
SIRT5 Alleviates Apoptosis of Vascular Endothelial Cells Under Simulated Microgravity via Desuccinylation of ERO1A

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
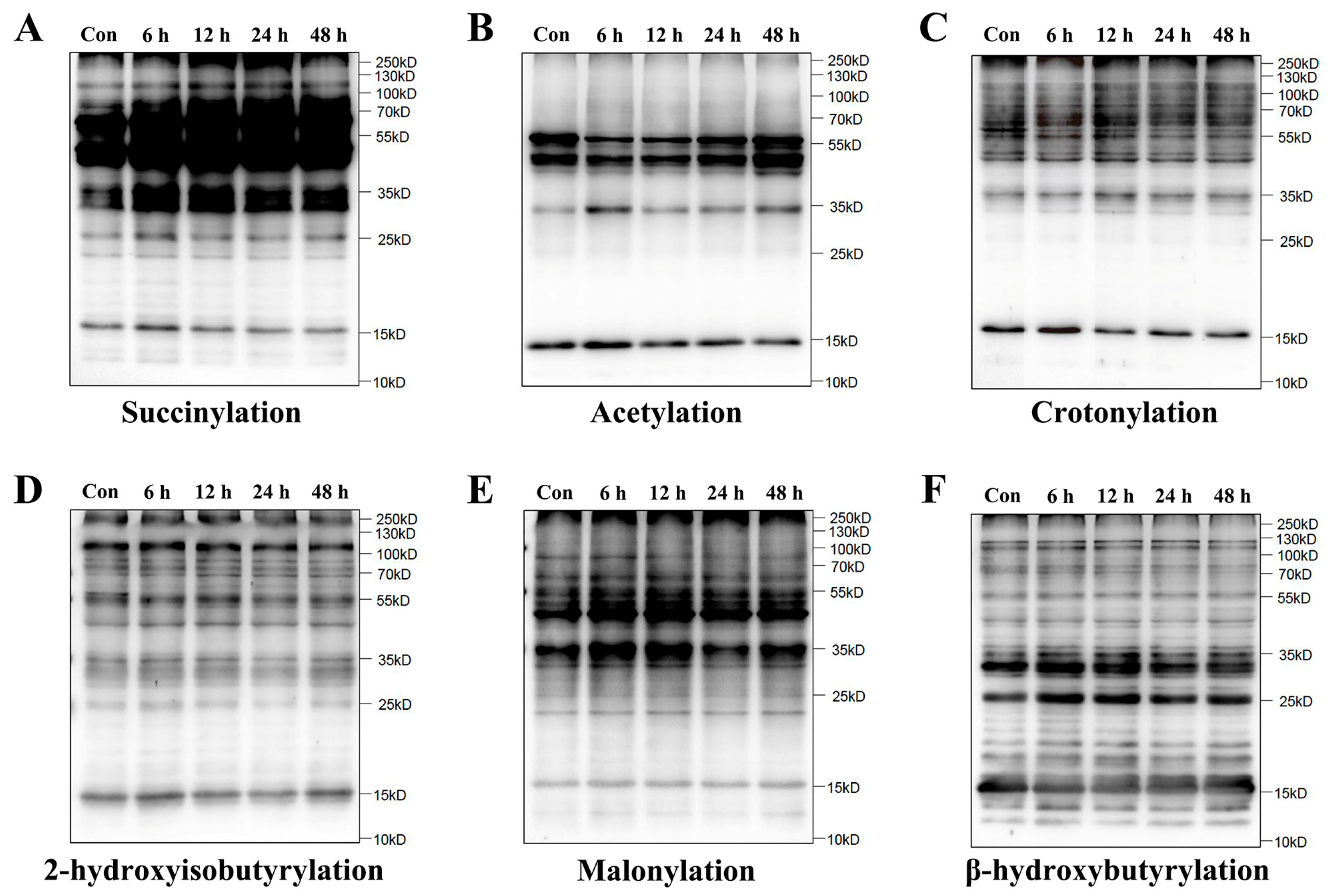
2.1. Increase in Lysine Succinylation in HUVECs Under Simulated Microgravity
2.2. Identification of Lysine-Succinylated Proteins and Sites in HUVECs Under Simulated Microgravity
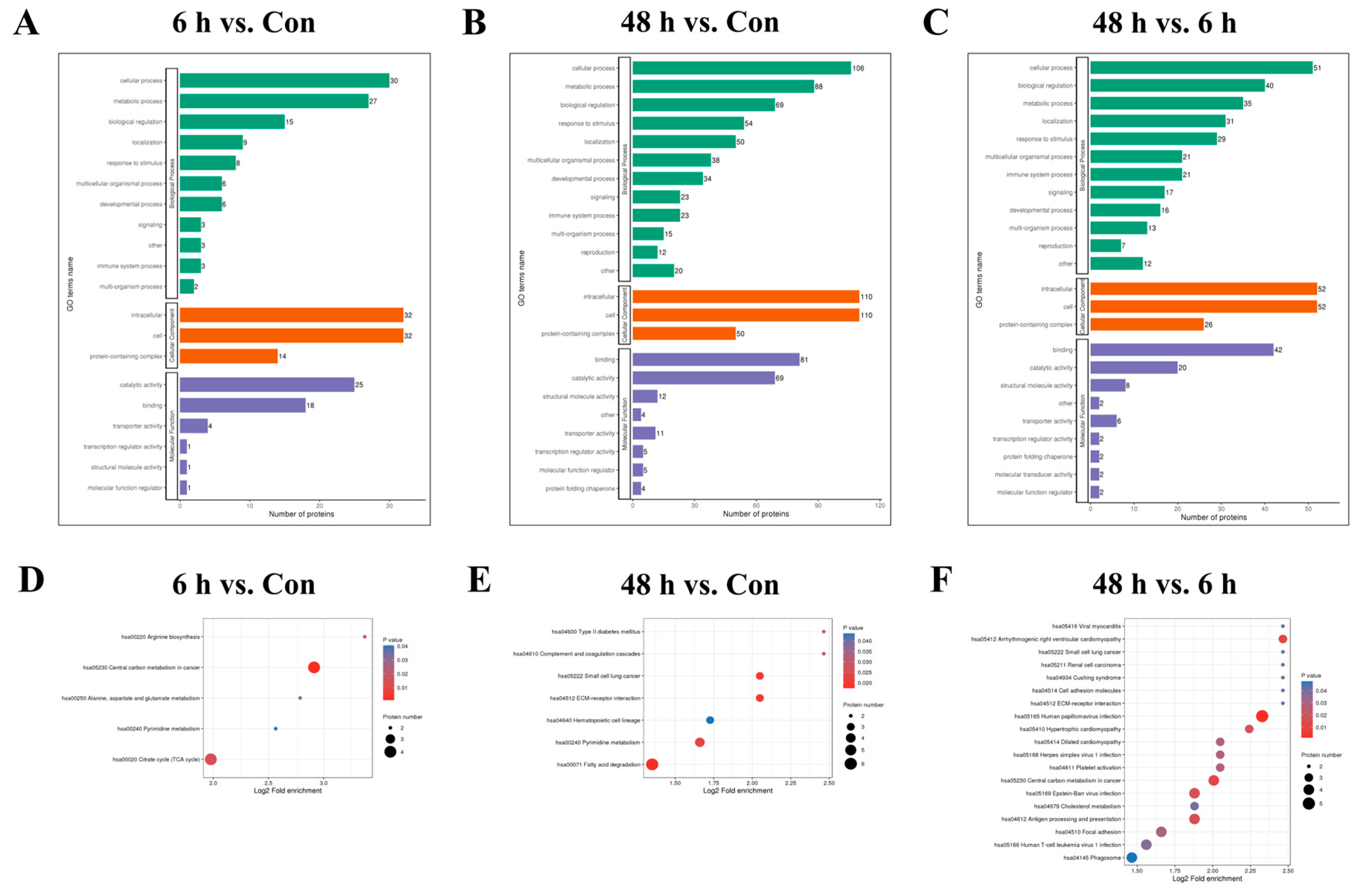
2.3. Functional Enrichment Analysis of Lysine-Succinylated Proteins
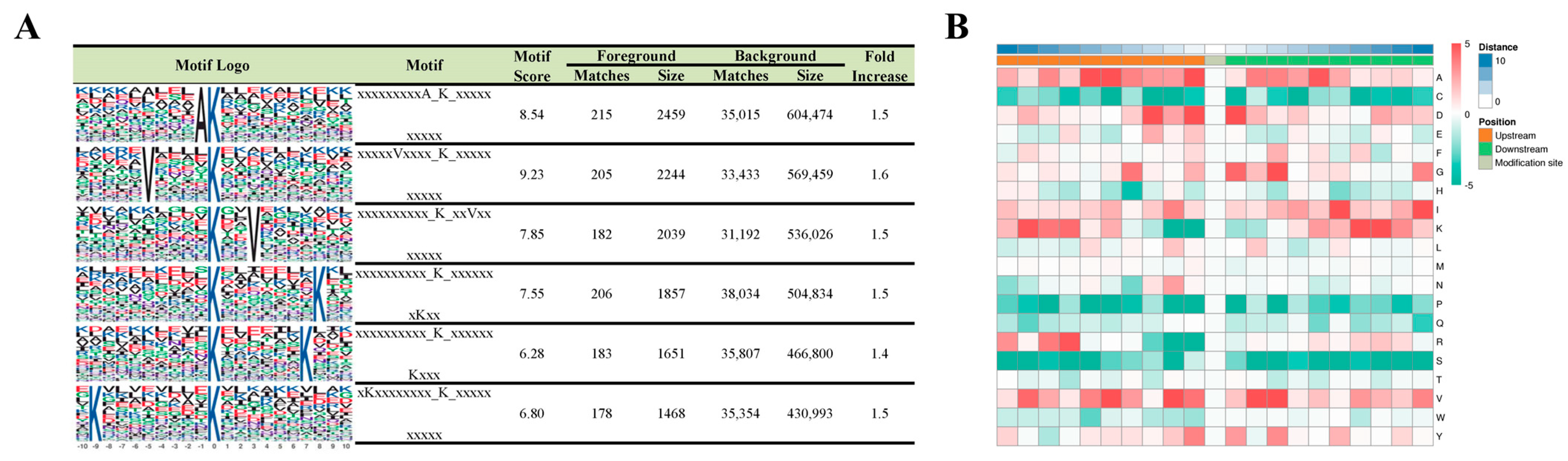
2.4. Motif Analysis of Lysine-Succinylated Proteins
2.5. SIRT5 Is Downregulated and Regulates Succinylation Modification Levels of HUVECs Under Simulated Microgravity
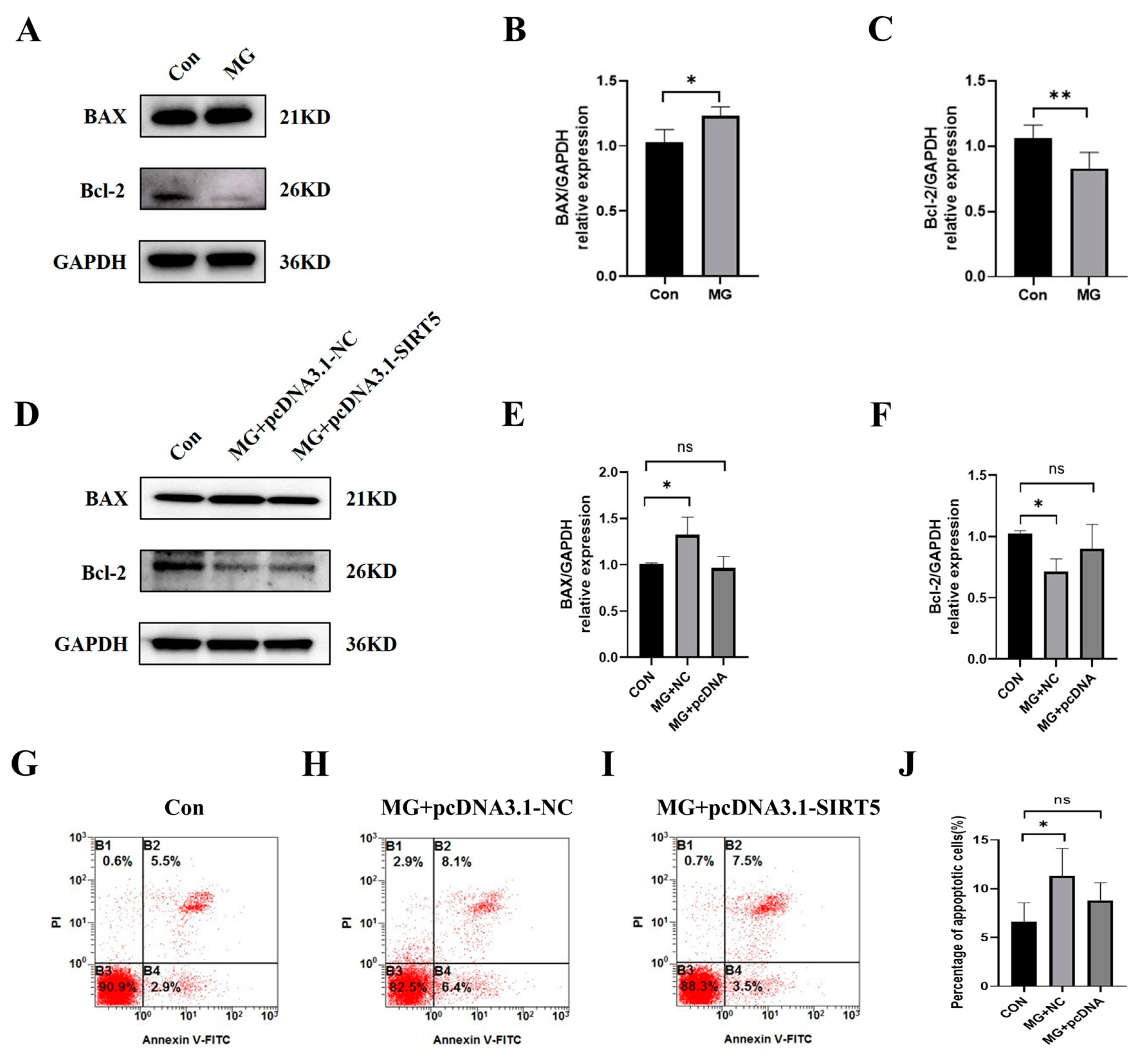
2.6. SIRT5 Protects HUVECs from Apoptosis Induced by Simulated Microgravity
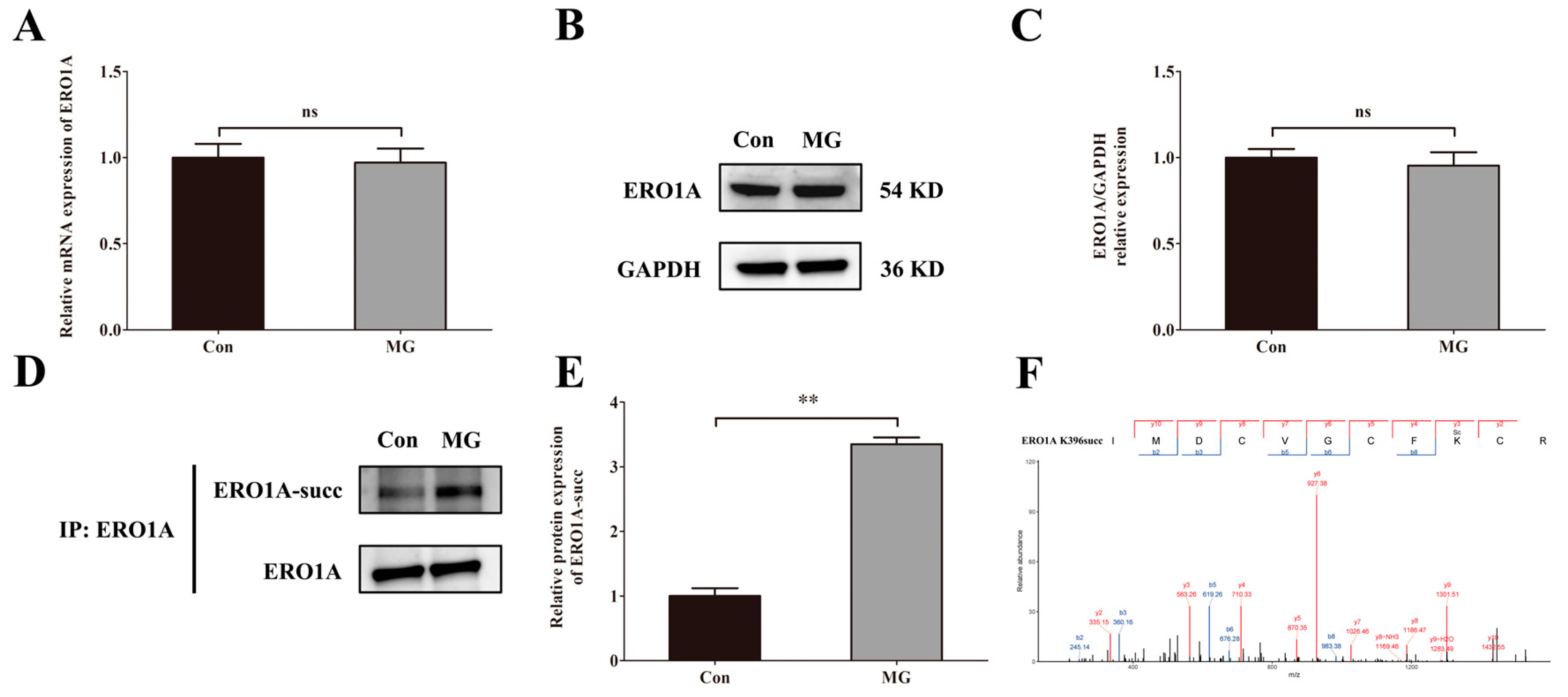
2.7. Simulated Microgravity Increases the Succinylation of ERO1A Protein at Site K396
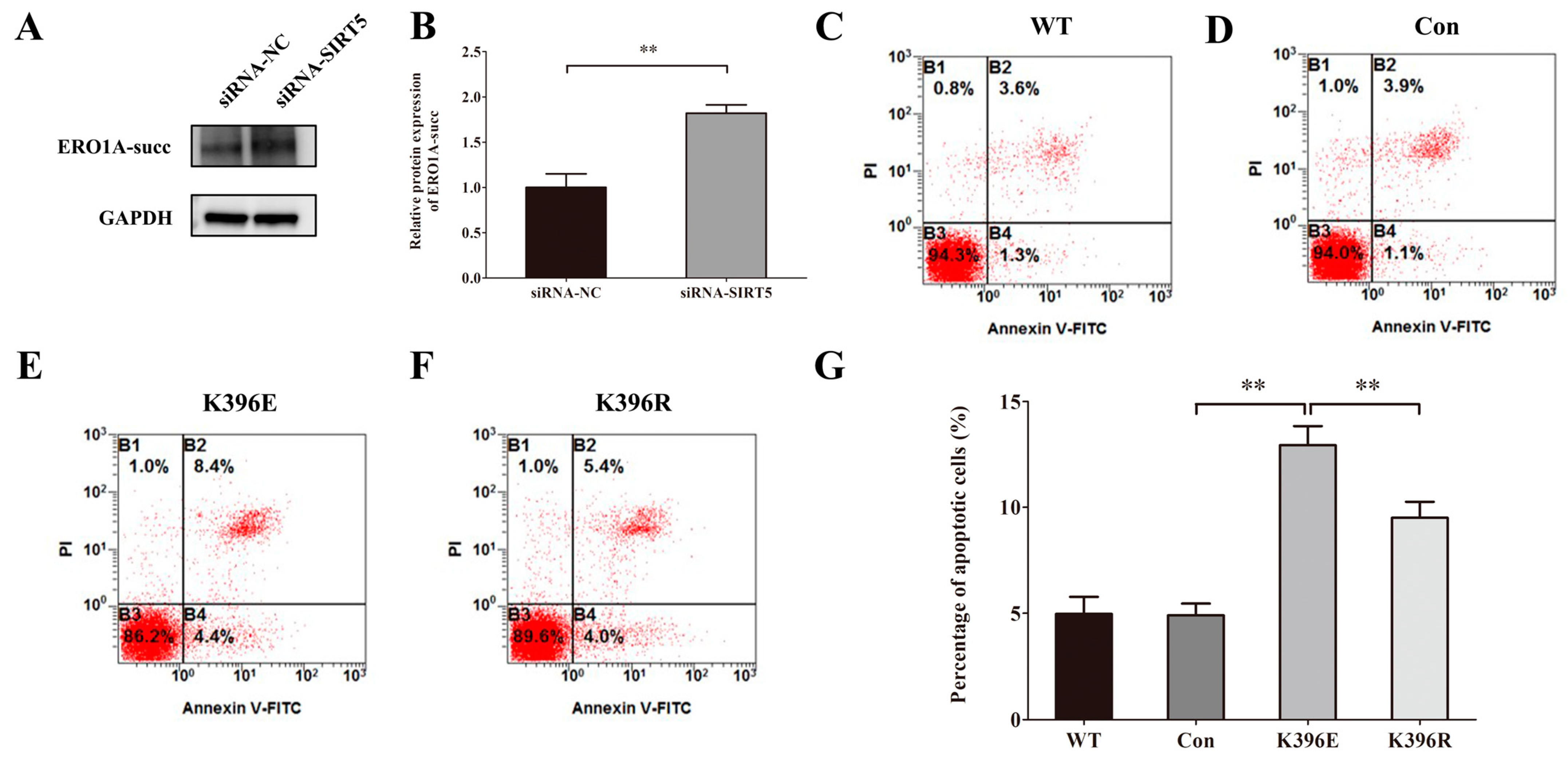
2.8. Knockdown of SIRT5 Induces Apoptosis of HUVECs Through the Succinylation of Lys396 in ERO1A
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Weightless Model Establishment
4.2. Cell Transfection and Treatment
4.3. Protein Extraction and Tryptic Digestion
