
Membrane Progesterone Receptor Beta Regulates the Decidualization of Endometrial Stromal Cells in Women with Endometriosis

 , , , , , , and
, , , , , , and
Abstract
1. Introduction
2. Results
2.1. Demographical and Clinical Data
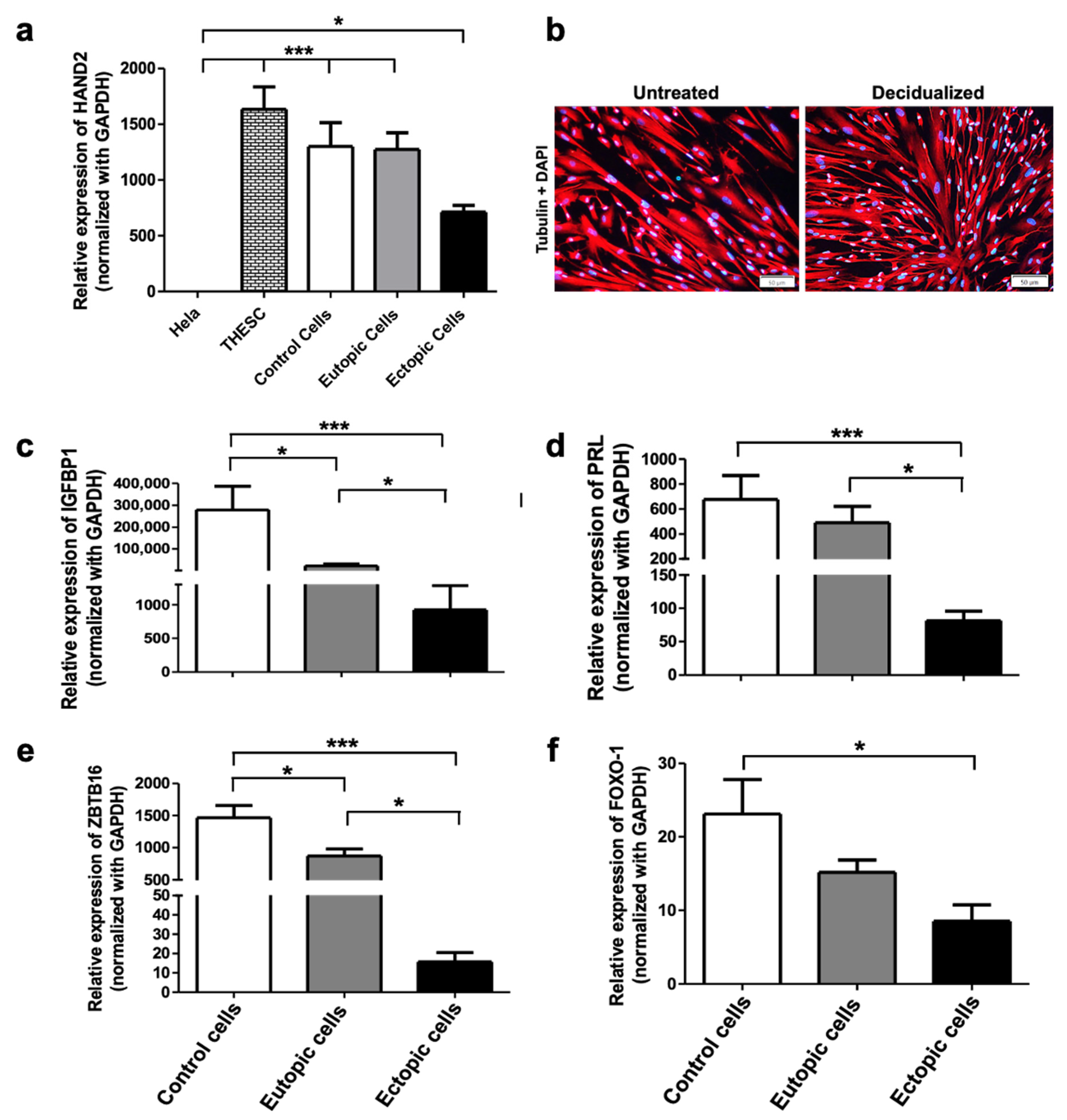
2.2. Decidualization of Human ESCs
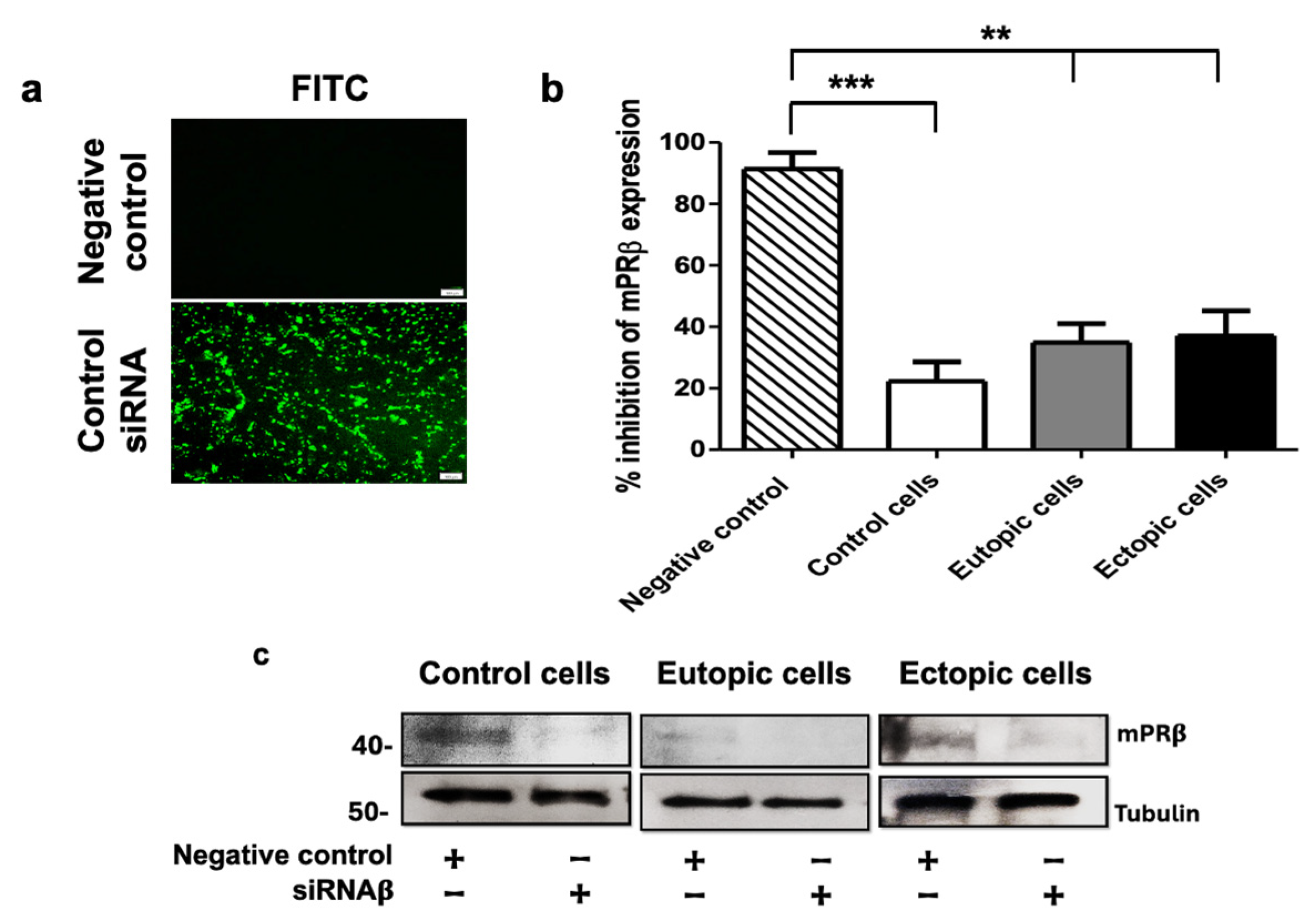
2.3. mPRs Participation in the Decidualization of Human Endometrial Stromal Cells
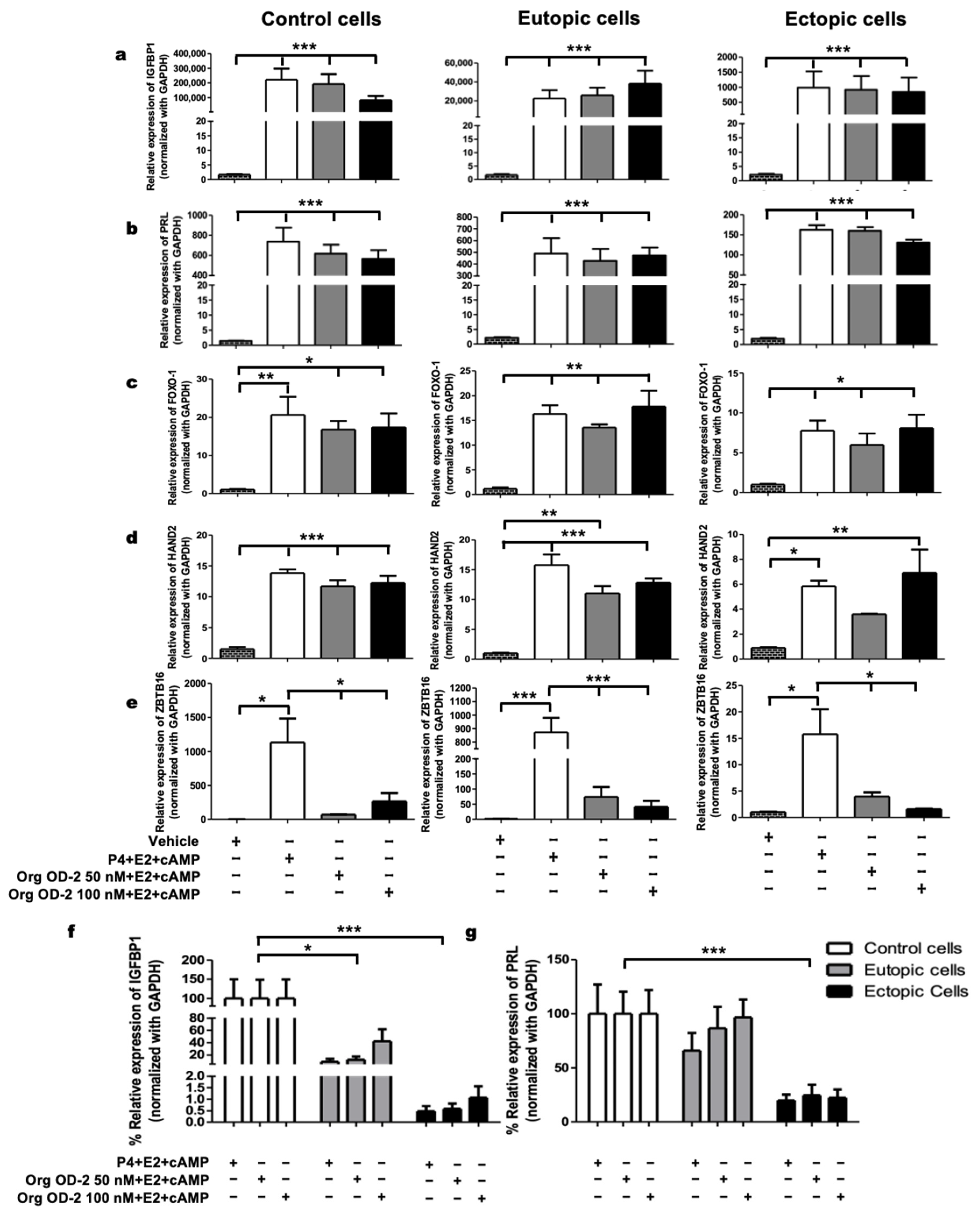
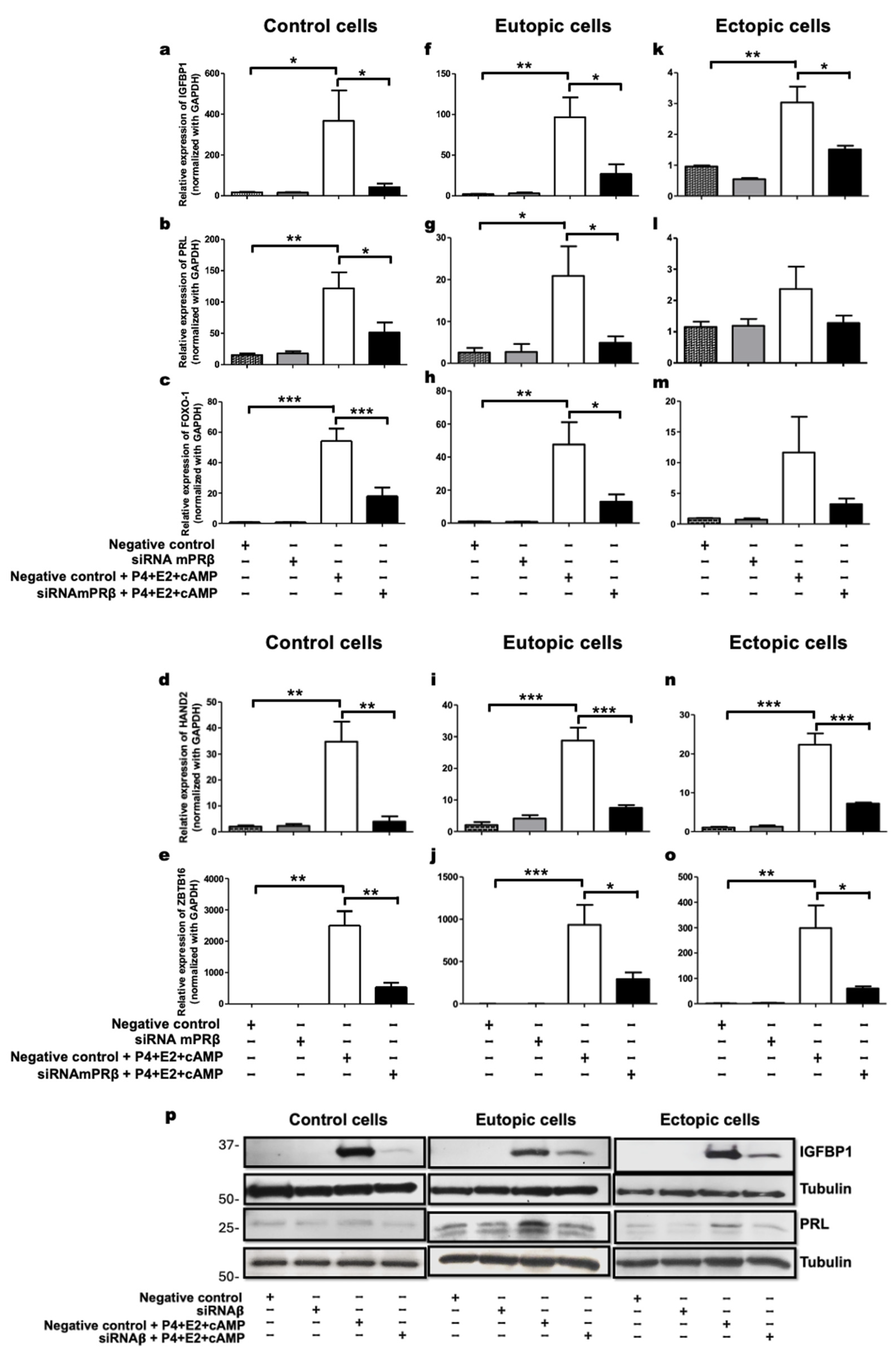
2.4. mPRβ Induces the Decidualization of Human Endometrial Stromal Cells
3. Discussion
4. Materials and Methods
4.1. Tissue Collection
4.2. Primary Cell Culture of ESCs
4.3. Cell Lines
4.4. Decidualization Treatments
4.5. RNA Isolation and Reverse Transcription-Quantitative PCR (RT-qPCR)
4.6. Immunofluorescence
4.7. mPRβ siRNA Transfection
4.8. Western Blot
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| mPRβ | Membrane Progesterone Receptor β |
| ESCs | Endometrial Stromal Cells |
| HAND2 | Heart and Neural Crest Derivatives-Expressed Transcript 2 |
| FOXO1 | Fork head Box O1 |
| BMP2 | Bone Morphogenetic Protein 2 a member of the Transforming Growth Factor-Beta (TGF-β) superfamily |
| HOXA10 | Homeobox A10 |
| IGFBP1 | Insulin-Like Growth Factor-Binding Protein 1 |
| PRL | Prolactin |
| ZBTB16 | Zinc Finger and BTB Domain-Containing 16 |
| PAQR | Progestin and AdipoQ Receptors |
| Org OD 02-0 | 10-Ethenyl-19-norprogesterone |
| cAMP | Cyclic Adenosine Monophosphate |
Appendix A
References
- Taylor, H.S.; Kotlyar, A.M.; Flores, V.A. Endometriosis is a chronic systemic disease: Clinical challenges and novel innovation. Lancet 2021, 397, 839–852. [Google Scholar] [CrossRef] [PubMed]
