
Repeated Head Exposures to a 5G-3.5 GHz Signal Do Not Alter Behavior but Modify Intracortical Gene Expression in Adult Male Mice

 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
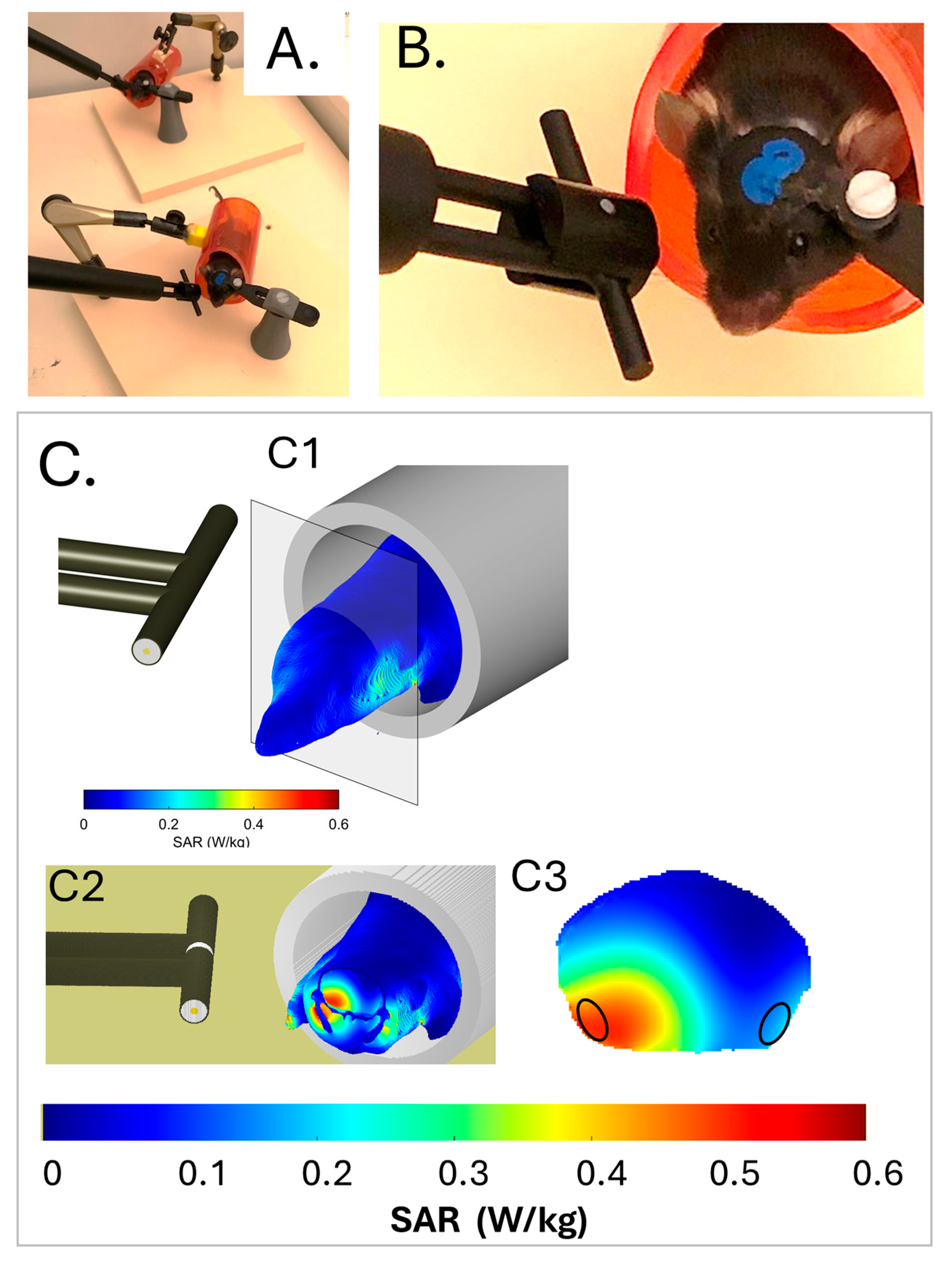
2.1. Development of a Calibrated Head Exposure System for Awake Mice
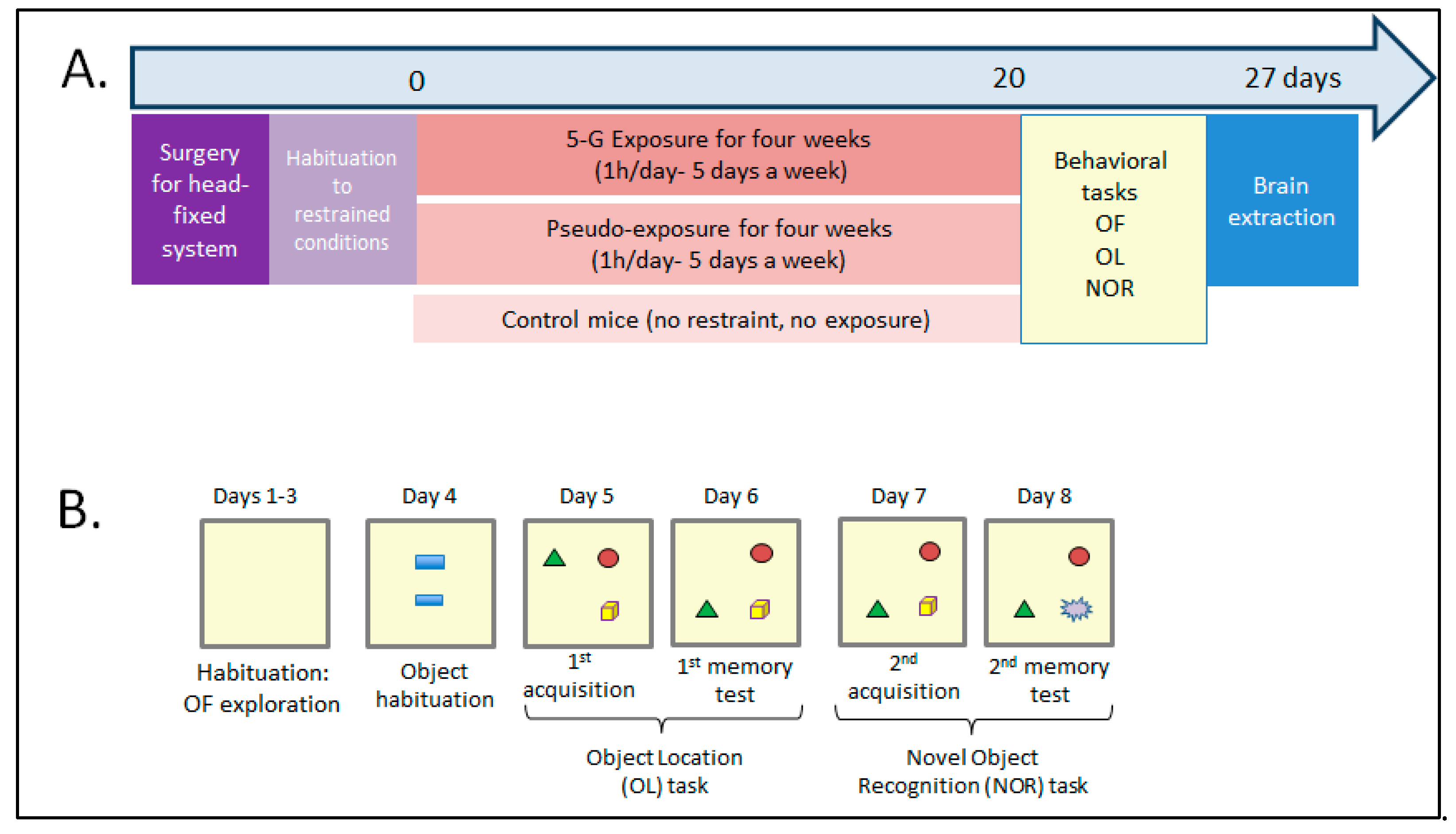
2.2. Assessing How Chronic 5G Exposure Affects Locomotion, Anxiety Level, or Memory
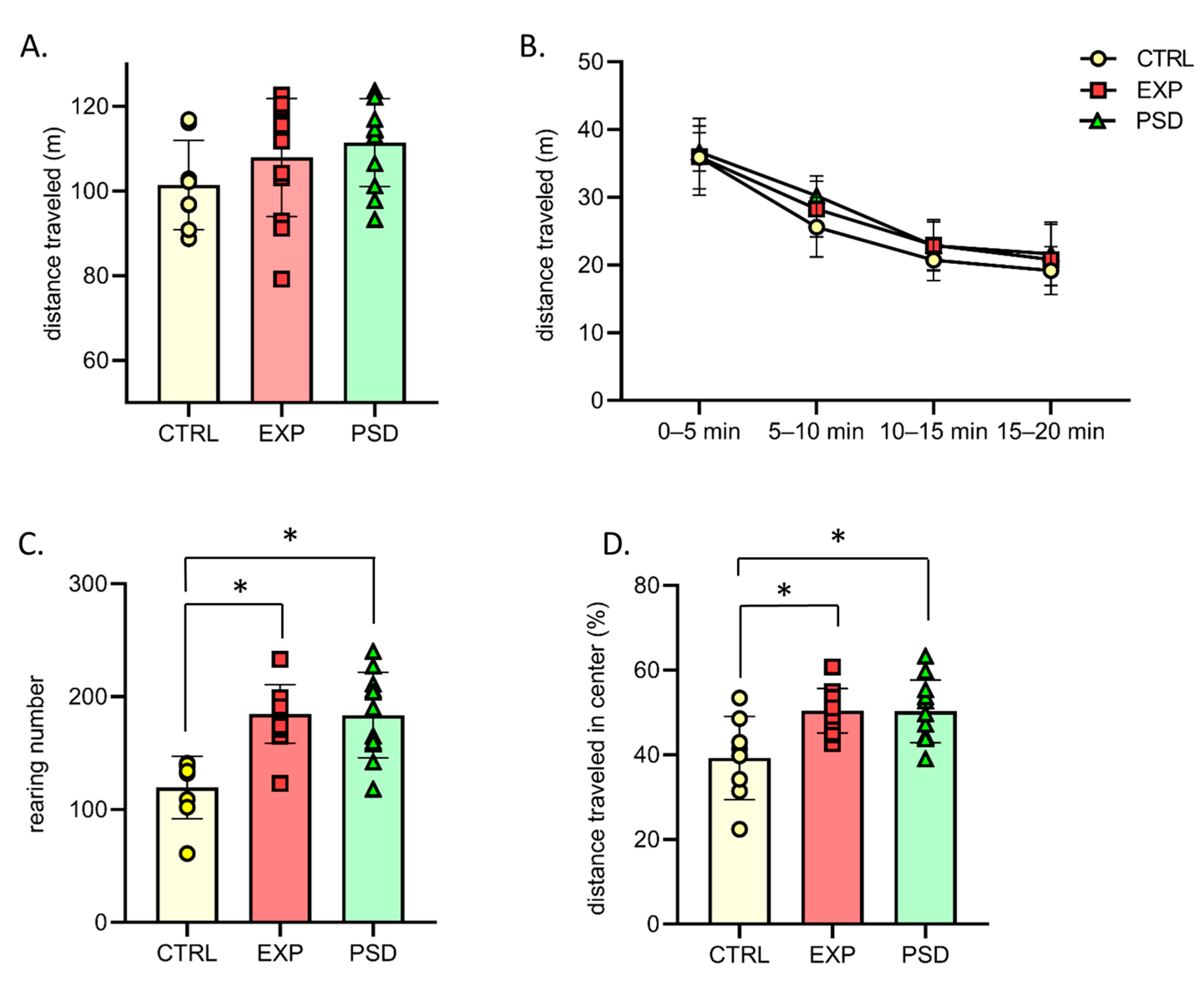
2.3. Chronic 5G Exposure Does Not Lead to Hyperactivity and Abnormal Anxiety
2.4. Chronic 5G Exposure Does Not Alter Object/Object Location Recognition Memory
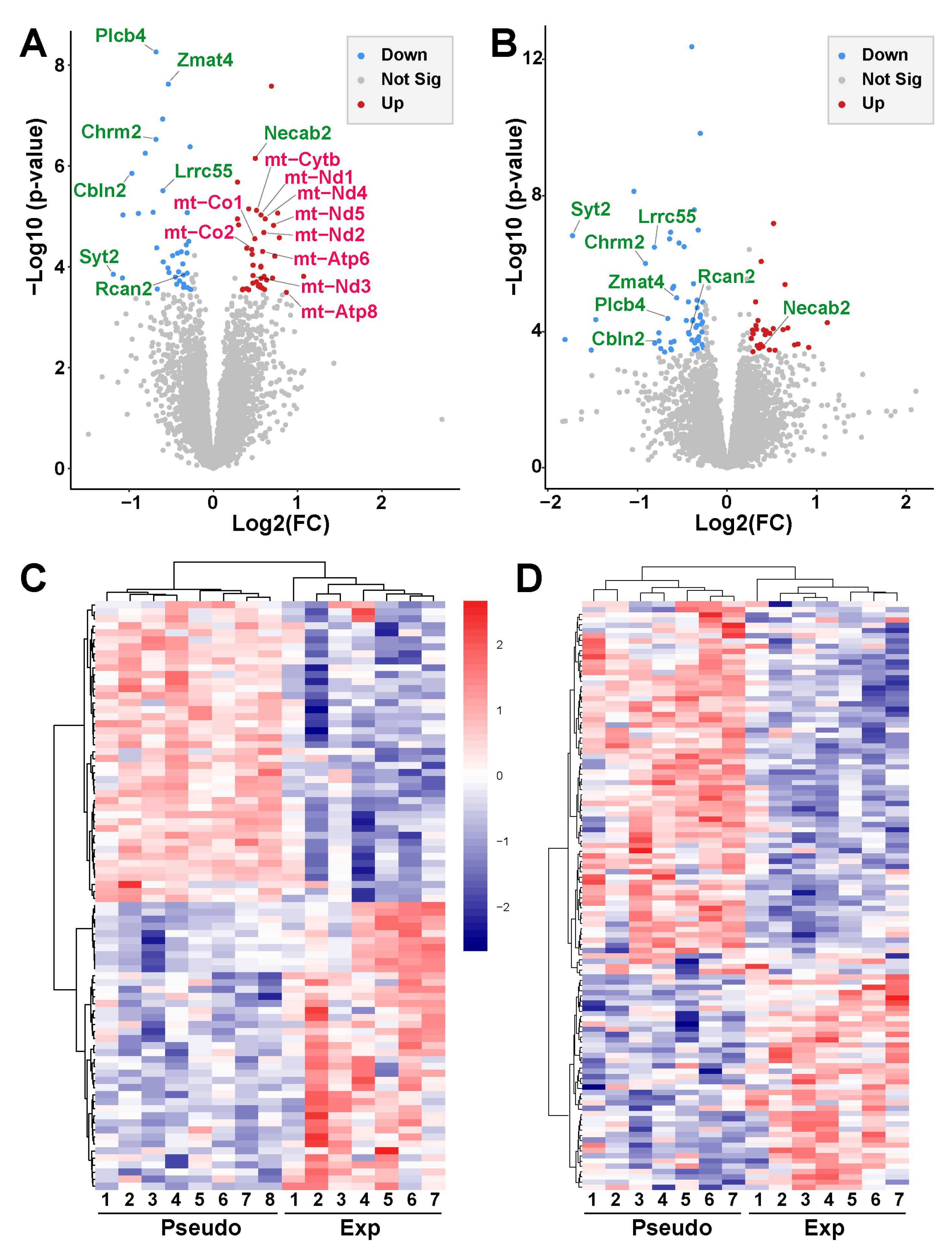
2.5. Effects of Chronic 5G Exposure on Gene Expression in the Cerebral Cortex
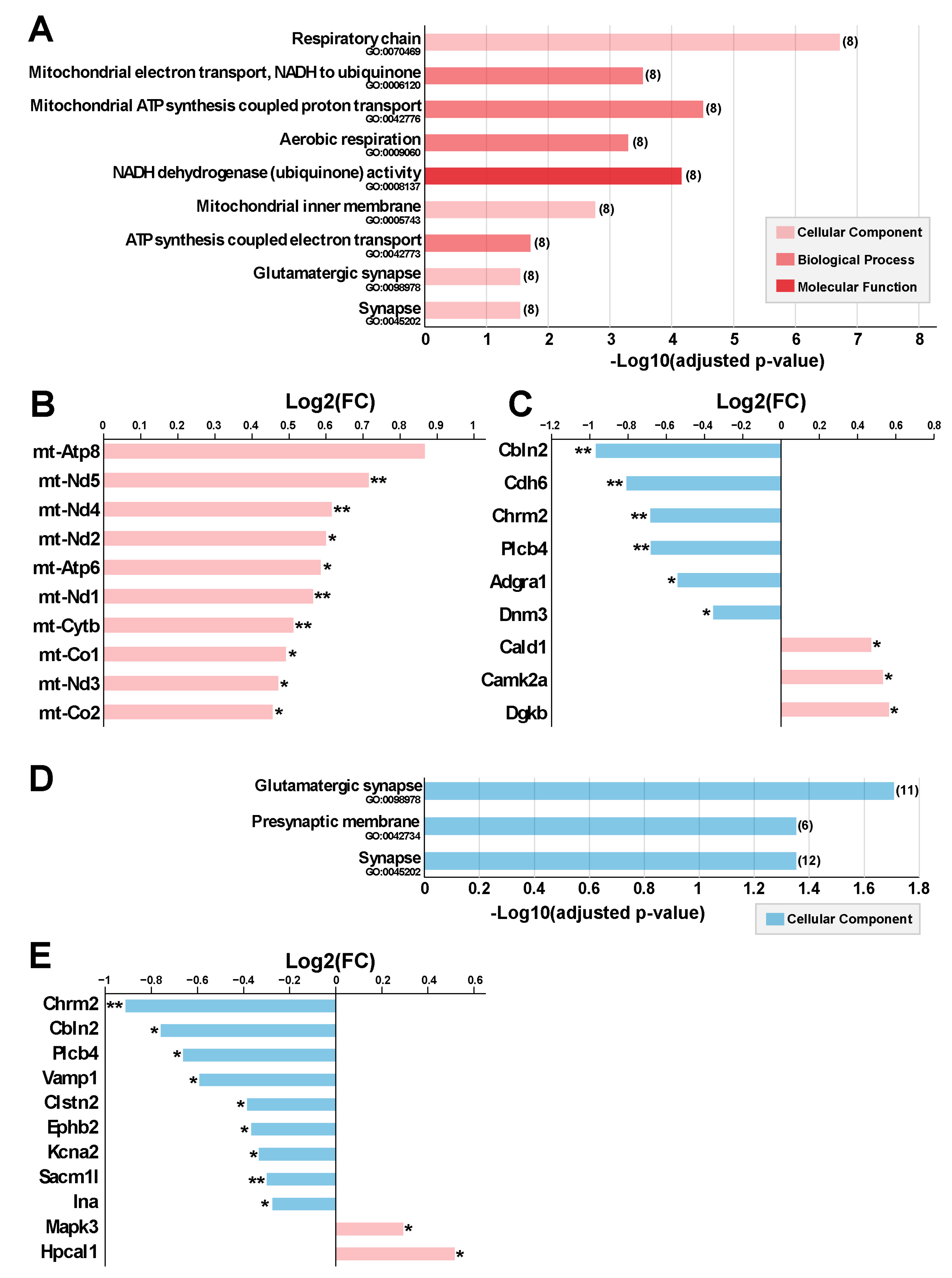
2.5.1. Chronic 5G Exposure Differentially Affects Gene Expression in the Right and Left Ent-Pir Cx
2.5.2. Chronic 5G Exposure Affects Genes Encoding Mitochondrial or Synapse Components
3. Discussion
4. Conclusions and Limitations
5. Materials and Methods
5.1. Subjects
5.2. Exposure System and Exposure Protocol to 5G-3.5 GHz Signals
5.3. Dosimetry
5.4. Behavioral Protocol
5.5. Statistical Analyses
5.6. Tissue Preparation for Transcriptome Analyses
5.7. RNA-Seq and Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ANOVA | Analysis of Variance |
| CTRL | Control |
| CNS | Central Nervous System |
| DEG | Differentially Expressed Genes |
| EMF | Electromagnetic Fields |
| Ent-Pir Cx | Entorhinal and Piriform Cortex |
| EXP | 5G-exposed |
| FC | Fold-Change |
| FDR | False Discovery Rate |
| GO | Gene Ontology |
| G | Generation |
| ITI | Inter Trial Interval |
| mt | Mitochondrial |
| NOR | Novel Object Recognition |
| OF | Open Field |
| OL | Object Location |
| OXPHOS | Oxidative Phosphorylation System |
| PSD | Pseudo |
| RF | Radiofrequency |
| SAR | Specific Absorption Rate |
References
- Kwon, M.S.; Hämäläinen, H. Effects of Mobile Phone Electromagnetic Fields: Critical Evaluation of Behavioral and Neurophysiological Studies. Bioelectromagnetics 2011, 32, 253–272. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, S.N.; Jetti, R.; Kesari, K.K.; Kumar, R.S.; Nayak, S.B.; Bhat, P.G. Radiofrequency Electromagnetic Radiation-Induced Behavioral Changes and Their Possible Basis. Environ. Sci. Pollut. Res. 2019, 26, 30693–30710. [Google Scholar] [CrossRef] [PubMed]