4.4. Lysine-Succinylated Peptide Affinity Enrichment
4.5. LC-MS/MS
4.6. Database Search
4.7. Bioinformatic Analysis
4.8. Western Blot Assay
4.9. Coimmunoprecipitation (Co-IP)
4.10. Apoptosis Assay
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ahmed, S.S.; Goswami, N.; Sirek, A.; Green, D.A.; Winnard, A.; Fiebig, L.; Weber, T. Systematic review of the effectiveness of standalone passive countermeasures on microgravity-induced physiologic deconditioning. npj Microgravity 2024, 10, 48. [Google Scholar] [CrossRef] [PubMed]
- Murgia, M.; Rittweger, J.; Reggiani, C.; Bottinelli, R.; Mann, M.; Schiaffino, S.; Narici, M.V. Spaceflight on the ISS changed the skeletal muscle proteome of two astronauts. npj Microgravity 2024, 10, 60. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhao, Y.; Zhong, G.; Xu, Q.; Tan, Y.; Xing, W.; Cao, D.; Wang, Y.; Liu, C.; Li, J.; et al. Vascular smooth muscle cell-specific miRNA-214 deficiency alleviates simulated microgravity-induced vascular remodeling. FASEB J. 2024, 38, e23369. [Google Scholar] [CrossRef]
- Navasiolava, N.; Yuan, M.; Murphy, R.; Robin, A.; Coupé, M.; Wang, L.; Alameddine, A.; Gauquelin-Koch, G.; Gharib, C.; Li, Y.; et al. Vascular and Microvascular Dysfunction Induced by Microgravity and Its Analogs in Humans: Mechanisms and Countermeasures. Front. Physiol. 2020, 11, 952. [Google Scholar] [CrossRef]