- Pasalic, E.; Tambuwala, M.M.; Jahjefendic-Hromic, A. Endometriosis: Classification, pathophysiology, and treatment options. Pathol. Res. Pract. 2023, 251, 154847. [Google Scholar] [CrossRef] [PubMed]
- Prescott, J.; Farland, L.V.; Tobias, D.K.; Gaskins, A.J.; Spiegelman, D.; Chavarro, J.E.; Rich-Edwards, J.W.; Barbieri, R.L.; Missmer, S.A. A prospective cohort study of endometriosis and subsequent risk of infertility. Hum. Reprod. 2016, 31, 1475–1482. [Google Scholar] [CrossRef]
- Feng, X.; Qi, L.; Xu, X.; Feng, Y.; Gong, X.; Aili, A.; Tong, X. Analysis of differences in the transcriptomic profiles of eutopic and ectopic endometriums in women with ovarian endometriosis. PeerJ 2021, 9, e11045. [Google Scholar] [CrossRef]
- Chantalat, E.; Valera, M.C.; Vaysse, C.; Noirrit, E.; Rusidze, M.; Weyl, A.; Vergriete, K.; Buscail, E.; Lluel, P.; Fontaine, C.; et al. Estrogen Receptors and Endometriosis. Int. J. Mol. Sci. 2020, 21, 2815. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Wang, G. Progesterone Resistance in Endometriosis: Current Evidence and Putative Mechanisms. Int. J. Mol. Sci. 2023, 24, 6992. [Google Scholar] [CrossRef]
- Ochoa-Bernal, M.A.; Fazleabas, A.T. Physiologic Events of Embryo Implantation and Decidualization in Human and Non-Human Primates. Int. J. Mol. Sci. 2020, 21, 1973. [Google Scholar] [CrossRef] [PubMed]
- Murata, H.; Tanaka, S.; Tsuzuki-Nakao, T.; Kido, T.; Kakita-Kobayashi, M.; Kida, N.; Hisamatsu, Y.; Tsubokura, H.; Hashimoto, Y.; Kitada, M.; et al. The transcription factor HAND2 up-regulates transcription of the IL15 gene in human endometrial stromal cells. J. Biol. Chem. 2020, 295, 9596–9605. [Google Scholar] [CrossRef]
