal, H.; deSilva, B.; Pan, Q.; Matrka, L. Immunological tolerance of low-risk HPV in recurrent respiratory papillomatosis. Clin. Exp. Immunol. 2020, 199, 131–142. [Google Scholar] [CrossRef]
- Paaso, A.; Jaakola, A.; Syrjänen, S.; Louvanto, K. From HPV Infection to Lesion Progression: The Role of HLA Alleles and Host Immunity. Acta Cytol. 2019, 63, 148–158. [Google Scholar] [CrossRef]
- Tarique, M.; Naz, H.; Suhail, M.; Turan, A.; Saini, C.; Muhammad, N.; Shankar, H.; Zughaibi, T.A.; Khan, T.H.; Khanna, N.; et al. Differential expression of programmed death 1 (PD-1) on various immune cells and its role in human leprosy. Front. Immunol. 2023, 14, 1138145. [Google Scholar] [CrossRef]
- Franceschi, D.S.; Mazini, P.S.; Rudnick, C.C.; Sell, A.M.; Tsuneto, L.T.; de Melo, F.C.; Braga, M.A.; Peixoto, P.R.; Visentainer, J.E. Association between killer-cell immunoglobulin-like receptor genotypes and leprosy in Brazil. Tissue Antigens 2008, 72, 478–482. [Google Scholar] [CrossRef] [PubMed]
- Jarduli, L.R.; Alves, H.V.; de Souza-Santana, F.C.; Marcos, E.V.; Pereira, A.C.; Dias-Baptista, I.M.; Fava, V.M.; Mira, M.T.; Moraes, M.O.; Virmond, M.C.; et al. Influence of KIR genes and their HLA ligands in the pathogenesis of leprosy in a hyperendemic population of Rondonópolis, Southern Brazil. BMC Infect. Dis. 2014, 14, 438. [Google Scholar] [CrossRef]
- Alves, H.V.; de Moraes, A.G.; Pepineli, A.C.; Tiyo, B.T.; de Lima Neto, Q.A.; Santos, T.D.S.; Teixeira, J.J.V.; Ambrosio-Albuquerque, E.P.; Sell, A.M.; Visentainer, J.E.L. The impact of KIR/HLA genes on the risk of developing multibacillary leprosy. PLoS Neglected Trop. Dis. 2019, 13, e0007696. [Google Scholar] [CrossRef]
- Martins, L.; Couto, A.R.; Bettencourt, B.F.; Santos, M. Genetic polymorphisms of killer cell immunoglobulin-like receptors are associated with susceptibility to leptospirosis—A preliminary study. Tissue Antigens 2006, 67, 9600–9606. [Google Scholar]
- Fialho, R.N.; Martins, L.; Pinheiro, J.P.; Bettencourt, B.F.; Couto, A.R.; Santos, M.R.; Peixoto, M.J.; Garrett, F.; Leal, J.; Tomás, A.M.; et al. Role of human leukocyte antigen, killer-cell immunoglobulin-like receptors, and cytokine gene polymorphisms in leptospirosis. Hum. Immunol. 2009, 70, 915–920. [Google Scholar] [CrossRef]
- Taniguchi, M.; Kawabata, M. KIR3DL1/S1 genotypes and KIR2DS4 allelic variants in the AB KIR genotypes are associated with Plasmodium-positive individuals in malaria infection. Immunogenetics 2009, 61, 717–730. [Google Scholar] [CrossRef]
- Lourembam, S.D.; Sawian, C.E.; Baruah, S. Differential association of KIR gene loci to risk of malaria in ethnic groups of Assam, Northeast India. Infect. Genet. Evol. 2011, 11, 1921–1928. [Google Scholar] [CrossRef]
- Hirayasu, K.; Ohashi, J.; Kashiwase, K.; Hananantachai, H.; Naka, I.; Ogawa, A.; Takanashi, M.; Satake, M.; Nakajima, K.; Parham, P.; et al. Significant association of KIR2DL3-HLA-C1 combination with cerebral malaria and implications for co-evolution of KIR and HLA. PLoS Pathog. 2012, 8, e1002565. [Google Scholar] [CrossRef]
- Yindom, L.M.; Forbes, R.; Aka, P.; Janha, O.; Jeffries, D.; Jallow, M.; Conway, D.J.; Walther, M. Killer-cell immunoglobulin-like receptors and malaria caused by Plasmodium falciparum in The Gambia. Tissue Antigens 2012, 79, 104–113. [Google Scholar] [CrossRef]
