
VMP1 Constitutive Expression in Mice Dampens Pancreatic and Systemic Histopathological Damage in an Experimental Model of Severe Acute Pancreatitis

 , , and
, , and
Abstract
1. Introduction
2. Results
2.1. Intrapancreatic Features
2.1.1. ElaI-VMP1 Mice Exhibit Lower Serum Enzyme Concentrations, Reduced Inflammatory Signs, and Less Necrosis During Experimental SAP
- Edema Analysis
- Inflammatory Infiltration
- Acinar Necrosis
- Fat Necrosis and Hemorrhage
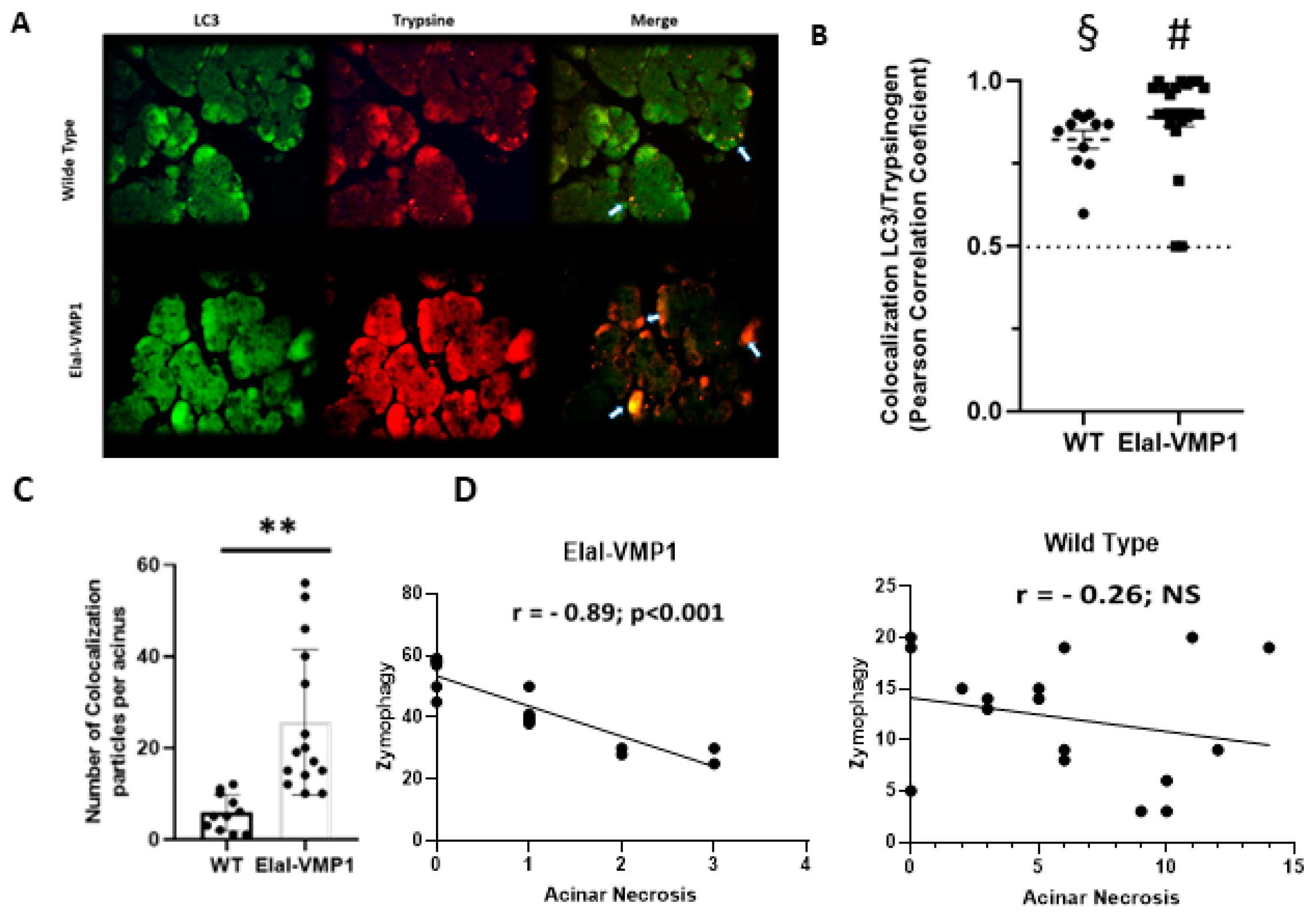
2.1.2. ElaI-VMP1 Mice Exhibit Higher Levels of Zymophagy, Which Significantly Correlates with Lower Acinar Necrosis During Experimental SAP
2.2. Extrapancreatic Features
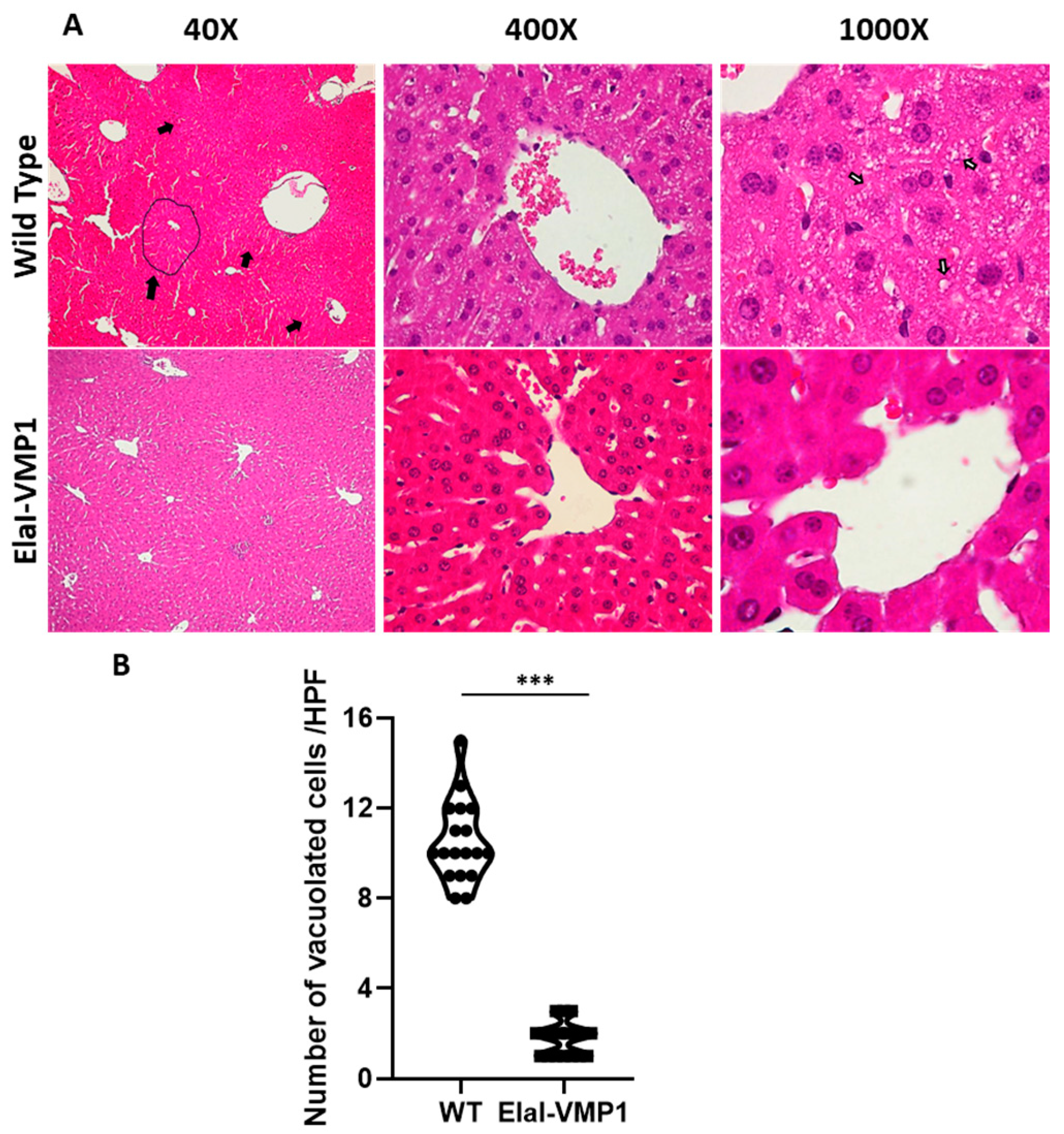
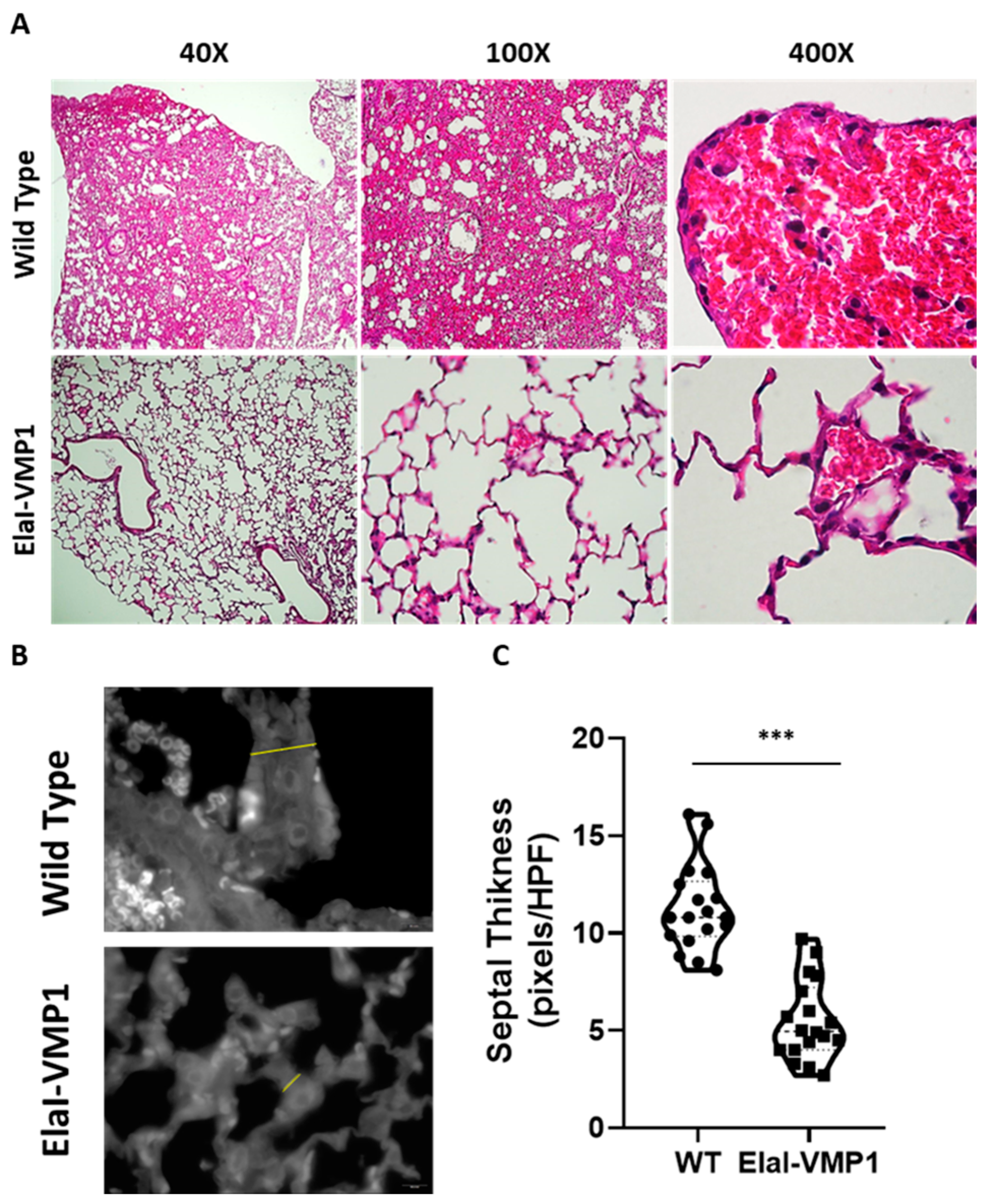
ElaI-VMP1 Mice Exhibited Preserved Liver Histology, Normal Kidney Histoarchitecture, and Clean Lung Tissue During Experimental SAP
- Liver
- Kidneys
- Lungs
3. Discussion
4. Materials and Methods
- Mice
- Transgenic Mice (ElaI-VMP1 Mice)
- CAE/EK-induced Pancreatitis Model
- Biochemical Markers
- Histological Studies
- ○
- Pancreatic Tissue
- ○
- Extra-pancreatic Tissue Damage
- Liver Damage: At LPF (i.e., 40×), the presence and localization of necrotic areas (e.g., centrilobular) were considered. At HPF, histological evidence of loss of hepatocyte membrane integrity, signs of tissue necrosis (e.g., cytoplasm swelling and/or intracellular vacuoles), and areas of leukocyte infiltration, especially in centrilobular areas, were examined;
- Kidney Damage: Kidney damage was assessed by examining areas of acute tubular necrosis with patchy loss of tubular epithelial cells, desquamation, pyknosis, and loss of nuclei, as well as the presence of intraluminal cellular debris;