- Grinius, L.; Kessler, C.; Schroeder, J.; Handwerger, S. Forkhead transcription factor FOXO1A is critical for induction of human decidualization. J. Endocrinol. 2006, 189, 179–187. [Google Scholar] [CrossRef]
- Huang, P.; Deng, W.; Bao, H.; Lin, Z.; Liu, M.; Wu, J.; Zhou, X.; Qiao, M.; Yang, Y.; Cai, H.; et al. SOX4 facilitates PGR protein stability and FOXO1 expression conducive for human endometrial decidualization. eLife 2022, 11, e72073. [Google Scholar] [CrossRef]
- Liao, Z.; Tang, S.; Jiang, P.; Geng, T.; Cope, D.I.; Dunn, T.N.; Guner, J.; Radilla, L.A.; Guan, X.; Monsivais, D. Impaired bone morphogenetic protein (BMP) signaling pathways disrupt decidulization in endometriosis. Commun. Biol. 2024, 7, 227. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Lu, J.; Wu, J.; Jiang, R.; Guo, C.; Tang, Y.; Wang, H.; Kong, S.; Wang, S. HOXA10 co-factor MEIS1 is required for the decidualization in human endometrial stromal cell. J. Mol. Endocrinol. 2020, 64, 249–258. [Google Scholar] [CrossRef]
- Ng, S.W.; Norwitz, G.A.; Pavlicev, M.; Tilburgs, T.; Simon, C.; Norwitz, E.R. Endometrial decidualization: The primary driver of pregnancy health. Int. J. Mol. Sci. 2020, 21, 4092. [Google Scholar] [CrossRef]
- Kommagani, R.; Szwarc, M.M.; Vasquez, Y.M.; Peavey, M.C.; Mazur, E.C.; Gibbons, W.E.; Lanz, R.B.; DeMayo, F.J.; Lydon, J.P. The Promyelocytic Leukemia Zinc Finger Transcription Factor Is Critical for Human Endometrial Stromal Cell Decidualization. PLoS Genet. 2016, 12, e1005937. [Google Scholar] [CrossRef] [PubMed]