- Sienkiewicz, Z.; Van Rongen, E. Can Low-Level Exposure to Radiofrequency Fields Effect Cognitive Behaviour in Laboratory Animals? A Systematic Review of the Literature Related to Spatial Learning and Place Memory. Int. J. Environ. Res. Public. Health 2019, 16, 1607. [Google Scholar] [CrossRef] [PubMed]
- Dubreuil, D.; Jay, T.; Edeline, J.-M. Does Head-Only Exposure to GSM-900 Electromagnetic Fields Affect the Performance of Rats in Spatial Learning Tasks? Behav. Brain Res. 2002, 129, 203–210. [Google Scholar] [CrossRef]
- Dubreuil, D.; Jay, T.; Edeline, J.-M. Head-Only Exposure to GSM 900-MHz Electromagnetic Fields Does Not Alter Rat’s Memory in Spatial and Non-Spatial Tasks. Behav. Brain Res. 2003, 145, 51–61. [Google Scholar] [CrossRef]
- Kumlin, T.; Iivonen, H.; Miettinen, P.; Juvonen, A.; Van Groen, T.; Puranen, L.; Pitkäaho, R.; Juutilainen, J.; Tanila, H. Mobile Phone Radiation and the Developing Brain: Behavioral and Morphological Effects in Juvenile Rats. Radiat. Res. 2007, 168, 471–479. [Google Scholar] [CrossRef]
- Abtin, S.; Seyedaghamiri, F.; Aalidaeijavadi, Z.; Farrokhi, A.M.; Moshrefi, F.; Ziveh, T.; Zibaii, M.I.; Aliakbarian, H.; Rezaei-Tavirani, M.; Haghparast, A. A Review on the Consequences of Molecular and Genomic Alterations Following Exposure to Electromagnetic Fields: Remodeling of Neuronal Network and Cognitive Changes. Brain Res. Bull. 2024, 217, 111090. [Google Scholar] [CrossRef]
- Jamal, L.; Yahia-Cherif, L.; Hugueville, L.; Mazet, P.; Lévêque, P.; Selmaoui, B. Assessment of Electrical Brain Activity of Healthy Volunteers Exposed to 3.5 GHz of 5G Signals within Environmental Levels: A Controlled–Randomised Study. Int. J. Environ. Res. Public. Health 2023, 20, 6793. [Google Scholar] [CrossRef]
- Lee, A.-K.; Choi, H.-D. Dosimetric Assessment in the Brain for Downlink EMF Exposure in Korean Mobile Communication Networks. Environ. Res. 2023, 234, 116542. [Google Scholar] [CrossRef]
- Canovi, A.; Orlacchio, R.; Poulletier De Gannes, F.; Lévêque, P.; Arnaud-Cormos, D.; Lagroye, I.; Garenne, A.; Percherancier, Y.; Lewis, N. In Vitro Exposure of Neuronal Networks to the 5G-3.5 GHz Signal. Front. Public Health 2023, 11, 1231360. [Google Scholar] [CrossRef]
