
Nuclear Receptors in Bladder Cancer: Insights into miRNA-Mediated Regulation and Potential Therapeutic Implications

 ,
,  , ,
, ,  , , and
, , and 
Abstract

1. Introduction
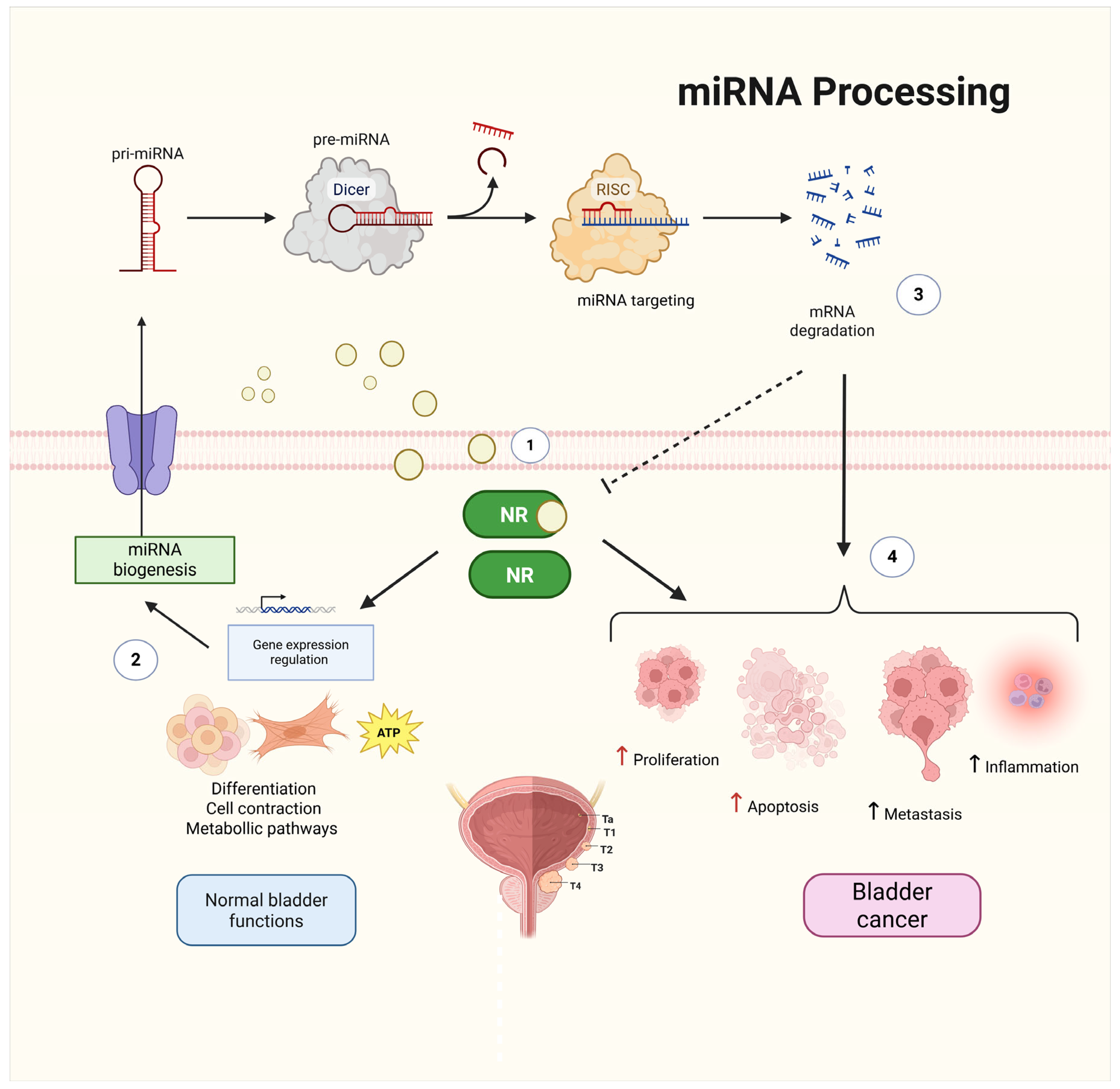
2. Nuclear Receptors and miRNAs as Key Regulators in BCa

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA (Expression) | Target/Regulation | Effect on BCa | References |
|---|---|---|---|
| miR-23a, miR-141b, miR-205 (↑) | Repress ZEB2, PTEN, and E-cadherin, respectively | Promote tumor invasion and progression | [31] |
| miR-525-5p, miR-144 (↑) | Repress SLPI; upregulate GRβ (Glucocorticoid Receptor β) | Promote metástasis | [52,53] |
| miR-92a, miR-19a, miR-130 (↑) | Repress DAB2IP, PTEN, RUNX3 | Promote tumor progression | [31,38] |
| miR-21, miR-221/222, miR-125b, miR-133b (↑) | Repress PTEN, PDCD4, p27Kip1, p57, E2F3; downregulate p-PI3K, p-AKT; upregulate AKT1 | Promote tumor growth and proliferation | [44,46,54] |
| miR-193a-3p (↑) | Repress Homeobox C9 (HOXC9) gene | Promote multidrug resistance | [55] |
| miR-29c (↓) | Regulates CDK6 | Regulate cell growth and invasion in vitro | [56] |
| miRNA (Expression) | Target/Regulation | Effect on BCa | References |
|---|---|---|---|
| miR-34a, miR-145 (↓) | Repress CD44, PD-L1 | Decrease adhesion, invasion, and immune evasion | [36,40,41] |
| miR-125a, miR-125b (↓) | Targets HK2 suppressing PI3K-Akt, targets FUT4 | Decreases migration, invasion, progression, and modulates apoptosis | [46,57] |
| miR-449a (↑) | Promotes AR degradation, targets CDK6 and CDC25a, and activates accumulation of the pocket proteins Rb and p130 | Decreases cell proliferation | [58,59] |
| miR-490-5p, miR-139 (↓) | Represses EGFR and MMP11expression | Reduces invasiveness | [60,61,65] |
| miR-101 (↓) | Target genes regulating the cell cycle | Decreases proliferation | [50] |
| miR-203a (↓) | Targets SIX4 | Decreases tumor growth | [51] |
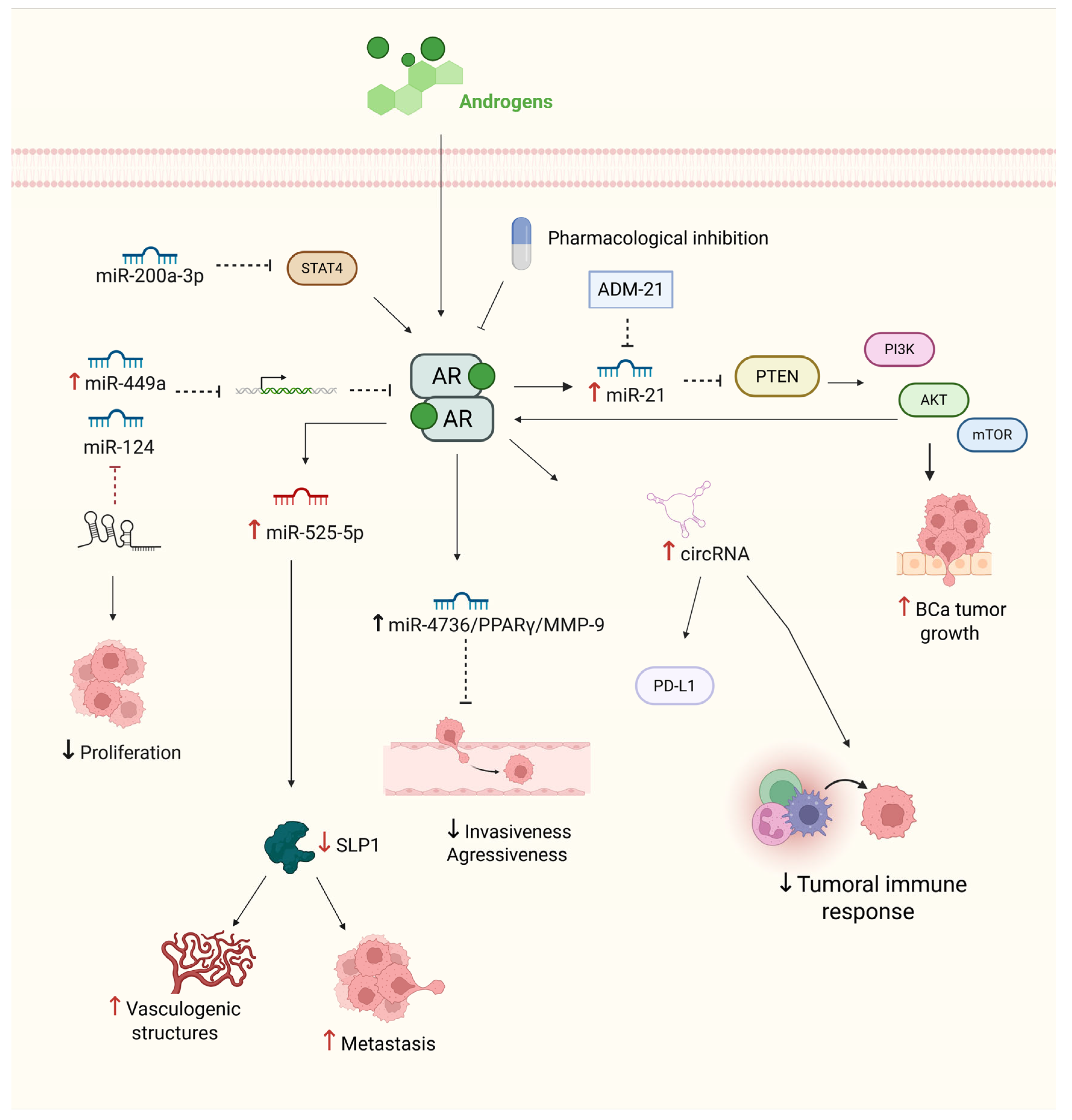
3. Androgen Receptor Signaling and Its Regulation by Non-Coding RNAs in BCa
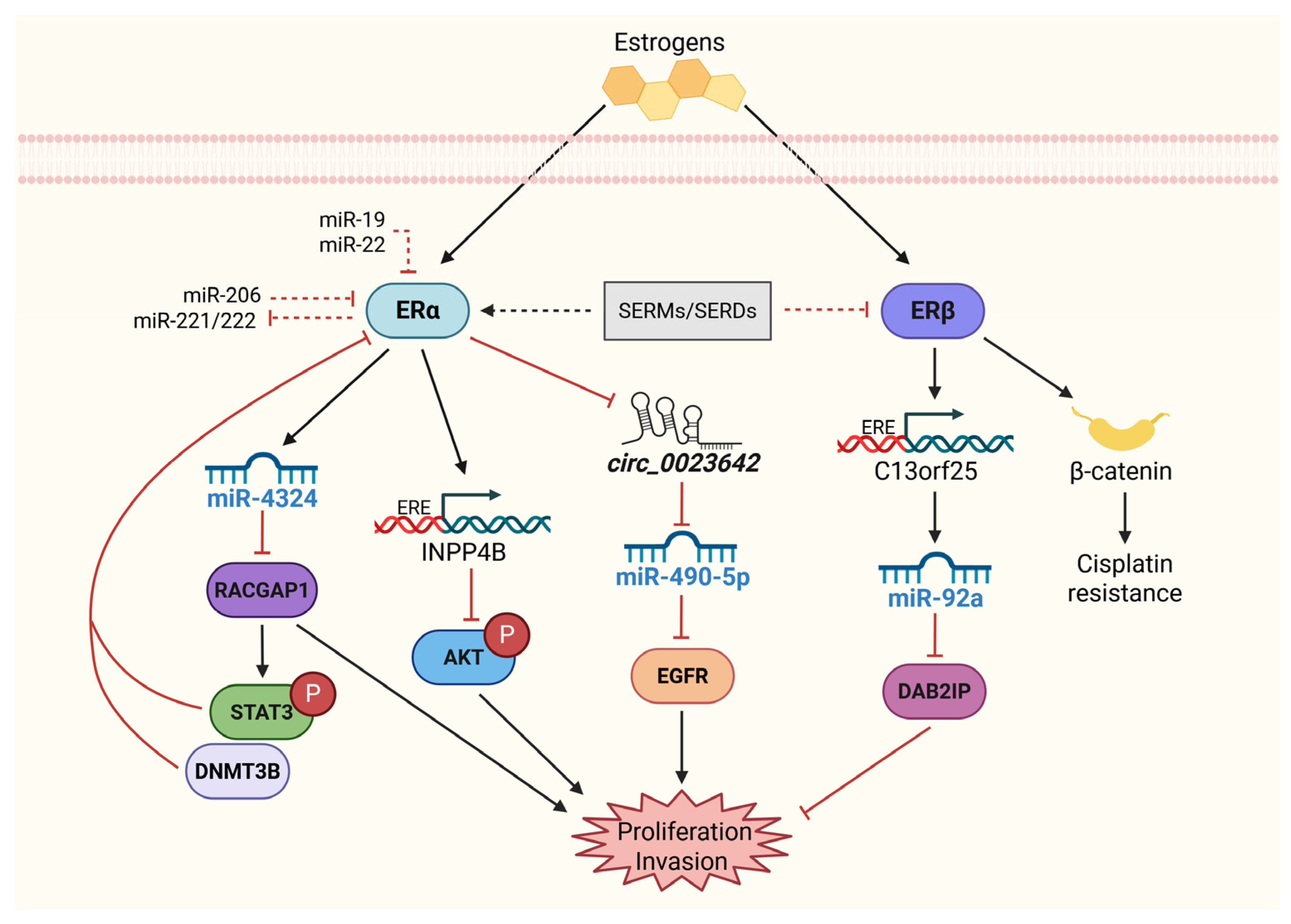
4. Molecular Roles of Estrogen Receptors in BCa: From Transcriptional Control to miRNA-Driven Networks
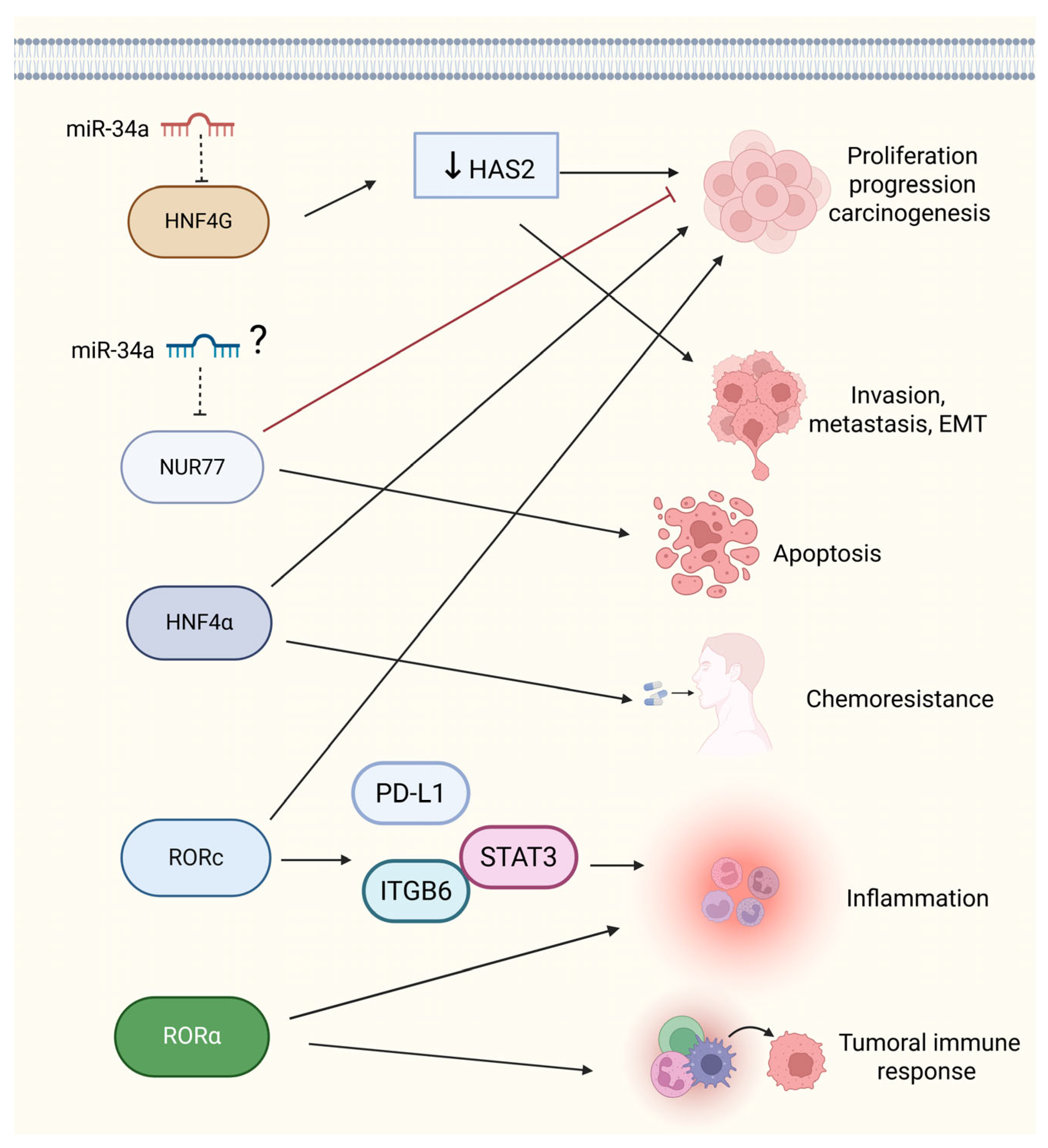
5. Molecular Functions of Orphan Nuclear Receptors in BCa: miRNA Regulatory Networks and Cancer Development and Progression
6. PPARγ and the Tumor Microenvironment in BCa: Molecular Regulation
7. Glucocorticoid Receptor Signaling in BCa: Isoform-Specific Roles and miRNA Interactions
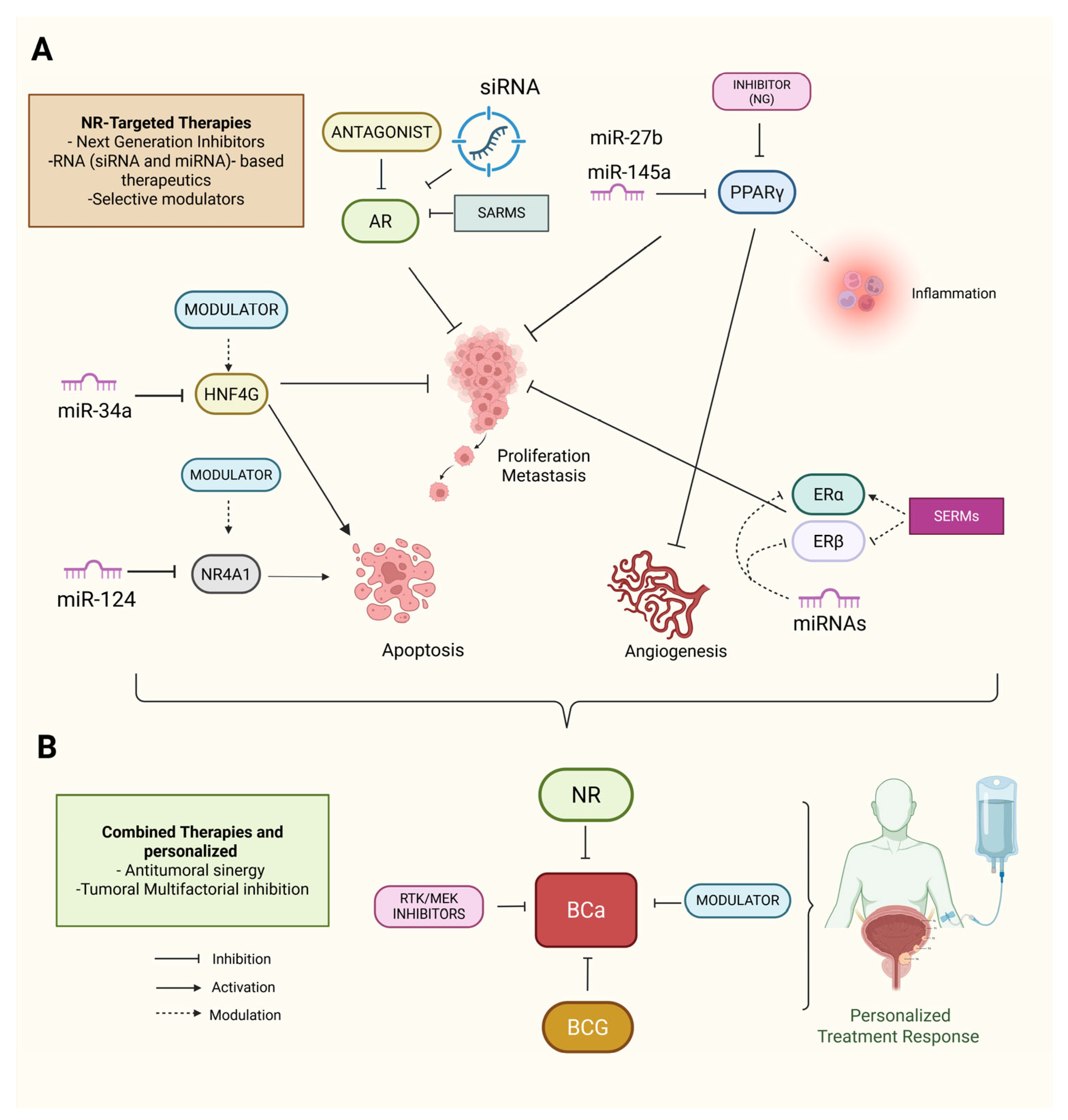
8. NR-Targeted and Therapeutic Implications in BCa
RNA-Based Therapeutics and Combination with miRNA-Based Therapies
9. Combination Therapies and Personalized Approaches
10. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BCa | Bladder cancer |
| NR | Nuclear receptor |
| BCG | Bacillus Calmette–Guérin |
| miRNAs | microRNAs |
| mRNAs | Messenger RNAs |
| PCa | Prostate cancer |
| MIBCa | Muscle-invasive BCa |
| NMIBCa | Non-muscle-invasive BCa |
| AR | Androgen receptor |
| AREs | Androgen response elements |
| UTR | Untranslated region |
| EMT | Epithelial-to-mesenchymal transition |
| ER | Estrogen receptors |
| PPARs | Peroxisome proliferator-activated receptors |
| ONRs | Orphan nuclear receptors |
| GCRs | Glucocorticoid receptors |
| VDR | Vitamin D Receptor |
References
- Dyrskjøt, L.; Hansel, D.E.; Efstathiou, J.A.; Knowles, M.A.; Galsky, M.D.; Teoh, J.; Theodorescu, D. Bladder cancer. Nat. Rev. Dis. Primers 2023, 9, 58. [Google Scholar] [CrossRef] [PubMed]
- Gopalakrishna, A.; Longo, T.A.; Fantony, J.J.; Van Noord, M.; Inman, B.A. Lifestyle factors and health-related quality of life in bladder cancer survivors: A systematic review. J. Cancer Surviv. 2016, 10, 874–882. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Cancer statistics for the year 2020: An overview. Int. J. Cancer 2021, 149, 778–789. [Google Scholar] [CrossRef]
- Kohada, Y.; Hayashi, T.; Hsi, R.S.; Yukihiro, K.; Sentani, K.; Goto, K.; Inoue, S.; Ohara, S.; Teishima, J.; Kajiwara, M.; et al. Recurrence- and progression-free survival in intermediate-risk non-muscle-invasive bladder cancer: The impact of conditional evaluation and subclassification. BJU Int. 2021, 127, 473–485. [Google Scholar] [CrossRef]
