

Confocal Laser Scanning Microscopy of Light-Independent ROS in Arabidopsis thaliana (L.) Heynh. TROL-FNR Mutants


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
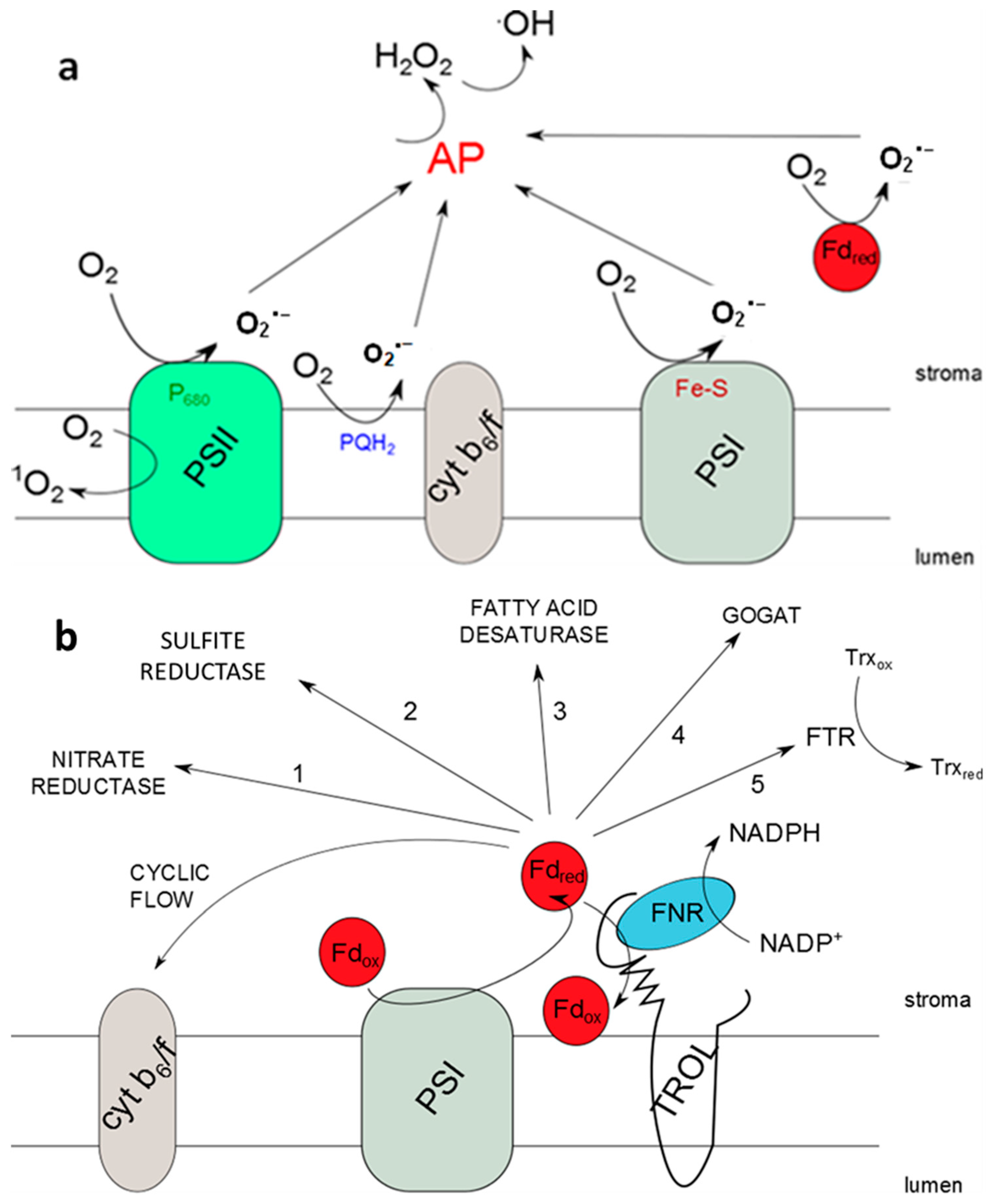
1. Introduction
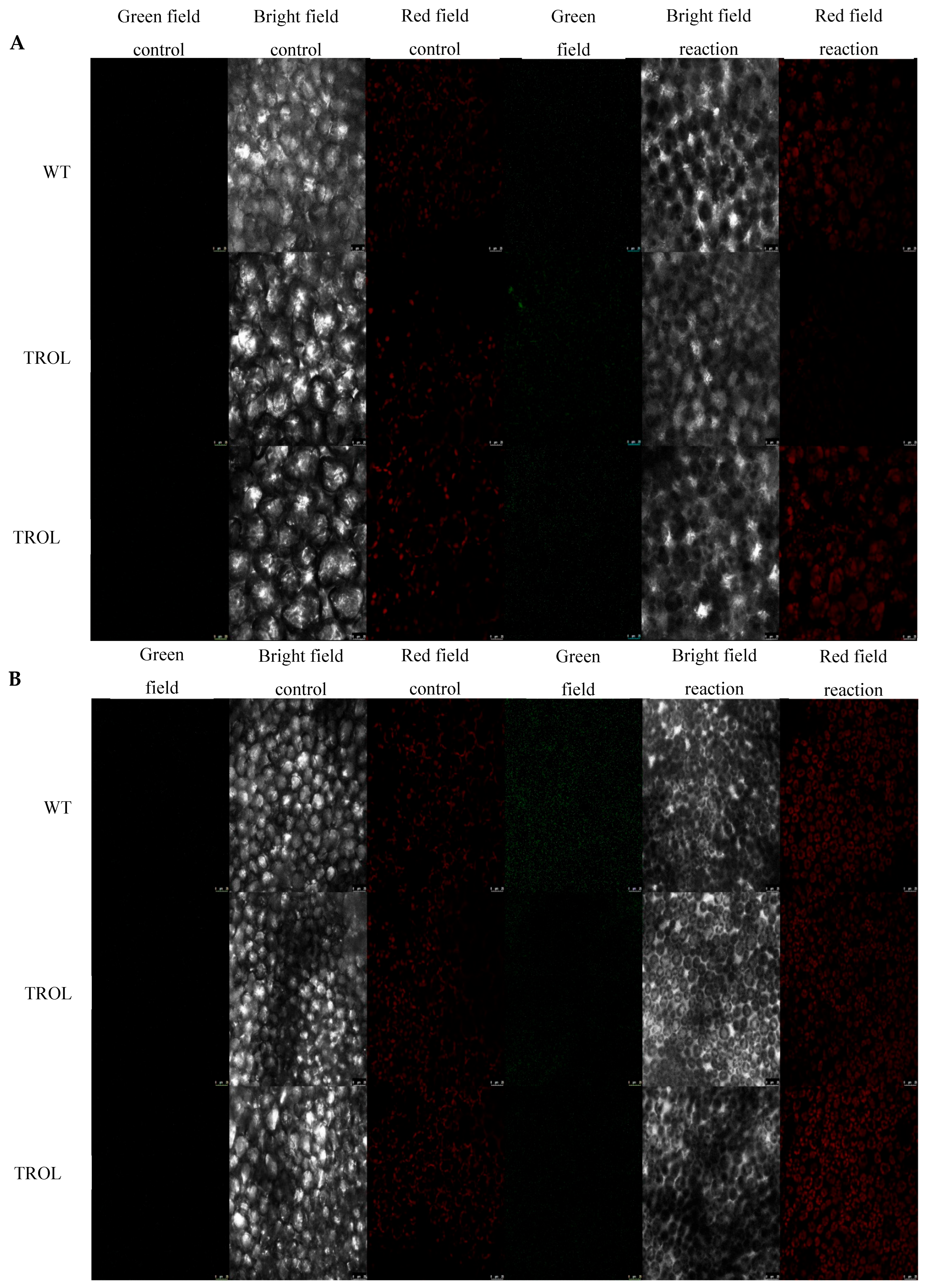
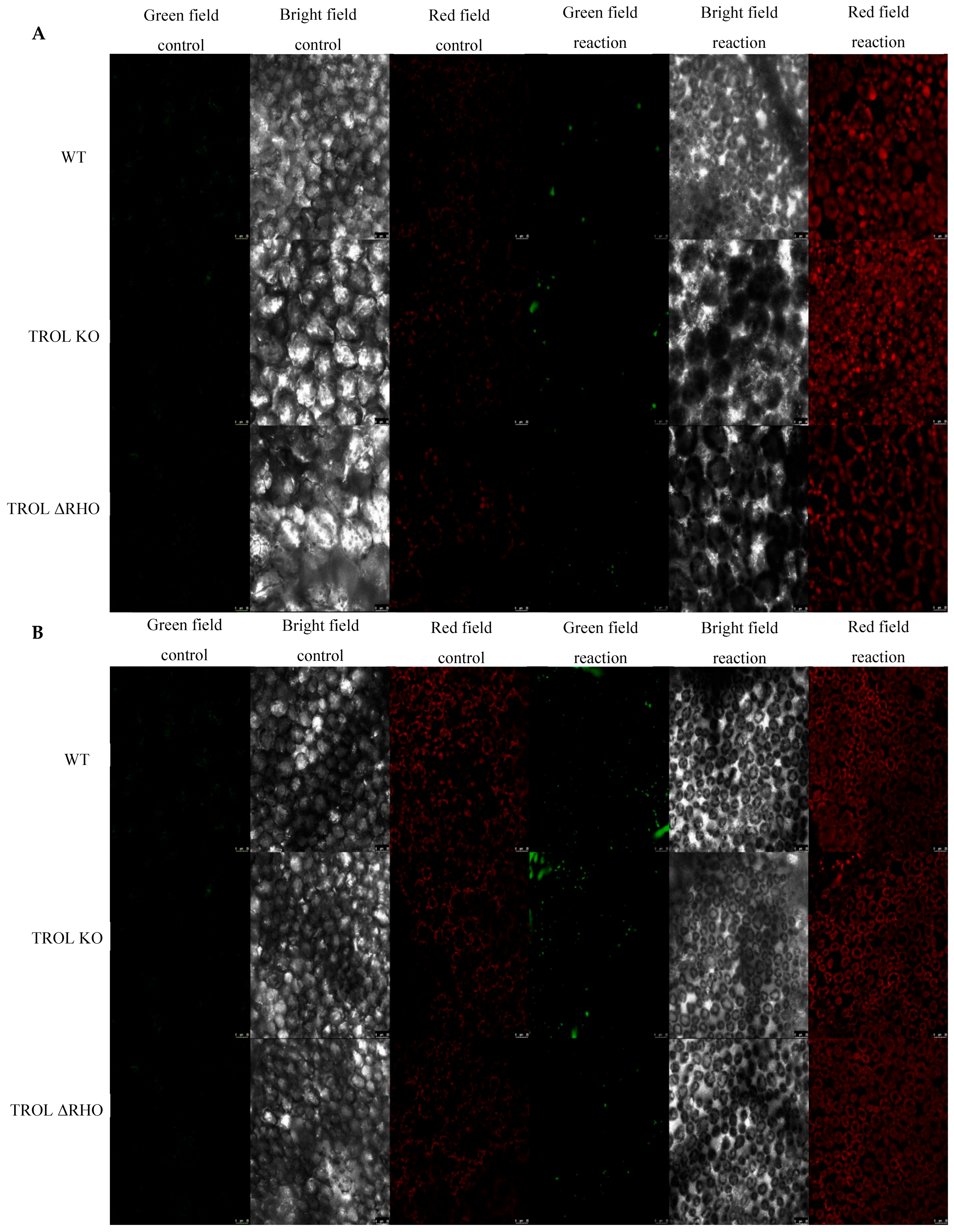
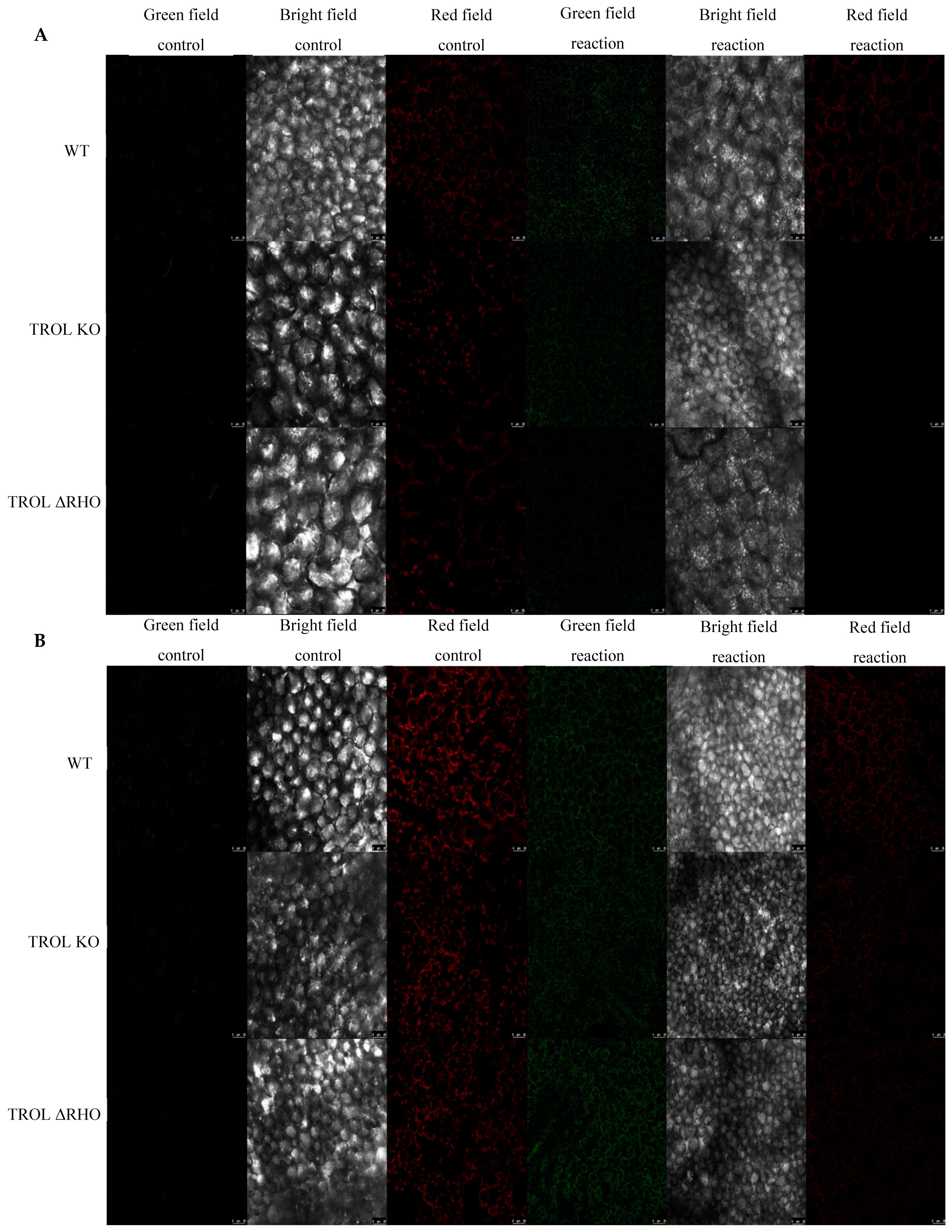
2. Results
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Chemicals
4.3. Sample Preparation
4.4. Confocal Microscopy
4.5. Image Analysis
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tanaka, A.; Makino, A. Photosynthetic research in plant science. Plant Cell Physiol. 2009, 50, 681–683. [Google Scholar] [CrossRef] [PubMed]
- Goss, T.; Hanke, G. The end of the line: Can ferredoxin and FxNADP(H) oxidoreductase determine the fate of photosynthetic electrons. Curr. Protein Pept. Sci. 2014, 15, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Benz, J.P.; Soll, J.; Bolter, B. Protein transport in organelles: The composition, function and regulation of Tic62 complex in chloroplast protein import. FEBS J. 2009, 276, 1166–1176. [Google Scholar] [CrossRef] [PubMed]
- Benz, J.P.; Stengel, A.; Lintala, M.; Lee, Y.H.; Weber, A.; Philippar, K.; Gügel, I.L.; Kaieda, S.; Ikegami, T.; Mulo, P.; et al. Arabidopsis Tic62 and ferredoxin-NADP(H) oxidoreductase form light-regulated complexes that are integrated into the chloroplast redox poise. Plant Cell 2009, 21, 3965–3983. [Google Scholar] [CrossRef] [PubMed]
