
Transcriptomic and Phenotypic Responses of Cucumber Trichome Density to Silver Nitrate and Sodium Thiosulfate Application

 , ,
, ,  and
and
Abstract
1. Introduction
2. Results
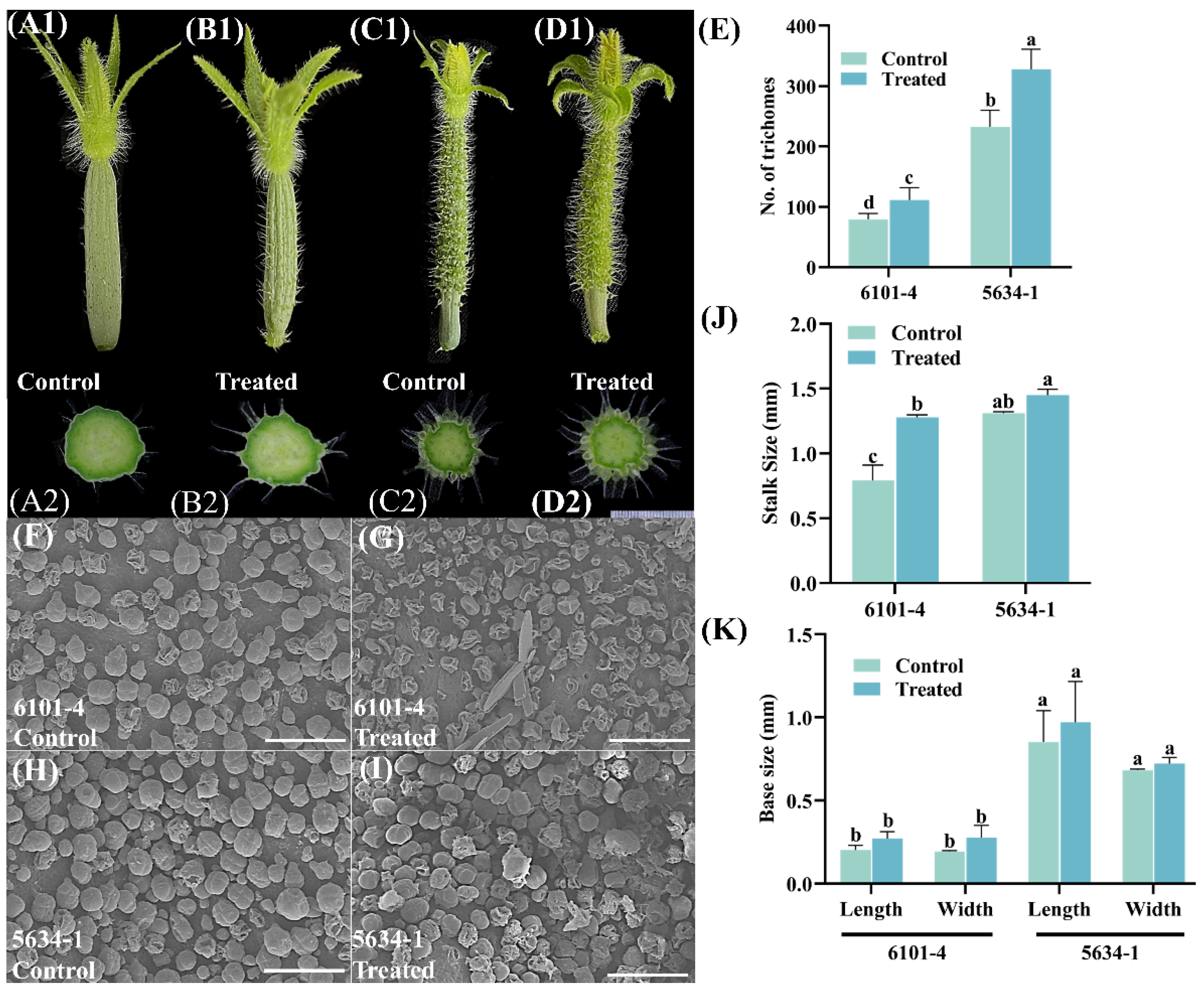
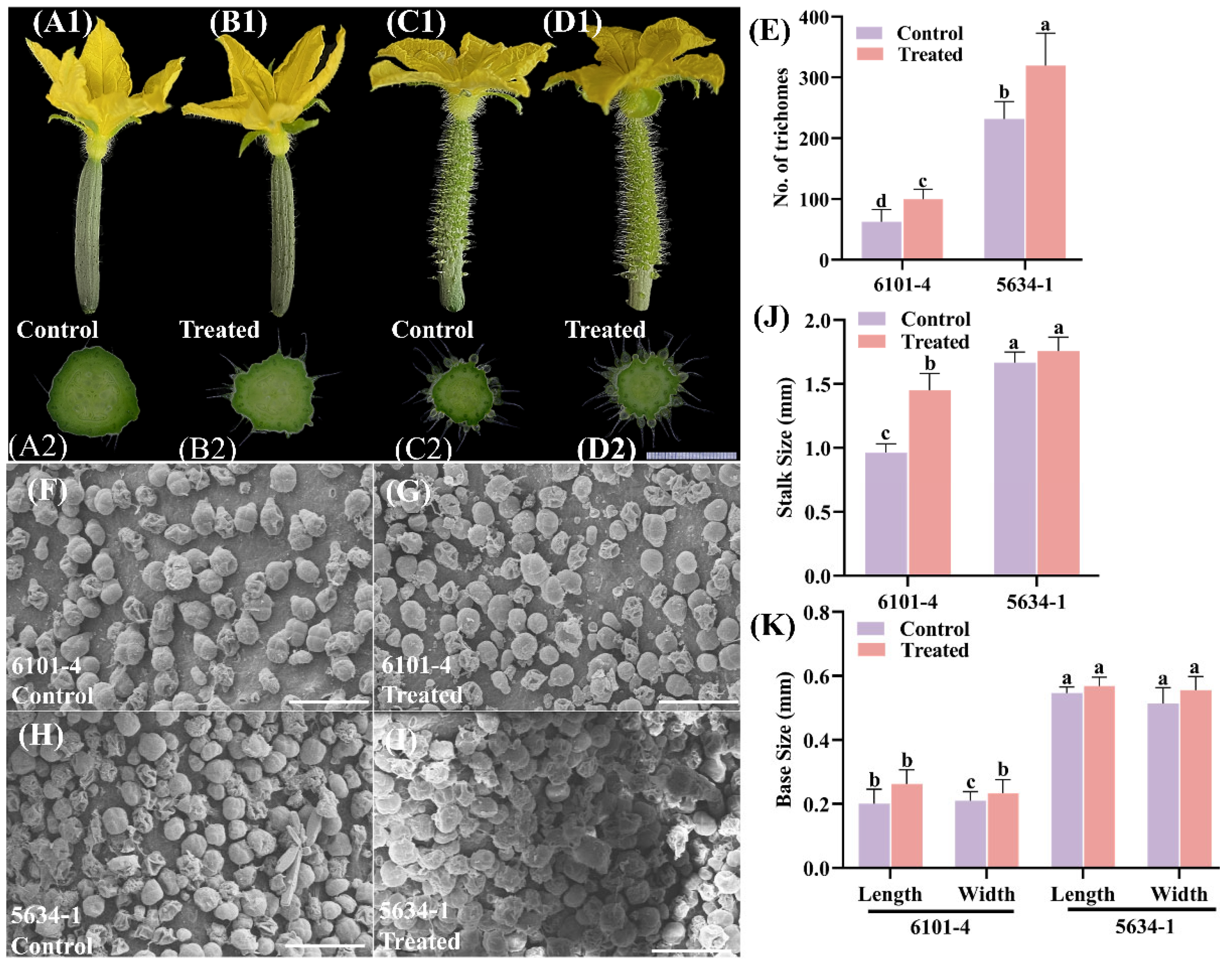
2.1. Trichome Density and Morphological Variations in Cucumber Genotypes at 5 DBF Stage
2.2. Trichome Density and Morphology of Immature Fruits at Flowering (DF)
2.3. Trichome Density and Morphology of Fruits 5 Days After Flowering (5 DAF)
2.4. Trichome Scanning Electron Microscopy (SEM) Analysis
2.5. Overview of RNA Sequencing
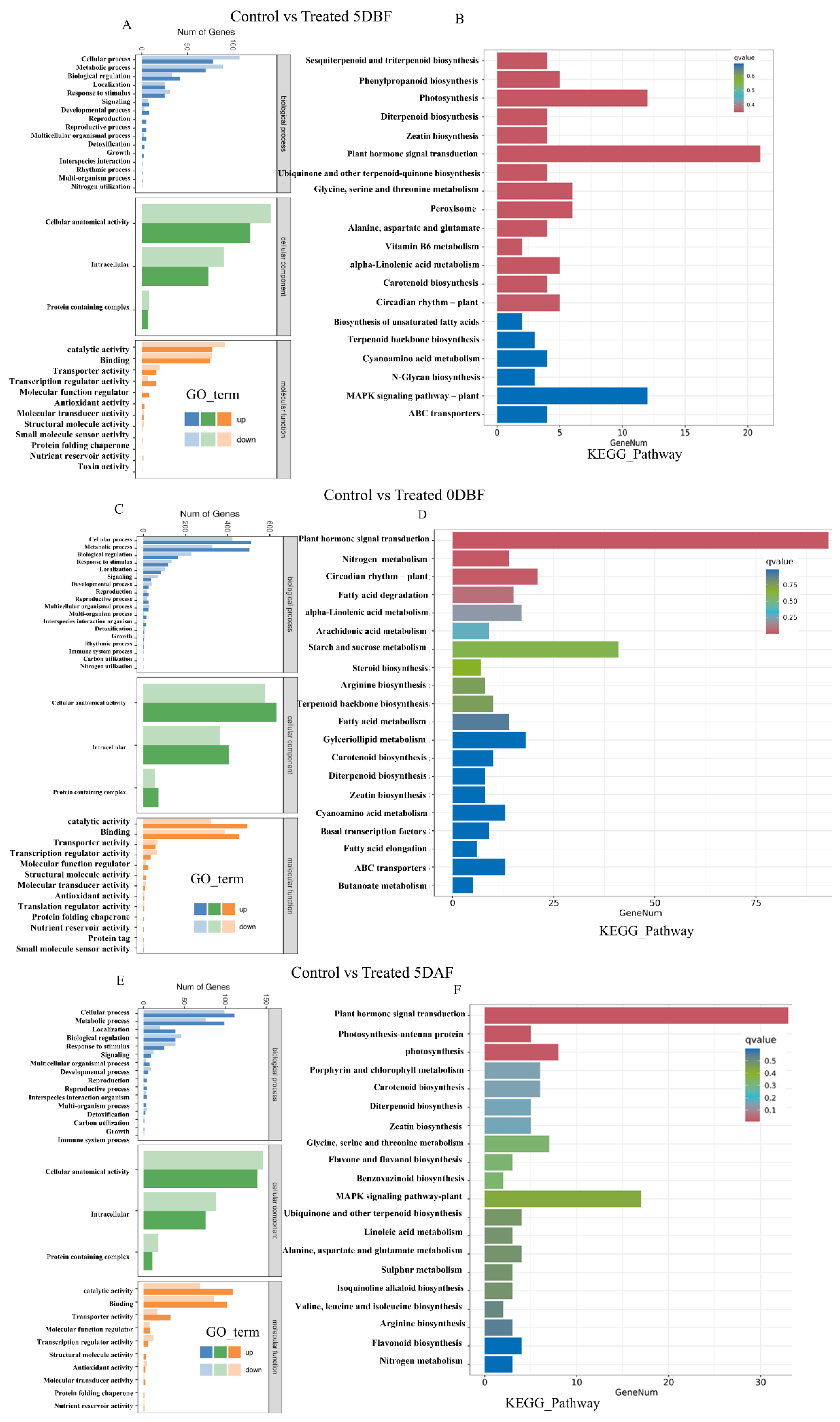
2.6. Differentially Expressed Gene (DEG) Analysis
2.7. GO and KEGG Enrichment Analysis
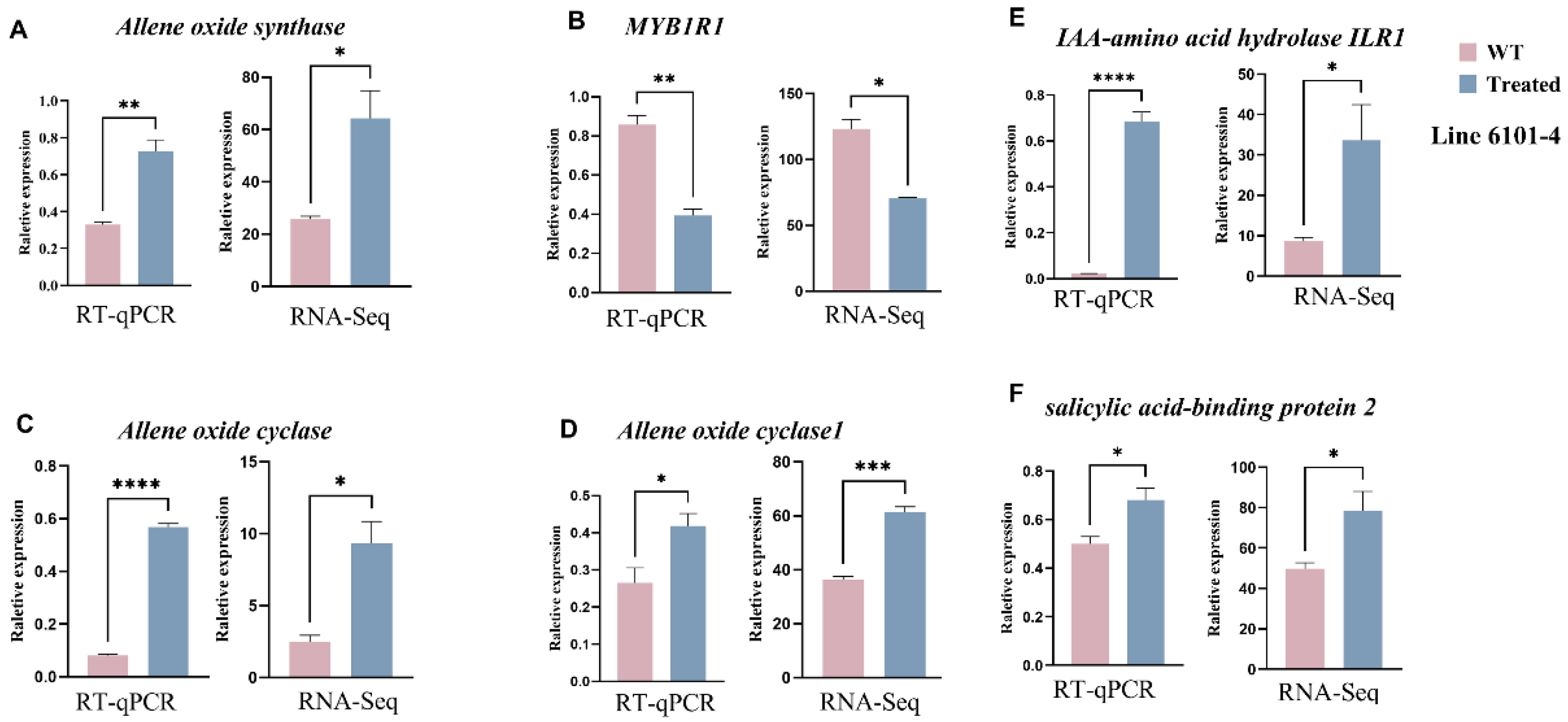
2.8. Gene Expression Analysis by RT-PCR
3. Discussion
4. Materials and Methods
4.1. Selection and Cultivation of Two Inbred Lines
4.2. Phenotypic and Scanning Electron Microscopy Data Collection
4.3. Foliar Application of Sodium Thiosulphate (Na2S2O3) and Silver Nitrate (AgNO3)
4.4. Sample Collection for Sequencing
4.5. RNA Extraction and Library Preparation
4.6. Analysis of Transcriptome Sequencing
4.7. Gene Ontology and KEGG Pathway Analysis
4.8. Quantitative Real-Time PCR (qRT-PCR) for Data Validation
4.9. Data Analysis and Interpretation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Renner, S.S.; Schaefer, H.; Grumet, R.; Katzir, N.; Garcia-Mas, J. Genetics and Genomics of Cucurbitaceae; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Chomicki, G.; Schaefer, H.; Renner, S.S. Origin and Domestication of Cucurbitaceae Crops: Insights from Phylogenies, Genomics and Archaeology. New Phytol. 2020, 226, 1240–1255. [Google Scholar] [CrossRef] [PubMed]