- Arlier, S.; Kayisli, U.A.; Semerci, N.; Ozmen, A.; Larsen, K.; Schatz, F.; Lockwood, C.J.; Guzeloglu-Kayisli, O. Enhanced ZBTB16 Levels by Progestin-Only Contraceptives Induces Decidualization and Inflammation. Int. J. Mol. Sci. 2023, 24, 10532. [Google Scholar] [CrossRef]
- Garrido-Gomez, T.; Dominguez, F.; Quiñonero, A.; Diaz-Gimeno, P.; Kapidzic, M.; Gormley, M.; Ona, K.; Padilla-Iserte, P.; McMaster, M.; Genbacev, O.; et al. Defective decidualization during and after severe preeclampsia reveals a possible maternal contribution to the etiology. Proc. Natl. Acad. Sci. USA 2017, 114, E8468–E8477. [Google Scholar] [CrossRef]
- Cope, D.I.; Monsivais, D. Progesterone Receptors Signaling in Uterus Is Essential for Pregnancy Success. Cell 2022, 11, 1474. [Google Scholar] [CrossRef] [PubMed]
- Medina-Laver, Y.; Rodríguez-Varela, C.; Salsano, S.; Labarta, E.; Domínguez, F. What Do We Know about Classical and Non-Classical Progesterone Receptors in the Human Female Reproductive Tract? A Review. Int. J. Mol. Sci. 2021, 22, 11278. [Google Scholar] [CrossRef]
- Kaya, H.S.; Hantak, A.M.; Stubbs, L.J.; Taylor, R.N.; Bagchi, I.C.; Bagchi, M.K. Roles of Progesterone Receptor A and B Isoforms During Human Endometrial Decidualization. Mol. Endocrinol. 2015, 29, 882–895. [Google Scholar] [CrossRef]
- Chi, R.A.; Wang, T.; Adams, N.; Wu, S.P.; Young, S.L.; Spencer, T.E.; DeMayo, F. Human Endometrial Transcriptome and Progesterone Receptor Cistrome Reveal Important Pathways and Epithelial Regulators. J. Clin. Endocrinol. Metab. 2020, 105, e1419–e1439. [Google Scholar] [CrossRef]