- Locatelli, L.; Maier, J.A.M. Cytoskeletal Remodeling Mimics Endothelial Response to Microgravity. Front. Cell Dev. Biol. 2021, 9, 733573. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Shi, Y.; Qiu, C.; Zhao, J.; Gong, Y.; Nie, C.; Wu, B.; Yang, Y.; Wang, F.; Luo, L. Effects of Simulated Microgravity on Ultrastructure and Apoptosis of Choroidal Vascular Endothelial Cells. Front. Physiol. 2021, 11, 577325. [Google Scholar] [CrossRef]
- Li, C.; Pan, Y.; Tan, Y.; Wang, Y.; Sun, X. PINK1-Dependent Mitophagy Reduced Endothelial Hyperpermeability and Cell Migration Capacity Under Simulated Microgravity. Front. Cell Dev. Biol. 2022, 10, 896014. [Google Scholar] [CrossRef]
- Xu, G.; Cong, Y.-S.; Das, S. Regulation of gene expression at the post-translational modification level. Biochim. Biophys. Acta Gene Regul. Mech. 2024, 1867, 195060. [Google Scholar] [CrossRef]
- Zhang, N.; Wu, J.; Zheng, Q. Chemical proteomics approaches for protein post-translational modification studies. Biochim. Biophys. Acta Proteins Proteom. 2024, 1872, 141017. [Google Scholar] [CrossRef]