- Lung Damage: For lung damage evaluation, particular emphasis was placed on septal integrity and thickness, as well as hemorrhagic foci in the alveolar spaces. The presence of leukocyte infiltration was noted, though it was not the central focus of the lung damage assessment.
- Antibodies
- IF Assays
- Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ANOVA | Analysis of Variance |
| AP | Acute Pancreatitis |
| CAE | Caerulein |
| CCK | Cholecystokinin |
| CCK-R | Cholecystokinin Receptor |
| EK | Enterokinase |
| ElaI-VMP1 | C57BL6J-ElaI-VMP1 transgenic mice |
| FXR | Farnesoid X receptor |
| H&E | Hematoxylin and Eosin |
| HPF | High-Power Field |
| IF | Immunofluorescence |
| IQR | Interquartile Range |
| Lamp2 | Lysosome-Associated Membrane Glycoprotein 2 |
| LC3 | Microtubule-Associated Protein 1A/1B-light chain 3 |
| LDH | Lactate Dehydrogenase |
| LPF | Low-Power Field |
| mTOR | Mammalian Target of Rapamycin |
| mTORC1 | Mammalian Target of Rapamycin Complex 1 |
| NF-κB | Nuclear Transcription Factor-κB |
| NS | Non-Significant |
| Osgin1 | Oxidative Stress-Induced Growth Inhibitor 1 |
| PBS | Phosphate-Buffered Saline |
| S6K1 | Serine/threonine Kinase 1 |
| SAP | Severe Acute Pancreatitis |
| SD | Standard Deviation |
| SEM | Standard Error of the Mean |
| VMP1 | Vacuole Membrane Protein 1 |
| WT | Wild-Type |
References
- Habtezion, A.; Gukovskaya, A.S.; Pandol, S.J. Acute Pancreatitis: A Multifaceted Set of Organelle and Cellular Interactions. Gastroenterology 2019, 156, 1941–1950. [Google Scholar] [CrossRef]
- Lee, P.J.; Papachristou, G.I. New insights into acute pancreatitis. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 479–496. [Google Scholar] [CrossRef]
- Banks, P.A.; Bollen, T.L.; Dervenis, C.; Gooszen, H.G.; Johnson, C.D.; Sarr, M.G.; Tsiotos, G.G.; Vege, S.S.; Acute Pancreatitis Classification Working Group. Classification of acute pancreatitis—2012: Revision of the Atlanta classification and definitions by international consensus. Gut 2013, 62, 102–111. [Google Scholar] [CrossRef]