- Retis-Resendiz, A.M.; Cid-Cruz, Y.; Velázquez-Hernández, D.M.; Romero-Reyes, J.; León-Juárez, M.; García-Gómez, E.; Camacho-Arroyo, I.; Vázquez-Martínez. cAMP regulates the progesterone receptor gene expression through the protein kinase A pathway during decidualization in human immortalized endometrial stromal cells. Steroids 2024, 203, 109363. [Google Scholar] [CrossRef]
- Salsano, S.; González-Martín, R.; Quiñonero, A.; Pérez-Debén, S.; Domínguez, F. Deciphering the Role of PGRMC1 During Human Decidualization Using an In Vitro Approach. J. Clin. Endocrinol. Metab. 2021, 106, 2313–2327. [Google Scholar] [CrossRef] [PubMed]
- Tsuru, A.; Yoshie, M.; Kojima, J.; Yonekawa, R.; Azumi, M.; Kusama, K.; Nishi, H.; Tamura, K. PGRMC1 Regulates Cellular Senescence via Modulating FOXO1 Expression in Decidualizing Endometrial Stromal Cells. Biomolecules 2022, 12, 1046. [Google Scholar] [CrossRef] [PubMed]
- Tomas, P. Membrane Progesterone Receptors (mPRs, PAQRs): Review of Structural and Signaling Characteristics. Cells 2022, 11, 1785. [Google Scholar] [CrossRef]
- Nader, N.; Dib, M.; Hodeify, R.; Courjaret, R.; Elmi, A.; Hammad, A.S.; Dey, R.; Huang, X.Y.; Machaca, K. Membrane progesterone receptor induces meiosis in Xenopus oocytes through endocytosis into signaling endosomes and interaction with APPL1 and Akt2. PLoS Biol. 2020, 18, e3000901. [Google Scholar] [CrossRef] [PubMed]
- Kowalik, K.; Dobrzyn, K.; Rekawiecki, R.; Kotwica, J. Expression of membrane progestin receptors (mPRs) α, β and γ in the bovine uterus during the oestrous cycle and pregnancy. Theriogenology 2019, 140, 171–179. [Google Scholar] [CrossRef]
- Fernandes, M.S.; Pierron, V.; Michalovich, D.; Astle, S.; Thornton, S.; Peltoketo, H.; Lam, E.W.; Gellersen, B.; Huhtaniemi, I.; Allen, J.; et al. Regulated expression of putative membrane progestin receptor homologues in human endometrium and gestational tissues. J. Endocrinol. 2005, 187, 89–101. [Google Scholar] [CrossRef]