- Zuniga, K.B.; Graff, R.E.; Feiger, D.B.; Meng, M.V.; Porten, S.P.; Kenfield, S.A. Lifestyle and Non-muscle Invasive Bladder Cancer Recurrence, Progression, and Mortality: Available Research and Future Directions. Bladder Cancer 2020, 6, 9–23. [Google Scholar] [CrossRef]
- Lee, C.U.; Song, W.; Koo, M.J.; Boo, Y.; Chung, J.H.; Kang, M.; Sung, H.H.; Jeon, H.G.; Jeong, B.C.; Seo, S.I.; et al. Utility of Contemporary Health Screening in the Diagnosis of Bladder Cancer. Diagnostics 2022, 12, 1040. [Google Scholar] [CrossRef]
- Yun, S.J.; Kim, S.K.; Kim, W.J. How do we manage high-grade T1 bladder cancer? Conservative or aggressive therapy? Investig. Clin. Urol. 2016, 57 (Suppl. S1), S44-51. [Google Scholar] [CrossRef]
- Massari, F.; Santoni, M.; Ciccarese, C.; Brunelli, M.; Conti, A.; Santini, D.; Montironi, R.; Cascinu, S.; Tortora, G. Emerging concepts on drug resistance in bladder cancer: Implications for future strategies. Crit. Rev. Oncol. Hematol. 2015, 96, 81–90. [Google Scholar] [CrossRef]
- Moras, D.; Gronemeyer, H. The nuclear receptor ligand-binding domain: Structure and function. Curr. Opin. Cell Biol. 1998, 10, 384–391. [Google Scholar] [CrossRef]
- Steinmetz, A.C.; Renaud, J.P.; Moras, D. Binding of ligands and activation of transcription by nuclear receptors. Annu. Rev. Biophys. Biomol. Struct. 2001, 30, 329–359. [Google Scholar] [CrossRef]
- Yang, Z.; Gimple, R.C.; Zhou, N.; Zhao, L.; Gustafsson, J.; Zhou, S. Targeting Nuclear Receptors for Cancer Therapy: Premises, Promises, and Challenges. Trends Cancer 2021, 7, 541–556. [Google Scholar] [CrossRef]
- El-Mahdy, H.A.; Elsakka, E.G.E.; El-Husseiny, A.A.; Ismail, A.; Yehia, A.M.; Abdelmaksoud, N.M.; Elshimy, R.A.A.; Noshy, M.; Doghish, A.S. miRNAs role in bladder cancer pathogenesis and targeted therapy: Signaling pathways interplay—A review. Pathol. Res. Pract. 2023, 242, 154316. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, G.; Guo, X.; Yao, H.; Wang, G.; Li, C. Non-coding RNA in bladder cancer. Cancer Lett. 2020, 485, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.; Chen, Y.; Meyer, C.; Geistlinger, T.; Lupien, M.; Wang, Q.; Liu, T.; Zhang, Y.; Brown, M.; Liu, X.S. A comprehensive view of nuclear receptor cancer cistromes. Cancer Res. 2011, 71, 6940–6947. [Google Scholar] [CrossRef] [PubMed]
- Dhiman, V.K.; Bolt, M.J.; White, K.P. Nuclear receptors in cancer—Uncovering new and evolving roles through genomic analysis. Nat. Rev. Genet. 2018, 19, 160–174. [Google Scholar] [CrossRef]
- Nakata, K.; Tanaka, Y.; Nakano, T.; Adachi, T.; Tanaka, H.; Kaminuma, T.; Ishikawa, T. Nuclear receptor-mediated transcriptional regulation in Phase I, II, and III xenobiotic metabolizing systems. Drug Metab. Pharmacokinet. 2006, 21, 437–457. [Google Scholar] [CrossRef]
- Vienberg, S.; Geiger, J.; Madsen, S.; Dalgaard, L.T. MicroRNAs in metabolism. Acta Physiol. 2017, 219, 346–361. [Google Scholar] [CrossRef]
- Chekulaeva, M.; Filipowicz, W. Mechanisms of miRNA-mediated post-transcriptional regulation in animal cells. Curr. Opin. Cell Biol. 2009, 21, 452–460. [Google Scholar] [CrossRef]
- Braicu, C.; Cojocneanu-Petric, R.; Chira, S.; Truta, A.; Floares, A.; Petrut, B.; Achimas-Cadariu, P.; Berindan-Neagoe, I. Clinical and pathological implications of miRNA in bladder cancer. Int. J. Nanomed. 2015, 10, 791–800. [Google Scholar] [CrossRef]
- Oliveto, S.; Mancino, M.; Manfrini, N.; Biffo, S. Role of microRNAs in translation regulation and cancer. World J. Biol. Chem. 2017, 8, 45–56. [Google Scholar] [CrossRef]
- Macfarlane, L.A.; Murphy, P.R. MicroRNA: Biogenesis, Function and Role in Cancer. Curr. Genom. 2010, 11, 537–561. [Google Scholar] [CrossRef]
- Tiwari, A.; Mukherjee, B.; Dixit, M. MicroRNA Key to Angiogenesis Regulation: MiRNA Biology and Therapy. Curr. Cancer Drug Targets 2018, 18, 266–277. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Malviya, R. miRNAs involvement in the etiology and targeted therapy of bladder cancer: Interaction between signaling pathway. Precis. Med. Sci. 2023, 12, 206–223. [Google Scholar] [CrossRef]