- Dumnicka, P.; Maduzia, D.; Ceranowicz, P.; Olszanecki, R.; Drożdż, R.; Kuśnierz-Cabala, B. The Interplay between Inflammation, Coagulation and Endothelial Injury in the Early Phase of Acute Pancreatitis: Clinical Implications. Int. J. Mol. Sci. 2017, 18, 354. [Google Scholar] [CrossRef]
- Huang, L.; Jiang, Y.; Sun, Z.; Gao, Z.; Wang, J.; Zhang, D. Autophagy Strengthens Intestinal Mucosal Barrier by Attenuating Oxidative Stress in Severe Acute Pancreatitis. Dig. Dis. Sci. 2018, 63, 910–919. [Google Scholar] [CrossRef]
- Zhang, Z.H.; Jhaveri, D.J.; Marshall, V.M.; Bauer, D.C.; Edson, J.; Narayanan, R.K.; Robinson, G.J.; Lundberg, A.E.; Bartlett, P.F.; Wray, N.R.; et al. A Comparative Study of Techniques for Differential Expression Analysis on RNA-Seq Data. PLoS ONE 2014, 9, e103207. [Google Scholar] [CrossRef]
- Hartwig, W.; Schimmel, E.; Hackert, T.; Fortunato, F.; Bergmann, F.; Baczako, A.; Strobel, O.; Büchler, M.W.; Werner, J. A novel animal model of severe pancreatitis in mice and its differences to the rat. Surgery 2008, 144, 394–403. [Google Scholar] [CrossRef]
- Hartwig, W.; Kolvenbach, M.; Hackert, T.; Fortunato, F.; Schneider, L.; Büchler, M.W.; Werner, J. Enterokinase induces severe necrosis and rapid mortality in cerulein pancreatitis: Characterization of a novel noninvasive rat model of necro-hemorrhagic pancreatitis. Surgery 2007, 142, 327–336. [Google Scholar] [CrossRef]
- Malla, S.R.; Krueger, B.; Wartmann, T.; Sendler, M.; Mahajan, U.M.; Weiss, F.U.; Thiel, F.G.; De Boni, C.; Gorelick, F.S.; Halangk, W.; et al. Early trypsin activation develops independently of autophagy in caerulein-induced pancreatitis in mice. Cell. Mol. Life Sci. 2020, 77, 1811–1825. [Google Scholar] [CrossRef]