- Qing, Z.; Shi, Y.; Han, L.; Li, P.; Zha, Z.; Liu, C.; Liu, X.; Huang, P.; Liu, Y.; Tang, Q. Identification of Seven Undescribed Cucurbitacins in Cucumis sativus (Cucumber) and Their Cytotoxic Activity. Phytochemistry 2022, 197, 113123. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Zhang, H.; Zhong, Y.; Jiang, N.; Zhong, X.; Zhang, Q.; Chai, S.; Li, H.; Zhang, Z. Comparative Genomics Analysis of BHLH Genes in Cucurbits Identifies a Novel Gene Regulating Cucurbitacin Biosynthesis. Hortic. Res. 2022, 9, uhac038. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, L.; Liu, L.; Guo, Y.; Ren, H. Alleviating Effect of Exogenous Nitric Oxide in Cucumber Seedling against Chilling Stress. African J. Biotechnol. 2011, 10, 4380–4386. [Google Scholar]
- Chen, C.; Liu, M.; Jiang, L.; Liu, X.; Zhao, J.; Yan, S.; Yang, S.; Ren, H.; Liu, R.; Zhang, X. Transcriptome Profiling Reveals Roles of Meristem Regulators and Polarity Genes during Fruit Trichome Development in Cucumber (Cucumis Sativus L.). J. Exp. Bot. 2014, 65, 4943–4958. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Hayashi, M.; Kanamaru, T.; Watanabe, T.; Mametsuka, S.; Tanaka, Y. Studies on Bloom on the Surface of Cucumber [Cucumis sativus] Fruits, 2: Relation between the Degree of Bloom Occurrence and Contents of Mineral Elements. In Bulletin of the Fukuoka Agricultural Research Center. Series B Horticulture; Fukuoka Agricultural Research Center: Chikushino, Japan, 1989. [Google Scholar]
- Li, Y.; Wen, C.; Weng, Y. Fine Mapping of the Pleiotropic Locus B for Black Spine and Orange Mature Fruit Color in Cucumber Identifies a 50 Kb Region Containing a R2R3-MYB Transcription Factor. Theor. Appl. Genet. 2013, 126, 2187–2196. [Google Scholar] [CrossRef]
- Wagner, G.J. Secreting Glandular Trichomes: More than Just Hairs. Plant Physiol. 1991, 96, 675–679. [Google Scholar] [CrossRef]
- Werker, E. Trichome Diversity and Development. Advances in Botanical Research. Plant Trichomes; Academic Press: Cambridge, MA, USA, 2000. [Google Scholar]
- Eisner, T.; Eisner, M.; Hoebeke, E.R. When Defense Backfires: Detrimental Effect of a Plant’s Protective Trichomes on an Insect Beneficial to the Plant. Proc. Natl. Acad. Sci. USA 1998, 95, 4410–4414. [Google Scholar] [CrossRef]
