
New Progress in Zebrafish Liver Tumor Models: Techniques and Applications in Hepatocellular Carcinoma Research



Abstract
1. Introduction
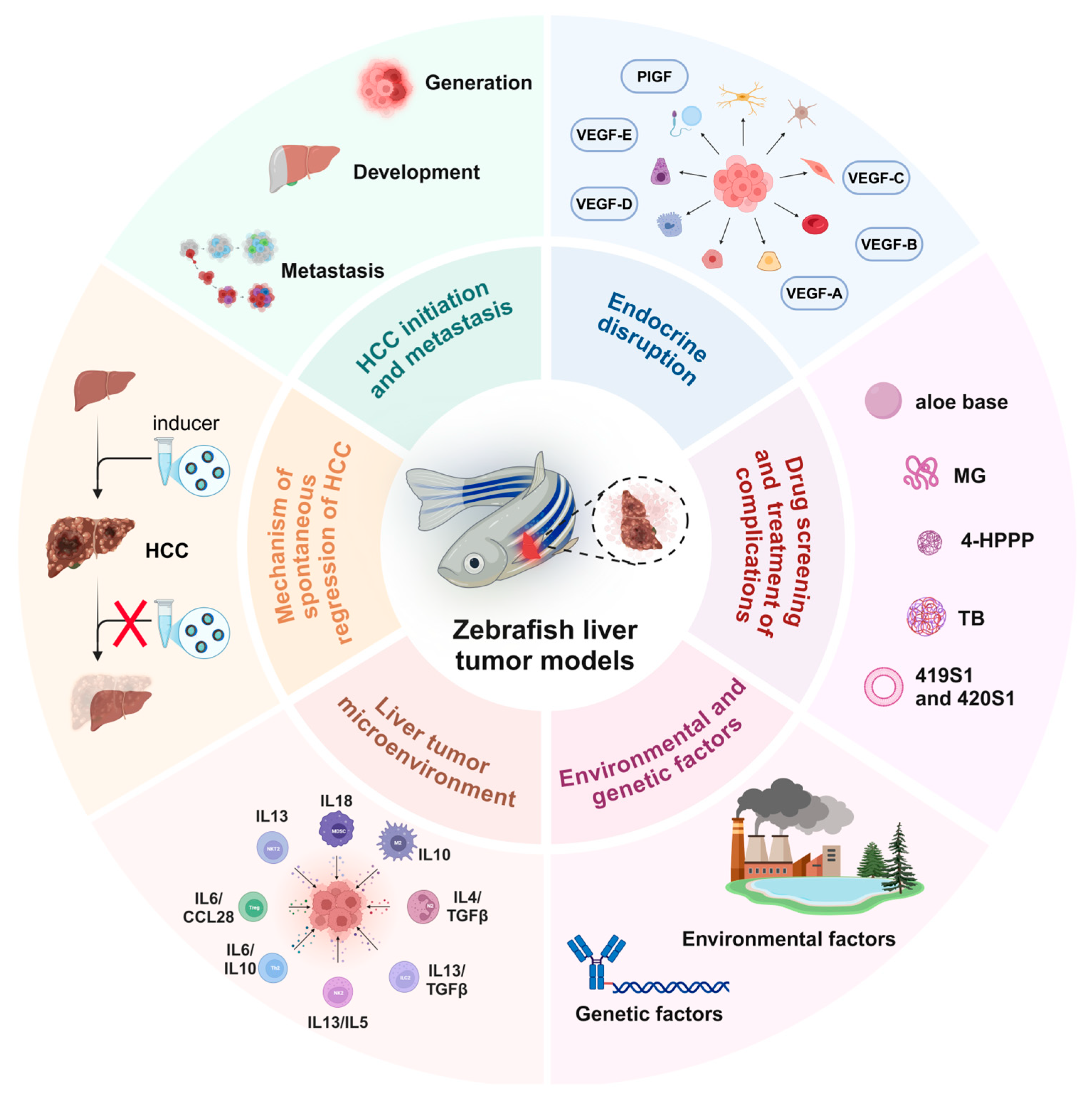
2. Application of the Zebrafish Liver Tumor Model in Different HCC Exploration Scenarios
2.1. Mechanisms of Occurrence and Metastasis of HCC
2.2. Research on the Mechanism of Spontaneous Regression in Liver Cancer
2.3. Liver Tumor Microenvironment
2.4. Drug Screening and Individualized Treatment of Complications
2.5. Interference of Sex Hormones on Liver Tumors
2.6. Research on the Impact of Genetic and Environmental Factors on Liver Cancer
3. Construction of a Zebrafish Liver Tumor Model Through Transplantation Methods
3.1. Transplantation Methods
3.2. Embryo Transfer Model
3.3. Adult Fish Transplant Model
3.4. Application Effects of Zebrafish Models Constructed Through Transplantation
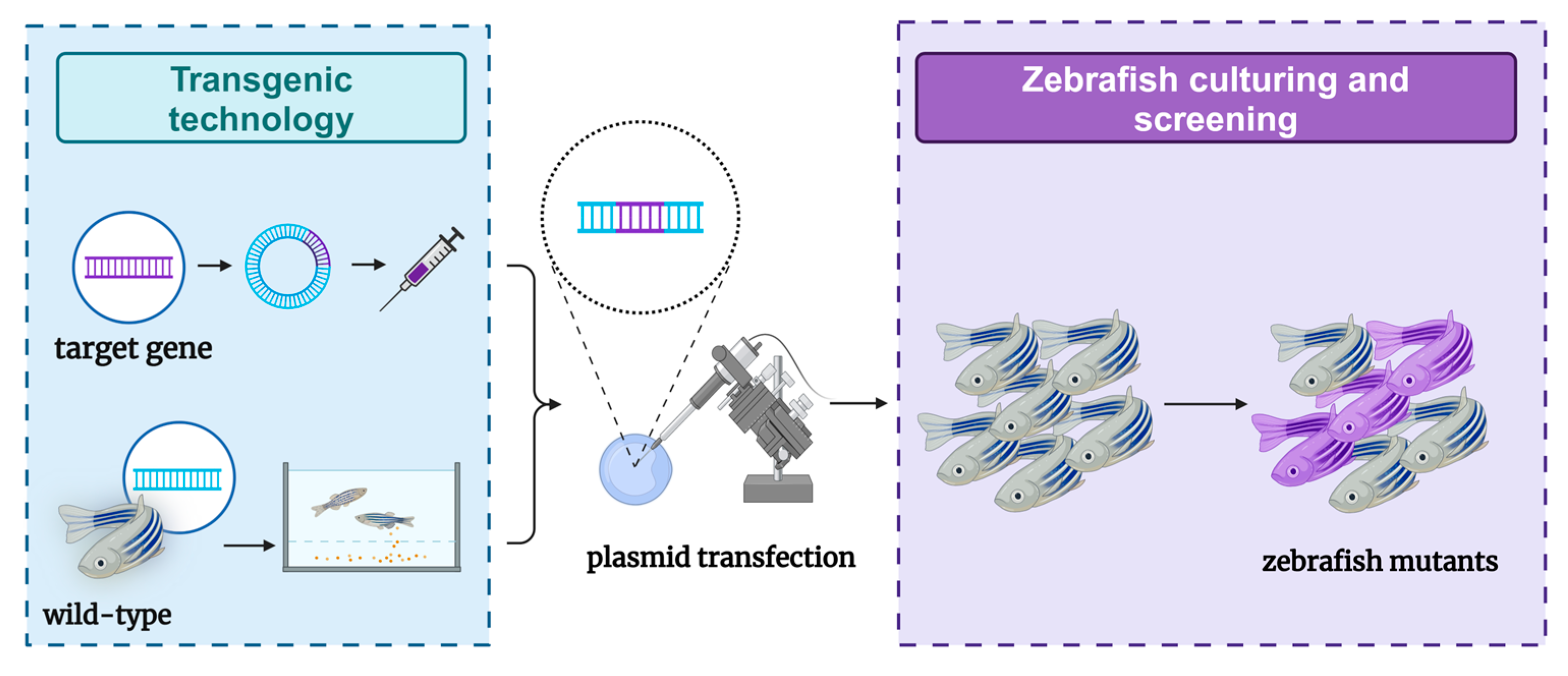
4. Transgenic Methods for Constructing a Zebrafish Liver Tumor Model
4.1. Transgenic Methods
4.2. krasV12 Gene Overexpression Model
4.3. xmrk Model
4.4. Myc Overexpression Model
4.5. Multitransgenic Model
4.6. Application Effects of Zebrafish Models Constructed Through Transgenic Methods
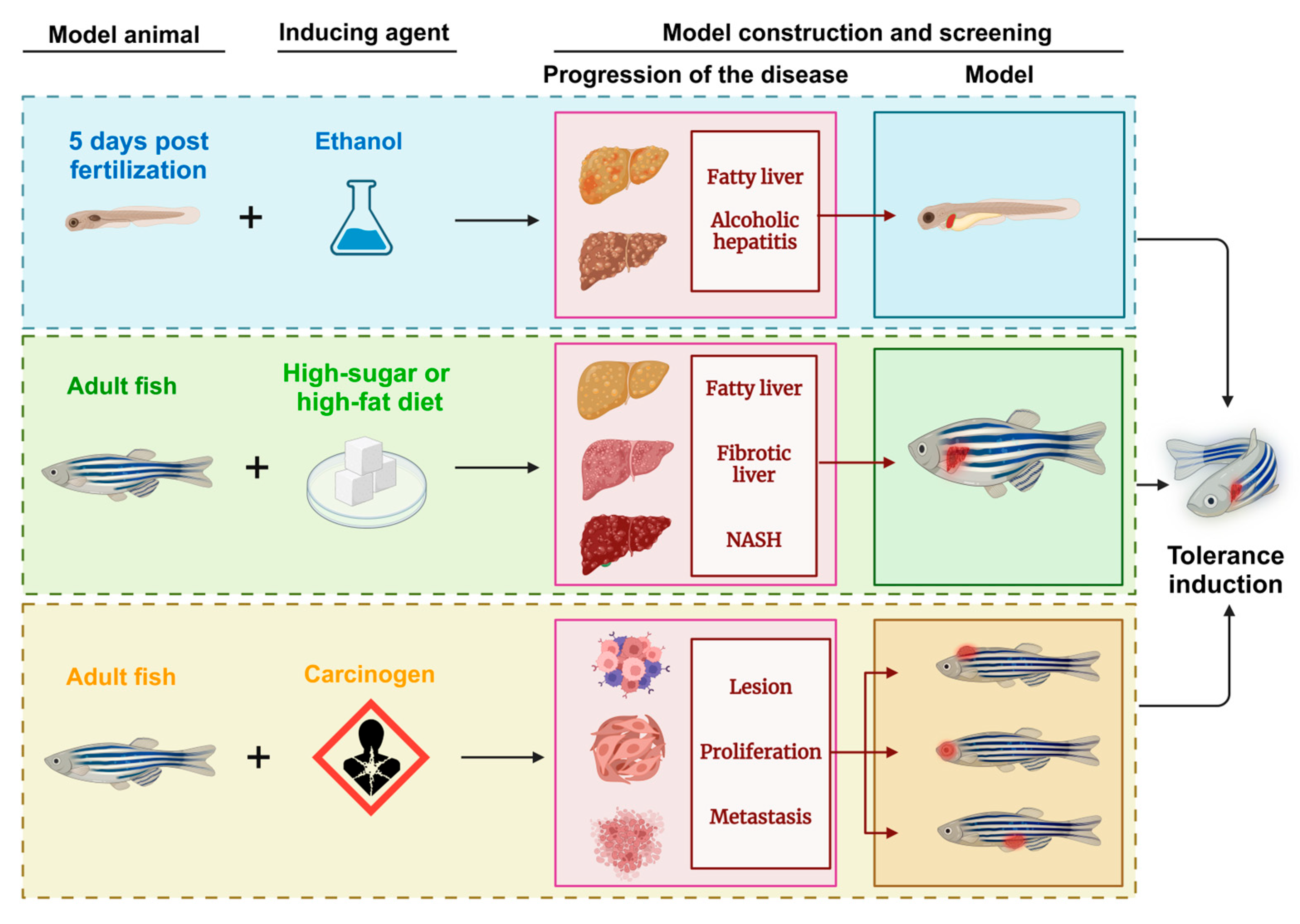
5. Construction of Zebrafish Liver Tumor Models Through Induction Methods
5.1. Induction Methods
5.2. Chemical Induction Model
5.3. Dietary Induction Model
5.4. Application Effects of Zebrafish Models Constructed Through Induction Methods
6. Gene Knockout Methods for Constructing Zebrafish Liver Tumor Models
6.1. Gene Knockout Methods
6.2. p53 Mutation Model
6.3. PTEN Knockout Model
6.4. Application Effects of Zebrafish Models Constructed Through Gene Knockout Methods
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| HCC | Hepatocellular carcinoma |
| WHO | World Health Organization |
| PET | Positron emission tomography |
| CT | Computed tomography |
| MRI | Resonance imaging |
| TERT | Telomerase reverse transcriptase |
| NAFLD | Non-alcoholic fatty liver disease |
| EMT | Epithelial-to-mesenchymal transition |
| TME | Tumor microenvironment |
| 4-HPPP | 4-[4-(4-hydroxyphenoxy)phenoxy]phenol |
| MG | Methyl gallate |
| TB | Theabrownin |
| CACS | Cancer-related anorexia cachexia syndrome |
| MFCSA | Muscle fiber cross-sectional area |
| Igf1 | Insulin-like growth factor 1 |
| KT11 | 11-ketotestosterone |
| fabp10 | Fatty acid binding protein 10 |
| EGFR | Epidermal growth factor receptor |
| hMOF | Human males absent on the first |
| IARC | International Agency for Research on Cancer |
| ADI | Aidi injection |
| SS | Sorbaria sorbifolia |
| PARP1 | Poly(adenosine diphosphate [ADP]-ribose) polymerase 1 |
| WNK1 | With-no-lysine (K)-1 |
| CP | Compound Phyllanthus urinaria L. |
| HBV | Hepatitis B virus |
| HCA | Hospital Corporation of America |
| LGDN | Low-grade dysplastic nodule |
| HGDN | High-grade dysplastic nodule |
| veHCC | Very early HCC |
| eHCC | Early HCC |
| aHCC | Advanced HCC |
| vaHCC | Very advanced HCC |
| MNNG | Methyl-N′-nitro-nitrosoguanidine |
| DEN | Diethyl nitrosamine |
| pGL-VP | Promoter-less green luciferase vector plasmid |
| ALD | Alcoholic liver disease |
| DIO | Diet-induced obesity |
| NASH | Non-alcoholic steatohepatitis |
| ZFNs | Zinc finger nucleases |
| TALENs | Transcription activator-like effector nucleases |
References
- Yang, J.D.; Hainaut, P.; Gores, G.J.; Amadou, A.; Plymoth, A.; Roberts, L.R. A global view of hepatocellular carcinoma: Trends, risk, prevention and management. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 589–604. [Google Scholar] [CrossRef]
- Tonon, F.; Farra, R.; Zennaro, C.; Pozzato, G.; Truong, N.; Parisi, S.; Rizzolio, F.; Grassi, M.; Scaggiante, B.; Zanconati, F.; et al. Xenograft Zebrafish Models for the Development of Novel Anti-Hepatocellular Carcinoma Molecules. Pharmaceuticals 2021, 14, 803. [Google Scholar] [CrossRef]
- Galvão, F.H.F.; Traldi, M.C.C.; Araújo, R.S.S.; Stefano, J.T.; D’albuquerque, L.A.C.; Oliveira, C.P. Preclinical models of liver câncer. Arq. Gastroenterol. 2023, 60, 383–392. [Google Scholar] [CrossRef]