- Yang, H.; Zhang, Y.; Wu, X.; Gan, P.; Luo, X.; Zhong, S.; Zuo, W. Effects of Acute Exposure to 3500 MHz (5G) Radiofrequency Electromagnetic Radiation on Anxiety-Like Behavior and the Auditory Cortex in Guinea Pigs. Bioelectromagnetics 2022, 43, 106–118. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, H.; Zhang, Z.; Sun, B.; Tang, C.; Zhang, L.; Jiang, Z.; Ding, B.; Liao, Y.; Cai, P. Simulated Mobile Communication Frequencies (3.5 GHz) Emitted by a Signal Generator Affects the Sleep of Drosophila Melanogaster. Environ. Pollut. 2021, 283, 117087. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jiang, Z.; Zhang, L.; Zhang, Z.; Liao, Y.; Cai, P. 3.5-GHz Radiofrequency Electromagnetic Radiation Promotes the Development of Drosophila Melanogaster. Environ. Pollut. 2022, 294, 118646. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, S.; Wang, G.; Simonich, M.T.; Zhang, T.; Truong, L.; Liu, H.; Tanguay, R.L. Impacts of High Dose 3.5 GHz Cellphone Radiofrequency on Zebrafish Embryonic Development. PLoS ONE 2020, 15, e0235869. [Google Scholar] [CrossRef]
- Dasgupta, S.; Leong, C.; Simonich, M.T.; Truong, L.; Liu, H.; Tanguay, R.L. Transcriptomic and Long-Term Behavioral Deficits Associated with Developmental 3.5 GHz Radiofrequency Radiation Exposures in Zebrafish. Environ. Sci. Technol. Lett. 2022, 9, 327–332. [Google Scholar] [CrossRef]
- Torres-Ruiz, M.; Suárez, O.J.; López, V.; Marina, P.; Sanchis, A.; Liste, I.; De Alba, M.; Ramos, V. Effects of 700 and 3500 MHz 5G Radiofrequency Exposure on Developing Zebrafish Embryos. Sci. Total Environ. 2024, 915, 169475. [Google Scholar] [CrossRef]
- Vercellino, I.; Sazanov, L.A. The Assembly, Regulation and Function of the Mitochondrial Respiratory Chain. Nat. Rev. Mol. Cell Biol. 2022, 23, 141–161. [Google Scholar] [CrossRef]
- Rackham, O.; Filipovska, A. Organization and Expression of the Mammalian Mitochondrial Genome. Nat. Rev. Genet. 2022, 23, 606–623. [Google Scholar] [CrossRef]
- Dong, L.L.; Fiete, I.R. Grid Cells in Cognition: Mechanisms and Function. Annu. Rev. Neurosci. 2024, 47, 345–368. [Google Scholar] [CrossRef]
- Zhou, G.; Wang, X.; Gao, P.; Qin, T.; Guo, L.; Zhang, Z.; Huang, Z.; Lin, J.; Jing, Y.; Wang, H.; et al. Intestinal Microbiota via NLRP3 Inflammasome Dependent Neuronal Pyroptosis Mediates Anxiety-like Behaviour in Mice Exposed to 3.5 GHz Radiofrequency Radiation. Sci. Total Environ. 2024, 927, 172391. [Google Scholar] [CrossRef]
- Ssimbwa, J.; Lim, B.; Lee, J.-H.; Ko, Y.-C. A Survey on Robust Modulation Requirements for the Next Generation Personal Satellite Communications. Front. Commun. Netw. 2022, 3, 850781. [Google Scholar] [CrossRef]
