
Identification of Novel 58-5p and SREBF1 Interaction and Effects on Apoptosis of Ovine Ovarian Granulosa Cell
 ,
,
Abstract
1. Introduction
2. Results
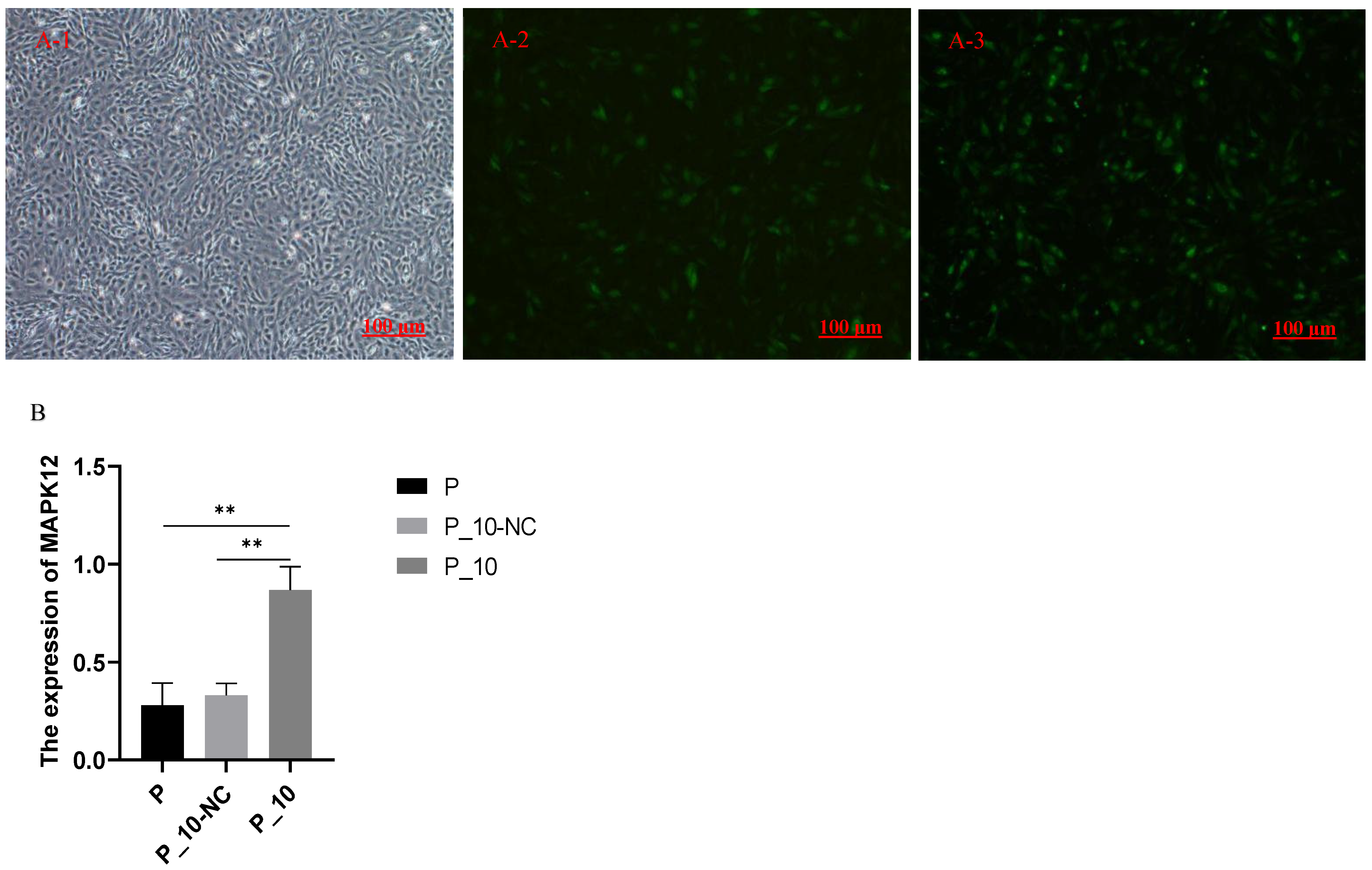
2.1. Expression of MAPK12 After Overexpression of MAPK12
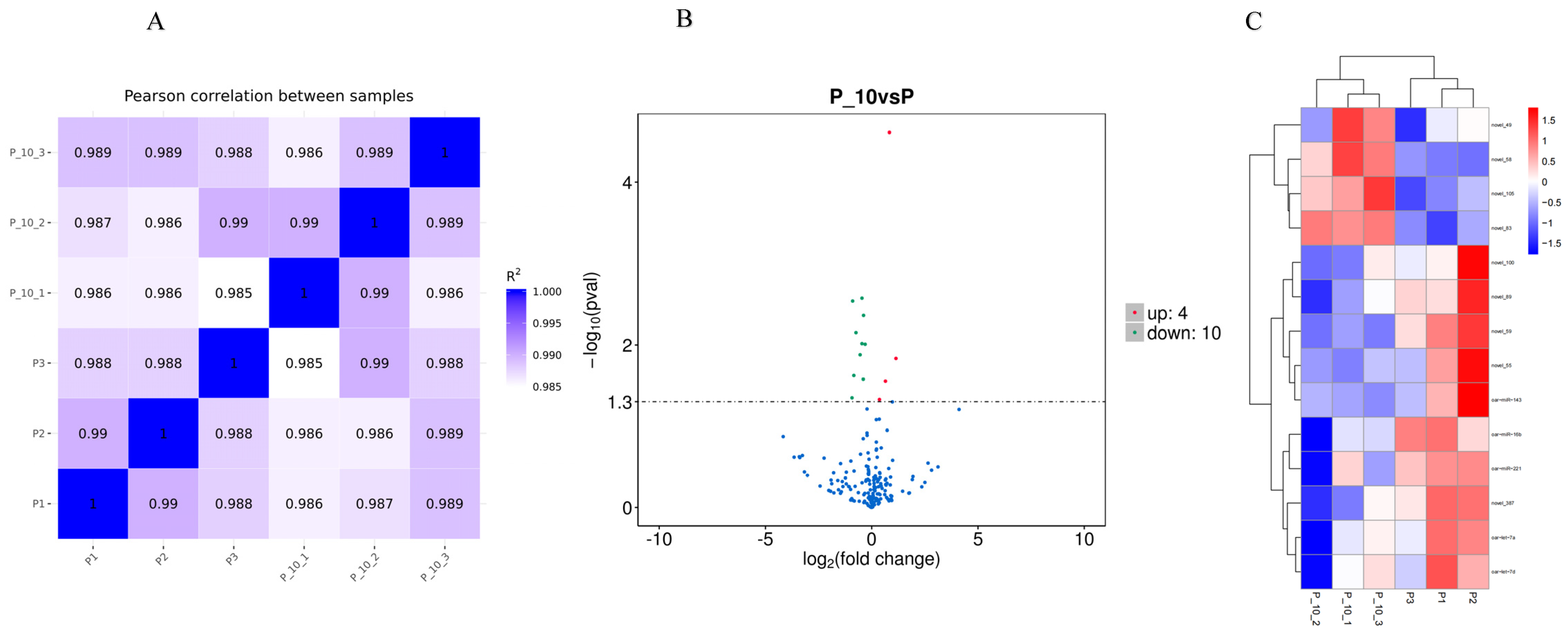
2.2. Small-RNA Identification and the Analysis of DE miRNAs
2.2.1. Small-RNA Identification
2.2.2. The Analysis of DE miRNAs
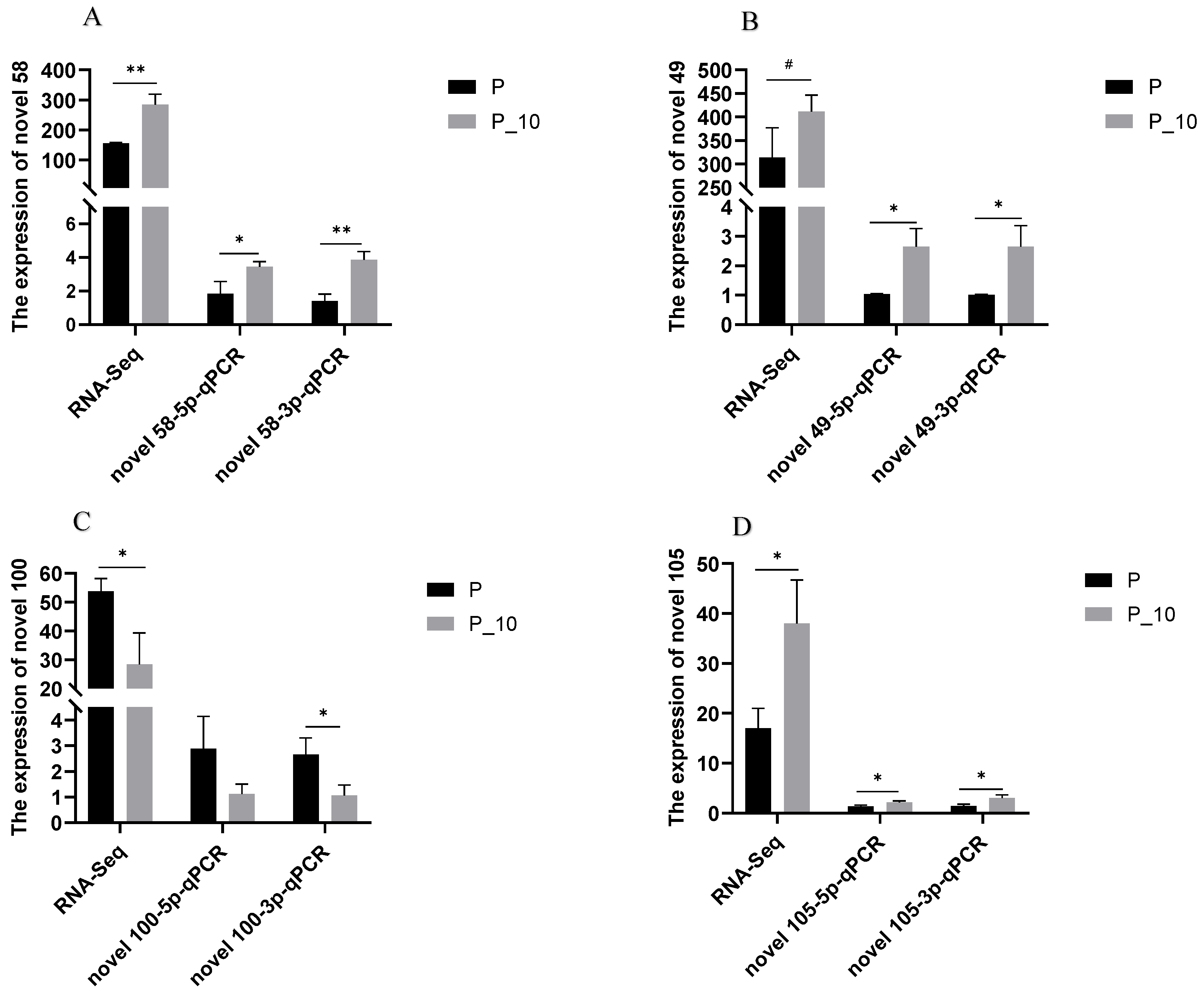
2.2.3. RT-qPCR Confirmation
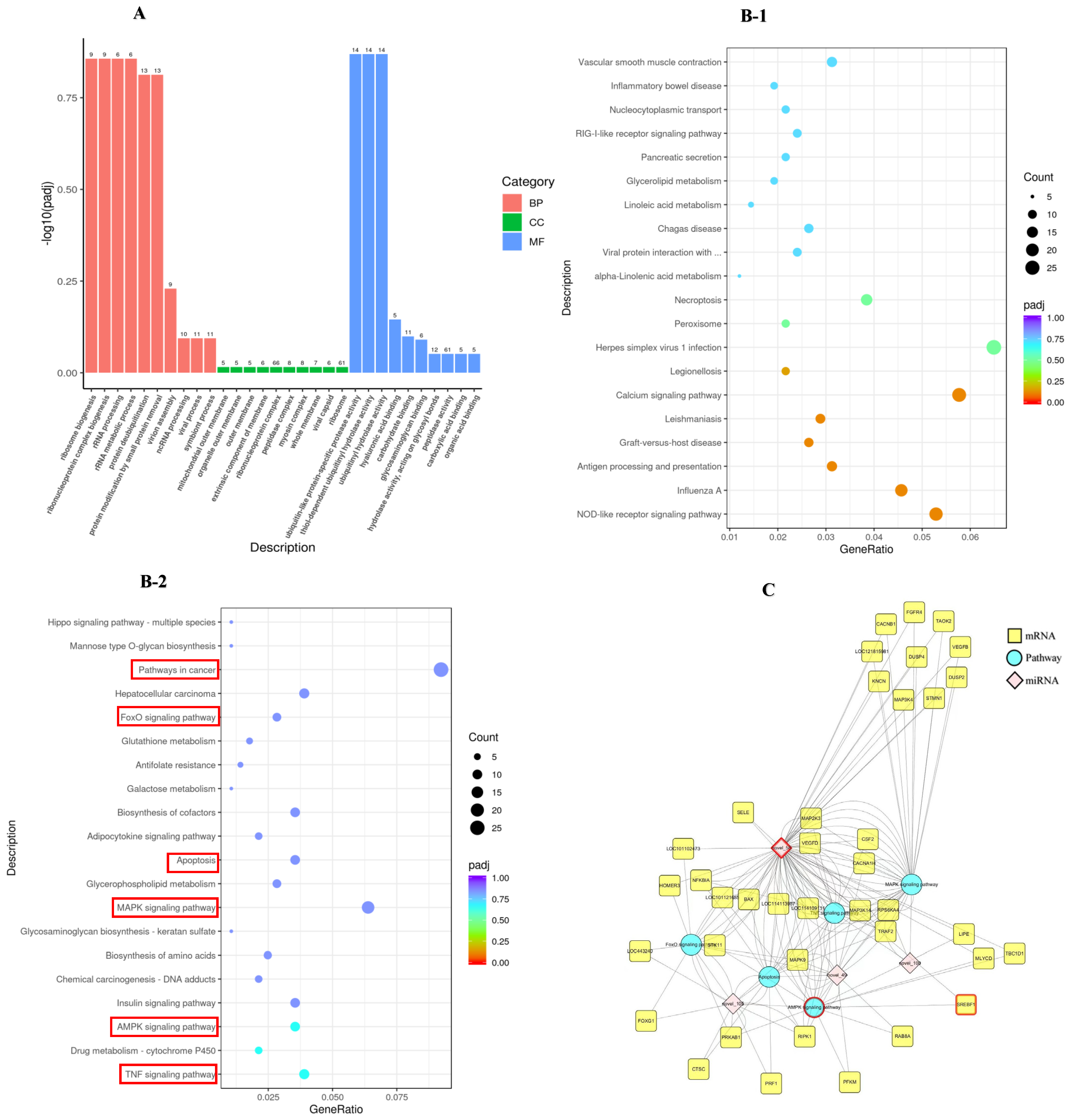
2.3. Functional Enrichment Analysis
2.3.1. GO Functional and KEGG Pathway Analysis
2.3.2. miRNAs-mRNAs-KEGG Pathway Interaction Network
2.4. The Effects of Novel 58 on Apoptosis in High PRL Concentration GCs
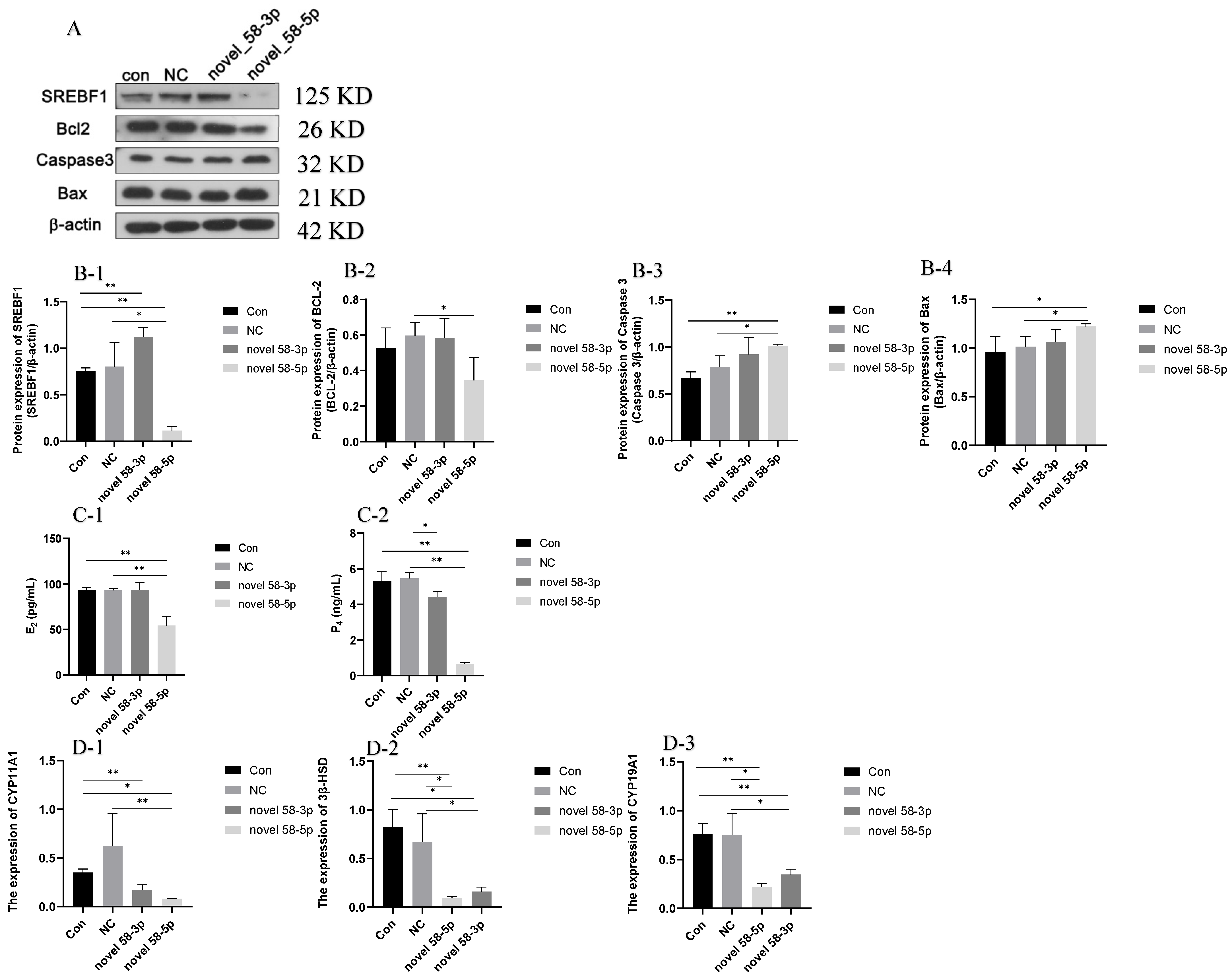
2.4.1. SREBF1 and Apoptosis-Related Proteins Expression Analysis
2.4.2. Steroid Hormone Secretion and Related Gene Expression
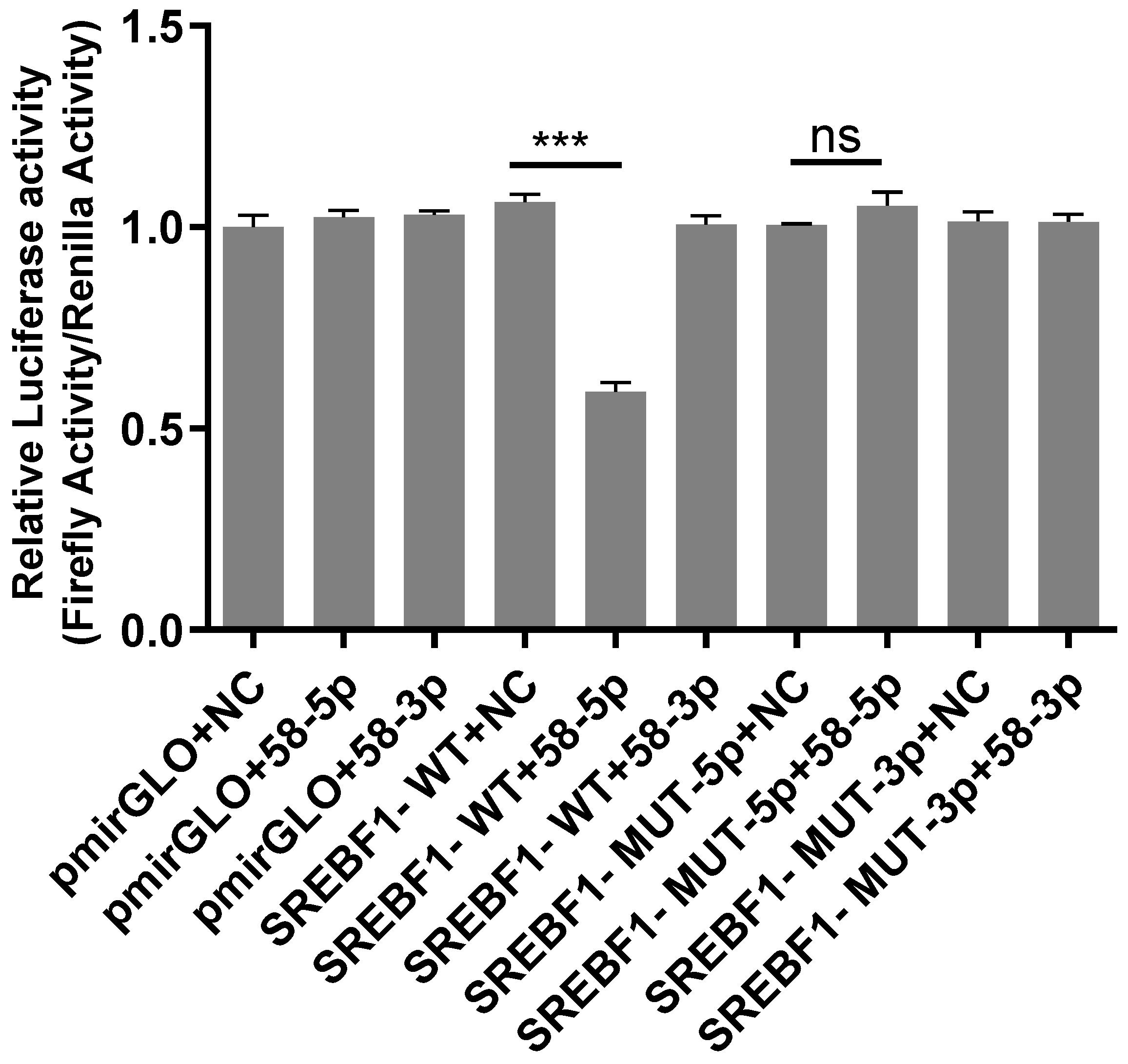
2.5. Analysis of Dual-Luciferase Reporter Gene
2.6. Bioanalysis of SREBF1
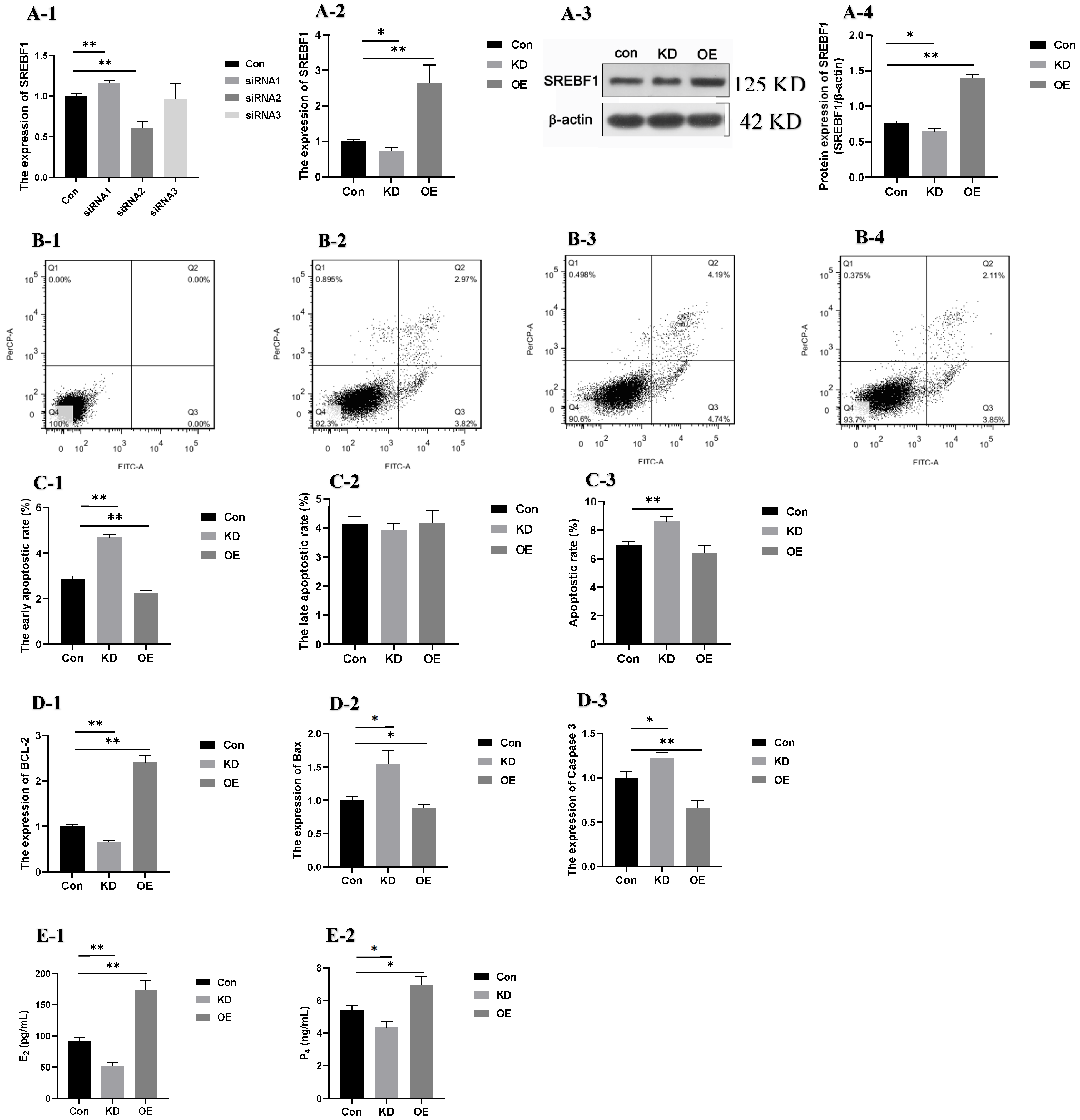
2.7. Apoptosis and the Secretion of Steroid Hormones After Interference and Overexpression of SREBF1 in GCs with High PRL Concentration
2.7.1. Apoptostic Rate and the Expression of Apoptosis-Related Genes
2.7.2. The Secretion of Steroid Hormones (E2 and P4)
3. Discussion
4. Materials and Methods
4.1. Experimental Design and Ovine Ovarian GCs Culture
4.2. GCs Infection and RT-qPCR
4.2.1. The GCs Model of Infection
4.2.2. RNA Extraction and RT-qPCR
4.3. RNA-Seq and Bioinformatics Analyses
4.3.1. RNA Extraction, Library Construction and Quality Control
4.3.2. Alignment and Identification of Small RNA
4.3.3. Differential Expression of miRNA and RT-qPCR Confirmation
4.3.4. Target Gene Prediction and Functional Enrichment Analysis
4.3.5. Construction of miRNAs-mRNAs-KEGG Pathway Interaction Network
4.4. Overexpression of Novel 58
4.4.1. The Transfection of Novel 58 Related RNA
4.4.2. Western Blotting
4.4.3. Steroid Hormone Secretion and Related Gene Expression
4.5. Dual-Luciferase Reporter Gene Assay
4.6. Functional Validation of SREBF1