- Olaniyan, S.A.; Amodu, O.K.; Yindom, L.M.; Conway, D.J.; Aka, P.; Bakare, A.A.; Omotade, O.O. Killer-cell immunoglobulin-like receptors and falciparum malaria in southwest Nigeria. Hum. Immunol. 2014, 75, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Prakash, S.; Ranjan, P.; Ghoshal, U.; Agrawal, S. KIR-like activating natural killer cell receptors and their association with complicated malaria in north India. Acta Trop. 2018, 178, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Digitale, J.C.; Callaway, P.C.; Martin, M.; Nelson, G.; Viard, M.; Rek, J.; Arinaitwe, E.; Dorsey, G.; Kamya, M.; Carrington, M.; et al. Association of inhibitory killer cell immunoglobulin-like receptor ligands with higher Plasmodium falciparum parasite prevalence. J. Infect. Dis. 2021, 224, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Perce-da-Silva, D.S.; Silva, L.A.; Lima-Junior, J.C.; Cardoso-Oliveira, J.; Ribeiro-Alves, M.; Santos, F.; Porto, L.C.; Oliveira-Ferreira, J.; Banic, D.M. Killer cell immunoglobulin-like receptor (KIR) gene diversity in a population naturally exposed to malaria in Porto Velho, Northern Brazil. Tissue Antigens 2015, 85, 190–199. [Google Scholar] [CrossRef]
- Tukwasibwe, S.; Traherne, J.A.; Chazara, O.; Jayaraman, J.; Trowsdale, J.; Moffett, A.; Jiang, W.; Nankabirwa, J.I.; Rek, J.; Arinaitwe, E.; et al. Diversity of KIR genes and their HLA-C ligands in Ugandan populations with historically varied malaria transmission intensity. Malar. J. 2021, 20, 111. [Google Scholar] [CrossRef]
- España, P.P.; Uranga, A.; Cillóniz, C.; Torres, A. Q Fever (Coxiella burnetii). Semin. Respir. Crit. Care Med. 2020, 41, 509–521. [Google Scholar] [CrossRef]
- Helbig, K.J.; Heatley, S.L.; Harris, R.J.; Mullighan, C.G.; Bardy, P.G.; Marmion, B.P. Variation in immune response genes and chronic Q fever. Concepts: Preliminary test with post-Q fever fatigue syndrome. Genes Immun. 2003, 4, 82–85. [Google Scholar] [CrossRef]
- Helbig, K.; Harris, R.; Ayres, J.; Dunckley, H.; Lloyd, A.; Robson, J.; Marmion, B.P. Immune response genes in the post-Q-fever fatigue syndrome, Q fever endocarditis and uncomplicated acute primary Q fever. QJM Int. J. Med. 2005, 98, 565–574. [Google Scholar] [CrossRef]
- Oliveira, L.M.; Portela, P.; Merzoni, J.; Lindenau, J.D.; Dias, F.S.; Beppler, J.; Graebin, P.; Alho, C.S.; Schwartsmann, G.; Dal-Pizzol, F.; et al. Reduced frequency of two activating KIR genes in patients with sepsis. Hum. Immunol. 2017, 78, 363–369. [Google Scholar] [CrossRef]
- Oliveira, L.M.; Portela, P.; Merzoni, J.; Beppler, J.; Dias, F.S.; Graebin, P.; Alho, C.S.; Schwartsmann, G.; Dal-Pizzol, F.; Jobim, L.F.; et al. KIR gene haplotype A is associated with hospital mortality in patients with sepsis. Clin. Immunol. 2019, 200, 37–38. [Google Scholar] [CrossRef]
- Zheng, M.; Gao, Y.; Wang, G.; Song, G.; Liu, S.; Sun, D.; Xu, Y.; Tian, Z. Functional exhaustion of antiviral lymphocytes in COVID-19 patients. Cell. Mol. Immunol. 2020, 17, 533–535. [Google Scholar] [CrossRef] [PubMed]
- Littera, R.; Chessa, L.; Deidda, S.; Angioni, G.; Campagna, M.; Lai, S.; Melis, M.; Cipri, S.; Firinu, D.; Santus, S.; et al. Natural killer-cell immunoglobulin-like receptors trigger differences in immune response to SARS-CoV-2 infection. PLoS ONE 2021, 16, e0255608. [Google Scholar] [CrossRef]