- Hinrikus, H.; Koppel, T.; Lass, J.; Orru, H.; Roosipuu, P.; Bachmann, M. Possible Health Effects on the Human Brain by Various Generations of Mobile Telecommunication: A Review Based Estimation of 5G Impact. Int. J. Radiat. Biol. 2022, 98, 1210–1221. [Google Scholar] [CrossRef] [PubMed]
- Belyaev, I.Y.; Koch, C.B.; Terenius, O.; Roxström-Lindquist, K.; Malmgren, L.O.G.; Sommer, W.H.; Salford, L.G.; Persson, B.R.R. Exposure of Rat Brain to 915 MHz GSM Microwaves Induces Changes in Gene Expression but Not Double Stranded DNA Breaks or Effects on Chromatin Conformation. Bioelectromagnetics 2006, 27, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Nittby, H.; Widegren, B.; Krogh, M.; Grafström, G.; Berlin, H.; Rehn, G.; Eberhardt, J.L.; Malmgren, L.; Persson, B.R.R.; Salford, L.G. Exposure to Radiation from Global System for Mobile Communications at 1,800 MHz Significantly Changes Gene Expression in Rat Hippocampus and Cortex. Environmentalist 2008, 28, 458–465. [Google Scholar] [CrossRef]
- Paparini, A.; Rossi, P.; Gianfranceschi, G.; Brugaletta, V.; Falsaperla, R.; De Luca, P.; Romano Spica, V. No Evidence of Major Transcriptional Changes in the Brain of Mice Exposed to 1800 MHz GSM Signal. Bioelectromagnetics 2008, 29, 312–323. [Google Scholar] [CrossRef]
- McNamee, J.P.; Bellier, P.V.; Konkle, A.T.M.; Thomas, R.; Wasoontarajaroen, S.; Lemay, E.; Gajda, G.B. Analysis of Gene Expression in Mouse Brain Regions after Exposure to 1.9 GHz Radiofrequency Fields. Int. J. Radiat. Biol. 2016, 92, 338–350. [Google Scholar] [CrossRef]
- Fragopoulou, A.F.; Polyzos, A.; Papadopoulou, M.; Sansone, A.; Manta, A.K.; Balafas, E.; Kostomitsopoulos, N.; Skouroliakou, A.; Chatgilialoglu, C.; Georgakilas, A.; et al. Hippocampal Lipidome and Transcriptome Profile Alterations Triggered by Acute Exposure of Mice to GSM 1800 MH z Mobile Phone Radiation: An Exploratory Study. Brain Behav. 2018, 8, e01001. [Google Scholar] [CrossRef]
- Lameth, J.; Arnaud-Cormos, D.; Lévêque, P.; Boillée, S.; Edeline, J.-M.; Mallat, M. Effects of a Single Head Exposure to GSM-1800 MHz Signals on the Transcriptome Profile in the Rat Cerebral Cortex: Enhanced Gene Responses Under Proinflammatory Conditions. Neurotox. Res. 2020, 38, 105–123. [Google Scholar] [CrossRef]
- Berretta, N.; Jones, R.S. A Comparison of Spontaneous EPSCs in Layer II and Layer IV-V Neurons of the Rat Entorhinal Cortex in Vitro. J. Neurophysiol. 1996, 76, 1089–1100. [Google Scholar] [CrossRef]