- Kong, Y.W.; Cannell, I.G.; de Moor, C.H.; Hill, K.; Garside, P.G.; Hamilton, T.L.; Meijer, H.A.; Dobbyn, H.C.; Stoneley, M.; Spriggs, K.A.; et al. The mechanism of micro-RNA-mediated translation repression is determined by the promoter of the target gene. Proc. Natl. Acad. Sci. USA 2008, 105, 8866–8871. [Google Scholar] [CrossRef]
- Seiler, R.; Thalmann, G.N.; Rotzer, D.; Perren, A.; Fleischmann, A. CCND1/CyclinD1 status in metastasizing bladder cancer: A prognosticator and predictor of chemotherapeutic response. Mod. Pathol. 2014, 27, 87–95. [Google Scholar] [CrossRef]
- Polo, A.; Marchese, S.; De Petro, G.; Montella, M.; Ciliberto, G.; Budillon, A.; Costantini, S. Identifying a panel of genes/proteins/miRNAs modulated by arsenicals in bladder, prostate, kidney cancers. Sci. Rep. 2018, 8, 10395. [Google Scholar] [CrossRef]
- Catto, J.W.; Miah, S.; Owen, H.C.; Bryant, H.; Myers, K.; Dudziec, E.; Larré, S.; Milo, M.; Rehman, I.; Rosario, D.J.; et al. Distinct microRNA alterations characterize high- and low-grade bladder cancer. Cancer Res. 2009, 69, 8472–8481. [Google Scholar] [CrossRef]
- Rosenberg, E.; Baniel, J.; Spector, Y.; Faerman, A.; Meiri, E.; Aharonov, R.; Margel, D.; Goren, Y.; Nativ, O. Predicting progression of bladder urothelial carcinoma using microRNA expression. BJU Int. 2013, 112, 1027–1034. [Google Scholar] [CrossRef]
- Awadalla, A.; Abol-Enein, H.; Hamam, E.T.; Ahmed, A.E.; Khirallah, S.M.; El-Assmy, A.; Mostafa, S.A.; Babalghith, A.O.; Ali, M.; Abdel-Rahim, M.; et al. Identification of Epigenetic Interactions between miRNA and Gene Expression as Potential Prognostic Markers in Bladder Cancer. Genes 2022, 13, 1629. [Google Scholar] [CrossRef]
- Otmani, K.; Lewalle, P. Tumor Suppressor miRNA in Cancer Cells and the Tumor Microenvironment: Mechanism of Deregulation and Clinical Implications. Front. Oncol. 2021, 11, 708765. [Google Scholar] [CrossRef]
- Braicu, C.; Buiga, R.; Cojocneanu, R.; Buse, M.; Raduly, L.; Pop, L.A.; Chira, S.; Budisan, L.; Jurj, A.; Ciocan, C.; et al. Connecting the dots between different networks: miRNAs associated with bladder cancer risk and progression. J. Exp. Clin. Cancer Res. 2019, 38, 433. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Pi, X.W.; Xin, C.Q.; Gong, C.; Guo, F.; Wang, Y.; Xiong, Y. Differential expression and functions of miRNAs in bladder cancer. Oncologie 2023, 25, 1–15. [Google Scholar] [CrossRef]
- Kim, J.; Yao, F.; Xiao, Z.; Sun, Y.; Ma, L. MicroRNAs and metastasis: Small RNAs play big roles. Cancer Metastasis Rev. 2018, 37, 5–15. [Google Scholar] [CrossRef]
- Enokida, H.; Yoshino, H.; Matsushita, R.; Nakagawa, M. The role of microRNAs in bladder cancer. Investig. Clin. Urol. 2016, 57, S60–S76. [Google Scholar] [CrossRef]
- Das, S.; Hayden, J.; Sullivan, T.; Rieger-Christ, K. The Roles of miRNAs in Predicting Bladder Cancer Recurrence and Resistance to Treatment. Int. J. Mol. Sci. 2023, 24, 964. [Google Scholar] [CrossRef]
- Ding, Z.S.; He, Y.H.; Deng, Y.S.; Peng, P.X.; Wang, J.F.; Chen, X.; Zhao, P.Y.; Zhou, X.F. MicroRNA-34a inhibits bladder cancer cell migration and invasion, and upregulates PTEN expression. Oncol. Lett. 2019, 18, 5549–5554. [Google Scholar] [CrossRef]
- Lin, F.; Hu, S.; Chen, J.; Li, H.; Li, M.; Li, R.; Xu, M.; Luo, M. MiR-125b suppresses bladder Cancer cell growth and triggers apoptosis by regulating IL-6/IL-6R/STAT3 axis in vitro and in vivo. Cytokine 2025, 190, 156926. [Google Scholar] [CrossRef]
- Ou, Z.; Wang, Y.; Chen, J.; Tao, L.; Zuo, L.; Sahasrabudhe, D.; Joseph, J.; Wang, L.; Yeh, S. Estrogen receptor β promotes bladder cancer growth and invasion via alteration of miR-92a/DAB2IP signals. Exp. Mol. Med. 2018, 50, 1–11. [Google Scholar] [CrossRef]
- Du, L.; Jiang, X.; Duan, W.; Wang, R.; Wang, L.; Zheng, G.; Yan, K.; Li, J.; Zhang, X.; Pan, H.; et al. Cell-free microRNA expression signatures in urine serve as novel noninvasive biomarkers for diagnosis and recurrence prediction of bladder cancer. Oncotarget 2017, 8, 40832–40842. [Google Scholar] [CrossRef]