- Han, Y.; Deng, X.; Chen, H.; Chen, J.; Xu, W.; Liu, L. Succinylation modification-mediated upregulation of Sp1 promotes hepatocellular carcinoma cell proliferation. Discov. Oncol. 2024, 15, 660. [Google Scholar] [CrossRef]
- Guo, Y.; Wen, H.; Chen, Z.; Jiao, M.; Zhang, Y.; Ge, D.; Liu, R.; Gu, J. Conjoint analysis of succinylome and phosphorylome reveals imbalanced HDAC phosphorylation-driven succinylayion dynamic contibutes to lung cancer. Brief. Bioinform. 2024, 25, bbae415. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Zhang, J.; Li, J.; Song, Y.; Pan, J.; Mei, C.; Cui, M.; He, Q.; Wang, H.; Li, H.; et al. Sirtuin 5-Mediated Desuccinylation of ALDH2 Alleviates Mitochondrial Oxidative Stress Following Acetaminophen-Induced Acute Liver Injury. Adv. Sci. 2024, 11, e2402710. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Xie, C.; Sun, S.; Zheng, Q.; Wang, J.; Wang, Z.; Man, C.-H.; Wang, H.; Yang, Y.; Wang, T.; et al. SUCLG1 restricts POLRMT succinylation to enhance mitochondrial biogenesis and leukemia progression. EMBO J. 2024, 43, 2337–2367. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, J.; Cheng, Y.; Chen, K.; Chen, Y.; Zhu, H.; Li, Z.; Liu, S.; Cao, X. Metabolic enzyme Suclg2 maintains tolerogenicity of regulatory dendritic cells diffDCs by suppressing Lactb succinylation. J. Autoimmun. 2023, 138, 103048. [Google Scholar] [CrossRef]