- Jurić, S.; Hazler-Pilepić, K.; Tomašić, A.; Lepeduš, H.; Jelić, B.; Puthiyaveetil, S.; Bionda, T.; Vojta, L.; Allen, J.F.; Schleiff, E.; et al. Tethering of ferredoxin: NADP+ oxidoreductase to thylakoid membranes is mediated by novel chloroplast protein TROL. Plant J. 2009, 6, 783–794. [Google Scholar] [CrossRef] [PubMed]
- Alte, F.; Stengel, A.; Benz, J.P.; Petersen, E.; Soll, J.; Groll, M.; Bölter, B. Ferredoxin: NADPH oxidoreductase is recruited to thylakoids by binding to a polyproline type II helix in a pH-dependent manner. Proc. Natl. Acad. Sci. USA 2010, 107, 19260–19265. [Google Scholar] [CrossRef] [PubMed]
- Lintala, M.; Allahverdiyeva, Y.; Kangasjärvi, S.; Lehtimäki, N.; Keränen, M.; Rintamäki, E.; Aro, E.M.; Mulo, P. Comparative analysis of leaf-type ferredoxin-NADP oxidoreductase isoforms in Arabidopsis thaliana. Plant J. 2007, 57, 1103–1115. [Google Scholar] [CrossRef] [PubMed]
- Vojta, L.; Carić, D.; Cesar, V.; Antunović Dunić, J.; Lepeduš, H.; Kveder, M.; Fulgosi, H. TROL-FNR interaction reveals alternative pathways of electron partitioning in photosynthesis. Sci. Rep. 2015, 5, 10085. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Foyer, C.H. Intracellular redox compartmentation and ROS-related communication in regulation and signaling. Plant Physiol. 2016, 171, 1581–1592. [Google Scholar] [CrossRef] [PubMed]