- Teshnizi, S.H.; Mirzazadeh, S.; Mashhadi, N.; Meri, S.; Kabelitz, D.; Kalantar, K. Association study between killer immunoglobulin-like receptor polymorphisms and susceptibility to COVID-19 disease: A systematic review and meta-analysis. Immunol. Res. 2024, 72, 175–184. [Google Scholar] [CrossRef]
- Saresella, M.; Trabattoni, D.; Marventano, I.; Piancone, F.; La Rosa, F.; Caronni, A.; Lax, A.; Bianchi, L.; Banfi, P.; Navarro, J.; et al. NK Cell Subpopulations and Receptor Expression in Recovering SARS-CoV-2 Infection. Mol. Neurobiol. 2021, 58, 6111–6120. [Google Scholar] [CrossRef]
- Hajeer, A.; Jawdat, D.; Massadeh, S.; Aljawini, N.; Abedalthagafi, M.S.; Arabi, Y.M.; Alaamery, M. Association of KIR gene polymorphisms with COVID-19 disease. Clin. Immunol. 2022, 234, 108911. [Google Scholar] [CrossRef] [PubMed]
- Balas, A.; Moreno-Hidalgo, M.Á.; de la Calle-Prieto, F.; Vicario, J.L.; Arsuaga, M.; Trigo, E.; de Miguel-Buckley, R.; Bellón, T.; Díaz-Menéndez, M. Coronavirus-19 disease risk and protective factors associated with HLA/KIR polymorphisms in Ecuadorian patients residing in Madrid. Hum. Immunol. 2023, 84, 571–577. [Google Scholar] [CrossRef]
- Farias, T.D.J.; Brugiapaglia, S.; Croci, S.; Magistroni, P.; Curcio, C.; Zguro, K.; Fallerini, C.; Fava, F.; Pettini, F.; Kichula, K.M.; et al. HLA-DPB113:01 associates with enhanced, and KIR2DS4001 with diminished protection from developing severe COVID-19. HLA 2024, 103, e15251. [Google Scholar] [CrossRef]
- Maruthamuthu, S.; Rajalingam, K.; Kaur, N.; Morvan, M.G.; Soto, J.; Lee, N.; Kong, D.; Hu, Z.; Reyes, K.; Ng, D.; et al. Individualized Constellation of Killer Cell Immunoglobulin-Like Receptors and Cognate HLA Class I Ligands that Controls Natural Killer Cell Antiviral Immunity Predisposes COVID-19. Front. Genet. 2022, 13, 845474. [Google Scholar] [CrossRef]
- Hu, S.; Shao, Z.; Ni, W.; Sun, P.; Qiao, J.; Wan, H.; Huang, Y.; Liu, X.; Zhai, H.; Xiao, M.; et al. The KIR2DL2/HLA-C1C1 Gene Pairing Is Associated With an Increased Risk of SARS-CoV-2 Infection. Front. Immunol. 2022, 13, 919110. [Google Scholar] [CrossRef]
- Alomar, S.; Alkhuriji, A.; Alkhulaifi, F.M.; Mansour, L.; Al-Jurayyan, A.; Aldossari, G.S.; Albalawi, A.E.; Alanazi, A.D. Relationship between KIR genotypes and HLA-ligands with SARS-CoV-2 infection in the Saudi population. J. King Saud Univ. Sci. 2023, 35, 102416. [Google Scholar] [CrossRef]
- Zhuang, Y.L.; Song, Y.; Zhu, C.; Zhang, Y.; Wang, D.; Nie, X.; Liu, Y.; Ren, G.J. Association of KIR genotypes and haplotypes with syphilis in a Chinese Han population. Scand. J. Immunol. 2012, 75, 361–367. [Google Scholar] [CrossRef]
- Zhuang, Y.L.; Zhu, C.F.; Zhang, Y.; Song, Y.H.; Wang, D.J.; Nie, X.M.; Liu, Y.; Ren, G.J. Association of KIR2DS4 and its variant KIR1D with syphilis in a Chinese Han population. Int. J. Immunogenet. 2012, 39, 114–118. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Y.L.; Ren, G.J.; Tian, K.L.; Li, X.Y.; Zhu, Y.B.; Liu, J.L.; Si, G.L.; Li, P.; Zhang, Y.; Wang, L.; et al. Human leukocyte antigen-C and killer cell immunoglobulin-like receptor gene polymorphisms among patients with syphilis in a Chinese Han population. APMIS 2012, 120, 828–835. [Google Scholar] [CrossRef]