- Riddick, E.W.; Simmons, A.M. Do Plant Trichomes Cause More Harm than Good to Predatory Insects? Pest Manag. Sci. 2014, 70, 1655–1665. [Google Scholar] [CrossRef]
- Glas, J.J.; Schimmel, B.C.J.; Alba, J.M.; Escobar-Bravo, R.; Schuurink, R.C.; Kant, M.R. Plant Glandular Trichomes as Targets for Breeding or Engineering of Resistance to Herbivores. Int. J. Mol. Sci. 2012, 13, 17077–17103. [Google Scholar] [CrossRef]
- Lange, B.M.; Ahkami, A. Metabolic Engineering of Plant Monoterpenes, Sesquiterpenes and Diterpenes—Current Status and Future Opportunities. Plant Biotechnol. J. 2013, 11, 169–196. [Google Scholar] [CrossRef] [PubMed]
- Loughner, R.; Goldman, K.; Loeb, G.; Nyrop, J. Influence of Leaf Trichomes on Predatory Mite (Typhlodromus pyri) Abundance in Grape Varieties. Exp. Appl. Acarol. 2008, 45, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Vosman, B.; Kashaninia, A.; van’t Westende, W.; Meijer-Dekens, F.; van Eekelen, H.; Visser, R.G.F.; de Vos, R.C.H.; Voorrips, R.E. QTL Mapping of Insect Resistance Components of Solanum Galapagense. Theor. Appl. Genet. 2019, 132, 531–541. [Google Scholar] [CrossRef] [PubMed]
- Shi, P.; Fu, X.; Shen, Q.; Liu, M.; Pan, Q.; Tang, Y.; Jiang, W.; Lv, Z.; Yan, T.; Ma, Y. The Roles of Aa MIXTA 1 in Regulating the Initiation of Glandular Trichomes and Cuticle Biosynthesis in Artemisia Annua. New Phytol. 2018, 217, 261–276. [Google Scholar] [CrossRef] [PubMed]
- Tissier, A. Glandular Trichomes: What Comes after Expressed Sequence Tags? Plant J. 2012, 70, 51–68. [Google Scholar] [CrossRef]
- Arpan, H. The Researches on Effect of 2-Chloroethylophosphonic Acid (Ethrel) on Gender Appearance and Some Other Characters of Cucumber. Fac. Agric. Publ. 1974, 165, 149–153. [Google Scholar]
- Dai, D.; Wang, L.; Liu, Y.; Chu, M.; Wang, J.; Ji, P.; Sheng, Y. Screening of Key Genes Promoting Stamen Formation Induced by Silver Nitrate in Gynoecious Melon. SSRN 2022, 4019483. [Google Scholar] [CrossRef]