- Basu, J.; Nagel, K. Neural Circuits for Goal-Directed Navigation across Species. Trends Neurosci. 2024, 47, 904–917. [Google Scholar] [CrossRef]
- Igarashi, K.M. Entorhinal Cortex Dysfunction in Alzheimer’s Disease. Trends Neurosci. 2023, 46, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Bontempi, B.; Lévêque, P.; Dubreuil, D.; Jay, T.M.; Edeline, J.-M. Effects of Head-Only Exposure to 900 MHz GSM Electromagnetic Fields in Rats: Changes in Neuronal Activity as Revealed by c-Fos Imaging without Concomitant Cognitive Impairments. Biomedicines 2024, 12, 1954. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Ning, W.; Xu, Z.; Zhou, S.; Chiang, H.; Luo, J. Chronic Exposure to GSM 1800-MHz Microwaves Reduces Excitatory Synaptic Activity in Cultured Hippocampal Neurons. Neurosci. Lett. 2006, 398, 253–257. [Google Scholar] [CrossRef] [PubMed]
- El Khoueiry, C.; Moretti, D.; Renom, R.; Camera, F.; Orlacchio, R.; Garenne, A.; Poulletier De Gannes, F.; Poque-Haro, E.; Lagroye, I.; Veyret, B.; et al. Decreased Spontaneous Electrical Activity in Neuronal Networks Exposed to Radiofrequency 1,800 MHz Signals. J. Neurophysiol. 2018, 120, 2719–2729. [Google Scholar] [CrossRef]
- Souffi, S.; Lameth, J.; Gaucher, Q.; Arnaud-Cormos, D.; Lévêque, P.; Edeline, J.-M.; Mallat, M. Exposure to 1800 MHz LTE Electromagnetic Fields under Proinflammatory Conditions Decreases the Response Strength and Increases the Acoustic Threshold of Auditory Cortical Neurons. Sci. Rep. 2022, 12, 4063. [Google Scholar] [CrossRef]
- Lameth, J.; Gervais, A.; Colin, C.; Lévêque, P.; Jay, T.M.; Edeline, J.-M.; Mallat, M. Acute Neuroinflammation Promotes Cell Responses to 1800 MHz GSM Electromagnetic Fields in the Rat Cerebral Cortex. Neurotox. Res. 2017, 32, 444–459. [Google Scholar] [CrossRef]
- Maussetbonnefont, A.; Hirbec, H.; Bonnefont, X.; Privat, A.; Vignon, J.; Deseze, R. Acute Exposure to GSM 900-MHz Electromagnetic Fields Induces Glial Reactivity and Biochemical Modifications in the Rat Brain. Neurobiol. Dis. 2004, 17, 445–454. [Google Scholar] [CrossRef]
- Occelli, F.; Lameth, J.; Adenis, V.; Huetz, C.; Lévêque, P.; Jay, T.M.; Edeline, J.-M.; Mallat, M. A Single Exposure to GSM-1800 MHz Signals in the Course of an Acute Neuroinflammatory Reaction Can Alter Neuronal Responses and Microglial Morphology in the Rat Primary Auditory Cortex. Neuroscience 2018, 385, 11–24. [Google Scholar] [CrossRef]