- Adwan, H.; Georges, R.; Pervaiz, A.; Berger, M.R. Investigation of Metastasis-Related Genes: A Rat Model Mimicking Liver Metastasis of Colorectal Carcinoma. Front. Oncol. 2017, 7, 152. [Google Scholar] [CrossRef]
- Pascale, F.; Pelage, J.-P.; Wassef, M.; Ghegediban, S.H.; Saint-Maurice, J.-P.; De Baere, T.; Denys, A.; Duran, R.; Deschamps, F.; Pellerin, O.; et al. Rabbit VX2 Liver Tumor Model: A Review of Clinical, Biology, Histology, and Tumor Microenvironment Characteristics. Front. Oncol. 2022, 12, 871829. [Google Scholar] [CrossRef]
- Feng, L.; Wang, Y.; Wang, X.; An, S.; Aizimuaji, Z.; Tao, C.; Zhang, K.; Cheng, S.; Wu, J.; Xiao, T.; et al. Integrated analysis of the rhesus monkey liver transcriptome during development and human primary HCC AFP-related gene expression. Mol. Ther. Nucleic Acids 2021, 25, 406–415. [Google Scholar] [CrossRef]
- Wang, L.; Piao, Y.; Guo, F.; Wei, J.; Chen, Y.; Dai, X.; Zhang, X. Current progress of pig models for liver cancer research. Biomed. Pharmacother. 2023, 165, 115256. [Google Scholar] [CrossRef]
- Targen, S.; Konu, O. Zebrafish Xenotransplantation Models for Studying Gene Function and Drug Treatment in Hepatocellular Carcinoma. J. Gastrointest. Cancer 2021, 52, 1248–1265. [Google Scholar] [CrossRef]
- Lu, J.-W.; Ho, Y.-J.; Yang, Y.-J.; Liao, H.-A.; Ciou, S.-C.; Lin, L.-I.; Ou, D.-L. Zebrafish as a disease model for studying human hepatocellular carcinoma. World J. Gastroenterol. 2015, 21, 12042–12058. [Google Scholar] [CrossRef]
- Wrighton, P.J.; Oderberg, I.M.; Goessling, W. There Is Something Fishy About Liver Cancer: Zebrafish Models of Hepatocellular Carcinoma. Cell Mol. Gastroenterol. Hepatol. 2019, 8, 347–363. [Google Scholar] [CrossRef]
- Tao, J.; Wei, Z.; Cheng, Y.; Xu, M.; Li, Q.; Lee, S.M.-Y.; Ge, W.; Luo, K.Q.; Wang, X.; Zheng, Y. Apoptosis-Sensing Xenograft Zebrafish Tumor Model for Anticancer Evaluation of Redox-Responsive Cross-Linked Pluronic Micelles. ACS Appl. Mater. Interfaces 2022, 14, 39775–39786. [Google Scholar] [CrossRef]
- White, R.M.; Sessa, A.; Burke, C.; Bowman, T.; LeBlanc, J.; Ceol, C.; Bourque, C.; Dovey, M.; Goessling, W.; Burns, C.E.; et al. Transparent adult zebrafish as a tool for in vivo transplantation analysis. Cell Stem Cell 2008, 2, 183–189. [Google Scholar] [CrossRef]
- Maciag, M.; Wnorowski, A.; Bednarz, K.; Plazinska, A. Evaluation of β-adrenergic ligands for development of pharmacological heart failure and transparency models in zebrafish. Toxicol. Appl. Pharmacol. 2022, 434, 115812. [Google Scholar] [CrossRef]
- Shimizu, N.; Shiraishi, H.; Hanada, T. Zebrafish as a Useful Model System for Human Liver Disease. Cells 2023, 12, 2246. [Google Scholar] [CrossRef]
- Oderberg, I.M.; Goessling, W. Biliary epithelial cells are facultative liver stem cells during liver regeneration in adult zebrafish. JCI Insight 2023, 8, e163929. [Google Scholar] [CrossRef]
- Li, Y.; Lee, A.Q.; Lu, Z.; Sun, Y.; Lu, J.-W.; Ren, Z.; Zhang, N.; Liu, D.; Gong, Z. Systematic Characterization of the Disruption of Intestine during Liver Tumor Progression in the xmrk Oncogene Transgenic Zebrafish Model. Cells 2022, 11, 1810. [Google Scholar] [CrossRef]
- Lin, H.-S.; Huang, Y.-L.; Wang, Y.-R.S.; Hsiao, E.; Hsu, T.-A.; Shiao, H.-Y.; Jiaang, W.-T.; Sampurna, B.P.; Lin, K.-H.; Wu, M.-S.; et al. Identification of Novel Anti-Liver Cancer Small Molecules with Better Therapeutic Index Than Sorafenib via Zebrafish Drug Screening Platform. Cancers 2019, 11, 739. [Google Scholar] [CrossRef]
- Lin, H.-D.; Tseng, Y.-K.; Yuh, C.-H.; Chen, S.-C. Low concentrations of 4-ABP promote liver carcinogenesis in human liver cells and a zebrafish model. J. Hazard. Mater. 2022, 423, 126954. [Google Scholar] [CrossRef]
- Lu, J.-W.; Lin, L.-I.; Sun, Y.; Liu, D.; Gong, Z. Effect of Lipopolysaccharides on Liver Tumor Metastasis of twist1a/krasV12 Double Transgenic Zebrafish. Biomedicines 2022, 10, 95. [Google Scholar] [CrossRef]
- Sun, L.; Nguyen, A.T.; Spitsbergen, J.M.; Gong, Z. Myc-Induced Liver Tumors in Transgenic Zebrafish Can Regress in tp53 Null Mutation. PLoS ONE 2015, 10, e0117249. [Google Scholar] [CrossRef]
