
Escalated Oxycodone Self-Administration Is Associated with Activation of Specific Gene Networks in the Rat Dorsal Striatum


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Rats Exposed to LgA Oxycodone Self-Administration Escalate Their Drug Intake over Time
2.2. RNA Sequencing Identifies Specific Differentially Expressed Genes in the Dorsal Striatum of LgA-H, LgA-L, and ShA Rats
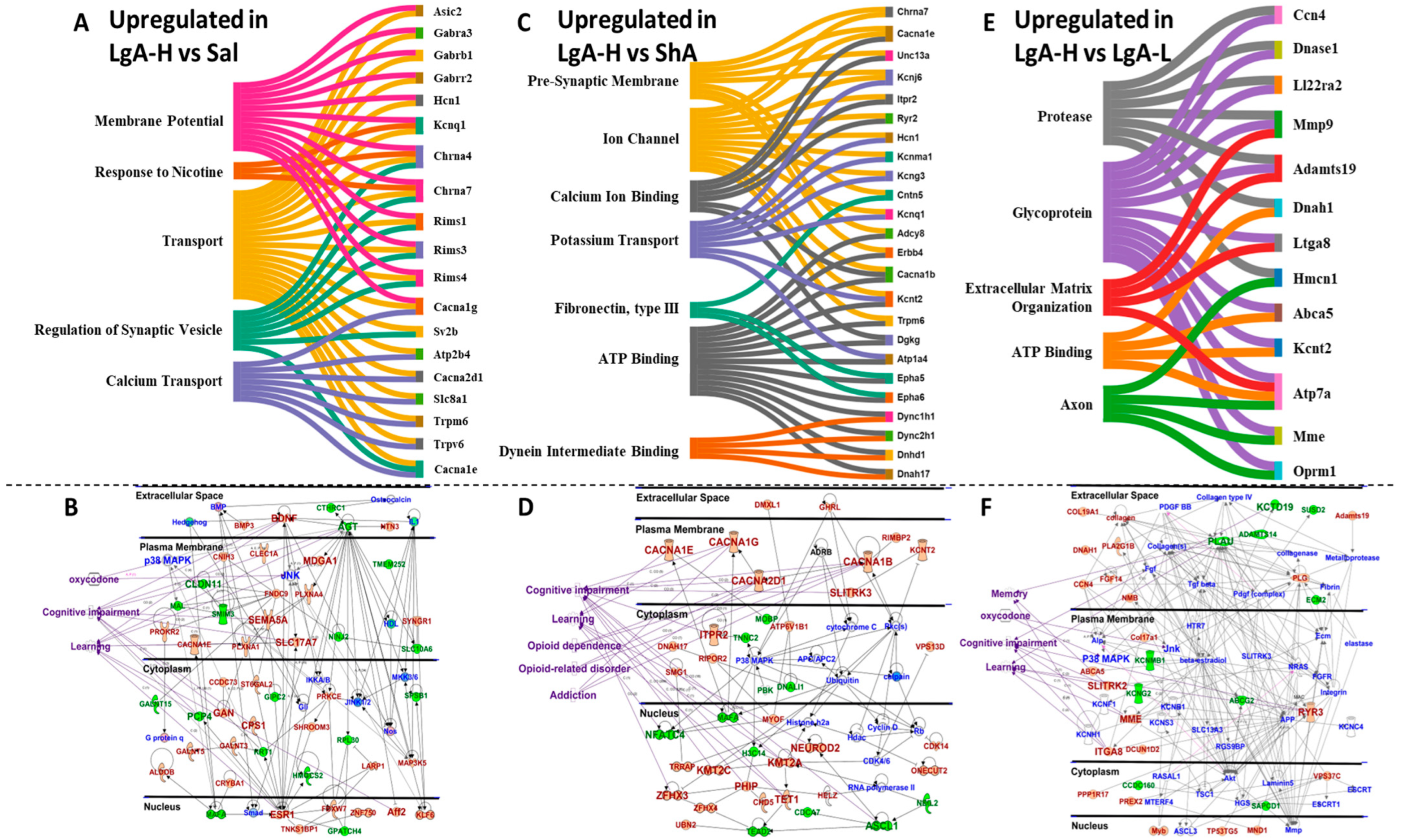
2.3. Striatal Genes Are Differentially Expressed in LgA-H
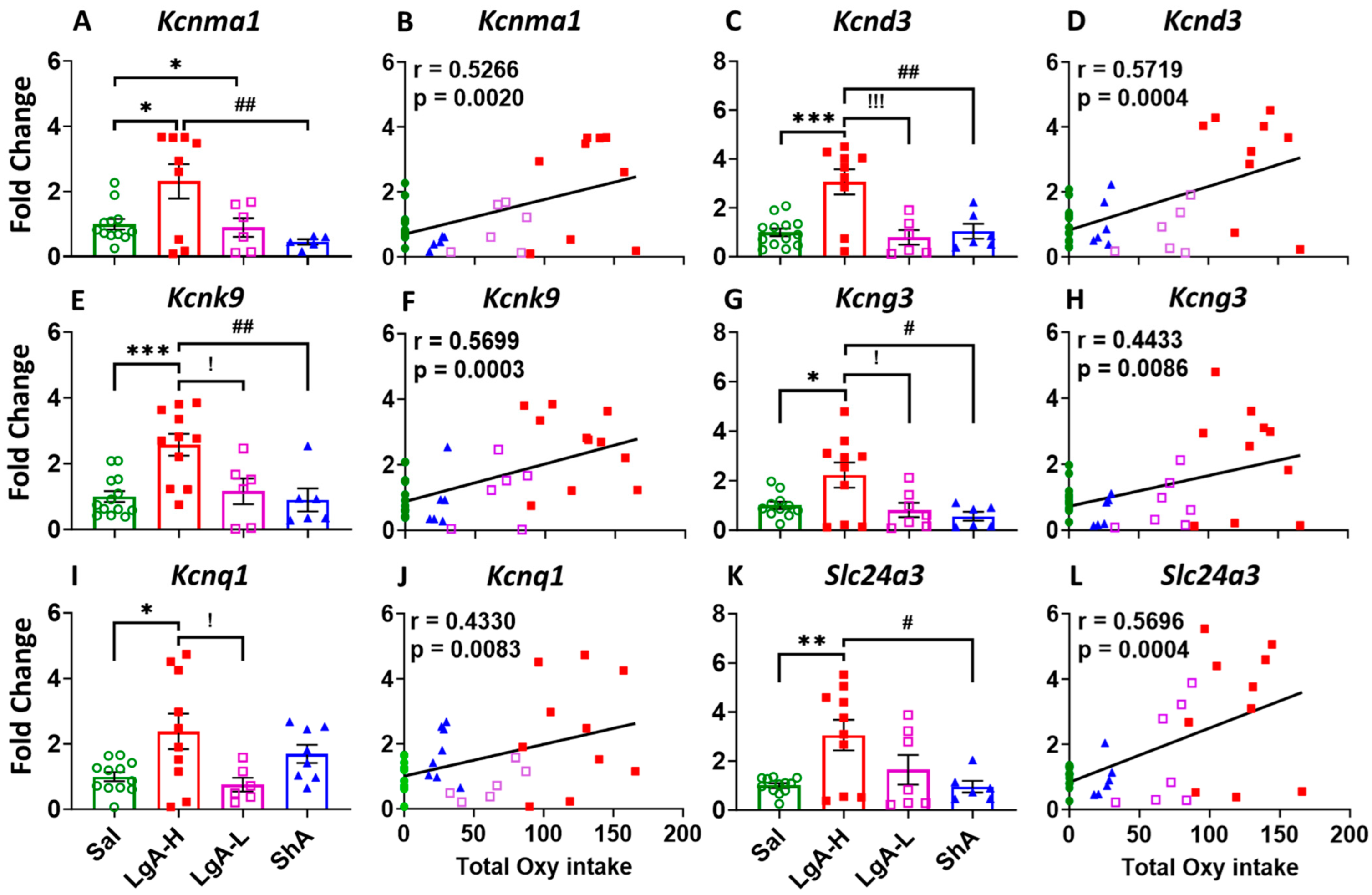
2.4. Increased Oxycodone Intake Is Associated with Upregulation of Potassium Channel Genes in the Dorsal Striatum of LgA-H Rats
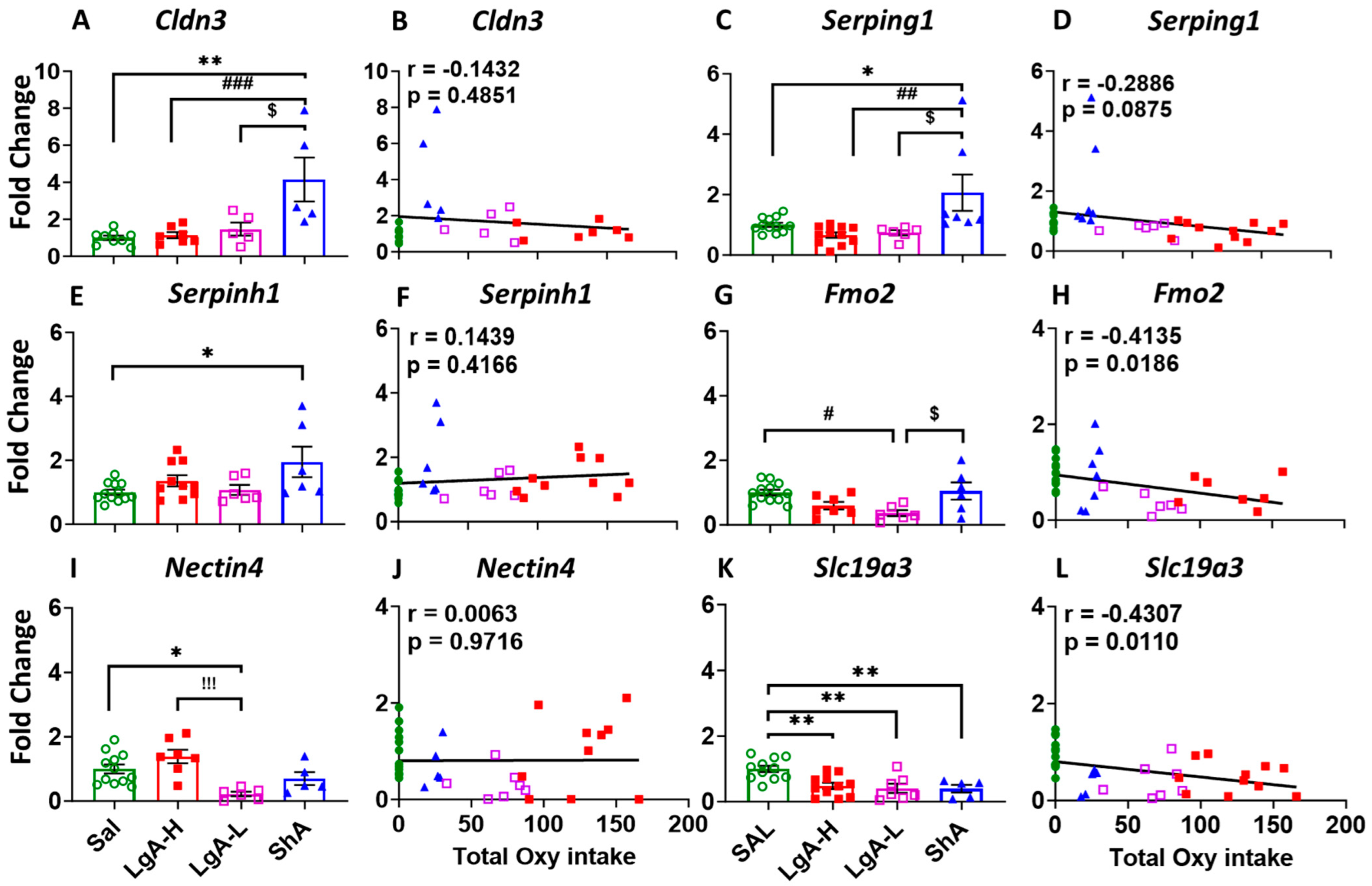
2.5. LgA-L and ShA Rats Showed Differential Gene Expression in the Dorsal Striatum
3. Discussion
3.1. Involvement of Potassium Channel Genes in High Oxycodone Intake
3.2. Molecular Mechanisms Associated with Low Oxycodone Intake
4. Materials and Methods
4.1. Animals
4.2. Intravenous Surgery
4.3. Oxycodone Self-Administration
4.4. RNA Extraction and Sequencing
4.5. Quantitative RT-PCR
4.6. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bergeria, C.L.; Strain, E.C. Opioid Use Disorder: Pernicious and Persistent. Am. J. Psychiatry 2022, 179, 708–714. [Google Scholar] [CrossRef]
- Boscarino, J.A.; Rukstalis, M.; Hoffman, S.N.; Han, J.J.; Erlich, P.M.; Gerhard, G.S.; Stewart, W.F. Risk factors for drug dependence among out-patients on opioid therapy in a large US health-care system. Addiction 2010, 105, 1776–1782. [Google Scholar] [CrossRef]
- King, S.J.; Reid, C.; Forbes, K.; Hanks, G. A systematic review of oxycodone in the management of cancer pain. Palliat. Med. 2011, 25, 454–740. [Google Scholar] [CrossRef]
- Marie, N.; Noble, F. Oxycodone, an opioid like the others? Front. Psychiatry 2023, 14, 1229439. [Google Scholar] [CrossRef]
- National Institute on Drug Abuse. Advancing Addiciton Science. Available online: https://nida.nih.gov/research-topics/trends-statistics/overdose-death-rates (accessed on 13 January 2025).
- Riley, J.; Eisenberg, E.; Muller-Schwefe, G.; Drewes, A.M.; Arendt-Nielsen, L. Oxycodone: A review of its use in the management of pain. Curr. Med. Res. Opin. 2008, 24, 175–192. [Google Scholar] [CrossRef] [PubMed]
- Hamina, A.; Muller, A.E.; Clausen, T.; Skurtveit, S.; Hesse, M.; Tjagvad, C.; Thylstrup, B.; Odsbu, I.; Zoega, H.; Jónsdóttir, H.L.; et al. Prescription opioids among older adults: Ten years of data across five countries. BMC Geriatr. 2022, 22, 429. [Google Scholar] [CrossRef] [PubMed]
- Ellis, M.S.; Cicero, T.J.; Dart, R.C.; Green, J.L. Understanding multi-pill ingestion of prescription opioids: Prevalence, characteristics, and motivation. Pharmacoepidemiol. Drug Saf. 2019, 28, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Kibaly, C.; Alderete, J.A.; Liu, S.H.; Nasef, H.S.; Law, P.Y.; Evans, C.J.; Cahill, C.M. Oxycodone in the Opioid Epidemic: High ’Liking’, ’Wanting’, and Abuse Liability. Cell. Mol. Neurobiol. 2021, 41, 899–926. [Google Scholar] [CrossRef]