- Lu, K.M.; Han, D. A review of the mechanism of succinylation in cancer. Medicine 2022, 101, e31493. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Miao, S.; Zhang, J.; Wang, P.; Liu, G.; Wang, J. The Growing Landscape of Succinylation Links Metabolism and Heart Disease. Epigenomics 2021, 13, 319–333. [Google Scholar] [CrossRef]
- Wang, S.; Li, Q.; Sun, X. SIRT5 participates in the suppressive tumor immune microenvironment of EGFR-mutant LUAD by regulating the succinylation of ACAT1. Heliyon 2024, 10, e39743. [Google Scholar] [CrossRef]
- Ramirez-Sagredo, A.; Sunny, A.T.; Cupp-Sutton, K.A.; Chowdhury, T.; Zhao, Z.; Wu, S.; Chiao, Y.A. Characterizing age-related changes in intact mitochondrial proteoforms in murine hearts using quantitative top-down proteomics. Clin. Proteom. 2024, 21, 57. [Google Scholar] [CrossRef]
- Oppedisano, F.; Nesci, S.; Spagnoletta, A. Mitochondrial sirtuin 3 and role of natural compounds: The effect of post-translational modifications on cellular metabolism. Crit. Rev. Biochem. Mol. Biol. 2024, 59, 199–220. [Google Scholar] [CrossRef]