- Hu, W.; Su, Y.; Zhou, J.; Zhu, H.; Guo, J.; Huo, H.; Gong, H. Foliar Application of Silicon and Selenium Improves the Growth, Yield and Quality Characteristics of Cucumber in Field Conditions. Sci. Hortic. 2022, 294, 110776. [Google Scholar] [CrossRef]
- Owen, L.C. Pollen Quality, Storage, Breeding, and Cytology of Cannabis sativa L.; North Carolina State University: Raleigh, NC, USA, 2023; ISBN 9798380713276. [Google Scholar]
- Chandler, J.W. The Hormonal Regulation of Flower Development. J. Plant Growth Regul. 2011, 30, 242–254. [Google Scholar] [CrossRef]
- Beyer, E.M., Jr. A Potent Inhibitor of Ethylene Action in Plants. Plant Physiol. 1976, 58, 268–271. [Google Scholar] [CrossRef]
- Rodrıguez, F.I.; Esch, J.J.; Hall, A.E.; Binder, B.M.; Schaller, G.E.; Bleecker, A.B. A Copper Cofactor for the Ethylene Receptor ETR1 from Arabidopsis. Science 1999, 283, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.-C.; Qu, X.; Mathews, D.E.; Schaller, G.E. Effect of Ethylene Pathway Mutations upon Expression of the Ethylene Receptor ETR1 from Arabidopsis. Plant Physiol. 2002, 130, 1983–1991. [Google Scholar] [CrossRef] [PubMed]
- Binder, B.M.; Rodriguez, F.I.; Bleecker, A.B.; Patterson, S.E. The Effects of Group 11 Transition Metals, Including Gold, on Ethylene Binding to the ETR1 Receptor and Growth of Arabidopsis Thaliana. FEBS Lett. 2007, 581, 5105–5109. [Google Scholar] [CrossRef] [PubMed]
- Kahana, A.; Silberstein, L.; Kessler, N.; Goldstein, R.S.; Perl-Treves, R. Expression of ACC Oxidase Genes Differs among Sex Genotypes and Sex Phases in Cucumber. Plant Mol. Biol. 1999, 41, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Duan, Q.-H.; Wang, D.-H.; Xu, Z.-H.; Bai, S.-N. Stamen Development in Arabidopsis Is Arrested by Organ-Specific Overexpression of a Cucumber Ethylene Synthesis Gene CsACO2. Planta 2008, 228, 537–543. [Google Scholar] [CrossRef]
- Singh, G.P.; Singh, R.K. Chemical Sex Modification and Its Effect on Fruiting in Cucumber (Cucumis sativus L.). South Indian Hortic. 1984, 32, 127–131. [Google Scholar]