- Chamakalayil, S.; Stohler, R.; Moldovanyi, A.; Gerber, M.; Brand, S.; Dürsteler, K.M. Neurocognitive performance of patients undergoing intravenous versus oral opioid agonist treatment: A prospective multicenter study on three-month treatment effects. Front. Psychiatry 2024, 15, 1375895. [Google Scholar] [CrossRef] [PubMed]
- Blackwood, C.A.; Cadet, J.L. The molecular neurobiology and neuropathology of opioid use disorder. Curr. Res. Neurobiol. 2021, 2, 100023. [Google Scholar] [CrossRef]
- Cadet, J.L.; Bisagno, V.; Milroy, C.M. Neuropathology of substance use disorders. Acta Neuropathol. 2014, 127, 91–107. [Google Scholar] [CrossRef] [PubMed]
- Sahebi-Fakhrabad, A.; Sadeghi, A.H.; Kemahlioglu-Ziya, E.; Handfield, R. Exploring Opioid Prescription Patterns and Overdose Rates in South Carolina (2017–2021): Insights into Rising Deaths in High-Risk Areas. Healthcare 2024, 12, 1268. [Google Scholar] [CrossRef] [PubMed]
- Tsang, A.; Rodda, L.N. Assessment of postmortem urine fentanyl detection by autopsy dipstick testing in accidental overdose deaths. J. Anal. Toxicol. 2024, 8, 667–671. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.M.; Chen, J.; Novak, P.J. Utilization of Buprenorphine for Opioid Use Disorder After the Practitioner Waiver Removal. Am. J. Prev. Med. 2024, 68, 207–209. [Google Scholar] [CrossRef]
- Grande, L.A.; Cundiff, D.; Greenwald, M.K.; Murray, M.; Wright, T.E.; Martin, S.A. Evidence on Buprenorphine Dose Limits: A Review. J. Addict. Med. 2023, 17, 509–516. [Google Scholar] [CrossRef]
- Hayes, C.J.; Raciborski, R.A.; Nowak, M.; Acharya, M.; Nunes, E.V., Jr.; Winhusen, T.J. Medications for opioid use disorder: Predictors of early discontinuation and reduction of overdose risk in US military veterans by medication type. Addiction 2024, 120, 138–151. [Google Scholar] [CrossRef]
- Olson, K.M.; Lei, W.; Keresztes, A.; LaVigne, J.; Streicher, J.M. Novel Molecular Strategies and Targets for Opioid Drug Discovery for the Treatment of Chronic Pain. Yale J. Biol. Med. 2017, 90, 97–110. [Google Scholar]
- Tamburello, A.; Martin, T.L. Dosing and Misuse of Buprenorphine in the New Jersey Department of Corrections. J. Am. Acad. Psychiatry Law. 2024, 52, 441–448. [Google Scholar]
- Blackwood, C.A.; Hoerle, R.; Leary, M.; Schroeder, J.; Job, M.O.; McCoy, M.T.; Ladenheim, B.; Jayanthi, S.; Cadet, J.L. Molecular Adaptations in the Rat Dorsal Striatum and Hippocampus Following Abstinence-Induced Incubation of Drug Seeking After Escalated Oxycodone Self-Administration. Mol. Neurobiol. 2019, 56, 3603–3615. [Google Scholar] [CrossRef]
- Salisbury, A.J.; Blackwood, C.A.; Cadet, J.L. Prolonged Withdrawal From Escalated Oxycodone Is Associated With Increased Expression of Glutamate Receptors in the Rat Hippocampus. Front. Neurosci. 2020, 14, 617973. [Google Scholar] [CrossRef]
- Vassoler, F.M.; Budge, K.E.; Isgate, S.B.; Gildawie, K.R.; Byrnes, E.M. Neuroplasticity-related genes correlate with individual differences in distinct phases of oxycodone self-administration in male rats. Neuropharmacology 2024, 254, 109972. [Google Scholar] [CrossRef]