- Wang, C.; Cui, W.; Yu, B.; Zhou, H.; Cui, Z.; Guo, P.; Yu, T.; Feng, Y. Role of succinylation modification in central nervous system diseases. Ageing Res. Rev. 2024, 95, 102242. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, F.; Chen, Y.; Ren, K.; Zhao, D.; Li, K.; Li, H.; Wan, X.; Peng, M.; Xiang, Z.; et al. Identification and analysis of drought-responsive F-box genes in upland rice and involvement of OsFBX148 in ABA response and ROS accumulation. BMC Plant Biol. 2024, 24, 1120. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zheng, Z.; Dai, X. SIRT5 induces autophagy and alleviates myocardial infarction via desuccinylation of TOM1. BMC Cardiovasc. Disord. 2024, 24, 464. [Google Scholar] [CrossRef]
- Chen, X.; Tian, M.; Sun, R.; Zhang, M.; Zhou, L.; Jin, L.; Chen, L.; Zhou, W.; Duan, K.; Chen, Y.; et al. SIRT5 inhibits peroxisomalACOX1 to prevent oxidative damage and is downregulated in liver cancer. Embo Rep. 2018, 19, e45124. [Google Scholar] [CrossRef]
- Mao, J.; Wang, D.; Wang, D.; Wu, Q.; Shang, Q.; Gao, C.; Wang, H.; Wang, H.; Du, M.; Peng, P.; et al. SIRT5-related desuccinylation modification of AIFM1 protects against compression-induced intervertebral disc degeneration by regulating mitochondrial homeostasis. Exp. Mol. Med. 2023, 55, 253–268. [Google Scholar] [CrossRef]