- Grasso, D.; Renna, F.J.; Vaccaro, M.I. Initial Steps in Mammalian Autophagosome Biogenesis. Front. Cell Dev. Biol. 2018, 6, 146. [Google Scholar] [CrossRef]
- Klionsky, D.J.; Abdel-Aziz, A.K.; Abdelfatah, S.; Abdellatif, M.; Abdoli, A.; Abel, S.; Abeliovich, H.; Abildgaard, M.H.; Abudu, Y.P.; Acevedo-Arozena, A.; et al. Guidelines for the use and interpretation of assays for monitoring autophagy (4th edition)1. Autophagy 2021, 17, 1–382. [Google Scholar] [CrossRef]
- Levine, B.; Packer, M.; Codogno, P. Development of autophagy inducers in clinical medicine. J. Clin. Investig. 2015, 125, 14–24. [Google Scholar] [CrossRef]
- Johansen, T.; Lamark, T. Selective Autophagy: ATG8 Family Proteins, LIR Motifs and Cargo Receptors. J. Mol. Biol. 2020, 432, 80–103. [Google Scholar] [CrossRef]
- Dusetti, N.J.; Jiang, Y.; Vaccaro, M.I.; Tomasini, R.; Samir, A.A.; Calvo, E.L.; Ropolo, A.; Fiedler, F.; Mallo, G.V.; Dagorn, J.-C.; et al. Cloning and Expression of the Rat Vacuole Membrane Protein 1 (VMP1), a New Gene Activated in Pancreas with Acute Pancreatitis, Which Promotes Vacuole Formation. Biochem. Biophys. Res. Commun. 2002, 290, 641–649. [Google Scholar] [CrossRef]
- Ropolo, A.; Grasso, D.; Pardo, R.; Sacchetti, M.L.; Archange, C.; Lo Re, A.; Seux, M.; Nowak, J.; Gonzalez, C.D.; Iovanna, J.L.; et al. The Pancreatitis-induced Vacuole Membrane Protein 1 Triggers Autophagy in Mammalian Cells. J. Biol. Chem. 2007, 282, 37124–37133. [Google Scholar] [CrossRef]
- MMolejon, I.; Ropolo, A.; Re, A.L.; Boggio, V.; Vaccaro, M.I. The VMP1-Beclin 1 interaction regulates autophagy induction. Sci. Rep. 2013, 3, 1055. [Google Scholar] [CrossRef]
- Nascimbeni, A.C.; Giordano, F.; Dupont, N.; Grasso, D.; Vaccaro, M.I.; Codogno, P.; Morel, E. ER–plasma membrane contact sites contribute to autophagosome biogenesis by regulation of local PI 3P synthesis. EMBO J. 2017, 36, 2018–2033. [Google Scholar] [CrossRef]