- Yu, G.; Yao, W.; Xiao, W.; Li, H.; Xu, H.; Lang, B. MicroRNA-34a functions as an anti-metastatic microRNA and suppresses angiogenesis in bladder cancer by directly targeting CD44. J. Exp. Clin. Cancer Res. 2014, 33, 779. [Google Scholar] [CrossRef]
- Zhu, J.; Li, Y.; Luo, Y.; Xu, J.; Liufu, H.; Tian, Z.; Huang, C.; Li, J. A Feedback Loop Formed by ATG7/Autophagy, FOXO3a/miR-145 and PD-L1 Regulates Stem-Like Properties and Invasion in Human Bladder Cancer. Cancers 2019, 11, 349. [Google Scholar] [CrossRef]
- Minami, K.; Taniguchi, K.; Sugito, N.; Kuranaga, Y.; Inamoto, T.; Takahara, K.; Takai, T.; Yoshikawa, Y.; Kiyama, S.; Akao, Y.; et al. MiR-145 negatively regulates Warburg effect by silencing KLF4 and PTBP1 in bladder cancer cells. Oncotarget 2017, 8, 33064–33077. [Google Scholar] [CrossRef]
- Zhang, H.; Jiang, M.; Liu, Q.; Han, Z.; Zhao, Y.; Ji, S. miR-145-5p inhibits the proliferation and migration of bladder cancer cells by targeting TAGLN2. Oncol. Lett. 2018, 16, 6355–6360. [Google Scholar] [CrossRef]
- Gottardo, F.; Liu, C.G.; Ferracin, M.; Calin, G.A.; Fassan, M.; Bassi, P.; Sevignani, C.; Byrne, D.; Negrini, M.; Pagano, F.; et al. Micro-RNA profiling in kidney and bladder cancers. Urol. Oncol. 2007, 25, 387–392. [Google Scholar] [CrossRef]
- Manickasamy, M.K.; Jayaprakash, S.; Girisa, S.; Kumar, A.; Lam, H.Y.; Okina, E.; Eng, H.; Alqahtani, M.S.; Abbas, M.; Sethi, G.; et al. Delineating the role of nuclear receptors in colorectal cancer, a focused review. Discov. Oncol. 2024, 15, 41. [Google Scholar] [CrossRef]
- Liu, S.; Chen, Q.; Wang, Y. MiR-125b-5p suppresses the bladder cancer progression via targeting HK2 and suppressing PI3K/AKT pathway. Hum. Cell 2020, 33, 185–194. [Google Scholar] [CrossRef]
- Huang, L.; Luo, J.; Cai, Q.; Pan, Q.; Zeng, H.; Guo, Z.; Dong, W.; Huang, J.; Lin, T. MicroRNA-125b suppresses the development of bladder cancer by targeting E2F3. Int. J. Cancer 2011, 128, 1758–1769. [Google Scholar] [CrossRef]
- Wang, F.; Wu, D.; Chen, J.; Chen, S.; He, F.; Fu, H.; Wu, Q.; Liu, S.; Li, X.; Wang, W. Long non-coding RNA HOXA-AS2 promotes the migration, invasion and stemness of bladder cancer via regulating miR-125b/Smad2 axis. Exp. Cell Res. 2019, 375, 1–10. [Google Scholar] [CrossRef]
- Xie, H.; Liao, X.; Chen, Z.; Fang, Y.; He, A.; Zhong, Y.; Gao, Q.; Xiao, H.; Li, J.; Huang, W.; et al. LncRNA MALAT1 Inhibits Apoptosis and Promotes Invasion by Antagonizing miR-125b in Bladder Cancer Cells. J. Cancer 2017, 8, 3803–3811. [Google Scholar] [CrossRef]
- Wang, K.; Ma, L.; Tang, J.; Yu, Q.; Shen, Y.; Wei, Y.; Zhu, C.; Deng, Z.; Zhang, W. LncRNA00518 promotes cell proliferation through regulating miR-101 in bladder cancer. J. Cancer 2020, 11, 1468–1477. [Google Scholar] [CrossRef]
- Na, X.Y.; Shang, X.S.; Zhao, Y.; Ren, P.P.; Hu, X.Q. MiR-203a functions as a tumor suppressor in bladder cancer by targeting SIX4. Neoplasma 2019, 66, 211–221. [Google Scholar] [CrossRef]
- Yang, Z.; Chen, J.; Xie, H.; Liu, T.; Chen, Y.; Ma, Z.; Pei, X.; Yang, W.; Li, L. Androgen receptor suppresses prostate cancer metastasis but promotes bladder cancer metastasis via differentially altering miRNA525-5p/SLPI-mediated vasculogenic mimicry formation. Cancer Lett. 2020, 473, 118–129. [Google Scholar] [CrossRef]
- McBeth, L.; Nwaneri, A.C.; Grabnar, M.; Demeter, J.; Nestor-Kalinoski, A.; Hinds, T.D. Glucocorticoid receptor beta increases migration of human bladder cancer cells. Oncotarget 2016, 7, 27313–27324. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.N.; Wang, K.F.; Xu, Z.Q.; Li, S.J.; Liu, Q.; Fu, D.H.; Wang, X.; Wu, B. MiR-133b regulates bladder cancer cell proliferation and apoptosis by targeting Bcl-w and Akt1. Cancer Cell Int. 2014, 14, 70. [Google Scholar] [CrossRef]
- Lv, L.; Li, Y.; Deng, H.; Zhang, C.; Pu, Y.; Qian, L.; Xiao, J.; Zhao, W.; Liu, Q.; Zhang, D.; et al. MiR-193a-3p promotes the multi-chemoresistance of bladder cancer by targeting the HOXC9 gene. Cancer Lett. 2015, 357, 105–113. [Google Scholar] [CrossRef]
- Zhao, X.; Li, J.; Huang, S.; Wan, X.; Luo, H.; Wu, D. MiRNA-29c regulates cell growth and invasion by targeting CDK6 in bladder cancer. Am. J. Transl. Res. 2015, 7, 1382–1389. [Google Scholar]