- Vojta, L.; Horvat, L.; Fulgosi, H. Balancing chloroplast redox status-regulation of FNR binding and release. Period. Biol. 2012, 114, 25–31. [Google Scholar]
- Pantoja-Uceda, D.; López-Méndez, B.; Koshiba, S. Solution structure of the rhodanese homology domain At4g01050 (175–295) from Arabidopsis thaliana. Protein Sci. 2005, 14, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Noctor, G. Redox homeostasis and antioxidant signalling: A metabolic interface between stress perception and physiological responses. Plant Cell 2005, 17, 1866–1875. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 1, 217037. [Google Scholar] [CrossRef]
- Prasad, A.; Sedlářová, M.; Balukova, A.; Rác, M.; Pospíšil, P. Reactive oxygen species as a response to wounding: In vivo imaging in Arabidopsis thaliana. Front. Plant Sci. 2020, 10, 1660. [Google Scholar] [CrossRef] [PubMed]
- Hüner, N.P.; Bode, R.; Dahal, K.; Hollis, L.; Rosso, D.; Krol, M.; Ivanov, A.G. Chloroplast redox imbalance governs phenotypic plasticity: The “grand design of photosynthesis” revisited. Front. Plant Sci. 2012, 3, 255. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Zhang, R.; Yang, M.; Law, Y.S.; Sun, F.; Hon, N.L.; Ngai, S.M.; Lim, B.L. A balance between the activities of chloroplasts and mitochondria is crucial for optimal plant growth. Antioxidants 2021, 10, 935. [Google Scholar] [CrossRef] [PubMed]