- Braun, K.; Wolfe, J.; Kiazyk, S.; Kaushal Sharma, M. Evaluation of host genetics on outcome of tuberculosis infection due to differences in killer immunoglobulin-like receptor gene frequencies and haplotypes. BMC Genet. 2015, 16, 63. [Google Scholar] [CrossRef]
- Omraninava, M.; Mehranfar, S.; Khosrojerdi, A.; Jamalzehi, S.; Karami, J.; Motallebnezhad, M.; Javan, M.R.; Aslani, S.; Mohammadi, H.; Kousha, A.; et al. Systematic review and meta-analytic findings on the association between killer-cell immunoglobulin-like receptor genes and susceptibility to pulmonary tuberculosis. Pathog. Glob. Health 2021, 115, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Esmaeili, S.; Mahmoudi, A.; Esmaeili, P.; Yousefi Ghalejoogh, Z.; Mordadi, A.; Ghasemi, A.; Mohammadi, A.; Bagheri, A.; Sohrabi, A.; Latifian, M.; et al. The surveillance of plague among rodents and dogs in Western Iran. PLoS Neglected Trop. Dis. 2023, 17, e0011722. [Google Scholar] [CrossRef]
- Immel, A.; Key, F.M.; Szolek, A.; Barquera, R.; Robinson, M.K.; Harrison, G.F.; Palmer, W.H.; Spyrou, M.A.; Susat, J.; Krause-Kyora, B.; et al. Analysis of genomic DNA from medieval plague victims suggests long-term effect of Yersinia pestis on human immunity genes. Mol. Biol. Evol. 2021, 38, 4059–4076. [Google Scholar] [CrossRef]
- Di, D.; Thomas, J.S.; Currat, M.; Nunes, J.M.; Sanchez-Mazas, A. Challenging ancient DNA results about putative HLA protection or susceptibility to Yersinia pestis. Mol. Biol. Evol. 2022, 39, msac073. [Google Scholar] [CrossRef]
- Spiroski, M.; Milenkovic, Z.; Petlichkovski, A.; Ivanovski, L.; Topuzovska, I.K.; Djulejic, E. Killer cell immunoglobulin-like receptor genes in four human West Nile virus infections reported 2011 in the Republic of Macedonia. Hum. Immunol. 2013, 74, 389–394. [Google Scholar] [CrossRef]
- Elpidio, L.N.S.; de Moraes, A.G.; Langer, I.B.V.; do Amaral, G.C.; Moretti, M.L.; Garcia, M.T.; Angerami, R.; Proenca-Modena, J.L.; Bispo-Dos-Santos, K.; Martini, M.C.; et al. Lack of association of the KIR and HLA class I ligands with ZIKV infection in south and southeast of Brazil. Mem. Inst. Oswaldo Cruz. 2022, 117, e210194. [Google Scholar] [CrossRef]
- Torres, A.R.; Westover, J.B.; Gibbons, C.; Johnson, R.C.; Ward, D.C. Activating killer-cell immunoglobulin-like receptors (KIR) and their cognate HLA ligands are significantly increased in autism. Brain Behav. Immun. 2012, 26, 1122–1127. [Google Scholar] [CrossRef] [PubMed]
- Torres, A.R.; Sweeten, T.L.; Johnson, R.C.; Odell, D.; Westover, J.B.; Bray-Ward, P.; Ward, D.C.; Davies, C.J.; Thomas, A.J.; Croen, L.A.; et al. Common genetic variants found in HLA and KIR immune genes in autism spectrum disorder. Front. Neurosci. 2016, 10, 463. [Google Scholar] [CrossRef]
- Guerini, F.R.; Bolognesi, E.; Chiappedi, M.; Manca, S.; Ghezzo, A.; Agliardi, C.; Zanette, M.; Littera, R.; Carcassi, C.; Sotgiu, S.; et al. Activating KIR molecules and their cognate ligands prevail in children with a diagnosis of ASD and in their mothers. Brain Behav. Immun. 2014, 36, 54–60. [Google Scholar] [CrossRef]