- Tan, B.G.; Gustafsson, C.M.; Falkenberg, M. Mechanisms and Regulation of Human Mitochondrial Transcription. Nat. Rev. Mol. Cell Biol. 2024, 25, 119–132. [Google Scholar] [CrossRef]
- Mustafa, M.F.; Fakurazi, S.; Abdullah, M.A.; Maniam, S. Pathogenic Mitochondria DNA Mutations: Current Detection Tools and Interventions. Genes 2020, 11, 192. [Google Scholar] [CrossRef]
- Stańczyk, M.; Szubart, N.; Maslanka, R.; Zadrag-Tecza, R. Mitochondrial Dysfunctions: Genetic and Cellular Implications Revealed by Various Model Organisms. Genes 2024, 15, 1153. [Google Scholar] [CrossRef] [PubMed]
- Monzel, A.S.; Enríquez, J.A.; Picard, M. Multifaceted Mitochondria: Moving Mitochondrial Science beyond Function and Dysfunction. Nat. Metab. 2023, 5, 546–562. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.P. How Mitochondria Produce Reactive Oxygen Species. Biochem. J. 2009, 417, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hernansanz-Agustín, P.; Enríquez, J.A. Generation of Reactive Oxygen Species by Mitochondria. Antioxidants 2021, 10, 415. [Google Scholar] [CrossRef]
- Schuermann, D.; Mevissen, M. Manmade Electromagnetic Fields and Oxidative Stress—Biological Effects and Consequences for Health. Int. J. Mol. Sci. 2021, 22, 3772. [Google Scholar] [CrossRef]
- Kesari, K.K.; Kumar, S.; Behari, J. 900-MHz Microwave Radiation Promotes Oxidation in Rat Brain. Electromagn. Biol. Med. 2011, 30, 219–234. [Google Scholar] [CrossRef]
- Megha, K.; Deshmukh, P.S.; Banerjee, B.D.; Tripathi, A.K.; Ahmed, R.; Abegaonkar, M.P. Low Intensity Microwave Radiation Induced Oxidative Stress, Inflammatory Response and DNA Damage in Rat Brain. NeuroToxicology 2015, 51, 158–165. [Google Scholar] [CrossRef]
- Hussein, S.; El-Saba, A.-A.; Galal, M.K. Biochemical and Histological Studies on Adverse Effects of Mobile Phone Radiation on Rat’s Brain. J. Chem. Neuroanat. 2016, 78, 10–19. [Google Scholar] [CrossRef]
- Sahin, D.; Ozgur, E.; Guler, G.; Tomruk, A.; Unlu, I.; Sepici-Dinçel, A.; Seyhan, N. The 2100MHz Radiofrequency Radiation of a 3G-Mobile Phone and the DNA Oxidative Damage in Brain. J. Chem. Neuroanat. 2016, 75, 94–98. [Google Scholar] [CrossRef]
- Alkis, M.E.; Bilgin, H.M.; Akpolat, V.; Dasdag, S.; Yegin, K.; Yavas, M.C.; Akdag, M.Z. Effect of 900-, 1800-, and 2100-MHz Radiofrequency Radiation on DNA and Oxidative Stress in Brain. Electromagn. Biol. Med. 2019, 38, 32–47. [Google Scholar] [CrossRef]