- Mantsch, J.R.; Yuferov, V.; Mathieu-Kia, A.M.; Ho, A.; Kreek, M.J. Effects of extended access to high versus low cocaine doses on self-administration, cocaine-induced reinstatement and brain mRNA levels in rats. Psychopharmacology 2004, 175, 26–36. [Google Scholar] [CrossRef]
- Ahmed, S.H.; Koob, G.F. Long-lasting increase in the set point for cocaine self-administration after escalation in rats. Psychopharmacology 1999, 146, 303–312. [Google Scholar] [CrossRef]
- Illenberger, J.M.; Flores-Ramirez, F.J.; Pascasio, G.; Franco, M.; Mendonsa, B.; Martin-Fardon, R. Pivotal role of orexin signaling in the posterior paraventricular nucleus of the thalamus during the stress-induced reinstatement of oxycodone-seeking behavior. J. Psychopharmacol. 2024, 38, 647–660. [Google Scholar] [CrossRef]
- Nguyen, J.D.; Grant, Y.; Taffe, M.A. Paradoxical changes in brain reward status during oxycodone self-administration in a novel test of the negative reinforcement hypothesis. Br. J. Pharmacol. 2021, 178, 3797–3812. [Google Scholar] [CrossRef] [PubMed]
- Samson, K.R.; Xu, W.; Kortagere, S.; España, R.A. Intermittent access to oxycodone decreases dopamine uptake in the nucleus accumbens core during abstinence. Addict. Biol. 2022, 27, e13241. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Duan, Y.; Fredriksson, I.; Tsai, P.J.; Batista, A.; Lu, H.; Shaham, Y.; Yang, Y. Role of dorsal striatum circuits in relapse to opioid seeking after voluntary abstinence. Neuropsychopharmacology 2024, 50, 452–460. [Google Scholar] [CrossRef] [PubMed]
- Malvaez, M.; Greenfield, V.Y.; Matheos, D.P.; Angelillis, N.A.; Murphy, M.D.; Kennedy, P.J.; Wood, M.A.; Wassum, K.M. Habits Are Negatively Regulated by Histone Deacetylase 3 in the Dorsal Striatum. Biol. Psychiatry 2018, 84, 383–392. [Google Scholar] [CrossRef]
- Phan, B.N.; Ray, M.H.; Xue, X.; Fu, C.; Fenster, R.J.; Kohut, S.J.; Bergman, J.; Haber, S.N.; McCullough, K.M.; Fish, M.K.; et al. Single nuclei transcriptomics in human and non-human primate striatum in opioid use disorder. Nat. Commun. 2024, 15, 878. [Google Scholar] [CrossRef]
- Gujar, V.V.; Daiwile, A.P.; Palande, V.; Cadet, J.L. RNA sequencing analysis identifies sex differences in transcriptional signatures in the dorsal striatum of female and male rats after withdrawal from methamphetamine self-administration. Neurochem. Int. 2025, 187, 105980. [Google Scholar] [CrossRef]
- Daws, S.E.; Whittard, J.D.; Jacobs, M.M.; Ren, Y.; Mazloom, A.R.; Caputi, F.F.; Horvath, M.; Keller, E.; Ma’ayan, A.; Pan, Y.-X.; et al. ELK1 transcription factor linked to dysregulated striatal mu opioid receptor signaling network and OPRM1 polymorphism in human heroin abusers. Biol. Psychiatry 2013, 74, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Elhadi, K.; Daiwile, A.P.; Cadet, J.L. Modeling methamphetamine use disorder and relapse in animals: Short- and long-term epigenetic, transcriptional., and biochemical consequences in the rat brain. Neurosci. Biobehav. Rev. 2023, 155, 105440. [Google Scholar] [CrossRef]
- McCoy, M.T.; Jayanthi, S.; Cadet, J.L. Potassium Channels and Their Potential Roles in Substance Use Disorders. Int. J. Mol. Sci. 2021, 22, 1249. [Google Scholar] [CrossRef]