- Zheng, D.; Zeng, Q.; He, D.; He, Y.; Yang, J. SIRT5 alleviates hepatic ischemia and reperfusion injury by diminishing oxidative stress and inflammation via elevating SOD1 and IDH2 expression. Exp. Cell Res. 2022, 419, 113319. [Google Scholar] [CrossRef] [PubMed]
- Zito, E.; Guarrera, L.; Janssen-Heininger, Y.M. Fingerprint of the oxido-reductase ERO1: A protein disulfide bond producer and supporter of cancer. Biochim. Biophys. Acta Rev. Cancer 2024, 1879, 189027. [Google Scholar] [CrossRef]
- Shergalis, A.G.; Hu, S.; Bankhead, A., 3rd; Neamati, N. Role of the ERO1-PDI interaction in oxidative protein folding and disease. Pharmacol. Ther. 2020, 210, 107525. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Zhang, L.; Miao, Y.; Yang, J.; Wang, X.; Wang, C.-C.; Feng, J.; Wang, L. Homocysteine causes vascular endothelial dysfunction by disrupting endoplasmic reticulum redox homeostasis. Redox Biol. 2019, 20, 46–59. [Google Scholar] [CrossRef]
- Pan, Y.-K.; Li, C.-F.; Gao, Y.; Wang, Y.-C.; Sun, X.-Q. Effect of miR-27b-5p on apoptosis of human vascular endothelial cells induced by simulated microgravity. Apoptosis 2019, 25, 73–91. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, Y.; Zhang, Q.; Li, C.; Li, X.; Li, S.; Wang, Y.; Wang, R.; Fan, J.; Tie, Y.; Zhao, X.; et al. SIRT5 Alleviates Apoptosis of Vascular Endothelial Cells Under Simulated Microgravity via Desuccinylation of ERO1A. Int. J. Mol. Sci. 2025, 26, 2908. https://doi.org/10.3390/ijms26072908
Pan Y, Zhang Q, Li C, Li X, Li S, Wang Y, Wang R, Fan J, Tie Y, Zhao X, et al. SIRT5 Alleviates Apoptosis of Vascular Endothelial Cells Under Simulated Microgravity via Desuccinylation of ERO1A. International Journal of Molecular Sciences. 2025; 26(7):2908. https://doi.org/10.3390/ijms26072908
Chicago/Turabian StylePan, Yikai, Qian Zhang, Chengfei Li, Xi Li, Shuhan Li, Yuan Wang, Ruonan Wang, Jieyi Fan, Yateng Tie, Xingcheng Zhao, and et al. 2025. "SIRT5 Alleviates Apoptosis of Vascular Endothelial Cells Under Simulated Microgravity via Desuccinylation of ERO1A" International Journal of Molecular Sciences 26, no. 7: 2908. https://doi.org/10.3390/ijms26072908
APA StylePan, Y., Zhang, Q., Li, C., Li, X., Li, S., Wang, Y., Wang, R., Fan, J., Tie, Y., Zhao, X., Gao, Y., Wang, Y., & Sun, X. (2025). SIRT5 Alleviates Apoptosis of Vascular Endothelial Cells Under Simulated Microgravity via Desuccinylation of ERO1A. International Journal of Molecular Sciences, 26(7), 2908. https://doi.org/10.3390/ijms26072908