- Zhang, Y.; Zhang, D.; Lv, J.; Wang, S.; Zhang, Q. MiR-125a-5p suppresses bladder cancer progression through targeting FUT4. Biomed. Pharmacother. 2018, 108, 1039–1047. [Google Scholar] [CrossRef]
- Guo, J.; Hu, J.; Cao, R.; Chen, Q.; Li, K. Androgen Receptor Is Inactivated and Degraded in Bladder Cancer Cells by Phenyl Glucosamine via miR-449a Restoration. Med. Sci. Monit. 2018, 24, 2294–2301. [Google Scholar] [CrossRef]
- Chen, H.; Lin, Y.W.; Mao, Y.Q.; Wu, J.; Liu, Y.F.; Zheng, X.Y.; Xie, L.P. MicroRNA-449a acts as a tumor suppressor in human bladder cancer through the regulation of pocket proteins. Cancer Lett. 2012, 320, 40–47. [Google Scholar] [CrossRef]
- Han, Y.; Chen, J.; Zhao, X.; Liang, C.; Wang, Y.; Sun, L.; Jiang, Z.; Zhang, Z.; Yang, R.; Li, Z.; et al. MicroRNA expression signatures of bladder cancer revealed by deep sequencing. PLoS ONE 2011, 6, e18286. [Google Scholar] [CrossRef]
- Wu, L.; Zhang, M.; Qi, L.; Zu, X.; Li, Y.; Liu, L.; Chen, M.; He, W.; Hu, X.; Mo, M.; et al. ERα-mediated alterations in circ_0023642 and miR-490-5p signaling suppress bladder cancer invasion. Cell Death Dis. 2019, 10, 635. [Google Scholar] [CrossRef]
- Maylin, Z.R.; Nicolescu, R.C.; Pandha, H.; Asim, M. Breaking androgen receptor addiction of prostate cancer by targeting different functional domains in the treatment of advanced disease. Transl. Oncol. 2021, 14, 101115. [Google Scholar] [CrossRef]
- Grosdidier, S.; Carbó, L.R.; Buzón, V.; Brooke, G.; Nguyen, P.; Baxter, J.D.; Bevan, C.; Webb, P.; Estébanez-Perpiñá, E.; Fernández-Recio, J. Allosteric conversation in the androgen receptor ligand-binding domain surfaces. Mol. Endocrinol. 2012, 26, 1078–1090. [Google Scholar] [CrossRef]
- Van-Duyne, G.; Blair, I.A.; Sprenger, C.; Moiseenkova-Bell, V.; Plymate, S.; Penning, T.M. The androgen receptor. Vitam. Horm. 2023, 123, 439–481. [Google Scholar] [CrossRef]
- Yonemori, M.; Seki, N.; Yoshino, H.; Matsushita, R.; Miyamoto, K.; Nakagawa, M.; Enokida, H. Dual tumor-suppressors miR-139-5p and miR-139-3p targeting matrix metalloprotease 11 in bladder cancer. Cancer Sci. 2016, 107, 1233–1242. [Google Scholar] [CrossRef]
- Barsky, S.T.; Monks, D.A. The role of androgens and global and tissue-specific androgen receptor expression on body composition, exercise adaptation, and performance. Biol. Sex Differ. 2025, 16, 28. [Google Scholar] [CrossRef]
- Chaudhary, P.; Singha, B.; Abdel-Hafiz, H.A.; Velegraki, M.; Sundi, D.; Satturwar, S.; Parwani, A.V.; Grivennikov, S.I.; You, S.; Goodridge, H.S.; et al. Sex differences in bladder cancer: Understanding biological and clinical implications. Biol. Sex Differ. 2025, 16, 31. [Google Scholar] [CrossRef]
- Doshi, B.; Athans, S.R.; Woloszynska, A. Biological differences underlying sex and gender disparities in bladder cancer: Current synopsis and future directions. Oncogenesis 2023, 12, 44. [Google Scholar] [CrossRef]
- Chang, C.; Lee, S.O.; Yeh, S.; Chang, T.M. Androgen receptor (AR) differential roles in hormone-related tumors including prostate, bladder, kidney, lung, breast and liver. Oncogene 2014, 33, 3225–3234. [Google Scholar] [CrossRef]
- Obeidat, F.; Al Khader, A.; Abdaljaleel, M.; Assaf, F.J.N.; Alsa’oudi, M.M. Expression of androgen receptor in bladder cancer: A tertiary care center study. Int. J. Surg. Open 2023, 56, 100645. [Google Scholar] [CrossRef]
- Hsieh, T.F.; Chen, C.C.; Ma, W.L.; Chuang, W.M.; Hung, X.F.; Tsai, Y.R.; Lin, M.H.; Zhang, Q.; Zhang, C.; Chang, C.; et al. Epidermal growth factor enhances androgen receptor-mediated bladder cancer progression and invasion via potentiation of AR transactivation. Oncol. Rep. 2013, 30, 2917–2922. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, Y.; Izumi, K.; Ishiguro, H.; Ye, B.; Li, F.; Miyamoto, H. Androgen activates β-catenin signaling in bladder cancer cells. Endocr. Relat. Cancer 2013, 20, 293–304. [Google Scholar] [CrossRef]
- Laval University. A Phase II Trial of Bicalutamide in Patients Receiving Intravesical BCG for Non-muscle Invasive Bladder Cancer (BicaBCa). Available online: https://ctv.veeva.com/study/a-phase-ii-trial-of-bicalutamide-in-patients-receiving-intravesical-bcg-for-non-muscle-invasive-blad (accessed on 19 July 2025).