- Yang, J.; Mani, S.A.; Donaher, J.L.; Ramaswamy, S.; Itzykson, R.A.; Come, C.; Savagner, P.; Gitelman, I.; Richardson, A.; Weinberg, R.A. Twist, a Master Regulator of Morphogenesis, Plays an Essential Role in Tumor Metastasis. Cell 2004, 117, 927–939. [Google Scholar] [CrossRef]
- Piccard, H.; Muschel, R.J.; Opdenakker, G. On the dual roles and polarized phenotypes of neutrophils in tumor development and progression. Crit. Rev. Oncol. Hematol. 2012, 82, 296–309. [Google Scholar] [CrossRef]
- Lai, K.P.; Gong, Z.; Tse, W.K.F. Zebrafish as the toxicant screening model: Transgenic and omics approaches. Aquat. Toxicol. 2021, 234, 105813. [Google Scholar] [CrossRef]
- Li, L.; Xiong, Y.; Cao, W.; Chen, Z.; He, L.; Tong, M.; Zhang, L.; Wu, M. Epidermal growth factor receptor promotes high-fructose nonalcoholic fatty liver disease by inducing mitochondrial fission in zebrafish. Biochem. Biophys. Res. Commun. 2023, 652, 112–120. [Google Scholar] [CrossRef]
- Morgan, K.J.; Doggett, K.; Geng, F.; Mieruszynski, S.; Whitehead, L.; Smith, K.A.; Hogan, B.M.; Simons, C.; Baillie, G.J.; Molania, R.; et al. ahctf1 and kras mutations combine to amplify oncogenic stress and restrict liver overgrowth in a zebrafish model of hepatocellular carcinoma. eLife 2023, 12, e73407. [Google Scholar] [CrossRef]
- Noordzij, J.G.; de Bruin-Versteeg, S.; Verkaik, N.S.; Vossen, J.M.J.J.; de Groot, R.; Bernatowska, E.; Langerak, A.W.; van Gent, D.C.; van Dongen, J.J.M. The immunophenotypic and immunogenotypic B-cell differentiation arrest in bone marrow of RAG-deficient SCID patients corresponds to residual recombination activities of mutated RAG proteins. Blood 2002, 100, 2145–2152. [Google Scholar]
- Tonon, F.; Zennaro, C.; Dapas, B.; Carraro, M.; Mariotti, M.; Grassi, G. Rapid and cost-effective xenograft hepatocellular carcinoma model in Zebrafish for drug testing. Int. J. Pharm. 2016, 515, 583–591. [Google Scholar] [CrossRef]
- Xu, Z.-H.; Dang, Y.; Dong, Y.; Dong, C.-Y.; Liu, Y.; Chen, X.; Yao, Z.; Shi, J.-P. Anti-hepatocellular carcinoma activity of Sorbaria sorbifolia by regulating VEGFR and c-Met/apoptotic pathway. J. Ethnopharmacol. 2024, 324, 117758. [Google Scholar] [CrossRef]
- Xu, J.; Xiao, X.; Yan, B.; Yuan, Q.; Dong, X.; Du, Q.; Zhang, J.; Shan, L.; Ding, Z.; Zhou, L.; et al. Green tea-derived theabrownin induces cellular senescence and apoptosis of hepatocellular carcinoma through p53 signaling activation and bypassed JNK signaling suppression. Cancer Cell Int. 2022, 22, 39. [Google Scholar] [CrossRef]
- Sun, C.; Jing, W.; Xiong, G.; Ma, D.; Lin, Y.; Lv, X.; Zhao, Y.; Ma, X.; Zhu, L.; Shen, X.; et al. Inhibiting Src-mediated PARP1 tyrosine phosphorylation confers synthetic lethality to PARP1 inhibition in HCC. Cancer Lett. 2022, 526, 180–192. [Google Scholar] [CrossRef]
- Meng, X.; Noyes, M.B.; Zhu, L.J.; Lawson, N.D.; Wolfe, S.A. Targeted gene inactivation in zebrafish using engineered zinc-finger nucleases. Nat. Biotechnol. 2008, 26, 695–701. [Google Scholar] [CrossRef]
- Huang, P.; Xiao, A.; Zhou, M.; Zhu, Z.; Lin, S.; Zhang, B. Heritable gene targeting in zebrafish using customized TALENs. Nat. Biotechnol. 2011, 29, 699–700. [Google Scholar] [CrossRef]
- Cao, Y.; Chen, Q.; Liu, Y.; Jin, L.; Peng, R. Research Progress on the Construction and Application of a Diabetic Zebrafish Model. Int. J. Mol. Sci. 2023, 24, 5195. [Google Scholar] [CrossRef]
- Ramakrishna, G.; Babu, P.E.; Singh, R.; Trehanpati, N. Application of CRISPR-Cas9 based gene editing to study the pathogenesis of colon and liver cancer using organoids. Hepatol. Int. 2021, 15, 1309–1317. [Google Scholar] [CrossRef]
- Lu, J.-W.; Sun, Y.; Lin, L.-I.; Liu, D.; Gong, Z. Exacerbation of Liver Tumor Metastasis in twist1a+/xmrk+ Double Transgenic Zebrafish following Lipopolysaccharide or Dextran Sulphate Sodium Exposure. Pharmaceuticals 2021, 14, 867. [Google Scholar] [CrossRef]
- Nguyen, A.T.; Emelyanov, A.; Koh, C.H.V.; Spitsbergen, J.M.; Parinov, S.; Gong, Z. An inducible krasV12 transgenic zebrafish model for liver tumorigenesis and chemical drug screening. Dis. Model. Mech. 2012, 5, 63–72. [Google Scholar] [CrossRef]