- Anderson, K.M.; Augusto, D.G.; Dandekar, R.; Shams, H.; Zhao, C.; Yusufali, T.; Montero-Martín, G.; Marin, W.M.; Nemat-Gorgani, N.; Creary, L.E.; et al. Killer Cell Immunoglobulin-like Receptor Variants Are Associated with Protection from Symptoms Associated with More Severe Course in Parkinson Disease. J. Immunol. 2020, 205, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- Yovel, G.; Sirota, P.; Mazeh, D.; Shakhar, G.; Rosenne, E.; Ben-Eliyahu, S. Higher natural killer cell activity in schizophrenic patients: The impact of serum factors, medication, and smoking. Brain Behav. Immun. 2000, 14, 153–169. [Google Scholar] [CrossRef]
- Wiśniewski, A.; Frydecka, D.; Nowak, I.; Majorczyk, E.; Senitzer, D.; Piotrowski, P.; Beszlej, A.; Kiejna, A.; Kuśnierczyk, P. Are KIR and HLA class I genes associated with schizophrenia? Tissue Antigens 2014, 84, 503–504. [Google Scholar] [CrossRef]
- Alsbrook, D.L.; Di Napoli, M.; Bhatia, K.; Biller, J.; Andalib, S.; Hinduja, A.; Rodrigues, R.; Rodriguez, M.; Sabbagh, S.Y.; Selim, M.; et al. Neuroinflammation in Acute Ischemic and Hemorrhagic Stroke. Curr. Neurol. Neurosci. Rep. 2023, 23, 407–431. [Google Scholar] [CrossRef]
- Tuttolondo, A.; Di Raimondo, D.; Pecoraro, R.; Casuccio, A.; Di Bona, D.; Aiello, A.; Accardi, G.; Arnao, V.; Clemente, G.; Corte, V.D.; et al. HLA and killer cell immunoglobulin-like receptor (KIRs) genotyping in patients with acute ischemic stroke. J. Neuroinflammation 2019, 16, 88. [Google Scholar] [CrossRef]
- Brézin, A.P.; Monnet, D.; Cohen, J.H.; Levinson, R.D. HLA-A29 and birdshot chorioretinopathy. Ocul. Immunol. Inflamm. 2011, 19, 397–400. [Google Scholar] [CrossRef]
- Levinson, R.D.; Du, Z.; Luo, L.; Monnet, D.; Tabary, T.; Brezin, A.P.; Zhao, L.; Gjertson, D.W.; Holland, G.N.; Reed, E.F.; et al. Combination of KIR and HLA gene variants augments the risk of developing birdshot chorioretinopathy in HLA-A*29-positive individuals. Genes Immun. 2008, 9, 249–258. [Google Scholar] [CrossRef]
- Abramiuk, M.; Grywalska, E.; Małkowska, P.; Sierawska, O.; Hrynkiewicz, R.; Niedźwiedzka-Rystwej, P. The Role of the Immune System in the Development of Endometriosis. Cells 2022, 11, 2028. [Google Scholar] [CrossRef]
- Kitawaki, J.; Xu, B.; Ishihara, H.; Fukui, M.; Hasegawa, G.; Nakamura, N.; Mizuno, S.; Ohta, M.; Obayashi, H.; Honjo, H.; et al. Association of killer cell immunoglobulin-like receptor genotypes with susceptibility to endometriosis. Am. J. Reprod. Immunol. 2007, 58, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Nowak, I.; Majorczyk, E.; Wiśniewski, A.; Pawlik, A.; Magott-Procelewska, M.; Passowicz-Muszyńska, E.; Malejczyk, J.; Płoski, R.; Giebel, S.; Barcz, E.; et al. Does the KIR2DS5 gene protect from some human diseases? PLoS ONE 2010, 5, e12381. [Google Scholar] [CrossRef]
- Chou, Y.C.; Chen, C.H.; Chen, M.J.; Chang, C.W.; Chen, P.H.; Yu, M.H.; Chen, Y.J.; Tsai, E.M.; Yang, P.S.; Lin, S.Y.; et al. Killer cell immunoglobulin-like receptors (KIR) and human leukocyte antigen-C (HLA-C) allorecognition patterns in women with endometriosis. Sci. Rep. 2020, 10, 4897. [Google Scholar] [CrossRef]
- Tufan, A.; Lachmann, H.J. Familial Mediterranean fever, from pathogenesis to treatment: A contemporary review. Turk. J. Med. Sci. 2020, 50, 1591–1610. [Google Scholar] [CrossRef] [PubMed]
- Mahfouz, R.A.; Sabbagh, A.S.; Shamseddine, W.; Bazarbachi, A.; Ibrahim, G.; Arayssi, T.; El Hajj, N.; Rayes, R.; Zaatari, G.S.; Khazen, G.; et al. Distribution of killer cell immunoglobulin-like receptor (KIR) genotypes in patients with familial Mediterranean fever. Genet. Test. Mol. Biomark. 2009, 13, 91–95. [Google Scholar] [CrossRef]