- Kaur, H.; Manchanda, P.; Dhall, R.K. Effect of Foliar Application of Plant Growth Regulators on Expression of Flowering Genes in Cucumber (Cucumis sativus L.). Nucleus 2024, 67, 1–11. [Google Scholar] [CrossRef]
- Karakaya, D.; PADEM, H. The Effects of Silver Nitrate Applications on the Flower Quantity of Cucumbers (Cucumis sativus L.). Not. Bot. Horti Agrobot. Cluj-Napoca 2011, 39, 139–143. [Google Scholar] [CrossRef]
- Levin, D.A. The Role of Trichomes in Plant Defense. Q. Rev. Biol. 1973, 48, 3–15. [Google Scholar] [CrossRef]
- Johnson, H.B. Plant Pubescence: An Ecological Perspective. Bot. Rev. 1975, 41, 233–258. [Google Scholar] [CrossRef]
- Crouzet, J.; Roland, J.; Peeters, E.; Trombik, T.; Ducos, E.; Nader, J.; Boutry, M. NtPDR1, a Plasma Membrane ABC Transporter from Nicotiana Tabacum, Is Involved in Diterpene Transport. Plant Mol. Biol. 2013, 82, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Li, Y.-C.; Zhou, X.-R.; Xu, X.-J.; Fu, Q.-Y.; Liu, C.-Z. Two Thymol Derivatives from the Flower Buds of Lonicera Japonica and Their Antibacterial Activity. Nat. Prod. Res. 2018, 32, 2238–2243. [Google Scholar] [CrossRef] [PubMed]
- Paudel, S.; Lin, P.-A.; Foolad, M.R.; Ali, J.G.; Rajotte, E.G.; Felton, G.W. Induced Plant Defenses against Herbivory in Cultivated and Wild Tomato. J. Chem. Ecol. 2019, 45, 693–707. [Google Scholar] [CrossRef] [PubMed]
- Kaur, I.; Kariyat, R. Trichomes Mediate Plant–Herbivore Interactions in Two Cucurbitaceae Species through Pre-and Post-Ingestive Ways. J. Pest Sci. 2023, 96, 1077–1089. [Google Scholar] [CrossRef] [PubMed]
- Watts, S.; Kariyat, R. Morphological Characterization of Trichomes Shows Enormous Variation in Shape, Density and Dimensions across the Leaves of 14 Solanum Species. AoB Plants 2021, 13, plab071. [Google Scholar] [CrossRef]
- Xue, S.; Dong, M.; Liu, X.; Xu, S.; Pang, J.; Zhang, W.; Weng, Y.; Ren, H. Classification of Fruit Trichomes in Cucumber and Effects of Plant Hormones on Type II Fruit Trichome Development. Planta 2019, 249, 407–416. [Google Scholar] [CrossRef]