- Rahnama, R.; Rafiee, M.; Fouladi, S.; Akbari-Fakhrabadi, M.; Mehrabian, F.; Rezaei, A. Gene expression analysis of membrane progesterone receptors in women with recurrent spontaneous abortion: A case control study. BMC Res. Notes 2019, 12, 790. [Google Scholar] [CrossRef]
- Vázquez-Martínez, E.R.; Bello-Alvarez, C.; Hermenegildo-Molina, A.L.; Solís-Paredes, M.; Parra-Hernández, S.; Cruz-Orozco, O.; Silvestri-Tomassoni, J.R.; Escobar-Ponce, L.F.; Hernández-López, L.A.; Reyes-Mayoral, C.; et al. Expression of Membrane Progesterone Receptors in Eutopic and Ectopic Endometrium of Women with Endometriosis. BioMed Res. Int. 2020, 2020, 2196024. [Google Scholar] [CrossRef]
- Tanaka, K.; Sakai, K.; Matsushima, M.; Matsuzawa, Y.; Izawa, T.; Nagashima, T.; Furukawa, S.; Kobayashi, Y.; Iwashita, M. Branched-chain amino acids regulate insulin-like growth factor-binding protein 1 (IGFBP1) production by decidua and influence trophoblast migration through IGFBP1. Mol. Hum. Reprod. 2016, 22, 890–899. [Google Scholar] [CrossRef]
- Bunch, K.; Tinnemore, D.; Huff, S.; Hoffer, Z.S.; Burney, R.O.; Stallings, J.D. Expression Patterns of Progesterone Receptor, Membrane Components 1 and 2 in Endometria From Women with and Without Endometriosis. Reprod. Sci. 2014, 21, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Xu, M.; Zhang, H.; Zhang, M.; Wang, J.; Mei, J.; Zhang, Y.; Zhou, J.; Zhen, X.; Kang, N.; et al. Endometrial stromal PRMT5 plays a crucial role in decidualization by regulating NF-κB signaling in endometriosis. Cell Death Discov. 2022, 8, 408. [Google Scholar] [CrossRef] [PubMed]