- Erken, E.; Goruroglu Ozturk, O.; Kudas, O.; Arslan Tas, D.; Demirtas, A.; Kibar, F.; Dinkci, S.; Erken, E. Killer Cell Immunoglobulin-Like Receptor (KIR) Genotype Distribution in Familial Mediterranean Fever (FMF) Patients. Med. Sci. Monit. 2015, 21, 3547–3554. [Google Scholar] [CrossRef]
- Stone, W.L.; Basit, H.; Mukkamalla, S.K.R.; Master, S.R. Gaucher Disease. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Vairo, F.; Portela, P.; Salim, P.H.; Jobim, M.; Netto, C.; Dorneles, A.; Mittlestadt, S.; Jobim, L.F.; Schwartz, I.V. Human leukocyte antigens and Gaucher disease. Blood Cells Mol. Dis. 2013, 50, 202–205. [Google Scholar] [CrossRef]
- Burstein, Y.; Zakuth, V.; Rechavi, G.; Spirer, Z. Abnormalities of cellular immunity and natural killer cells in Gaucher’s disease. J. Clin. Lab. Immunol. 1987, 23, 149–151. [Google Scholar]
- Vairo, F.; Portela, P.; Salim, P.H.; Jobim, M.; Netto, C.; Dorneles, A.; Mittlestadt, S.; Jobim, L.F.; Schwartz, I.V. KIR genes and HLA class I ligands in Gaucher disease. Gene 2013, 516, 53–57. [Google Scholar] [CrossRef]
- Landgren, O.; Turesson, I.; Gridley, G.; Caporaso, N.E. Risk of malignant disease among 1525 adult male US veterans with Gaucher disease. Arch. Intern. Med. 2007, 167, 1189–1194. [Google Scholar] [CrossRef]
- Hill, A.; DeZern, A.E.; Kinoshita, T.; Brodsky, R.A. Paroxysmal nocturnal haemoglobinuria. Nat. Rev. Dis. Primers 2017, 3, 17028. [Google Scholar] [CrossRef] [PubMed]
- Cosentini, E.; Gargiulo, L.; Bruno, P.; Lastraioli, S.; Risitano, A.; Camerlingo, R.; Luongo, V.; Serra, M.; Sica, M.; Garzillo, C.; et al. Killer immunoglobulin-like receptors (KIR) and their HLA-ligands in Italian paroxysmal nocturnal haemoglobinuria (PNH) patients. Tissue Antigens 2012, 80, 322–327. [Google Scholar] [CrossRef]
- Meier, R.K. Polycystic Ovary Syndrome. Nurs. Clin. N. Am. 2018, 53, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Sala, E.L.N.; de Alencar, J.B.; Tsuneto, P.Y.; Alves, H.V.; Trento Toretta, M.; It Taura, S.K.; Laguila Visentainer, J.E.; Sell, A.M. Killer-cell immunoglobulin-like receptors associated with polycystic ovary syndrome. J. Reprod. Immunol. 2018, 130, 1–6. [Google Scholar] [CrossRef]
- Dimitriadis, E.; Rolnik, D.L.; Zhou, W.; Estrada-Gutierrez, G.; Koga, K.; Francisco, R.P.V.; Whitehead, C.; Hyett, J.; da Silva Costa, F.; Nicolaides, K.; et al. Pre-eclampsia. Nat. Rev. Dis. Primers 2023, 9, 8, Erratum in Nat. Rev. Dis. Primers 2023, 9, 35. https://doi.org/10.1038/s41572-023-00451-4. [Google Scholar] [CrossRef]
- Hiby, S.; Walker, J.J.; O’Shaughnessy, K.M.; Redman, C.W.; Carrington, M.; Trowsdale, J.; Moffett, A. Combinations of maternal KIR and fetal HLA-C genes influence the risk of preeclampsia and reproductive success. J. Exp. Med. 2004, 200, 957–965. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhao, Y.H.; Zhang, W.Y.; Cui, M.H. Correlation between killer cell immunoglobulin-like receptors genes and pre-eclampsia. Zhonghua Fu Chan Ke Za Zhi 2008, 43, 909–912. [Google Scholar] [PubMed]
- Wang, D.; Tian, Y.; Zhao, Y.; Liu, L.; Liu, X.; Wu, F. KIR2DL4 expression rather than its single nucleotide polymorphisms correlates with pre-eclampsia. Int. J. Clin. Exp. Pathol. 2015, 8, 14535–14541. [Google Scholar]