4.6.1. Bioanalysis of SREBF1
4.6.2. Gene Editing of SREBF1 in GCs with a High PRL Concentration
4.6.3. Apoptosis Assay
4.6.4. Steroid Hormones Secretion
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bush, S.J.; McCulloch, M.E.B.; Muriuki, C.; Salavati, M.; Davis, G.M.; Farquhar, I.L.; Lisowski, Z.M.; Archibald, A.L.; Hume, D.A.; Clark, E.L. Comprehensive transcriptional profiling of the gastrointestinal tract of ruminants from birth to adulthood reveals strong developmental stage specific gene expression. G3 Genes Genomes Genet. 2019, 9, 359–373. [Google Scholar] [CrossRef] [PubMed]
- Kaltungo, B.Y.; Musa, I.W. A Review of Some Protozoan Parasites Causing Infertility in Farm Animals. ISRN 2013, 2013, 1–6. [Google Scholar] [CrossRef]
- Carson, S.A.; Kallen, A.N. Diagnosis and Management of Infertility: A Review. JAMA 2021, 326, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Quirk, S.M.; Cowan, R.G.; Harman, R.M.; Hu, C.L.; Porter, D.A. Ovarian follicular growth and atresia: The relationship between cell proliferation and survival. J. Anim. Sci. 2004, 82 (Suppl. S13), E40–E52. [Google Scholar] [CrossRef] [PubMed]
- Albertini, D.F.; Combelles, C.M.H.; Benecchi, E.; Carabatsos, M.J. Cellular basis for paracrine regulation of ovarian follicle development. Reproduction 2001, 121, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Dumesic, D.A.; Meldrum, D.R.; Katz-Jaffe, M.G.; Krisher, R.L.; Schoolcraft, W.B. Oocyte environment: Follicular fluid and cumulus cells are critical for oocyte health. Fertil. Steril. 2015, 103, 303–316. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xu, Y.; Liu, H.; Pan, Z. MicroRNAs in ovarian follicular atresia and granulosa cell apoptosis. Reprod. Biol. Endocrinol. 2019, 17, 9. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.M.; Li, Y.Y.; Fei, C.F.; Zhou, L.Q. Insm2 deficiency results in female infertility by disturbing steroid pathway and decreasing ovarian reserve in mice. Cell Cycle 2022, 21, 2255–2267. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Duan, H.; You, X.; Peng, Q.; Yuan, N.; Sha, R.; Xie, Z.; Feng, Y. Deoxynivalenol triggers mitotic catastrophe and apoptosis in C2C12 myoblasts. Ecotoxicol. Environ. Saf. 2024, 281, 116607. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Ye, Z.; Xu, C.; Chen, H.; Dai, S.; Chen, W.; Bao, G. Corosolic acid enhances oxidative stress-induced apoptosis and senescence in pancreatic cancer cells by inhibiting the JAK2/STAT3 pathway. Mol. Biol. Rep. 2024, 51, 176. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, R.; Li, M.; Sun, J.; Liu, J.; Xie, M.; Chen, Y.; Tang, S. Zuojin Pill inhibits the proliferation of esophageal cancer cells by inducing ROS production and apoptosis via regulation of PI3K/Akt signalling. J. Taibah Univ. Sci. 2024, 18, 2361991. [Google Scholar] [CrossRef]
- Wang, X.; Wang, T.; Yao, J.; Li, X.; Deng, R.; Zheng, P. Bta-miR-149-3p suppresses inflammatory response in bovine Sertoli cells exposed to microcystin-leucine arginine (MC-LR) through TLR4/NF-kB signaling pathway. Ecotoxicol. Environ. Saf. 2024, 281, 116636. [Google Scholar]
- Yang, R.C.; Duan, C.H.; Zhang, S.; Liu, Y.Q.; Zhang, Y.J. Prolactin Regulates Ovine Ovarian Granulosa Cell Apoptosis by Affecting the Expression of MAPK12 Gene. Int. J. Mol. Sci. 2023, 24, 10269. [Google Scholar] [CrossRef] [PubMed]
- Ruan, K.; Fang, X.; Ouyang, G.L. MicroRNAs: Novel regulators in the hallmarks of human cancer. Cancer Lett. 2009, 285, 116–126. [Google Scholar] [CrossRef] [PubMed]
- Shiomi, E.; Sugai, T.; Ishida, K.; Osakabe, M.; Tsuyukubo, T.; Kato, Y.; Takata, R.; Obara, W. Analysis of Expression Patterns of MicroRNAs That Are Closely Associated with Renal Carcinogenesis. Front. Oncol. 2019, 9, 431. [Google Scholar] [CrossRef]
- Sun, X.F.; Li, Y.P.; Pan, B.; Wang, Y.F.; Li, J.; Shen, W. Molecular regulation of miR-378 on the development of mouse follicle and the maturation of oocyte in vivo. Cell Cycle 2018, 17, 2230–2242. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, R.; Hao, J.; Huang, X.; Liu, M.; Lv, M.; Su, C.; Mu, Y.L. miRNA-122-5p in POI ovarian-derived exosomes promotes granulosa cell apoptosis by regulating BCL9. Cancer Med. 2022, 11, 2414–2426. [Google Scholar] [CrossRef] [PubMed]