- Antsiferova, M.; Berrera, M.; Zagdoun, A.-C.; Raauf, M.; Nguyen, T.T.; Murgia, C.; Appelt, B.; Trumpfheller, C.; Gasser, S.; Pilet, S.; et al. Novel immunodominant neoepitope in a KPC mouse model of pancreatic cancer allowing identification of tumor-specific T cells. Oncoimmunology 2025, 14, 2489815. [Google Scholar] [CrossRef]
- Jan, L.Y.; Jan, Y.N. Voltage-gated potassium channels and the diversity of electrical signalling. J. Physiol. 2012, 590, 2591–2599. [Google Scholar] [CrossRef]
- Hodgkin, A.L.; Huxley, A.F. Currents carried by sodium and potassium ions through the membrane of the giant axon of Loligo. J. Physiol. 1952, 116, 449–472. [Google Scholar] [CrossRef]
- Holly, E.N.; Davatolhagh, M.F.; Choi, K.; Alabi, O.O.; Vargas Cifuentes, L.; Fuccillo, M.V. Striatal Low-Threshold Spiking Interneurons Regulate Goal-Directed Learning. Neuron 2019, 103, 92–101.e6. [Google Scholar] [CrossRef] [PubMed]
- Hogarth, L.; Balleine, B.W.; Corbit, L.H.; Killcross, S. Associative learning mechanisms underpinning the transition from recreational drug use to addiction. Ann. N. Y. Acad. Sci. 2013, 1282, 12–24. [Google Scholar] [CrossRef]
- Malvaez, M.; Wassum, K.M. Regulation of habit formation in the dorsal striatum. Curr. Opin. Behav. Sci. 2018, 20, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Blackwood, C.A.; McCoy, M.T.; Ladenheim, B.; Cadet, J.L. Escalated Oxycodone Self-Administration and Punishment: Differential Expression of Opioid Receptors and Immediate Early Genes in the Rat Dorsal Striatum and Prefrontal Cortex. Front. Neurosci. 2019, 13, 1392. [Google Scholar] [CrossRef]
- Daiwile, A.P.; Jayanthi, S.; Ladenheim, B.; McCoy, M.T.; Brannock, C.; Schroeder, J.; Cadet, J.L. Sex Differences in Escalated Methamphetamine Self-Administration and Altered Gene Expression Associated With Incubation of Methamphetamine Seeking. Int. J. Neuropsychopharmacol. 2019, 22, 710–723. [Google Scholar] [CrossRef] [PubMed]
- Daiwile, A.P.; Jayanthi, S.; Cadet, J.L. Sex- and Brain Region-specific Changes in Gene Expression in Male and Female Rats as Consequences of Methamphetamine Self-administration and Abstinence. Neuroscience 2021, 452, 265–279. [Google Scholar] [CrossRef]
- Daiwile, A.P.; Sullivan, P.; Jayanthi, S.; Goldstein, D.S.; Cadet, J.L. Sex-Specific Alterations in Dopamine Metabolism in the Brain after Methamphetamine Self-Administration. Int. J. Mol. Sci. 2022, 23, 4353. [Google Scholar] [CrossRef] [PubMed]
- de Guglielmo, G.; Carrette, L.; Kallupi, M.; Brennan, M.; Boomhower, B.; Maturin, L.; Conlisk, D.; Sedighim, S.; Tieu, L.; Fannon, M.J.; et al. Large-scale characterization of cocaine addiction-like behaviors reveals that escalation of intake, aversion-resistant responding, and breaking-points are highly correlated measures of the same construct. Elife 2024, 12, RP90422. [Google Scholar] [CrossRef]
- Lobo, M.K.; Nestler, E.J. The striatal balancing act in drug addiction: Distinct roles of direct and indirect pathway medium spiny neurons. Front. Neuroanat. 2011, 5, 41. [Google Scholar] [CrossRef]