- Lv, D.; Wang, G.; Zhang, Q.; Yu, Y.; Qin, P.-C.; Pang, J.-A.; Sun, J.-X.; Zhang, K.-Y.; He, H.-L.; Cai, R. Comparative Transcriptome Analysis of Hard and Tender Fruit Spines of Cucumber to Identify Genes Involved in the Morphological Development of Fruit Spines. Front. Plant Sci. 2022, 13, 797433. [Google Scholar] [CrossRef]
- Zhang, L.; Lv, D.; Pan, J.; Zhang, K.; Wen, H.; Chen, Y.; Du, H.; He, H.; Cai, R.; Pan, J. A SNP of HD-ZIP I Transcription Factor Leads to Distortion of Trichome Morphology in Cucumber (Cucumis sativus L.). BMC Plant Biol. 2021, 21, 1–13. [Google Scholar] [CrossRef]
- Kundan, M.; Gani, U.; Nautiyal, A.K.; Misra, P. Molecular Biology of Glandular Trichomes and Their Functions in Environmental Stresses. In Molecular Approaches in Plant Biology and Environmental Challenges; Springer: Singapore, 2019; pp. 365–393. [Google Scholar]
- Jain, S.; Nidhi, N.; Kale, S.; Rathod, M.; Dhurve, L.; Mehara, H.; Baidya, B.K. A Comprehensive Review on Role of Bio-Regulators in the Growth and Development of Fruit and Vegetable Crops. Int. J. Environ. Clim. Change 2023, 13, 2879–2892. [Google Scholar] [CrossRef]
- Juturu, V.N.; Mekala, G.K.; Kirti, P.B. Current Status of Tissue Culture and Genetic Transformation Research in Cotton (Gossypium Spp.). Plant Cell Tissue Organ Cult. 2015, 120, 813–839. [Google Scholar] [CrossRef]
- Spyropoulou, E.A.; Haring, M.A.; Schuurink, R.C. RNA Sequencing on Solanum Lycopersicum Trichomes Identifies Transcription Factors That Activate Terpene Synthase Promoters. BMC Genom. 2014, 15, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Zhang, Y.; Ding, Q.; Wang, H.; Meng, X.; Fan, H.; Yu, Y.; Cui, N. The SlMYC1-TOR Module Regulates Trichome Formation and Terpene Biosynthesis in Tomatoes (Solanum lycopersicum L.). J. Plant Growth Regul. 2024, 43, 3282–3294. [Google Scholar] [CrossRef]