- Bahmyari, S.; Jamali, Z.; Khatami, S.H.; Vakili, O.; Roozitalab, M.; Savardashtaki, A.; Solati, A.; Mousavi, P.; Shabaninejad, Z.; Vakili, S. microRNAs in female infertility: An overview. Cell Biochem. Funct. 2021, 39, 955–969. [Google Scholar] [CrossRef]
- Peng, W.; Sha, H.; Sun, X.; Zou, R.; Zhu, Y.; Zhou, G.; Feng, J. Role and mechanism of miR-187 in human cancer. Am. J. Transl. Res. 2020, 12, 4873–4884. [Google Scholar] [PubMed]
- Withers, H.G. Functional Genomic Screens for Novel Metastasis Suppressor Genes in Breast and Prostate Cancer Models. Ph.D. Thesis, State University of New York at Buffalo, Buffalo, NY, USA, 2018. [Google Scholar]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef] [PubMed]
- Wen, M.; Shen, Y.; Shi, S.; Tang, T. miREvo. an integrative microRNA evolutionary analysis platform for next-generation sequencing experiments. BMC Bioinform. 2012, 13, 140. [Google Scholar] [CrossRef]
- Friedlaender, M.R.; Mackowiak, S.D.; Li, N.; Chen, W.; Rajewsky, N. miRDeep2 accurately identifies known and hundreds of novel microRNA genes in seven animal clades. Nucleic Acids Res. 2012, 40, 37–52. [Google Scholar] [CrossRef] [PubMed]
- Enright, A.; John, B.; Gaul, U.; Tuschl, T.; Sander, C.; Marks, D.G. MicroRNA targets in Drosophila. Genome Biol. 2003, 5, R1. [Google Scholar] [CrossRef] [PubMed]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A.R.B. goseq: Gene Ontology testing for RNA-seq datasets. Biology 2012. [Google Scholar]
- Liu, S.; Li, L.; Li, M.; Zhang, J. Effect of miR-26b-5p on cis-diamine dichloroplatinum-induced ovarian granulosa cell injury by targeting MAP3K9. Vitr. Cell Dev. Biol. Anim. 2020, 56, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhao, W.; Wang, L.; Luo, Q.; Yin, N.; Lu, X.; Hou, Y.; Cui, J.; Zhang, H. Human placenta-derived mesenchymal stem cells inhibit apoptosis of granulosa cells induced by IRE1α pathway in autoimmune POF mice. Cell Biol. Int. 2019, 43, 899–909. [Google Scholar] [CrossRef]