- van Huijstee, A.N.; Mansvelder, H.D. Glutamatergic synaptic plasticity in the mesocorticolimbic system in addiction. Front. Cell. Neurosci. 2014, 8, 466. [Google Scholar] [CrossRef]
- Yager, L.M.; Garcia, A.F.; Wunsch, A.M.; Ferguson, S.M. The ins and outs of the striatum: Role in drug addiction. Neuroscience 2015, 301, 529–541. [Google Scholar] [CrossRef]
- Volkow, N.D.; Michaelides, M.; Baler, R. The Neuroscience of Drug Reward and Addiction. Physiol. Rev. 2019, 99, 2115–2140. [Google Scholar] [CrossRef]
- Ciccocioppo, R.; Sanna, P.P.; Weiss, F. Cocaine-predictive stimulus induces drug-seeking behavior and neural activation in limbic brain regions after multiple months of abstinence: Reversal by D(1) antagonists. Proc. Natl. Acad. Sci. USA 2001, 98, 1976–1981. [Google Scholar] [CrossRef]
- Joho, R.H.; Hurlock, E.C. The role of Kv3-type potassium channels in cerebellar physiology and behavior. Cerebellum 2009, 8, 323–333. [Google Scholar] [CrossRef] [PubMed]
- Llinás, R.R. The intrinsic electrophysiological properties of mammalian neurons: Insights into central nervous system function. Science 1988, 242, 1654–1664. [Google Scholar] [CrossRef] [PubMed]
- Noh, W.; Pak, S.; Choi, G.; Yang, S.; Yang, S. Transient Potassium Channels: Therapeutic Targets for Brain Disorders. Front. Cell. Neurosci. 2019, 13, 265. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Fernández, D.; Lamas, J.A. Metabotropic Modulation of Potassium Channels During Synaptic Plasticity. Neuroscience 2021, 456, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Hoffman, D.A. Potassium channels: Newly found players in synaptic plasticity. Neuroscientist 2008, 14, 276–286. [Google Scholar] [CrossRef]
- Bausch, A.E.; Dieter, R.; Nann, Y.; Hausmann, M.; Meyerdierks, N.; Kaczmarek, L.K.; Ruth, P.; Lukowski, R. The sodium-activated potassium channel Slack is required for optimal cognitive flexibility in mice. Learn. Mem. 2015, 22, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Parakramaweera, R.; Teng, S.; Gowda, M.; Sharad, Y.; Thakker-Varia, S.; Alder, J.; Sesti, F. Oxidation of KCNB1 Potassium Channels Causes Neurotoxicity and Cognitive Impairment in a Mouse Model of Traumatic Brain Injury. J. Neurosci. 2016, 36, 11084–11096. [Google Scholar] [CrossRef]
- Yu, W.; Zhang, H.; Shin, M.R.; Sesti, F. Oxidation of KCNB1 potassium channels in the murine brain during aging is associated with cognitive impairment. Biochem. Biophys. Res. Commun. 2019, 512, 665–669. [Google Scholar] [CrossRef]
- Alam, K.A.; Svalastoga, P.; Martinez, A.; Glennon, J.C.; Haavik, J. Potassium channels in behavioral brain disorders. Molecular mechanisms and therapeutic potential: A narrative review. Neurosci. Biobehav. Rev. 2023, 152, 105301. [Google Scholar] [CrossRef]
- Blum, K.; Chen, A.L.; Thanos, P.K.; Febo, M.; Demetrovics, Z.; Dushaj, K.; Kovoor, A.; Baron, D.; Smith, D.E.; Roy, A.K., III; et al. Genetic addiction risk score (GARS)™, a predictor of vulnerability to opioid dependence. Front. Biosci. (Elite Ed.) 2018, 10, 175–196. [Google Scholar] [CrossRef]