- Deng, G.; Wang, R.; Sun, Y.; Huang, C.P.; Yeh, S.; You, B.; Feng, C.; Li, G.; Ma, S.; Chang, C. Targeting androgen receptor (AR) with antiandrogen Enzalutamide increases prostate cancer cell invasion yet decreases bladder cancer cell invasion via differentially altering the AR/circRNA-ARC1/miR-125b-2-3p or miR-4736/PPARγ/MMP-9 signals. Cell Death Differ. 2021, 28, 2145–2159. [Google Scholar] [CrossRef]
- Kourbanhoussen, K.; McMartin, C.; Lodde, M.; Zlotta, A.; Bryan, R.T.; Toren, P. Switching Cancers: A Systematic Review Assessing the Role of Androgen Suppressive Therapy in Bladder Cancer. Eur. Urol. Focus 2021, 7, 1044–1051. [Google Scholar] [CrossRef]
- Xiang, P.; Du, Z.; Hao, Y.; Guan, D.; Liu, D.; Yan, W.; Wang, M.; Liu, Y.; Ping, H. Impact of Androgen Suppression Therapy on the Risk and Prognosis of Bladder Cancer: A Systematic Review and Meta-Analysis. Front. Oncol. 2021, 11, 784627. [Google Scholar] [CrossRef]
- Li, P.; Chen, J.; Miyamoto, H. Androgen Receptor Signaling in Bladder Cancer. Cancers 2017, 9, 20. [Google Scholar] [CrossRef]
- Xiong, Y.; Wang, L.; Li, Y.; Chen, M.; He, W.; Qi, L. The Long Non-Coding RNA XIST Interacted with MiR-124 to Modulate Bladder Cancer Growth, Invasion and Migration by Targeting Androgen Receptor (AR). Cell Physiol. Biochem. 2017, 43, 405–418. [Google Scholar] [CrossRef]
- Cao, Z.; Xu, L.; Zhao, S.; Zhu, X. The functions of microRNA-124 on bladder cancer. Onco Targets Ther. 2019, 12, 3429–3439. [Google Scholar] [CrossRef]
- Feng, T.; Shao, F.; Wu, Q.; Zhang, X.; Xu, D.; Qian, K.; Xie, Y.; Wang, S.; Xu, N.; Wang, Y.; et al. miR-124 downregulation leads to breast cancer progression via LncRNA-MALAT1 regulation and CDK4/E2F1 signal activation. Oncotarget 2016, 7, 16205–16216. [Google Scholar] [CrossRef]
- Bautista-Sánchez, D.; Arriaga-Canon, C.; Pedroza-Torres, A.; De La Rosa-Velázquez, I.A.; González-Barrios, R.; Contreras-Espinosa, L.; Montiel-Manríquez, R.; Castro-Hernández, C.; Fragoso-Ontiveros, V.; Álvarez-Gómez, R.M.; et al. The Promising Role of miR-21 as a Cancer Biomarker and Its Importance in RNA-Based Therapeutics. Mol. Ther. Nucleic Acids 2020, 20, 409–420. [Google Scholar] [CrossRef] [PubMed]
- Yong-Ming, H.; Ai-Jun, J.; Xiao-Yue, X.; Jian-Wei, L.; Chen, Y.; Ye, C. miR-449a: A potential therapeutic agent for cancer. Anticancer Drugs 2017, 28, 1067–1078. [Google Scholar] [CrossRef]
- Li, M.; Li, J.; Ye, C.; Wu, W.; Cheng, Y. miR-200a-3p predicts prognosis and inhibits bladder cancer cell proliferation by targeting STAT4. Arch. Med. Sci. 2023, 19, 724–735. [Google Scholar] [CrossRef]
- Liu, Q.; You, B.; Meng, J.; Huang, C.P.; Dong, G.; Wang, R.; Chou, F.; Gao, S.; Chang, C.; Yeh, S.; et al. Targeting the androgen receptor to enhance NK cell killing efficacy in bladder cancer by modulating ADAR2/circ_0001005/PD-L1 signaling. Cancer Gene Ther. 2022, 29, 1988–2000. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, S.; Kanthaje, S.; Rekha, P.D.; Mujeeburahiman, M.; Ratnacaram, C.K. Expanding frontiers in liquid biopsy-discovery and validation of circulating biomarkers in renal cell carcinoma and bladder cancer. Int. Rev. Cell Mol. Biol. 2025, 391, 135–197. [Google Scholar] [CrossRef] [PubMed]
- Godoy, G.; Gakis, G.; Smith, C.L.; Fahmy, O. Effects of Androgen and Estrogen Receptor Signaling Pathways on Bladder Cancer Initiation and Progression. Bladder Cancer 2016, 2, 127–137. [Google Scholar] [CrossRef]
- Huang, W.; Chen, Y.; Liu, Y.; Zhang, Q.; Yu, Z.; Mou, L.; Wu, H.; Zhao, L.; Long, T.; Qin, D.; et al. Roles of ERβ and GPR30 in Proliferative Response of Human Bladder Cancer Cell to Estrogen. Biomed Res. Int. 2015, 2015, 251780. [Google Scholar] [CrossRef]
- Thomas, C.; Gustafsson, J. The different roles of ER subtypes in cancer biology and therapy. Nat. Rev. Cancer 2011, 11, 597–608. [Google Scholar] [CrossRef]
- Hsu, I.; Chuang, K.L.; Slavin, S.; Da, J.; Lim, W.X.; Pang, S.T.; O‘Brien, J.H.; Yeh, S. Suppression of ERβ signaling via ERβ knockout or antagonist protects against bladder cancer development. Carcinogenesis 2014, 35, 651–661. [Google Scholar] [CrossRef]
- Goto, T.; Kashiwagi, E.; Jiang, G.; Nagata, Y.; Teramoto, Y.; Baras, A.S.; Yamashita, S.; Ito, A.; Arai, Y.; Miyamoto, H. Estrogen receptor-β signaling induces cisplatin resistance in bladder cancer. Am. J. Cancer Res. 2020, 10, 2523–2534. [Google Scholar]
- Hsu, I.; Yeh, C.R.; Slavin, S.; Miyamoto, H.; Netto, G.J.; Tsai, Y.C.; Muyan, M.; Wu, X.R.; Messing, E.M.; Guancial, E.A.; et al. Estrogen receptor alpha prevents bladder cancer via INPP4B inhibited akt pathway in vitro and in vivo. Oncotarget 2014, 5, 7917–7935. [Google Scholar] [CrossRef]
- Lappano, R.; Todd, L.A.; Stanic, M.; Cai, Q.; Maggiolini, M.; Marincola, F.; Pietrobon, V. Multifaceted Interplay between Hormones, Growth Factors and Hypoxia in the Tumor Microenvironment. Cancers 2022, 14, 539. [Google Scholar] [CrossRef]
- Sato, N.; Ise, K.; Hata, S.; Yamashita, S.; Ito, A.; Sasano, H.; Nakamura, Y. Clinicopathological Significance of Estrogen Receptor. Pathol. Oncol. Res. 2021, 27, 589649. [Google Scholar] [CrossRef]
- Ide, H.; Miyamoto, H. Sex Hormone Receptor Signaling in Bladder Cancer: A Potential Target for Enhancing the Efficacy of Conventional Non-Surgical Therapy. Cells 2021, 10, 1169. [Google Scholar] [CrossRef] [PubMed]
- Ge, Q.; Lu, M.; Ju, L.; Qian, K.; Wang, G.; Wu, C.L.; Liu, X.; Xiao, Y.; Wang, X. miR-4324-RACGAP1-STAT3-ESR1 feedback loop inhibits proliferation and metastasis of bladder cancer. Int. J. Cancer 2019, 144, 3043–3055. [Google Scholar] [CrossRef]
- Huang, B.; Zhai, W.; Hu, G.; Huang, C.; Xie, T.; Zhang, J.; Xu, Y. MicroRNA-206 acts as a tumor suppressor in bladder cancer via targeting YRDC. Am. J. Transl. Res. 2016, 8, 4705–4715. [Google Scholar]
- Fu, B.; Wang, Y.; Zhang, X.; Lang, B.; Zhou, X.; Xu, X.; Zeng, T.; Liu, W.; Guo, J.; Wang, G. MiR-221-induced PUMA silencing mediates immune evasion of bladder cancer cells. Int. J. Oncol. 2015, 46, 1169–1180. [Google Scholar] [CrossRef]
- Wu, D.; Cheung, A.; Wang, Y.; Yu, S.; Chan, F.L. The emerging roles of orphan nuclear receptors in prostate cancer. Biochim. Biophys. Acta 2016, 1866, 23–36. [Google Scholar] [CrossRef]
- Cao, D.; Qi, Z.; Pang, Y.; Li, H.; Xie, H.; Wu, J.; Huang, Y.; Zhu, Y.; Shen, Y.; Dai, B.; et al. Retinoic Acid-Related Orphan Receptor C Regulates Proliferation, Glycolysis, and Chemoresistance via the PD-L1/ITGB6/STAT3 Signaling Axis in Bladder Cancer. Cancer Res. 2019, 79, 2604–2618. [Google Scholar] [CrossRef]