- Sharma, S.; Shukla, S. Effect of Electromagnetic Radiation on Redox Status, Acetylcholine Esterase Activity and Cellular Damage Contributing to the Diminution of the Brain Working Memory in Rats. J. Chem. Neuroanat. 2020, 106, 101784. [Google Scholar] [CrossRef] [PubMed]
- Bektas, H.; Algul, S.; Altindag, F.; Yegin, K.; Akdag, M.Z.; Dasdag, S. Effects of 3.5 GHz Radiofrequency Radiation on Ghrelin, Nesfatin-1, and Irisin Level in Diabetic and Healthy Brains. J. Chem. Neuroanat. 2022, 126, 102168. [Google Scholar] [CrossRef] [PubMed]
- Rana, J.N.; Mumtaz, S.; Choi, E.H.; Han, I. ROS Production in Response to High-Power Microwave Pulses Induces P53 Activation and DNA Damage in Brain Cells: Radiosensitivity and Biological Dosimetry Evaluation. Front. Cell Dev. Biol. 2023, 11, 1067861. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Fabuel, I.; Le Douce, J.; Logan, A.; James, A.M.; Bonvento, G.; Murphy, M.P.; Almeida, A.; Bolaños, J.P. Complex I Assembly into Supercomplexes Determines Differential Mitochondrial ROS Production in Neurons and Astrocytes. Proc. Natl. Acad. Sci. USA 2016, 113, 13063–13068. [Google Scholar] [CrossRef]
- Lalonde, R.; Strazielle, C. Relations between Open-Field, Elevated plus-Maze, and Emergence Tests as Displayed by C57/BL6J and BALB/c Mice. J. Neurosci. Methods 2008, 171, 48–52. [Google Scholar] [CrossRef]
- ICNIRP. Guidelines for Limiting Exposure to Electromagnetic Fields (100 kHz to 300 GHz). Health Phys. 2020, 118, 483–524. [Google Scholar] [CrossRef]
- Leveque, P.; Dale, C.; Veyret, B.; Wiart, J. Dosimetric Analysis of a 900-MHz Rat Head Exposure System. IEEE Trans. Microw. Theory Tech. 2004, 52, 2076–2083. [Google Scholar] [CrossRef]
- Watilliaux, A.; Edeline, J.-M.; Lévêque, P.; Jay, T.M.; Mallat, M. Effect of Exposure to 1,800 MHz Electromagnetic Fields on Heat Shock Proteins and Glial Cells in the Brain of Developing Rats. Neurotox. Res. 2011, 20, 109–119. [Google Scholar] [CrossRef]
- Yee, K. Numerical Solution of Initial Boundary Value Problems Involving Maxwell’s Equations in Isotropic Media. IEEE Trans. Antennas Propag. 1966, 14, 302–307. [Google Scholar] [CrossRef]
- Kunz, K.S.; Luebbers, R.J. The Finite Difference Time Domain Method for Electromagnetics; CRC Press: Boca Raton, FL, USA, 1993; ISBN 978-0-8493-8657-2. [Google Scholar]
- Taflove, A.; Hagness, S.C. Computational Electrodynamics: The Finite-Difference Time-Domain Method, 3rd ed.; Artech House Antennas and Propagation Library; [Nachdr.]; Artech House: Boston, MA, USA, 2010; ISBN 978-1-58053-832-9. [Google Scholar]