- Cloninger, C.R. Neurogenetic adaptive mechanisms in alcoholism. Science 1987, 236, 410–416. [Google Scholar] [CrossRef]
- Kashani, J.H.; Solomon, N.A.; Dugan, K.; Joy, F. Differences between early and late onset of substance abuse: An inpatient experience. South. Med. J. 1987, 80, 554–557. [Google Scholar] [CrossRef]
- Ball, S.A.; Carroll, K.M.; Babor, T.F.; Rounsaville, B.J. Subtypes of cocaine abusers: Support for a type A-type B distinction. J. Consult. Clin. Psychol. 1995, 63, 115–124. [Google Scholar] [CrossRef]
- Debeck, K.; Kerr, T.; Marshall, B.D.; Simo, A.; Montaner, J.; Wood, E. Risk factors for progression to regular injection drug use among street-involved youth in a Canadian setting. Drug Alcohol. Depend. 2013, 133, 468–472. [Google Scholar] [CrossRef]
- Drossel, G.; Brucar, L.R.; Rawls, E.; Hendrickson, T.J.; Zilverstand, A. Subtypes in addiction and their neurobehavioral profiles across three functional domains. Transl. Psychiatry 2023, 13, 127. [Google Scholar] [CrossRef]
- Kelly, T.H.; Robbins, G.; Martin, C.A.; Fillmore, M.T.; Lane, S.D.; Harrington, N.G.; Rush, C.R. Individual differences in drug abuse vulnerability: D-amphetamine and sensation-seeking status. Psychopharmacology 2006, 189, 17–25. [Google Scholar] [CrossRef]
- Gipson, C.D.; Beckmann, J.S.; Adams, Z.W.; Marusich, J.A.; Nesland, T.O.; Yates, J.R.; Kelly, T.H.; Bardo, M.T. A translational behavioral model of mood-based impulsivity: Implications for substance abuse. Drug Alcohol. Depend. 2012, 122, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Harvanko, A.; Martin, C.; Lile, J.; Kryscio, R.; Kelly, T.H. Individual differences in the reinforcing and subjective effects of d-amphetamine: Dimensions of impulsivity. Exp. Clin. Psychopharmacol. 2016, 24, 436–446. [Google Scholar] [CrossRef] [PubMed]
- Joseph, J.E.; Zhu, X.; Corbly, C.R.; DeSantis, S.; Lee, D.C.; Baik, G.; Kiser, S.; Jiang, Y.; Lynam, D.R.; Kelly, T.H. Influence of neurobehavioral incentive valence and magnitude on alcohol drinking behavior. Neuroimage 2015, 104, 373–385. [Google Scholar] [CrossRef] [PubMed]
- Daiwile, A.P.; McCoy, M.T.; Ladenheim, B.; Subramaniam, J.; Cadet, J.L. Incubation of methamphetamine craving in punishment-resistant individuals is associated with activation of specific gene networks in the rat dorsal striatum. Mol. Psychiatry 2024, 29, 1990–2000. [Google Scholar] [CrossRef]
- González, C.; Baez-Nieto, D.; Valencia, I.; Oyarzún, I.; Rojas, P.; Naranjo, D.; Latorre, R. K(+) channels: Function-structural overview. Compr. Physiol. 2012, 2, 2087–2149. [Google Scholar] [CrossRef]
- Liu, M.; Si, Z. An update: Epigenetic mechanisms underlying methamphetamine addiction. Front. Cell Dev. Biol. 2024, 12, 1494557. [Google Scholar] [CrossRef] [PubMed]
- Linden, A.M.; Sandu, C.; Aller, M.I.; Vekovischeva, O.Y.; Rosenberg, P.H.; Wisden, W.; Korpi, E.R. TASK-3 knockout mice exhibit exaggerated nocturnal activity, impairments in cognitive functions, and reduced sensitivity to inhalation anesthetics. J. Pharmacol. Exp. Ther. 2007, 323, 924–934. [Google Scholar] [CrossRef]