- Fan, J.; Lv, Z.; Yang, G.; Liao, T.T.; Xu, J.; Wu, F.; Huang, Q.; Guo, M.; Hu, G.; Zhou, M.; et al. Retinoic Acid Receptor-Related Orphan Receptors: Critical Roles in Tumorigenesis. Front. Immunol. 2018, 9, 1187. [Google Scholar] [CrossRef]
- Wu, L.; Chen, L. Characteristics of Nur77 and its ligands as potential anticancer compounds (Review). Mol. Med. Rep. 2018, 18, 4793–4801. [Google Scholar] [CrossRef]
- Cho, S.D.; Lee, S.O.; Chintharlapalli, S.; Abdelrahim, M.; Khan, S.; Yoon, K.; Kamat, A.M.; Safe, S. Activation of nerve growth factor-induced B alpha by methylene-substituted diindolylmethanes in bladder cancer cells induces apoptosis and inhibits tumor growth. Mol. Pharmacol. 2010, 77, 396–404. [Google Scholar] [CrossRef]
- Guo, Q.; Zhang, T.; Gong, Y.; Tao, Y.; Gao, Y.; Wang, Y.; Tian, J.; Zhang, S.; Wang, H.; Rodriguez, R.; et al. Aldehyde dehydrogenase 6 family member A1 negatively regulates cell growth and to cisplatin sensitivity in bladder cancer. Mol. Carcinog. 2022, 61, 690–701. [Google Scholar] [CrossRef] [PubMed]
- Lv, D.D.; Zhou, L.Y.; Tang, H. Hepatocyte nuclear factor 4α and cancer-related cell signaling pathways: A promising insight into cancer treatment. Exp. Mol. Med. 2021, 53, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Oh, M.; Kim, S.Y.; Gil, J.E.; Byun, J.S.; Cha, D.W.; Ku, B.; Lee, W.; Kim, W.K.; Oh, K.J.; Lee, E.W.; et al. Nurr1 performs its anti-inflammatory function by regulating RasGRP1 expression in neuro-inflammation. Sci. Rep. 2020, 10, 10755. [Google Scholar] [CrossRef]
- Mao, W.; Xiong, G.; Wu, Y.; Wang, C.; St. Clair, D.; Li, J.D.; Xu, R. RORα Suppresses Cancer-Associated Inflammation by Repressing Respiratory Complex I-Dependent ROS Generation. Int. J. Mol. Sci. 2021, 22, 10665. [Google Scholar] [CrossRef]
- Sun, H.; Tian, J.; Xian, W.; Xie, T.; Yang, X. miR-34a inhibits proliferation and invasion of bladder cancer cells by targeting orphan nuclear receptor HNF4G. Dis. Markers 2015, 2015, 879254. [Google Scholar] [CrossRef]
- Zhang, L.; Liao, Y.; Tang, L. MicroRNA-34 family: A potential tumor suppressor and therapeutic candidate in cancer. J. Exp. Clin. Cancer Res. 2019, 38, 53. [Google Scholar] [CrossRef]
- Li, S.; Wei, X.; He, J.; Cao, Q.; Du, D.; Zhan, X.; Zeng, Y.; Yuan, S.; Sun, L. The comprehensive landscape of miR-34a in cancer research. Cancer Metastasis Rev. 2021, 40, 925–948. [Google Scholar] [CrossRef]
- Wu, J.; Liu, J.; Jia, R.; Song, H. Nur77 inhibits androgen-induced bladder cancer growth. Cancer Invest. 2013, 31, 654–660. [Google Scholar] [CrossRef]
- Ye, X.; Guo, J.; Zhang, H.; Meng, Q.; Ma, Y.; Lin, R.; Yi, X.; Lu, H.; Bai, X.; Cheng, J. The enhanced expression of estrogen-related receptor α in human bladder cancer tissues and the effects of estrogen-related receptor α knockdown on bladder cancer cells. J. Cell Biochem. 2019, 120, 13841–13852. [Google Scholar] [CrossRef]
- Lemberger, T.; Braissant, O.; Juge-Aubry, C.; Keller, H.; Saladin, R.; Staels, B.; Auwerx, J.; Burger, A.G.; Meier, C.A.; Wahli, W. PPAR tissue distribution and interactions with other hormone-signaling pathways. Ann. N. Y. Acad Sci. 1996, 804, 231–251. [Google Scholar] [CrossRef]
- Tate, T.; Xiang, T.; Wobker, S.E.; Zhou, M.; Chen, X.; Kim, H.; Batourina, E.; Lin, C.S.; Kim, W.Y.; Lu, C.; et al. Pparg signaling controls bladder cancer subtype and immune exclusion. Nat. Commun. 2021, 12, 6160. [Google Scholar] [CrossRef]
- Varley, C.L.; Stahlschmidt, J.; Smith, B.; Stower, M.; Southgate, J. Activation of peroxisome proliferator-activated receptor-gamma reverses squamous metaplasia and induces transitional differentiation in normal human urothelial cells. Am. J. Pathol. 2004, 164, 1789–1798. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.P.; Lapi, E.; Martínez de Villarreal, J.; Álvaro-Espinosa, L.; Fernández-Barral, A.; Barbáchano, A.; Domínguez, O.; Laughney, A.M.; Megías, D.; Muñoz, A.; et al. Urothelial organoids originating from Cd49f. Nat. Commun. 2019, 10, 4407. [Google Scholar] [CrossRef]
- Lehrke, M.; Lazar, M.A. The many faces of PPARgamma. Cell 2005, 123, 993–999. [Google Scholar] [CrossRef]
- Biton, A.; Bernard-Pierrot, I.; Lou, Y.; Krucker, C.; Chapeaublanc, E.; Rubio-Pérez, C.; López-Bigas, N.; Kamoun, A.; Neuzillet, Y.; Gestraud, P.; et al. Independent component analysis uncovers the landscape of the bladder tumor transcriptome and reveals insights into luminal and basal subtypes. Cell Rep. 2014, 9, 1235–1245. [Google Scholar] [CrossRef]
- Korpal, M.; Puyang, X.; Wu, Z.J.; Seiler, R.; Furman, C.; Oo, H.Z.; Seiler, M.; Irwin, S.; Subramanian, V.; Joshi, J.J.; et al. Evasion of immunosurveillance by genomic alterations of PPARγ/RXRα in bladder cancer. Nat. Commun. 2017, 8, 103. [Google Scholar] [CrossRef]
- Goldstein, J.T.; Berger, A.C.; Shih, J.; Duke, F.F.; Furst, L.; Kwiatkowski, D.J.; Cherniack, A.D.; Meyerson, M.; Strathdee, C.A. Genomic activation of PPARG reveals a candidate therapeutic axis in bladder cancer. Cancer Res. 2017, 77, 6987–6998. [Google Scholar] [CrossRef]
- Garcia-Bates, T.M.; Bernstein, S.H.; Phipps, R.P. Peroxisome proliferator-activated receptor gamma overexpression suppresses growth and induces apoptosis in human multiple myeloma cells. Clin. Cancer Res. 2008, 14, 6414–6425. [Google Scholar] [CrossRef]
- Fauconnet, S.; Lascombe, I.; Chabannes, E.; Adessi, G.L.; Desvergne, B.; Wahli, W.; Bittard, H. Differential regulation of vascular endothelial growth factor expression by peroxisome proliferator-activated receptors in bladder cancer cells. J. Biol. Chem. 2002, 277, 23534–23543. [Google Scholar] [CrossRef]
- Peters, J.M.; Shah, Y.M.; Gonzalez, F.J. The role of peroxisome proliferator-activated receptors in carcinogenesis and chemoprevention. Nat. Rev. Cancer 2012, 12, 181–195. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Li, H.; Liu, S.; Xie, X.; Zhai, W.; Pan, J. Long noncoding RNA UCA1 inhibits epirubicin-induced apoptosis by activating PPARα-mediated lipid metabolism. Exp. Cell Res. 2024, 442, 114271. [Google Scholar] [CrossRef] [PubMed]
- Ide, H.; Inoue, S.; Miyamoto, H. The Role of Glucocorticoid Receptor Signaling in Bladder Cancer Progression. Cancers 2018, 10, 484. [Google Scholar] [CrossRef]