- Váňová, L. Of Dissertation: Use of in vitro Culture for Risk Assessment of Pahs in Plants. J. Exp. Bot 2009, 51, 1861–1866. [Google Scholar]
- Chandra-kuntal, K. Ethylene and ROS Crosstalk in Plant Developmental Processes. In Ethylene in Plant Biology; Wiley: Hoboken, NJ, USA, 2022; pp. 125–177. [Google Scholar]
- Goossens, J.; Mertens, J.; Goossens, A. Role and Functioning of BHLH Transcription Factors in Jasmonate Signalling. J. Exp. Bot. 2017, 68, 1333–1347. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie Enables Improved Reconstruction of a Transcriptome from RNA-Seq Reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene Ontology Analysis for RNA-Seq: Accounting for Selection Bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef]
- Mao, X.; Cai, T.; Olyarchuk, J.G.; Wei, L. Automated Genome Annotation and Pathway Identification Using the KEGG Orthology (KO) as a Controlled Vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef]
- Wan, H.; Zhao, Z.; Qian, C.; Sui, Y.; Malik, A.A.; Chen, J. Selection of Appropriate Reference Genes for Gene Expression Studies by Quantitative Real-Time Polymerase Chain Reaction in Cucumber. Anal. Biochem. 2010, 399, 257–261. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Total Clean Reads (M) | Clean Reads Q20 (%) | Clean Reads Q30 (%) | Total Mapping (%) | Unique Mapping (%) |
|---|---|---|---|---|---|
| Treated-5 DBF-6101-4 | 59.65 | 97.96 | 94.11 | 95.97 | 91.99 |
| Treated-0 DBF-6101-4 | 43.45 | 97.53 | 93.05 | 97.16 | 94.06 |
| Treated-5 DAF-6101-4 | 57.38 | 97.81 | 93.84 | 96.93 | 94.02 |
| Contro-5 DBF-6101-4 | 49.10 | 97.73 | 93.52 | 97.61 | 94.65 |
| Contro-0 DBF-6101-4 | 42.31 | 97.73 | 93.51 | 96.44 | 92.72 |
| Contro-5 DAF-6101-4 | 43.96 | 97.73 | 93.51 | 97.46 | 94.60 |
| Treated-5 DBF-5634-1 | 45.01 | 97.59 | 93.21 | 97.07 | 93.65 |
| Treated-0 DBF-5634-1 | 42.88 | 97.71 | 93.48 | 97.30 | 94.25 |
| Treated-5 DAF-5634-1 | 47.51 | 97.76 | 93.58 | 97.12 | 94.57 |
| Contro-5 DBF-5634-1 | 43.72 | 97.61 | 93.31 | 97.16 | 85.61 |