- Grasso, D.; Ropolo, A.; Ré, A.L.; Boggio, V.; Molejón, M.I.; Iovanna, J.L.; Gonzalez, C.D.; Urrutia, R.; Vaccaro, M.I. Zymophagy, a Novel Selective Autophagy Pathway Mediated by VMP1-USP9x-p62, Prevents Pancreatic Cell Death*. J. Biol. Chem. 2011, 286, 8308–8324. [Google Scholar] [CrossRef]
- Gukovskaya, A.S.; Gukovsky, I. Autophagy and pancreatitis. Am. J. Physiol.-Gastrointest. Liver Physiol. 2012, 303, G993–G1003. [Google Scholar] [CrossRef]
- Mayerle, J.; Sendler, M.; Hegyi, E.; Beyer, G.; Lerch, M.M.; Sahin-Tóth, M. Genetics, Cell Biology, and Pathophysiology of Pancreatitis. Gastroenterology 2019, 156, 1951–1968.e1. [Google Scholar] [CrossRef]
- Murr, M.M.; Yang, J.; Fier, A.; Kaylor, P.; Mastorides, S.; Norman, J.G. Pancreatic Elastase Induces Liver Injury by Activating Cytokine Production Within Kupffer Cells via Nuclear Factor-Kappa B. J. Gastrointest. Surg. 2002, 6, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Kazantsev, G.B.; Hecht, D.W.; Rao, R.; Fedorak, I.J.; Gattuso, P.; Thompson, K.; Djuricin, G.; Prinz, R.A. Plasmid labeling confirms bacterial translocation in pancreatitis. Am. J. Surg. 1994, 167, 201–207. [Google Scholar] [CrossRef]
- Kes, P.; VuČIČEviĆ, Ž.; RatkoviĆ-GusiĆ, I.; Fotivec, A. Acute Renal Failure Complicating Severe Acute Pancreatitis. Ren. Fail. 1996, 18, 621–628. [Google Scholar] [CrossRef]
- Ruan, Q.; Lu, H.; Zhu, H.; Guo, Y.; Bai, Y. A network-regulative pattern in the pathogenesis of kidney injury following severe acute pancreatitis. Biomed. Pharmacother. 2020, 125, 109978. [Google Scholar] [CrossRef]
- Bahtia, M. Apoptosis versus necrosis in acute pancreatitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 286, G189–G196. [Google Scholar] [CrossRef]