- Iscan, E.; Ekin, U.; Yildiz, G.; Oz, O.; Keles, U.; Suner, A.; Cakan-Akdogan, G.; Ozhan, G.; Nekulova, M.; Vojtesek, B.; et al. TAp73β Can Promote Hepatocellular Carcinoma Dedifferentiation. Cancers 2021, 13, 783. [Google Scholar] [CrossRef]
- Passeri, M.J.; Cinaroglu, A.; Gao, C.; Sadler, K.C. Hepatic Steatosis in Response to Acute Alcohol Exposure in Zebrafish requires Srebp Activation. Hepatology 2009, 49, 443–452. [Google Scholar] [CrossRef]
- Yan, C.; Huo, X.; Wang, S.; Feng, Y.; Gong, Z. Stimulation of hepatocarcinogenesis by neutrophils upon induction of oncogenic kras expression in transgenic zebrafish. J. Hepatol. 2015, 63, 420–428. [Google Scholar] [CrossRef]
- de Oliveira, S.; Houseright, R.A.; Graves, A.L.; Golenberg, N.; Korte, B.G.; Miskolci, V.; Huttenlocher, A. Metformin modulates innate immune inflammation and early progression of NAFLD-associated hepatocellular carcinoma in zebrafish. J. Hepatol. 2019, 70, 710–721. [Google Scholar] [CrossRef]
- Yan, C.; Yang, Q.; Gong, Z. Tumor-Associated Neutrophils and Macrophages Promote Gender Disparity in Hepatocellular Carcinoma in Zebrafish. Cancer Res. 2017, 77, 1395–1407. [Google Scholar] [CrossRef]
- Rosa, M.E.; Rebouças, L.M.; Marques, S.P.; Silva, L.M.; Cunha, F.E.; Costa, P.M.; de Assis, D.A.; Silveira, K.B.; Muniz, C.R.; Trevisan, M.T.; et al. Sodium hyaluronate microcapsules to promote antitumor selectivity of anacardic acid. Int. J. Biol. Macromol. 2025, 296, 139616. [Google Scholar] [CrossRef]
- Li, Z.; Huang, X.; Zhan, H.; Zeng, Z.; Li, C.; Spitsbergen, J.M.; Meierjohann, S.; Schartl, M.; Gong, Z. Inducible and repressable oncogene-addicted hepatocellular carcinoma in Tet-on xmrk transgenic zebrafish. J. Hepatol. 2012, 56, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Elabd, S.; Jabeen, N.A.; Gerber, V.; Peravali, R.; Bourdon, J.-C.; Kancherla, S.; Vallone, D.; Blattner, C. Delay in development and behavioural abnormalities in the absence of p53 in zebrafish. PLoS ONE 2019, 14, e0220069. [Google Scholar] [CrossRef] [PubMed]
- Hollander, M.C.; Blumenthal, G.M.; Dennis, P.A. PTEN loss in the continuum of common cancers, rare syndromes and mouse models. Nat. Rev. Cancer 2011, 11, 289–301. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Lu, C.; Feng, M.; Dai, L.; Wang, M.; Qiu, Y.; Zheng, H.; Liu, Y.; Li, L.; Tang, B.; et al. Cooperation between liver-specific mutations of pten and tp53 genetically induces hepatocarcinogenesis in zebrafish. J. Exp. Clin. Cancer Res. CR 2021, 40, 262. [Google Scholar] [CrossRef]
- Zhou, X.; Wu, D.; Mi, T.; Li, R.; Guo, T.; Li, W. Icaritin activates p53 and inhibits aerobic glycolysis in liver cancer cells. Chem. Biol. Interact. 2024, 392, 110926. [Google Scholar] [CrossRef]
- Hill, R.J.; Bona, N.; Smink, J.; Webb, H.K.; Crisp, A.; Garaycoechea, J.I.; Crossan, G.P. p53 regulates diverse tissue-specific outcomes to endogenous DNA damage in mice. Nat. Commun. 2024, 15, 2518. [Google Scholar] [CrossRef]
- Lu, J.-W.; Yang, W.-Y.; Tsai, S.-M.; Lin, Y.-M.; Chang, P.-H.; Chen, J.-R.; Wang, H.-D.; Wu, J.-L.; Jin, S.-L.C.; Yuh, C.-H. Liver-specific expressions of HBx and src in the p53 mutant trigger hepatocarcinogenesis in zebrafish. PLoS ONE 2013, 8, e76951. [Google Scholar] [CrossRef]
- Li, Y.; Li, H.; Spitsbergen, J.M.; Gong, Z. Males develop faster and more severe hepatocellular carcinoma than females in krasV12 transgenic zebrafish. Sci. Rep. 2017, 7, 41280. [Google Scholar] [CrossRef]
- Fei, F.; Sun, S.; Li, Q.; Pei, Z.; Wang, L.; Zhang, R.; Luo, F.; Yu, M.; Wang, X. Combinatorial Normalization of Liver-Derived Cytokine Pathways Alleviates Hepatic Tumor-Associated Cachexia in Zebrafish. Cancer Res. 2021, 81, 873–884. [Google Scholar] [CrossRef] [PubMed]