- Chen, H.; Wang, X.; Guo, F.; Li, P.; Peng, D.; He, J. Impact of p38 gamma mitogen-activated protein kinase (MAPK) on MDA-MB-231 breast cancer cells using metabolomic approach. Int. J. Biochem. Cell Biol. 2019, 107, 6–13. [Google Scholar] [CrossRef]
- Gui, Z.L.; Wu, T.L.; Zhao, G.C.; Lin, Z.X.; Xu, H.G. MicroRNA-497 suppress osteosarcoma by targeting MAPK/Erk pathway. Bratisl. Med. J. 2017, 118, 449–452. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.; Wang, J.; Han, X.; Wang, Q.; Zhang, S.; Liang, S.; Li, H.; Meng, L.; Zhang, C.; Chen, H. Identification of small extracellular vesicle-linked miRNA specifically derived from intrafollicular cells in women with polycystic ovary syndrome. Reprod. Biomed. Online 2021, 42, 870–880. [Google Scholar] [CrossRef] [PubMed]
- Bakhshalizadeh, S.; Amidi, F.; Shirazi, R.; Nashtaei, M.S. Vitamin D3 regulates steroidogenesis in granulosa cells through AMP-activated protein kinase (AMPK) activation in a mouse model of polycystic ovary syndrome. Cell Biochem. Funct. 2018, 36, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Zha, J.K.; Shu, H.B. Molecular mechanism of signaling by tumor necrosis factor. Sci. China Ser. C 2002, 45, 113–119. [Google Scholar] [CrossRef]
- Fu, X.; He, Y.; Wang, X.; Peng, D.; Chen, X.; Li, X.; Wang, Q. Overexpression of miR-21 in stem cells improves ovarian structure and function in rats with chemotherapy-induced ovarian damage by targeting PDCD4 and PTEN to inhibit granulosa cell apoptosis. Stem Cell Res. Ther. 2017, 8, 187. [Google Scholar] [CrossRef] [PubMed]
- Payne, A.H.; Hales, D.B. Overview of steroidogenic enzymes in the pathway from cholesterol to active steroid hormones. Endocr. Rev. 2004, 25, 947–970. [Google Scholar] [CrossRef]
- Ebeling, S.; Topfer, D.; Meinecke, B. Steroidogenesis and the Influence of MAPK Activity During In Vitro Maturation of Porcine Cumulus Oocyte Complexes. Reprod. Domest. Anim. 2011, 46, 513–519. [Google Scholar] [CrossRef]