- Kramer, D.M.; Evans, J.R. The importance of energy balance in improving photosynthetic productivity. Plant Physiol. 2011, 155, 70–78. [Google Scholar] [CrossRef] [PubMed]
- Lehtimaki, N.; Lintala, M.; Allahverdiyeva, Y.; Aro, E.M.; Mulo, P. Drought stress-induced upregulation of components involved in ferredoxin-dependent cyclic electron transfer. J. Plant Physiol. 2010, 167, 1018–1022. [Google Scholar] [CrossRef] [PubMed]
- Sandalio, L.M.; Peláez-Vico, M.A.; Molina-Moya, E.; Romero-Puertas, M.C. Peroxisomes as redox-signaling nodes in intracellular communication and stress responses. Plant Physiol. 2021, 186, 22–35. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, N.; Ceccarelli, E.A. Open questions in ferredoxin-NADP(+) reductase catalytic mechanism. Eur. J. Biochem. 2003, 270, 1900–1915. [Google Scholar] [CrossRef] [PubMed]
- Cejudo, F.J.; González, M.C.; Pérez-Ruiz, J.M. Redox regulation of chloroplast metabolism. Plant Physiol. 2021, 186, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Vojta, L.; Rac-Justament, A.; Zechmann, B.; Fulgosi, H. Thylakoid rhodanese-like protein–ferredoxin: NADP+ oxidoreductase interaction is integrated into plant redox homeostasis system. Antioxidants 2023, 12, 1838. [Google Scholar] [CrossRef] [PubMed]