- Poirier, R.; Jacquot, S.; Vaillend, C.; Soutthiphong, A.A.; Libbey, M.; Davis, S.; Laroche, S.; Hanauer, A.; Welzl, H.; Lipp, H.-P.; et al. Deletion of the Coffin–Lowry Syndrome Gene Rsk2 in Mice Is Associated With Impaired Spatial Learning and Reduced Control of Exploratory Behavior. Behav. Genet. 2007, 37, 31–50. [Google Scholar] [CrossRef]
- Andrews, S. FastQC: A Quality Control Analysis Tool for High Throughput Sequence Data [Online]. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 21 February 2025).
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and Integrative Analysis of Large Gene Lists Using DAVID Bioinformatics Resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: A Web Server for Functional Enrichment Analysis and Functional Annotation of Gene Lists (2021 Update). Nucleic Acids Res. 2022, 50, W216–W221. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lameth, J.; Royer, J.; Martin, A.; Marie, C.; Arnaud-Cormos, D.; Lévêque, P.; Poirier, R.; Edeline, J.-M.; Mallat, M. Repeated Head Exposures to a 5G-3.5 GHz Signal Do Not Alter Behavior but Modify Intracortical Gene Expression in Adult Male Mice. Int. J. Mol. Sci. 2025, 26, 2459. https://doi.org/10.3390/ijms26062459
Lameth J, Royer J, Martin A, Marie C, Arnaud-Cormos D, Lévêque P, Poirier R, Edeline J-M, Mallat M. Repeated Head Exposures to a 5G-3.5 GHz Signal Do Not Alter Behavior but Modify Intracortical Gene Expression in Adult Male Mice. International Journal of Molecular Sciences. 2025; 26(6):2459. https://doi.org/10.3390/ijms26062459
Chicago/Turabian StyleLameth, Julie, Juliette Royer, Alexandra Martin, Corentine Marie, Délia Arnaud-Cormos, Philippe Lévêque, Roseline Poirier, Jean-Marc Edeline, and Michel Mallat. 2025. "Repeated Head Exposures to a 5G-3.5 GHz Signal Do Not Alter Behavior but Modify Intracortical Gene Expression in Adult Male Mice" International Journal of Molecular Sciences 26, no. 6: 2459. https://doi.org/10.3390/ijms26062459
APA StyleLameth, J., Royer, J., Martin, A., Marie, C., Arnaud-Cormos, D., Lévêque, P., Poirier, R., Edeline, J.-M., & Mallat, M. (2025). Repeated Head Exposures to a 5G-3.5 GHz Signal Do Not Alter Behavior but Modify Intracortical Gene Expression in Adult Male Mice. International Journal of Molecular Sciences, 26(6), 2459. https://doi.org/10.3390/ijms26062459