- Wang, W.; Sun, Y.; Liu, J.; Li, Y.; Li, H.; Xiao, S.; Weng, S.; Zhang, W. Soy isoflavones administered to rats from weaning until sexual maturity affect ovarian follicle development by inducing apoptosis. Food Chem. Toxicol. 2014, 72, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, F.; Inoue, N.; Manabe, N.; Ohkura, S. Follicular Growth and Atresia in Mammalian Ovaries: Regulation by Survival and Death of Granulosa Cells. J. Reprod. Dev. 2012, 58, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Shang, J.; Chen, Z.Z.; Wang, Z.H.; Wei, T.N.; Wu, W.B.; Chen, W.M. Effect of Expression Levels of MiRNA-132, -125, -143 and -145 on Autophagy and Apoptosis of Multiple Myeloma Cells. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2018, 26, 1688–1694. [Google Scholar] [PubMed]
- Hu, Z.; Shen, W.J.; Cortez, Y.; Tang, X.; Liu, L.F.; Kraemer, F.B.; Azhar, S. Hormonal Regulation of MicroRNA Expression in Steroid Producing Cells of the Ovary, Testis and Adrenal Gland. PLoS ONE 2013, 8, e78040. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, S.; Cai, Z.; Dong, W.; Ye, J.; Cai, Z.; Han, Z.; Liang, Y.; Zhuo, Y.; Luo, Y. Down-regulation of ACACA suppresses the malignant progression of Prostate Cancer through inhibiting mitochondrial potential. J. Cancer 2021, 12, 232–243. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.Y.; Xu, J.J.; Li, C.; Yu, J.L.; Wu, Y.T.; Huang, H.F. A PPARG Splice Variant in Granulosa Cells Is Associated with Polycystic Ovary Syndrome. J. Clin. Med. 2022, 11, 7285. [Google Scholar] [CrossRef]
- Shafiee, M.N.; Mongan, N.; Seedhouse, C.; Chapman, C.; Deen, S.; Abu, J.; Atiomo, W. Sterol regulatory element binding protein-1 (SREBP1) gene expression is similarly increased in polycystic ovary syndrome and endometrial cancer. Acta Obstet. Gynecol. Scand. 2017, 96, 556–562. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; He, W.; Luo, M.; Zhou, Y.; Chang, G.; Ren, W.; Wu, K.; Li, X.; Shen, J.; Zhao, X. SREBP1 regulates tumorigenesis and prognosis of pancreatic cancer through targeting lipid metabolism. Tumor Biol. 2015, 36, 4133–4141. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Xia, H.; Jiang, J.; Xu, X.; Li, M.; Chen, Z.; Sun, Y.; Zhang, H.; Yang, Z. MicroRNA-212 targets SIRT2 to influence lipogenesis in bovine mammary epithelial cell line. J. Dairy. Res. 2020, 87, 232–238. [Google Scholar] [CrossRef]