- Sánchez-Rodríguez, E.N.; Nava-Salazar, S.; Mendoza-Rodríguez, C.A.; Moran, C.; Romero-Arauz, J.F.; Ortega, E.; Granados, J.; Cervantes-Peredo, A.; Cerbón, M. Persistence of decidual NK cells and KIR genotypes in healthy pregnant and preeclamptic women: A case-control study in the third trimester of gestation. Reprod. Biol. Endocrinol. 2011, 9, 8. [Google Scholar] [CrossRef]
- Yusrizal, F. Genetic and preeclampsia. Pregnancy Hypertens. 2014, 4, 243. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Pan, N.; Shen, Y.; Jin, S.; Zhai, J.; Qiao, D.; Shen, Y.; Miao, F.; Wang, L.; He, Y.; et al. Interaction of parental KIR and fetal HLA-C genotypes with the risk of preeclampsia. Hypertens. Pregnancy 2014, 33, 402–411. [Google Scholar] [CrossRef] [PubMed]
- Nakimuli, A.; Chazara, O.; Hiby, S.E.; Farrell, L.; Tukwasibwe, S.; Jayaraman, J.; Traherne, J.A.; Trowsdale, J.; Colucci, F.; Lougee, E.; et al. A KIR B centromeric region present in Africans but not Europeans protects pregnant women from pre-eclampsia. Proc. Natl. Acad. Sci. USA 2015, 112, 845–850. [Google Scholar] [CrossRef]
- Long, W.; Shi, Z.; Fan, S.; Liu, L.; Lu, Y.; Guo, X.; Rong, C.; Cui, X.; Ding, H. Association of maternal KIR and fetal HLA-C genes with the risk of preeclampsia in the Chinese Han population. Placenta 2015, 36, 433–437. [Google Scholar] [CrossRef] [PubMed]
- Huhn, O.; Chazara, O.; Ivarsson, M.A.; Retière, C.; Venkatesan, T.C.; Norman, P.J.; Hilton, H.G.; Jayaraman, J.; Traherne, J.A.; Trowsdale, J.; et al. High-resolution genetic and phenotypic analysis of KIR2DL1 alleles and their association with pre-eclampsia. J. Immunol. 2018, 201, 2593–2601. [Google Scholar] [CrossRef]
- Ishmail, H.; Khaliq, O.P.; Ngene, N.C. The role of genetics in maternal susceptibility to preeclampsia in women of African ancestry. J. Reprod. Immunol. 2023, 160, 104139. [Google Scholar] [CrossRef]
- Somashekar, A.; Leung, Y.T. Updates in the diagnosis and management of Takayasu’s arteritis. Postgrad. Med. 2023, 135, 14–21. [Google Scholar] [CrossRef]
- Beigmohammadi, F.; Aslani, S.; Kavosi, H.; Javinani, A.; Mostafaei, S.; Pournazari, M.; Tasorian, B.; Farhadi, E.; Hajiabbasi, A.; Zayeni, H.; et al. Association of Killer Cell Immunoglobulin-like Receptor (KIR) Genes and their HLA Ligands with Susceptibility to Takayasu Arteritis in the Iranian Population. Iran J. Allergy Asthma Immunol. 2023, 22, 25–33. [Google Scholar] [CrossRef]
- Black, J.O. Xeroderma Pigmentosum. Head Neck Pathol. 2016, 10, 139–144. [Google Scholar] [CrossRef]
- Giraldo, G.; Degos, L.; Beth, E.; Gharbi, R.M.; Day, N.K.; Dastot, H.; Haus, M.; Reboul, M.; Schmid, M. HLA antigens in 16 families with xeroderma pigmentosum. Tissue Antigens 1977, 9, 167–170. [Google Scholar] [CrossRef]
- Freund-Brown, J.; Chirino, L.; Kambayashi, T. Strategies to Enhance NK Cell Function for the Treatment of Tumors and Infections. Crit. Rev. Immunol. 2018, 38, 105–130. [Google Scholar] [CrossRef] [PubMed]
- Vey, N.; Karlin, L.; Sadot-Lebouvier, S.; Broussais, F.; Berton-Rigaud, D.; Rey, J.; Charbonnier, A.; Marie, D.; André, P.; Paturel, C.; et al. A Phase 1 Study of Lirilumab (Antibody Against Killer Immunoglobulin-Like Receptor Antibody KIR2D.; IPH2102) in Patients with Solid Tumors and Hematologic Malignancies. Oncotarget 2018, 9, 17675–17688. [Google Scholar] [CrossRef]