- Shah, N.H.; Aizenman, E. Voltage-gated potassium channels at the crossroads of neuronal function, ischemic tolerance, and neurodegeneration. Transl. Stroke Res. 2014, 5, 38–58. [Google Scholar] [CrossRef]
- Pollini, L.; Galosi, S.; Tolve, M.; Caputi, C.; Carducci, C.; Angeloni, A.; Leuzzi, V. KCND3-Related Neurological Disorders: From Old to Emerging Clinical Phenotypes. Int. J. Mol. Sci. 2020, 21, 5802. [Google Scholar] [CrossRef]
- Cadet, J.L.; Brannock, C.; Krasnova, I.N.; Jayanthi, S.; Ladenheim, B.; McCoy, M.T.; Walther, D.; Godino, A.; Pirooznia, M.; Lee, R.S. Genome-wide DNA hydroxymethylation identifies potassium channels in the nucleus accumbens as discriminators of methamphetamine addiction and abstinence. Mol. Psychiatry 2017, 22, 1196–1204. [Google Scholar] [CrossRef]
- Lin, M.A.; Cannon, S.C.; Papazian, D.M. Kv4.2 autism and epilepsy mutation enhances inactivation of closed channels but impairs access to inactivated state after opening. Proc. Natl. Acad. Sci. USA 2018, 115, E3559–E3568. [Google Scholar] [CrossRef]
- Rui, Y.; Liu, S.; Liu, S. Kir4.1 channel and voltage-gated calcium channel of astrocyte account for the transition dynamics of seizures. J. Theor. Biol. 2025, 604, 112082. [Google Scholar] [CrossRef]
- Gilron, I.; Dickenson, A.H. Emerging drugs for neuropathic pain. Expert. Opin. Emerg. Drugs 2014, 19, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Krueger, S.K.; Williams, D.E. Mammalian flavin-containing monooxygenases: Structure/function, genetic polymorphisms and role in drug metabolism. Pharmacol. Ther. 2005, 106, 357–387. [Google Scholar] [CrossRef] [PubMed]
- Perez-Paramo, Y.X.; Lazarus, P. Pharmacogenetics factors influencing smoking cessation success; the importance of nicotine metabolism. Expert. Opin. Drug Metab. Toxicol. 2021, 17, 333–349. [Google Scholar] [CrossRef]
- Krasnova, I.N.; Gerra, M.C.; Walther, D.; Jayanthi, S.; Ladenheim, B.; McCoy, M.T.; Brannock, C.; Cadet, J.L. Compulsive methamphetamine taking in the presence of punishment is associated with increased oxytocin expression in the nucleus accumbens of rats. Sci. Rep. 2017, 7, 8331. [Google Scholar] [CrossRef]
- Cadet, J.L.; Brannock, C.; Ladenheim, B.; McCoy, M.T.; Beauvais, G.; Hodges, A.B.; Lehrmann, E.; Wood, W.H., 3rd; Becker, K.G.; Krasnova, I.N. Methamphetamine preconditioning causes differential changes in striatal transcriptional responses to large doses of the drug. Dose Response 2011, 9, 165–181. [Google Scholar] [CrossRef]
- Crittenden, J.R.; Gipson, T.A.; Smith, A.C.; Bowden, H.A.; Yildirim, F.; Fischer, K.B.; Yim, M.; Housman, D.E.; Graybiel, A.M. Striatal transcriptome changes linked to drug-induced repetitive behaviors. Eur. J. Neurosci. 2021, 53, 2450–2468. [Google Scholar] [CrossRef]
- Browne, C.J.; Futamura, R.; Minier-Toribio, A.; Hicks, E.M.; Ramakrishnan, A.; Martínez-Rivera, F.J.; Estill, M.; Godino, A.; Parise, E.M.; Torres-Berrío, A.; et al. Transcriptional signatures of heroin intake and relapse throughout the brain reward circuitry in male mice. Sci. Adv. 2023, 9, eadg8558. [Google Scholar] [CrossRef]