- Klemmt, P.A.; Carver, J.G.; Kennedy, S.H.; Koninckx, P.R.; Mardon, H.J. Stromal cells from endometriotic lesions and endometrium from women with endometriosis have reduced decidualization capacity. Fertil. Steril. 2006, 85, 564–572. [Google Scholar] [CrossRef] [PubMed]
- Chae, U.; Min, J.Y.; Kim, S.H.; Ihm, H.J.; Oh, Y.S.; Park, S.Y.; Chae, H.D.; Kim, C.H.; Kang, B.M. Decreased progesterone receptor B/A ratio in endometrial cells by tumor necrosis factor-alpha and peritoneal fluid from patients with endometriosis. Yonsei Med. J. 2016, 57, 1468–1474. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Characteristics | Control (n = 20) | Women with Endometriosis (n = 20) |
|---|---|---|
| Age years (mean ± SD) | 38.4 (± 6.6) | 33.22 (± 9.7) |
| Number of women with full-term pregnancies | 20 | 4 |
| Number of women with abortions | 2 | 11 |
| Number of women in the proliferative phase during biopsy collection | 13 | 11 |
| Number of women in the secretory phase during biopsy collection | 7 | 9 |
| GENE | FORWARD 5′3′ | REVERSE 5′3′ |
|---|---|---|
| HAND 2 | GAGGAAGAAGGAGCTGAACGAA | GTCCGGCCTTTGGTTTTCTTG |
| IGFBP1 | TCCTTTGGGACGCCATCAGTAC | GATGTCTCCTGTGCCTTGGCTA |
| PRL | CATATTGCGATCCTGGAATGAGC | TCCTCAATCTCTACAGCTTTGGA |
| FOXO-1 | AGGDTTAGTGAGCAGGTTACAC | TGGACTGCTTCTCTCAGTTCC |
| ZBTB16 | TTGTCTGTGATCAGTGCGGT | GCCATGTCAGTGCCAGTATG |
| PAQR8 | AGGACGCAGCAAACAGGACA | GCC AACACAGGCAGGAATAA |