- Kucuksezer, U.C.; Aktas Cetin, E.; Esen, F.; Tahrali, I.; Akdeniz, N.; Gelmez, M.Y.; Deniz, G. The Role of Natural Killer Cells in Autoimmune Diseases. Front. Immunol. 2021, 12, 622306. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Basar, R.; Wang, G.; Liu, E.; Moyes, J.S.; Li, L.; Kerbauy, L.N.; Uprety, N.; Fathi, M.; Rezvan, A.; et al. KIR-Based Inhibitory CARs Overcome CAR-NK Cell Trogocytosis-Mediated Fratricide and Tumor Escape. Nat. Med. 2022, 28, 2133–2144. [Google Scholar] [CrossRef]
- Khan, T.H.; Muhammad, N.; Tarique, M.; Usmani, D.; Naz, H.; Sarode, A. The Role of Cancer-Specific Target Antigens in CAR T Cell Therapy in Hematological Malignancies. Curr. Tissue Microenviron. Rep. 2024, 5, 61–67. [Google Scholar] [CrossRef]
- Khawar, M.B.; Sun, H. CAR-NK Cells: From Natural Basis to Design for Kill. Front. Immunol. 2021, 12, 707542. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Sferruzza, G.; Yang, L.; Zhou, L.; Chen, S. CAR-T and CAR-NK as Cellular Cancer Immunotherapy for Solid Tumors. Cell Mol. Immunol. 2024, 21, 1089–1108. [Google Scholar] [CrossRef]
- Karmakar, S.; Pal, P.; Lal, G. Key Activating and Inhibitory Ligands Involved in the Mobilization of Natural Killer Cells for Cancer Immunotherapies. Immunotargets Ther. 2021, 10, 387–407. [Google Scholar] [CrossRef]
- Graham, L.V.; Fisher, J.G.; Khakoo, S.I.; Blunt, M.D. Targeting KIR as a Novel Approach to Improve CAR-NK Cell Function. J. Transl. Genet. Genom. 2023, 7, 230–235. [Google Scholar] [CrossRef]
- Rettman, P.; Blunt, M.D.; Fulton, R.J.; Vallejo, A.F.; Bastidas-Legarda, L.Y.; España-Serrano, L.; Polak, M.E.; Al-Shamkhani, A.; Retiere, C.; Khakoo, S.I. Peptide: MHC-Based DNA Vaccination Strategy to Activate Natural Killer Cells by Targeting Killer Cell Immunoglobulin-Like Receptors. J. Immunother. Cancer 2021, 9, e001912. [Google Scholar] [CrossRef]

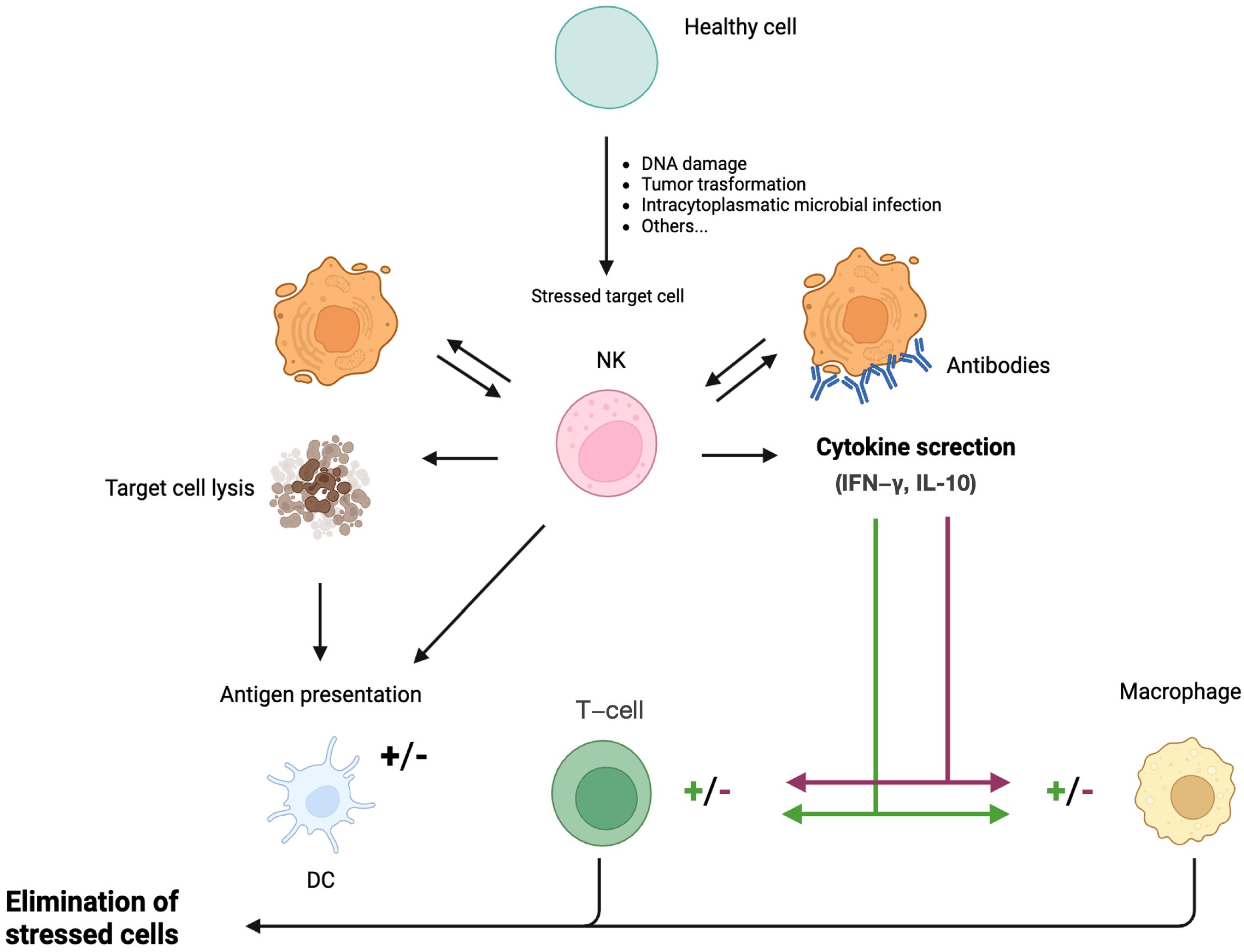

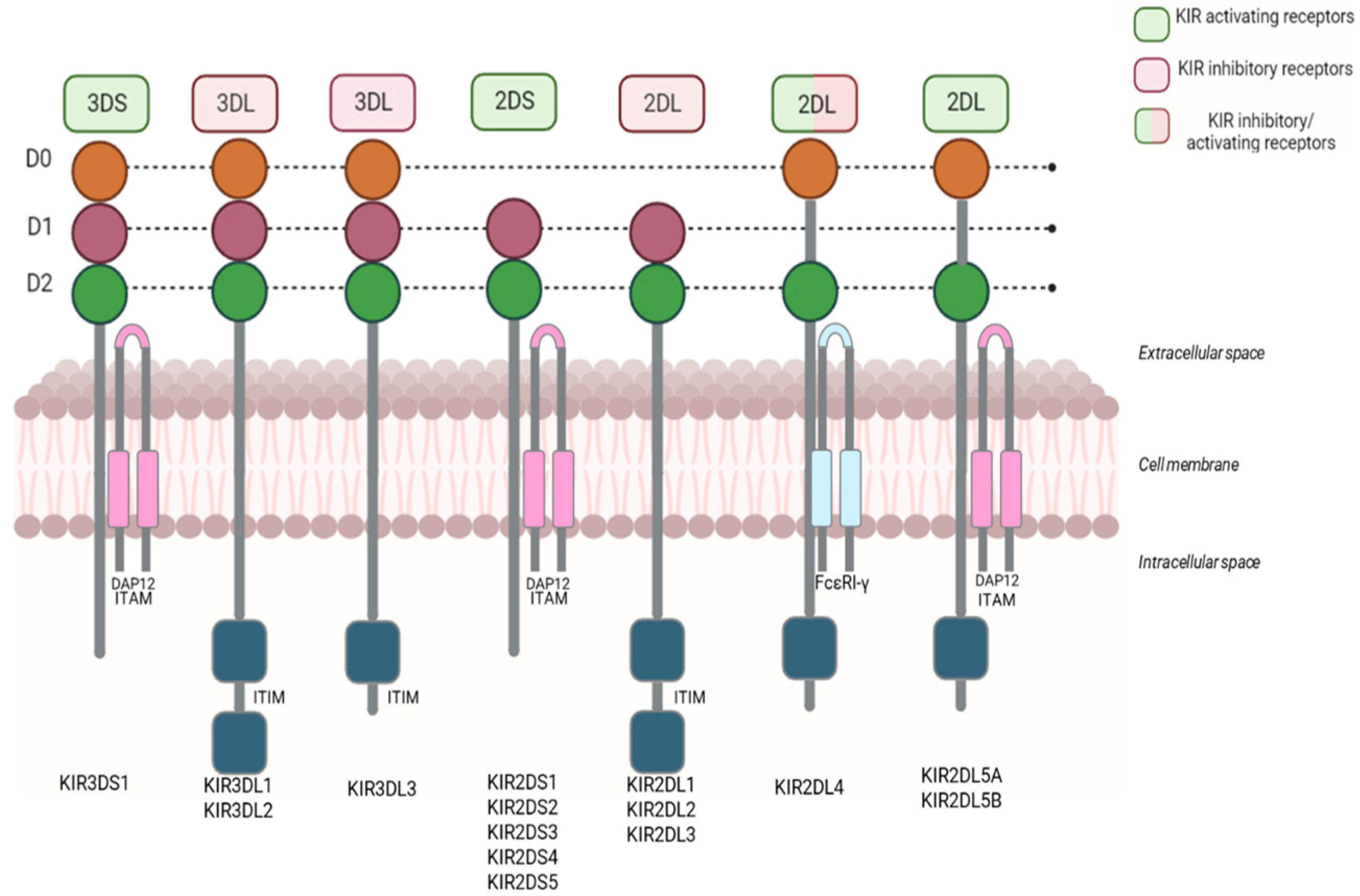
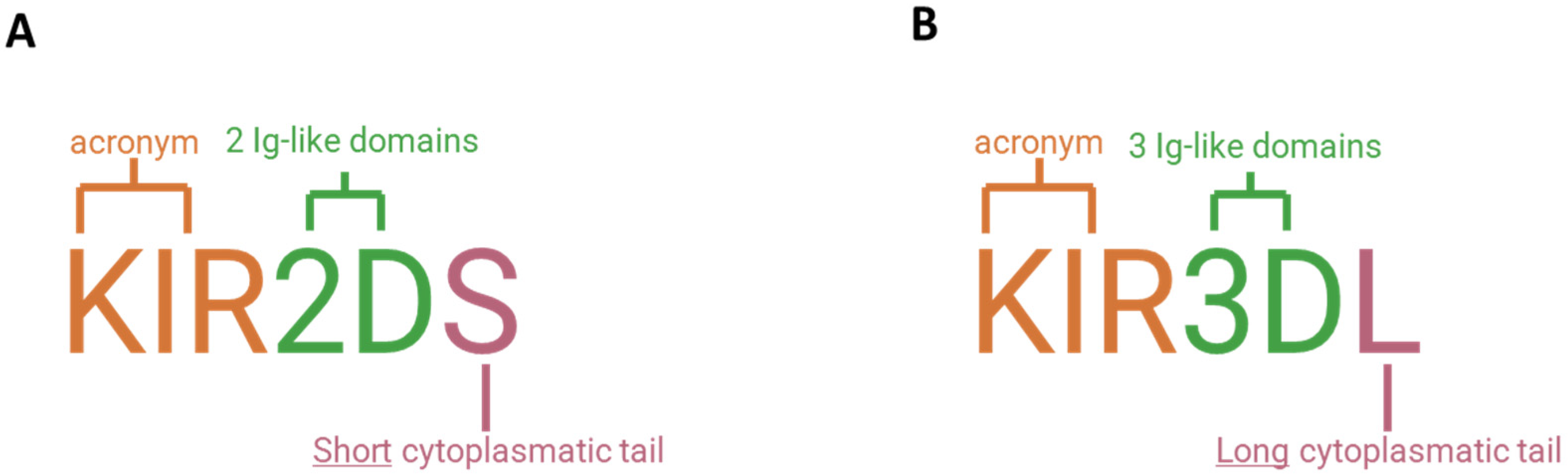



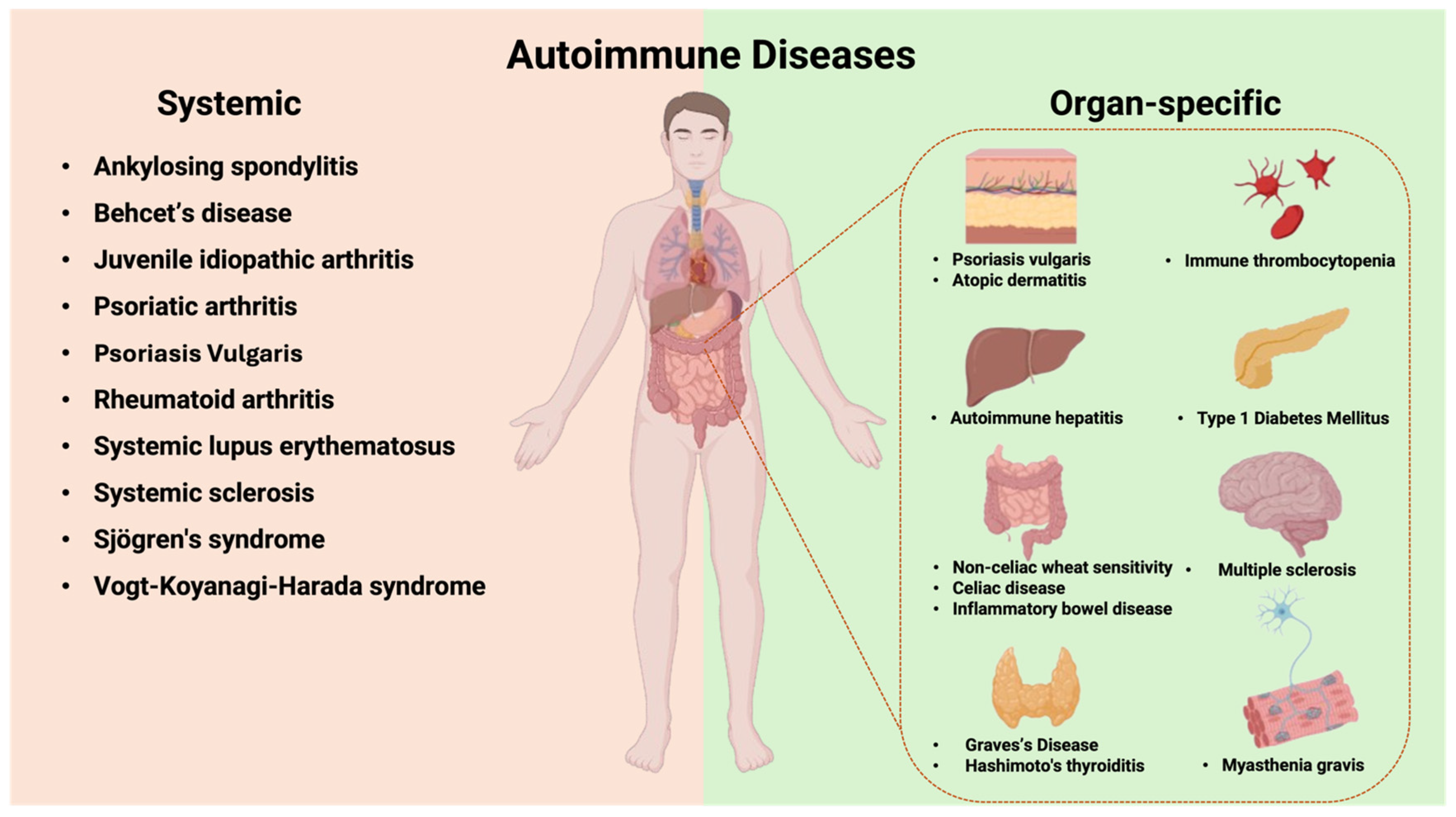
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agnello, L.; Masucci, A.; Tamburello, M.; Vassallo, R.; Massa, D.; Giglio, R.V.; Midiri, M.; Gambino, C.M.; Ciaccio, M. The Role of Killer Ig-like Receptors in Diseases from A to Z. Int. J. Mol. Sci. 2025, 26, 3242. https://doi.org/10.3390/ijms26073242
Agnello L, Masucci A, Tamburello M, Vassallo R, Massa D, Giglio RV, Midiri M, Gambino CM, Ciaccio M. The Role of Killer Ig-like Receptors in Diseases from A to Z. International Journal of Molecular Sciences. 2025; 26(7):3242. https://doi.org/10.3390/ijms26073242
Chicago/Turabian StyleAgnello, Luisa, Anna Masucci, Martina Tamburello, Roberta Vassallo, Davide Massa, Rosaria Vincenza Giglio, Mauro Midiri, Caterina Maria Gambino, and Marcello Ciaccio. 2025. "The Role of Killer Ig-like Receptors in Diseases from A to Z" International Journal of Molecular Sciences 26, no. 7: 3242. https://doi.org/10.3390/ijms26073242
APA StyleAgnello, L., Masucci, A., Tamburello, M., Vassallo, R., Massa, D., Giglio, R. V., Midiri, M., Gambino, C. M., & Ciaccio, M. (2025). The Role of Killer Ig-like Receptors in Diseases from A to Z. International Journal of Molecular Sciences, 26(7), 3242. https://doi.org/10.3390/ijms26073242