- Fang, C.P.; Liu, T.H.; Chung, R.H.; Tsou, H.H.; Kuo, H.W.; Wang, S.C.; Liu, C.C.; Liu, S.C.; Chen, A.C.H.; Liu, Y.L.; et al. Genetic variants in NECTIN4 encoding an adhesion molecule are associated with continued opioid use. PLoS ONE 2020, 15, e0234549. [Google Scholar] [CrossRef]
- Liu, Y.L.; Kuo, H.W.; Fang, C.P.; Tsung, J.H.; Chen, A.C.H. Adhesion Molecules as Potential Novel Biomarkers for Opioid Dependence. Curr. Pharm. Des. 2020, 26, 253–259. [Google Scholar] [CrossRef]
- Chang, H.M.; Chen, P.Y.; Fang, C.P.; Liu, T.H.; Wu, C.T.; Hsu, Y.C.; Kuo, H.W.; Liu, Y.L.; Huang, M.C. Increased Nectin-4 levels in chronic ketamine abusers and the relationship with lower urinary tract symptoms. Environ. Toxicol. Pharmacol. 2021, 87, 103714. [Google Scholar] [CrossRef]
- McCoy, M.T.; Daiwile, A.P.; Ladenheim, B.; Cadet, J.L. Sex-specific Alterations in the mRNA Expression of Histone Deacetylases (HDACs) in the Rat Brain Following Prolonged Abstinence from Methamphetamine Self-administration. Mol. Neurobiol. 2025, 62, 9759–9771. [Google Scholar] [CrossRef] [PubMed]
- Krasnova, I.N.; Marchant, N.J.; Ladenheim, B.; McCoy, M.T.; Panlilio, L.V.; Bossert, J.M.; Shaham, Y.; Cadet, J.L. Incubation of methamphetamine and palatable food craving after punishment-induced abstinence. Neuropsychopharmacology 2014, 39, 2008–2016. [Google Scholar] [CrossRef] [PubMed]
- Tella, S.R.; Ladenheim, B.; Andrews, A.M.; Goldberg, S.R.; Cadet, J.L. Differential reinforcing effects of cocaine and GBR-12909: Biochemical evidence for divergent neuroadaptive changes in the mesolimbic dopaminergic system. J. Neurosci. 1996, 16, 7416–7427. [Google Scholar] [CrossRef] [PubMed]
- Guha, S.K.; Alonso-Caraballo, Y.; Driscoll, G.S.; Babb, J.A.; Neal, M.; Constantino, N.J.; Lintz, T.; Kinard, E.; Chartoff, E.H. Ranking the contribution of behavioral measures comprising oxycodone self-administration to reinstatement of drug-seeking in male and female rats. Front. Behav. Neurosci. 2022, 16, 1035350. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wabreha, A.Y.; McCoy, M.T.; Cadet, J.L.; Daiwile, A.P. Escalated Oxycodone Self-Administration Is Associated with Activation of Specific Gene Networks in the Rat Dorsal Striatum. Int. J. Mol. Sci. 2025, 26, 7356. https://doi.org/10.3390/ijms26157356
Wabreha AY, McCoy MT, Cadet JL, Daiwile AP. Escalated Oxycodone Self-Administration Is Associated with Activation of Specific Gene Networks in the Rat Dorsal Striatum. International Journal of Molecular Sciences. 2025; 26(15):7356. https://doi.org/10.3390/ijms26157356
Chicago/Turabian StyleWabreha, Ammanuel Y., Michael T. McCoy, Jean Lud Cadet, and Atul P. Daiwile. 2025. "Escalated Oxycodone Self-Administration Is Associated with Activation of Specific Gene Networks in the Rat Dorsal Striatum" International Journal of Molecular Sciences 26, no. 15: 7356. https://doi.org/10.3390/ijms26157356
APA StyleWabreha, A. Y., McCoy, M. T., Cadet, J. L., & Daiwile, A. P. (2025). Escalated Oxycodone Self-Administration Is Associated with Activation of Specific Gene Networks in the Rat Dorsal Striatum. International Journal of Molecular Sciences, 26(15), 7356. https://doi.org/10.3390/ijms26157356