| Contro-0 DBF-5634-1 | 46.22 | 98.04 | 94.24 | 97.35 | 94.42 |
| Contro-5 DAF-5634-1 | 41.76 | 97.77 | 93.66 | 96.96 | 93.37 |
| Cucumber Lines | Comparison | Total Unigenes Identified | Upregulated | Downregulated |
|---|---|---|---|---|
| 6101-4 | control_5 DBF vs. treated_5 DBF | 479 | 233 | 246 |
| 6101-4 | control_0 DBF vs. treated_0 DBF | 2280 | 1266 | 1014 |
| 6101-4 | control_5 DAF vs. treated_5 DAF | 536 | 270 | 266 |
| 5634-1 | control_5 DBF vs. treated_5 DBF | 132 | 53 | 79 |
| 5634-1 | control_0 DBF vs. treated_0 DBF | 346 | 168 | 178 |
| 5634-1 | control_5 DAF vs. treated_5 DAF | 946 | 513 | 433 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmad, M.; Li, S.; Shan, L.; Yang, S.; Wang, Y.; Fan, S.; An, M.; Shi, Y.; Xu, Y.; Pei, T.; et al. Transcriptomic and Phenotypic Responses of Cucumber Trichome Density to Silver Nitrate and Sodium Thiosulfate Application. Int. J. Mol. Sci. 2025, 26, 1298. https://doi.org/10.3390/ijms26031298
Ahmad M, Li S, Shan L, Yang S, Wang Y, Fan S, An M, Shi Y, Xu Y, Pei T, et al. Transcriptomic and Phenotypic Responses of Cucumber Trichome Density to Silver Nitrate and Sodium Thiosulfate Application. International Journal of Molecular Sciences. 2025; 26(3):1298. https://doi.org/10.3390/ijms26031298
Chicago/Turabian StyleAhmad, Muhammad, Sen Li, Li Shan, Songlin Yang, Yaru Wang, Shanshan Fan, Menghang An, Yingqi Shi, Yifan Xu, Tiantian Pei, and et al. 2025. "Transcriptomic and Phenotypic Responses of Cucumber Trichome Density to Silver Nitrate and Sodium Thiosulfate Application" International Journal of Molecular Sciences 26, no. 3: 1298. https://doi.org/10.3390/ijms26031298
APA StyleAhmad, M., Li, S., Shan, L., Yang, S., Wang, Y., Fan, S., An, M., Shi, Y., Xu, Y., Pei, T., Ma, X., Zhao, Y., Xue, H., Liu, X., & Ren, H. (2025). Transcriptomic and Phenotypic Responses of Cucumber Trichome Density to Silver Nitrate and Sodium Thiosulfate Application. International Journal of Molecular Sciences, 26(3), 1298. https://doi.org/10.3390/ijms26031298