- Jessy, T. Immunity over inability: The spontaneous regression of cancer. J. Nat. Sci. Biol. Med. 2011, 2, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Luo, H.; Li, C.; Huo, X.; Yan, C.; Huang, X.; Al-Haddawi, M.; Mathavan, S.; Gong, Z. Transcriptomic analysis of a transgenic zebrafish hepatocellular carcinoma model reveals a prominent role of immune responses in tumour progression and regression. Int. J. Cancer 2014, 135, 1564–1573. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-Limón, G.; Zappia, J.; Muller, M. A realistic mixture of ubiquitous persistent organic pollutants affects bone and cartilage development in zebrafish by interaction with nuclear receptor signaling. PLoS ONE 2024, 19, e0298956. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yang, Z.; Liu, Y.; Sun, P.; Wu, B.; Chen, L. Combined effects of polyvinyl chloride or polypropylene microplastics with cadmium on the intestine of zebrafish at environmentally relevant concentrations. Sci. Total Environ. 2024, 954, 176289. [Google Scholar] [CrossRef]
- Tank, M.R.; Patel, H.B.; Patel, H.R.; Delvadiya, R.S.; Patel, U.D.; Fefar, D.T.; Chauhan, J.M. Long-term exposure to bisphenol-A causes oxidative stress-related alterations at the genetic and cellular levels in the mature ovary of adult zebrafish. Drug Chem. Toxicol. 2024, 1–11. [Google Scholar] [CrossRef]
- Yang, H.; Kong, L.; Chen, Z.; Wu, J. Effect of functional groups of polystyrene nanoplastics on the neurodevelopmental toxicity of acrylamide in the early life stage of zebrafish. Aquat. Toxicol. 2025, 278, 107177. [Google Scholar] [CrossRef]
- Wu, Z.; Zhang, H.; Li, R.; Sun, N.; Liu, Z.; He, F.; Yang, B. Progress in the application of zebra fish model in cancer research. China Med. Her. 2023, 15, 45–48+60. [Google Scholar]
- Lee, L.M.J.; Seftor, E.A.; Bonde, G.; Cornell, R.A.; Hendrix, M.J.C. The fate of human malignant melanoma cells transplanted into zebrafish embryos: Assessment of migration and cell division in the absence of tumor formation. Dev. Dyn. 2005, 233, 1560–1570. [Google Scholar] [CrossRef]
- Yang, J.; Pei, H.; Luo, H.; Fu, A.; Yang, H.; Hu, J.; Zhao, C.; Chai, L.; Chen, X.; Shao, X.; et al. Non-toxic dose of liposomal honokiol suppresses metastasis of hepatocellular carcinoma through destabilizing EGFR and inhibiting the downstream pathways. Oncotarget 2016, 8, 915–932. [Google Scholar] [CrossRef]
- Lam, S.H.; Chua, H.L.; Gong, Z.; Lam, T.J.; Sin, Y.M. Development and maturation of the immune system in zebrafish, Danio rerio: A gene expression profiling, in situ hybridization and immunological study. Dev. Comp. Immunol. 2004, 28, 9–28. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Huang, J.; Zhang, J.; Wu, C.; Huang, Z.; Tao, X.; You, L.; Stalin, A.; Chen, M.; Li, J.; et al. The anti-hepatocellular carcinoma effect of Aidi injection was related to the synergistic action of cantharidin, formononetin, and isofraxidin through BIRC5, FEN1, and EGFR. J. Ethnopharmacol. 2024, 319, 117209. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Yin, Z.; Wu, Z.; Sheng, Z.; Ma, C.; Chen, R.; Zhang, X.; Tang, K.; Fei, J.; Cao, Z. Probing Synergistic Targets by Natural Compounds for Hepatocellular Carcinoma. Front. Cell Dev. Biol. 2021, 9, 715762. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Yang, B.; Yao, Y.; Liao, M.; Zhang, Y.; Zeng, Y.; Zhang, F.; Wang, N.; Tong, G. Autophagic Inhibition of Caveolin-1 by Compound Phyllanthus urinaria L. Activates Ubiquitination and Proteasome Degradation of β-catenin to Suppress Metastasis of Hepatitis B-Associated Hepatocellular Carcinoma. Front. Pharmacol. 2021, 12, 659325. [Google Scholar] [CrossRef] [PubMed]
- Akbari, S.; Kunter, I.; Azbazdar, Y.; Ozhan, G.; Atabey, N.; Karagonlar, Z.F.; Erdal, E. LGR5/R-Spo1/Wnt3a axis promotes stemness and aggressive phenotype in hepatoblast-like hepatocellular carcinoma cell lines. Cell. Signal. 2021, 82, 109972. [Google Scholar] [CrossRef]
- Stuart, G.W.; V1Elkind, J.R.; Mcmurray, J.V.; Westerfield, M. Stable lines of transgenic zebrafish exhibit reproducible patterns of transgene expression. Development 1990, 109, 577–584. [Google Scholar] [CrossRef]
- Iribarne, M. Editorial: Advances in morphogenesis and patterning: Zebrafish as a model organism. Front. Cell Dev. Biol. 2024, 12, 1376663. [Google Scholar] [CrossRef]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The zebrafish reference genome sequence and its relationship to the human genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef]