- Hartwig, W.; Jimenez, R.E.; Werner, J.; Lewandrowski, K.B.; Warshaw, A.L.; Castillo, C.F. Interstitial trypsinogen release and its relevance to the transformation of mild into necrotizing pancreatitis in rats. Gastroenterology 1999, 117, 717–725. [Google Scholar] [CrossRef]
- Hartwig, W.; Werner, J.; Jimenez, R.E.; Z’Graggen, K.; Weimann, J.; Lewandrowski, K.B.; Warshaw, A.L.; Castillo, C.F.-D. Trypsin and activation of circulating trypsinogen contribute to pancreatitis-associated lung injury. Am. J. Physiol.-Gastrointest. Liver Physiol. 1999, 277, G1008–G1016. [Google Scholar] [CrossRef]
- Zheng, Y.; Sun, W.; Wang, Z.; Liu, J.; Shan, C.; He, C.; Li, B.; Hu, X.; Zhu, W.; Liu, L.; et al. Activation of Pancreatic Acinar FXR Protects against Pancreatitis via Osgin1-Mediated Restoration of Efficient Autophagy. Research 2022, 2022, 978081. [Google Scholar] [CrossRef]
- Kataura, T.; Tashiro, E.; Nishikawa, S.; Shibahara, K.; Muraoka, Y.; Miura, M.; Sakai, S.; Katoh, N.; Totsuka, M.; Onodera, M.; et al. A chemical genomics-aggrephagy integrated method studying functional analysis of autophagy inducers. Autophagy 2020, 17, 1856–1872. [Google Scholar] [CrossRef]
- Zhao, Z.; Han, L.; Xin, M.; Zhou, L.; Jiang, K.; Huang, Q.; Dai, R. Therapeutic TP effect of ucMSC-Ex via regulating mTOR to enhance autophagy. J. Cell Mol. Med. 2024, 28, e18120. [Google Scholar] [CrossRef]
- Mei, Q.; Zeng, Y.; Huang, C.; Zheng, J.; Guo, Y.; Fan, J.; Fu, X.; Wang, X.; Lu, Y. Rapamycin Alleviates Hypertriglyceridemia-Related Acute Pancreatitis via Restoring Autophagy Flux and Inhibiting Endoplasmic Reticulum Stress. Inflammation 2020, 43, 1510–1523. [Google Scholar] [CrossRef]
- Renna, F.; Steinberg, J.H.E.; Gonzalez, C.D.; Manifava, M.; Tadic, M.S.; Orquera, T.; Vecino, C.V.; Ropolo, A.; Guardavaccaro, D.; Rossi, M.; et al. Ubiquitination Is a Novel Post-Translational Modification of VMP1 in Autophagy of Human Tumor Cells. Int. J. Mol. Sci. 2023, 24, 12981. [Google Scholar] [CrossRef]