| GAPDH | GACAGTCAGCCGCATCTTCT | GCCCAATACGAC CAAATCCGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Velázquez-Hernández, D.M.; Vázquez-Martínez, E.R.; Cruz-Orozco, O.; Silvestri-Tomassoni, J.R.; Sánchez-Ramírez, B.; Olguín-Ortega, A.; Escobar-Ponce, L.F.; Rodríguez-Dorantes, M.; Camacho-Arroyo, I. Membrane Progesterone Receptor Beta Regulates the Decidualization of Endometrial Stromal Cells in Women with Endometriosis. Int. J. Mol. Sci. 2025, 26, 7297. https://doi.org/10.3390/ijms26157297
Velázquez-Hernández DM, Vázquez-Martínez ER, Cruz-Orozco O, Silvestri-Tomassoni JR, Sánchez-Ramírez B, Olguín-Ortega A, Escobar-Ponce LF, Rodríguez-Dorantes M, Camacho-Arroyo I. Membrane Progesterone Receptor Beta Regulates the Decidualization of Endometrial Stromal Cells in Women with Endometriosis. International Journal of Molecular Sciences. 2025; 26(15):7297. https://doi.org/10.3390/ijms26157297
Chicago/Turabian StyleVelázquez-Hernández, Dora Maria, Edgar Ricardo Vázquez-Martínez, Oliver Cruz-Orozco, José Roberto Silvestri-Tomassoni, Brenda Sánchez-Ramírez, Andrea Olguín-Ortega, Luis F. Escobar-Ponce, Mauricio Rodríguez-Dorantes, and Ignacio Camacho-Arroyo. 2025. "Membrane Progesterone Receptor Beta Regulates the Decidualization of Endometrial Stromal Cells in Women with Endometriosis" International Journal of Molecular Sciences 26, no. 15: 7297. https://doi.org/10.3390/ijms26157297
APA StyleVelázquez-Hernández, D. M., Vázquez-Martínez, E. R., Cruz-Orozco, O., Silvestri-Tomassoni, J. R., Sánchez-Ramírez, B., Olguín-Ortega, A., Escobar-Ponce, L. F., Rodríguez-Dorantes, M., & Camacho-Arroyo, I. (2025). Membrane Progesterone Receptor Beta Regulates the Decidualization of Endometrial Stromal Cells in Women with Endometriosis. International Journal of Molecular Sciences, 26(15), 7297. https://doi.org/10.3390/ijms26157297