- Nguyen, A.T.; Emelyanov, A.; Koh, C.H.V.; Spitsbergen, J.M.; Lam, S.H.; Mathavan, S.; Parinov, S.; Gong, Z. A high level of liver-specific expression of oncogenic Kras(V12) drives robust liver tumorigenesis in transgenic zebrafish. Dis. Model. Mech. 2011, 4, 801–813. [Google Scholar] [CrossRef]
- Zheng, W.; Li, Z.; Nguyen, A.T.; Li, C.; Emelyanov, A.; Gong, Z. Xmrk, Kras and Myc Transgenic Zebrafish Liver Cancer Models Share Molecular Signatures with Subsets of Human Hepatocellular Carcinoma. PLoS ONE 2014, 9, e91179. [Google Scholar] [CrossRef]
- Li, Z.; Zheng, W.; Wang, Z.; Zeng, Z.; Zhan, H.; Li, C.; Zhou, L.; Yan, C.; Spitsbergen, J.M.; Gong, Z. A transgenic zebrafish liver tumor model with inducible Myc expression reveals conserved Myc signatures with mammalian liver tumors. Dis. Model. Mech. 2013, 6, 414–423. [Google Scholar] [CrossRef] [PubMed]
- Kaposi-Novak, P.; Libbrecht, L.; Woo, H.G.; Lee, Y.-H.; Sears, N.C.; Coulouarn, C.; Conner, E.A.; Factor, V.M.; Roskams, T.; Thorgeirsson, S.S. Central role of c-Myc during malignant conversion in human hepatocarcinogenesis. Cancer Res. 2009, 69, 2775–2782. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.; Arvanitis, C.; Chu, K.; Dewey, W.; Leonhardt, E.; Trinh, M.; Sundberg, C.D.; Bishop, J.M.; Felsher, D.W. Sustained loss of a neoplastic phenotype by brief inactivation of MYC. Science 2002, 297, 102–104. [Google Scholar] [CrossRef] [PubMed]
- Monroe, J.D.; Fraher, D.; Huang, X.; Mellett, N.A.; Meikle, P.J.; Sinclair, A.J.; Lirette, S.T.; Maihle, N.J.; Gong, Z.; Gibert, Y. Identification of novel lipid biomarkers in xmrk- and Myc-induced models of hepatocellular carcinoma in zebrafish. Cancer Metab. 2022, 10, 7. [Google Scholar] [CrossRef]
- Miyoshi, Y.; Iwao, K.; Nagasawa, Y.; Aihara, T.; Sasaki, Y.; Imaoka, S.; Murata, M.; Shimano, T.; Nakamura, Y. Activation of the beta-catenin gene in primary hepatocellular carcinomas by somatic alterations involving exon 3. Cancer Res. 1998, 58, 2524–2527. [Google Scholar]
- Hsiao, H.-Y.; Hsu, P.-J.; Sampurna, B.P.; Lin, Y.-J.; Lin, K.-H.; Zhao, Y.-N.; Wang, H.-D.; Yuh, C.-H. The Comprehensive Effects of Carassius auratus Complex Formula against Lipid Accumulation, Hepatocarcinogenesis, and COVID-19 Pathogenesis via Stabilized G-Quadruplex and Reduced Cell Senescence. Adv. Biol. 2023, 7, 2200310. [Google Scholar] [CrossRef]
- Wu, S.; Yang, W.; Cheng, C.; Lin, K.; Sampurna, B.P.; Chan, S.; Yuh, C. Low molecular weight fucoidan inhibits hepatocarcinogenesis and nonalcoholic fatty liver disease in zebrafish via ASGR/STAT3/HNF4A signaling. Clin. Transl. Med. 2017, 10, e252. [Google Scholar] [CrossRef]
- Li, H.; Lu, J.-W.; Huo, X.; Li, Y.; Li, Z.; Gong, Z. Effects of sex hormones on liver tumor progression and regression in Myc/xmrk double oncogene transgenic zebrafish. Gen. Comp. Endocrinol. 2019, 277, 112–121. [Google Scholar] [CrossRef]
- Zhang, R.; Yuan, J.; Liu, S.; Torraca, V.; Liao, Z.; Wu, Y.; Tan, H.; Yao, X.; Hou, X.; Lyu, H.; et al. ILKAP Promotes the Metastasis of Hepatocellular Carcinoma Cells by Inhibiting β-Catenin Degradation and Enhancing the WNT Signaling Pathway. Adv. Biol. 2024, 8, 2300117. [Google Scholar] [CrossRef]
- Lee, A.Q.; Li, Y.; Gong, Z. Inducible Liver Cancer Models in Transgenic Zebrafish to Investigate Cancer Biology. Cancers 2021, 13, 5148. [Google Scholar] [CrossRef]
- Sotty, J.; Bablon, P.; Weiss, P.-H.; Soussan, P. Diethylnitrosamine Induction of Hepatocarcinogenesis in Mice. Methods Mol. Biol. 2024, 2769, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Srimathi Devi, J.; Haripriya, D.; Arul, S.; Saravanan, K.M.; Bupesh, G. Evaluation of anti-cancer effect of zerumbone and cisplatin on N-nitrosodiethylamine induced hepatic cancer in freshwater fish (Danio rerio). Nat. Prod. Res. 2022, 36, 4794–4798. [Google Scholar] [CrossRef] [PubMed]
- Reboredo, M.; Kramer, M.G.; Smerdou, C.; Prieto, J.; Rivas, J.D.L. Transcriptomic Effects of Tet-On and Mifepristone-Inducible Systems in Mouse Liver. Hum. Gene Ther. 2008, 19, 1233–1248. [Google Scholar] [CrossRef] [PubMed]