- Dai, J.; Jiang, M.; Hu, Y.; Xiao, J.; Hu, B.; Xu, J.; Han, X.; Shen, S.; Li, B.; Wu, Z. Dysregulated SREBP1c/miR-153 signaling induced by hypertriglyceridemia worsens acute pancreatitis and delays tissue repair. JCI Insight 2021, 6, 138584. [Google Scholar] [CrossRef] [PubMed]
- Ortega, R.; Liu, B.; Persaud, S.J. Effects of miR-33 Deficiency on Metabolic and Cardiovascular Diseases: Implications for Therapeutic Intervention. Int. J. Mol. Sci. 2023, 24, 10777. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Sequence (5′-3′) | Tm (°C) |
|---|---|---|
| P53 | F: AAGCAGGGCTCATTCTAGCC R: GGCCCTTCTCTCTTGAGCAT | 60 |
| MAPK12 | F: GCAGGCAGACAGCGAGAT R: GGTCAGGACGGAGGCAAA | 62 |
| CYP11A1 | F: GTTTCGCTTTGCCTTTGAGTC R: ACAGTTCTGGAGGGAGGTTGA | 60 |
| 3β-HSD | F: CAGTCTATGTTGGCAATGTGGC R: CGGTTGAAGCAGGGGTGGTAT | 60 |
| CYP19A1 | F: GCTTTTGGAAGTGCTGAACCC R: CATGCCGATGAACTGCAACC | 60 |
| SREBF1 | F: TTGGAGCGAGCACTGAATTG R: GTTCAGCAGCTGCAGGTATC | 60 |
| BCL-2 | F: CCTTTGTGGAGCTGTATGGC R: CCTTTGTGGAGCTGTATGGC | 60 |
| Bax | F: CATCATGGGCTGGACATTGG R: GTGGGTGTCCCAAAGTAGGA | 60 |
| Caspase3 | F: TTCAGAGGGGACTGTTGCAG R: CAGTCCAGTTCTGTGCCTCG | 60 |
| β-actin | F: TCAGCAAGCAGGAGTACGAC R: GGGTGTAACGCAGCTAACAG | 60 |
| Name | Sequence |
|---|---|
| novel_58-5p | novel_58-5p RT: GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACAACAAC |
| novel_58-5p (F): GCGTGGCAGTGTCTTAGCTG | |
| novel_58-3p | novel_58-3p RT: GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACTAGGGC |
| novel_58-3p (F): CGCGCAATCAGCAAGTATACT | |
| novel_49-5p | novel_49-5p RT: GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACGGAGGC |
| novel_49-5p (F): GCGTATTGCACTCGTCCCG | |
| novel_49-3p | novel_49-3p RT: GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACACACTG |
| novel_49-3p (F): GGGACGGGACGCGGTG | |
| novel_100-5p | novel_100-5p RT: GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACAGTCAG |
| novel_100-5p (F): GCGCGTGGATAACGCGT | |
| novel_100-3p | novel_100-3p RT: GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACAAAACA |
| novel_100-3p (F): CGCGGGGAGACGCGTG | |
| novel_105-5p | novel_105-5p RT: GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACGCAATC |
| novel_105-5p (F): CGCGAGGCAGTGTAGTTAGCT | |
| novel_105-3p | novel_105-3p RT: GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACCCTGGC |
| novel_105-3p (F): CGCGAATCACTAACCACACG | |
| U6 | U6 (F): GCGCGCTCGCTTCGGC |
| U6 (R): AGTGCAGGGTCCGAGGTATT |
| Name | Sequence (5′-3′) |
|---|---|
| mimics NC | Sense: UUGUACUACACAAAAGUACUG Antisense: GUACUUUUGUGUAGUACAAUU |
| Novel 58-3p mimics | Sense: CAAUCAGCAAGUAUACUGCCCUA Antisense: GGGCAGUAUACUUGCUGAUUGUU |
| Novel 58-5p mimics | Sense: UGGCAGUGUCUUAGCUGGUUGUU Antisense: CAACCAGCUAAGACACUGCCAUU |
| Name | Sequence (5′-3′) |
|---|---|
| siRNA1 | F: GCCGCGGAACCAUGGACUGCACGUU R: AACGUGCAGUCCAUGGUUCCGCGGC |
| siRNA2 | F: UCCUUCCACGAUGAGCUCCUCACUU R: AAGUGAGGAGCUCAUCGUGGAAGGA |
| siRNA3 | F: CGGAGCUCUCCUGCCGCAGAGUGUU R: AACACUCUGCGGCAGGAGAGCUCCG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, R.; Wang, Y.; Yue, S.; Liu, Y.; Zhang, Y.; Duan, C. Identification of Novel 58-5p and SREBF1 Interaction and Effects on Apoptosis of Ovine Ovarian Granulosa Cell. Int. J. Mol. Sci. 2025, 26, 576. https://doi.org/10.3390/ijms26020576
Yang R, Wang Y, Yue S, Liu Y, Zhang Y, Duan C. Identification of Novel 58-5p and SREBF1 Interaction and Effects on Apoptosis of Ovine Ovarian Granulosa Cell. International Journal of Molecular Sciences. 2025; 26(2):576. https://doi.org/10.3390/ijms26020576
Chicago/Turabian StyleYang, Ruochen, Yong Wang, Sicong Yue, Yueqin Liu, Yingjie Zhang, and Chunhui Duan. 2025. "Identification of Novel 58-5p and SREBF1 Interaction and Effects on Apoptosis of Ovine Ovarian Granulosa Cell" International Journal of Molecular Sciences 26, no. 2: 576. https://doi.org/10.3390/ijms26020576
APA StyleYang, R., Wang, Y., Yue, S., Liu, Y., Zhang, Y., & Duan, C. (2025). Identification of Novel 58-5p and SREBF1 Interaction and Effects on Apoptosis of Ovine Ovarian Granulosa Cell. International Journal of Molecular Sciences, 26(2), 576. https://doi.org/10.3390/ijms26020576