- Ide, H.; Inoue, S.; Mizushima, T.; Kashiwagi, E.; Zheng, Y.; Miyamoto, H. Role of glucocorticoid signaling in urothelial tumorigenesis: Inhibition by prednisone presumably through inducing glucocorticoid receptor transrepression. Mol. Carcinog. 2019, 58, 2297–2305. [Google Scholar] [CrossRef]
- Lockett, J.; Inder, W.J.; Clifton, V.L. The Glucocorticoid Receptor: Isoforms, Functions, and Contribution to Glucocorticoid Sensitivity. Endocr. Rev. 2024, 45, 593–624. [Google Scholar] [CrossRef]
- Ramos-Ramírez, P.; Tliba, O. Glucocorticoid Receptor β (GRβ): Beyond Its Dominant-Negative Function. Int. J. Mol. Sci. 2021, 22, 3649. [Google Scholar] [CrossRef]
- Shi, W.; Wang, D.; Yuan, X.; Liu, Y.; Guo, X.; Li, J.; Song, J. Glucocorticoid receptor-IRS-1 axis controls EMT and the metastasis of breast cancers. J. Mol. Cell Biol. 2019, 11, 1042–1055. [Google Scholar] [CrossRef]
- Habib, T.; Sadoun, A.; Nader, N.; Suzuki, S.; Liu, W.; Jithesh, P.V.; Kino, T. AKT1 has dual actions on the glucocorticoid receptor by cooperating with 14-3-3. Mol. Cell Endocrinol. 2017, 439, 431–443. [Google Scholar] [CrossRef]
- Bo, J.; Yang, G.; Huo, K.; Jiang, H.; Zhang, L.; Liu, D.; Huang, Y. microRNA-203 suppresses bladder cancer development by repressing bcl-w expression. FEBS J. 2011, 278, 786–792. [Google Scholar] [CrossRef]
- Saini, S.; Arora, S.; Majid, S.; Shahryari, V.; Chen, Y.; Deng, G.; Yamamura, S.; Ueno, K.; Dahiya, R. Curcumin modulates microRNA-203-mediated regulation of the Src-Akt axis in bladder cancer. Cancer Prev. Res. 2011, 4, 1698–1709. [Google Scholar] [CrossRef]
- Lynch, J.T.; Rajendran, R.; Xenaki, G.; Berrou, I.; Demonacos, C.; Krstic-Demonacos, M. The role of glucocorticoid receptor phosphorylation in Mcl-1 and NOXA gene expression. Mol. Cancer 2010, 9, 38. [Google Scholar] [CrossRef] [PubMed]
- BharathwajChetty, B.; Sajeev, A.; Vishwa, R.; Aswani, B.S.; Alqahtani, M.S.; Abbas, M.; Kunnumakkara, A.B. Dynamic interplay of nuclear receptors in tumor cell plasticity and drug resistance: Shifting gears in malignant transformations and applications in cancer therapeutics. Cancer Metastasis Rev. 2024, 43, 321–362. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, D.J.; Missiaen, R.; Skuli, N.; Steger, D.J.; Simon, M.C. Cell-Intrinsic Tumorigenic Functions of PPARγ in Bladder Urothelial Carcinoma. Mol. Cancer Res. 2021, 19, 598–611. [Google Scholar] [CrossRef]
- Safe, S.; Jin, U.H.; Hedrick, E.; Reeder, A.; Lee, S.O. Minireview: Role of orphan nuclear receptors in cancer and potential as drug targets. Mol. Endocrinol. 2014, 28, 157–172. [Google Scholar] [CrossRef]
- Okegawa, T.; Ushio, K.; Imai, M.; Morimoto, M.; Hara, T. Orphan nuclear receptor HNF4G promotes bladder cancer growth and invasion through the regulation of the hyaluronan synthase 2 gene. Oncogenesis 2013, 2, e58. [Google Scholar] [CrossRef]
- Wu, J.T.; Han, B.M.; Yu, S.Q.; Wang, H.P.; Xia, S.J. Androgen receptor is a potential therapeutic target for bladder cancer. Urology 2010, 75, 820–827. [Google Scholar] [CrossRef]
- Hoffman, K.L.; Lerner, S.P.; Smith, C.L. Raloxifene inhibits growth of RT4 urothelial carcinoma cells via estrogen receptor-dependent induction of apoptosis and inhibition of proliferation. Horm. Cancer 2013, 4, 24–35. [Google Scholar] [CrossRef]
- Goto, T.; Miyamoto, H. The Role of Estrogen Receptors in Urothelial Cancer. Front. Endocrinol. 2021, 12, 643870. [Google Scholar] [CrossRef]
- Whang, Y.M.; Jin, S.B.; Park, S.I.; Chang, I.H. MEK inhibition enhances efficacy of bacillus Calmette-Guérin on bladder cancer cells by reducing release of Toll-like receptor 2-activated antimicrobial peptides. Oncotarget 2017, 8, 53168–53179. [Google Scholar] [CrossRef]
- Bourn, J.; Cekanova, M. Cyclooxygenase inhibitors potentiate receptor tyrosine kinase therapies in bladder cancer cells in vitro. Drug Des. Devel. Ther. 2018, 12, 1727–1742. [Google Scholar] [CrossRef]
- Wang, S.; Wu, G.; Han, Y.; Song, P.; Chen, J.; Wu, Y.; Yang, J.; Liang, P. miR-124 regulates STAT3-mediated cell proliferation, migration and apoptosis in bladder cancer. Oncol Lett. 2018, 16, 5875–5881. [Google Scholar] [CrossRef]
- Lee, J.J.; Drakaki, A.; Iliopoulos, D.; Struhl, K. MiR-27b targets PPARγ to inhibit growth, tumor progression and the inflammatory response in neuroblastoma cells. Oncogene 2012, 31, 3818–3825. [Google Scholar] [CrossRef]
- Alexa, F. The Roles of Nuclear Receptor NR4A1 in Cancer Cell Proliferation and Skeletal Muscle Differentiation. Ph.D. Thesis, University of Tennessee Health Science Center, Memphis, TN, USA, 2016. [Google Scholar]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flores-Estrada, J.J.; Jiménez, A.; Victoria-Acosta, G.; Cortés-Malagón, E.M.; Ortiz-López, M.G.; Alvarez-Sánchez, M.E.; Nuñez-Olvera, S.I.; Pérez-Navarro, Y.F.; Morales-Reyna, M.; Puente-Rivera, J. Nuclear Receptors in Bladder Cancer: Insights into miRNA-Mediated Regulation and Potential Therapeutic Implications. Int. J. Mol. Sci. 2025, 26, 7340. https://doi.org/10.3390/ijms26157340
Flores-Estrada JJ, Jiménez A, Victoria-Acosta G, Cortés-Malagón EM, Ortiz-López MG, Alvarez-Sánchez ME, Nuñez-Olvera SI, Pérez-Navarro YF, Morales-Reyna M, Puente-Rivera J. Nuclear Receptors in Bladder Cancer: Insights into miRNA-Mediated Regulation and Potential Therapeutic Implications. International Journal of Molecular Sciences. 2025; 26(15):7340. https://doi.org/10.3390/ijms26157340
Chicago/Turabian StyleFlores-Estrada, José Javier, Adriana Jiménez, Georgina Victoria-Acosta, Enoc Mariano Cortés-Malagón, María Guadalupe Ortiz-López, María Elizbeth Alvarez-Sánchez, Stephanie I. Nuñez-Olvera, Yussel Fernando Pérez-Navarro, Marcos Morales-Reyna, and Jonathan Puente-Rivera. 2025. "Nuclear Receptors in Bladder Cancer: Insights into miRNA-Mediated Regulation and Potential Therapeutic Implications" International Journal of Molecular Sciences 26, no. 15: 7340. https://doi.org/10.3390/ijms26157340
APA StyleFlores-Estrada, J. J., Jiménez, A., Victoria-Acosta, G., Cortés-Malagón, E. M., Ortiz-López, M. G., Alvarez-Sánchez, M. E., Nuñez-Olvera, S. I., Pérez-Navarro, Y. F., Morales-Reyna, M., & Puente-Rivera, J. (2025). Nuclear Receptors in Bladder Cancer: Insights into miRNA-Mediated Regulation and Potential Therapeutic Implications. International Journal of Molecular Sciences, 26(15), 7340. https://doi.org/10.3390/ijms26157340