- Deodato, D.; Asad, N.; Dore, T.M. Photorearrangement of Quinoline-Protected Dialkylanilines and the Photorelease of Aniline-Containing Biological Effectors. J. Org. Chem. 2019, 84, 7342–7353. [Google Scholar] [CrossRef]
- Zhu, Y.; Yang, D.; Duan, X.; Zhang, Y.; Chen, D.; Gong, Z.; Liu, C. Perfluorooctane sulfonate promotes doxycycline-induced liver tumor progression in male Krasv12 transgenic zebrafish. Environ. Res. 2021, 196, 110962. [Google Scholar] [CrossRef]
- Scharfenberger, L.; Hennerici, T.; Király, G.; Kitzmüller, S.; Vernooij, M.; Zielinski, J.G. Transgenic Mouse Technology in Skin Biology: Generation of Complete or Tissue-Specific Knockout Mice. J. Investig. Dermatol. 2014, 134, 1–5. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Model Animal | Advantages | Disadvantages | Main Applications | References |
|---|---|---|---|---|---|
| Mammals | Mouse | Similar genetics to humans, mature gene editing | Liver structure and cancer microenvironment differ from those of humans | Liver cancer mechanisms, tumor growth, new drug screening | [3] |
| Rat | Closer to human structure, suitable for complex surgeries and toxicology studies | Challenging gene editing, high cost | Liver cancer mechanisms, tumor treatment | [3,4] | |
| Rabbit | Moderate size, structure similar to humans | Limited gene editing, low liver regeneration | Drug metabolism, toxicity studies | [5] | |
| Rhesus monkey | Genetic and biological characteristics close to humans | Ethical and cost limitations | Liver cancer mechanisms, drug evaluation | [6] | |
| Pig | Liver size and function similar to humans | High cost, slow reproduction | Liver physiology, surgery, transplantation | [7] | |
| Non-mammals | Zebrafish | Small size, fast reproduction, transparent embryos, rapid gene editing | Different liver structure, does not fully replicate human liver processes | Liver cancer mechanisms, drug screening | [8,9,10] |
| Cell Line | Injection Method | Application | Main Outcome Assessment | References |
|---|---|---|---|---|
| Hep 3B2.1-7 and Li-7 | Microinjection | ADI efficacy against HCC | ADI inhibits BIRC5, FEN1, and the EGFR/PI3K/AKT signaling pathway | [61] |
| HepG2 | Microinjection | HCC cell migration rate | MAT2B knockdown reduces HCC cell migration | [26] |
| HepG2 | Microinjection (yolk sac) | Sorbaria sorbifolia (SS) anti-tumor efficacy | SS downregulates p-Met and p-AKT, inhibiting tumor growth | [27] |
| SK-Hep-1 | Microinjection (yolk sac) | Effect of TB on HCC | TB induces apoptosis in cancer cells | [9] |
| Huh7 (EGFP overexpressed) | Microinjection (yolk sac) | Src and PARP1 combination therapy | Src and PARP1 inhibitors show synergistic lethal effects on HCC | [60] |
| Hep3B_Lifeact-RFP | Microinjection (yolk sac) | Impact of With-no-lysine (K)-1 (WNK1) overexpression | WNK1 knockdown reduces tumor angiogenesis and cell proliferation | [62] |
| BEL-7402 | Microinjection (yolk sac) | NF-κB and c-JUN synergy verification | NF-κB and c-JUN are potential HCC therapy inducers | [63] |
| HepG2-HBx | Microinjection | Anti-metastatic effect of the compound Phyllanthus urinaria L. (CP) | CP inhibits metastasis in HBV-related HCC | [64] |
| HuH-7-Lgr5 | Microinjection (yolk sac) | Metastatic potential of LGR5 cells | LGR5 overexpression enhances metastatic potential | [65] |
| Hep3B-TAp73β | Microinjection (yolk sac) | TAp73β impact on HCC cell migration | TAp73β induces a twofold increase in migration ability | [37] |
| Gene | Construction Method | Specific Findings | Outcome Assessment | References |
|---|---|---|---|---|
| CTNNB1 | Phosphorylation site mutation (4-point) | Activates β-catenin, influences specific lipid metabolism | Common oncogenic mutation in 30% of HCC tumors | [19,74] |
| CD36 | Inject expression vector with Tol2 transposase mRNA | Anti-HCC effect of oligofucose polysaccharides in CD36 model | Anti-HCC, anti-steatosis, anti-fibrosis | [75] |
| CD36/tert | Microinjection and hybridization | Anti-HCC effect of Carassius auratus complex formula (CACF) dose | Anti-HCC in zebrafish xenograft model | [76] |
| krasV12 | Mutant allele introduction into EGFP-krasV12 zebrafish | Affects HCC stress response and liver growth | Mutation reduces liver cancer cell growth and survival | [77] |