- Carrascal, M.; Areny-Balagueró, A.; De-Madaria, E.; Cárdenas-Jaén, K.; García-Rayado, G.; Rivera, R.; Mateos, R.M.M.; Pascual-Moreno, I.; Gironella, M.; Abian, J.; et al. Inflammatory capacity of exosomes released in the early stages of acute pancreatitis predicts the severity of the disease. J. Pathol. 2022, 256, 83–92. [Google Scholar] [CrossRef]
- Tadic, M.; Renna, F.J.; López, M.H.; Orquera, T.; Mingorance, F.L.; Ropolo, A.; Vaccaro, M.I. Unconventional secretion of the autophagy-related protein vmp1 via extracellular vesicles as a potential biomarker for pancreatic cell injury. bioRxiv 2024, bioRxiv:2024.10.31.615473. [Google Scholar] [CrossRef]
- Fernandez-Zapico, M.E.; Gonzalez-Paz, N.C.; Weiss, E.; Savoy, D.N.; Molina, J.R.; Fonseca, R.; Smyrk, T.C.; Chari, S.T.; Urrutia, R.; Billadeau, D.D. Ectopic expression of VAV1 reveals an unexpected role in pancreatic cancer tumorigenesis. Cancer Cell 2005, 7, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J.A.; Rattner, D.W.; Lewandrowski, K.; Compton, C.C.; Mandavilli, U.; Knoefel, W.T.; Warshaw, A.L. A Better Model of Acute Pancreatitis for Evaluating Therapy. Ann. Surg. 1992, 215, 44. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Histopathological Score | 0 | 1 | 2 | 3 | 4 |
|---|---|---|---|---|---|
| Edema | Absent | Diffuse expansion of Interlobar septae | “1” + Diffuse expansion of Interlobular septae | “2” + Diffuse expansion of Interacinar septae | “3” + Diffuse expansion of intercellular spaces |
| Inflammation and perivascular infiltrate | 0–5 intralobular or perivascular leukocytes/HPF | 6–15 intralobular or perivascular leukocytes/HPF | 16–25 intralobular or perivascular leukocytes/HPF | 26–35 intralobular or perivascular leukocytes/HPF | >35 intralobular or perivascular leukocytes/HPF |