| Xmrk | Tet-on system for transgenic zebrafish | Rapid HCC regression upon inducer withdrawal | Xmrk model shows tumor spontaneous regression | [43] |
| twist1a+/kras+ | Doxycycline and 4-hydroxytamoxifen induction | Significant role of LPS in double-transgenic zebrafish | Lipopolysaccharide (LPS) may exacerbate HCC metastasis | [23] |
| twist1a+/xmrk+ | Same as above to create double-transgenic zebrafish | High-dose Dox induces liver tumor metastasis | Stronger metastatic ability in transgenic zebrafish | [25] |
| Myc and Ras | Hybrid generation of double-transgenic zebrafish | Exhibits anorexia/cachexia-like phenotype | Severe muscle atrophy in Tg (Myc and Ras) | [43] |
| Tg (Myc and Ras Mosaic) | Random UAS promoter silencing for new transgenic model | Transforms the entire liver, simulating real HCC heterogeneity | Stable muscle and fat atrophy in Tg (Myc and Ras Mosaic) | [43] |
| CreER/xmrk | Cre/loxP method for new transgenic strain | Single oncogene induction triggers tumors | Dox and 4-Hydroxytamoxifen(4-OHT) treatments achieved time control of Cre-mediated recombination | [35] |
| Myc/xmrk | Hybrid double-transgenic zebrafish model | Androgen KT11 stimulates HCC progression | Significant sex-based differences in HCC progression | [51] |
| Inducer | Construction Method | Application | Outcome Assessment | References |
|---|---|---|---|---|
| Ethanol | Ethanol-induced zebrafish | Alcoholic liver disease (ALD) model | Acute effects reversible, larvae show feeding difficulties, no advanced fibrosis | [14] |
| Mifepristone | LexPR system for liver-specific EGFP-krasV12 expression | Tumor progression and regression in liver | Applicable for tumor progression analysis and anticancer drug screening | [36] |
| Fructose | 4% fructose in culture medium | Induces NAFLD, progresses to HCC | Poor repeatability and stability for primary liver tumor induction in NAFLD model | [24] |
| Doxycycline | Tetracycline-inducible krasV12 transgenic zebrafish | Study of sex differences in HCC development | Examines perfluorooctane sulfonate effects on male HCC progression | [85] |
| High-Fat Diet (HFD) | High-cholesterol diet from days 5–12 | Angiogenesis in HCC | Short-term HFD induces malignant cell and nuclear changes | [40] |
| Gene | Construction Method | Application Direction | Outcome Assessment | References |
|---|---|---|---|---|
| Pten/tp53 | Gene knockout using CRISPR/Cas9 system | Study the roles of the Pten and Tp53 pathways in liver cancer | High tumor incidence and severe clinical symptoms in double-mutant zebrafish | [46] |
| Tg(fabp10a, src, p53) | Triple transgenic zebrafish with diet-induced obesity | Evaluate the impact of 4-Aminobiphenyl (4-ABP) on liver cancer | Activation of the Ras–ERK pathway observed with repeated 4-ABP exposure, promoting liver cancer | [18] |
| tp53−/− | Hybridization of MycAG fish and tp53−/− mutants, PCR genotyping | Study the role of Myc-induced liver tumors in tp53 mutations | tp53 mutation reduces apoptosis and accelerates tumor progression; tumors regress after stopping the inducer | [20] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, Q.; Jin, L.; Peng, R. New Progress in Zebrafish Liver Tumor Models: Techniques and Applications in Hepatocellular Carcinoma Research. Int. J. Mol. Sci. 2025, 26, 780. https://doi.org/10.3390/ijms26020780
Lin Q, Jin L, Peng R. New Progress in Zebrafish Liver Tumor Models: Techniques and Applications in Hepatocellular Carcinoma Research. International Journal of Molecular Sciences. 2025; 26(2):780. https://doi.org/10.3390/ijms26020780
Chicago/Turabian StyleLin, Qizhuan, Libo Jin, and Renyi Peng. 2025. "New Progress in Zebrafish Liver Tumor Models: Techniques and Applications in Hepatocellular Carcinoma Research" International Journal of Molecular Sciences 26, no. 2: 780. https://doi.org/10.3390/ijms26020780
APA StyleLin, Q., Jin, L., & Peng, R. (2025). New Progress in Zebrafish Liver Tumor Models: Techniques and Applications in Hepatocellular Carcinoma Research. International Journal of Molecular Sciences, 26(2), 780. https://doi.org/10.3390/ijms26020780