| Acinar necrosis | Absent or Focal occurrence of 0–4 necrotic cells/HPF | Diffuse occurrence of 0–4 necrotic cells/HPF or Focal occurrence of 5–10 necrotic cells/HPF | Diffuse occurrence of 5–10 necrotic cells/HPF or Focal occurrence of 11–16 necrotic cells/HPF | Diffuse occurrence of 11–16 necrotic cells/HPF or Focal occurrence of >16 necrotic cells/HPF | >16 necrotic cells/HPF |
| Hemorrhage and fat necrosis | Absent | 1–3 foci | 4–5 foci | 6–7 foci | ≥8 foci |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boggio, V.; Gonzalez, C.D.; Zotta, E.; Ropolo, A.; Vaccaro, M.I. VMP1 Constitutive Expression in Mice Dampens Pancreatic and Systemic Histopathological Damage in an Experimental Model of Severe Acute Pancreatitis. Int. J. Mol. Sci. 2025, 26, 3196. https://doi.org/10.3390/ijms26073196
Boggio V, Gonzalez CD, Zotta E, Ropolo A, Vaccaro MI. VMP1 Constitutive Expression in Mice Dampens Pancreatic and Systemic Histopathological Damage in an Experimental Model of Severe Acute Pancreatitis. International Journal of Molecular Sciences. 2025; 26(7):3196. https://doi.org/10.3390/ijms26073196
Chicago/Turabian StyleBoggio, Veronica, Claudio Daniel Gonzalez, Elsa Zotta, Alejandro Ropolo, and Maria Ines Vaccaro. 2025. "VMP1 Constitutive Expression in Mice Dampens Pancreatic and Systemic Histopathological Damage in an Experimental Model of Severe Acute Pancreatitis" International Journal of Molecular Sciences 26, no. 7: 3196. https://doi.org/10.3390/ijms26073196
APA StyleBoggio, V., Gonzalez, C. D., Zotta, E., Ropolo, A., & Vaccaro, M. I. (2025). VMP1 Constitutive Expression in Mice Dampens Pancreatic and Systemic Histopathological Damage in an Experimental Model of Severe Acute Pancreatitis. International Journal of Molecular Sciences, 26(7), 3196. https://doi.org/10.3390/ijms26073196