- Tripathy, B.C.; Oelmüller, R. Reactive oxygen species generation and signaling in plants. Plant Signal. Behav. 2012, 7, 1621–1633. [Google Scholar] [CrossRef] [PubMed]
- Sedlářová, M.; Luhová, L. Re-evaluation of imaging methods of reactive oxygen and nitrogen species in plants and fungi: Influence of cell wall composition. Front. Physiol. 2017, 8, 826. [Google Scholar] [CrossRef] [PubMed]
- Exposito-Rodriguez, M.; Laissue, P.P.; Yvon-Durocher, G.; Smirnoff, N.; Mullineaux, P.M. Photosynthesis-dependent H2O2 transfer from chloroplasts to nuclei provides a high-light signalling mechanism. Nat. Commun. 2017, 8, 49. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Hanke, G. ROS production and signalling in chloroplasts: Cornerstones and evolving concepts. Plant J. 2022, 111, 642–661. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R.; Vanderauwera, S.; Suzuki, N.; Miller, G.A.D.; Tognetti, V.B.; Vandepoele, K.; Van Breusegem, F. ROS signaling: The new wave? Trends Plant Sci. 2011, 16, 300–309. [Google Scholar] [CrossRef] [PubMed]
- Quan, L.J.; Zhang, B.; Shi, W.W.; Li, H.Y. Hydrogen peroxide in plants: A versatile molecule of the reactive oxygen species network. J. Integr. Plant Biol. 2008, 50, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Duanghathaipornsuk, S.; Farrell, E.J.; Alba-Rubio, A.C.; Zelenay, P.; Kim, D.S. Detection technologies for reactive oxygen species: Fluorescence and electrochemical methods and their applications. Biosensors 2021, 11, 30. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Jockusch, S.; Zhou, Z.; Blanchard, S.C. The contribution of reactive oxygen species to the photobleaching of organic fluorophores. Photochem. Photobiol. 2014, 90, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Winterbourn, C.C. The challenges of using fluorescent probes to detect and quantify specific reactive oxygen species in living cells. BBA-Gen. Subj. 2014, 1840, 730–738. [Google Scholar] [CrossRef] [PubMed]
- Pospíšil, P. Production of reactive oxygen species by photosystem II as a response to light and temperature stress. Front. Plant Sci. 2016, 7, 1950. [Google Scholar] [CrossRef] [PubMed]
- Goltsev, V.; Zaharieva, I.; Chernev, P.; Strasser, R. Delayed chlorophyll fluorescence as a monitor for physiological state of photosynthetic apparatus. Biotechnol. Biotechnol. Equip. 2009, 23, 452–457. [Google Scholar] [CrossRef]
- Goltsev, V.; Zaharieva, I.; Chernev, P.; Strasser, R.J. Delayed fluorescence in photosynthesis. Photosynth. Res. 2009, 101, 217–232. [Google Scholar] [CrossRef] [PubMed]
- Peterkovic, Z.; Upadhya, A.; Perrella, C.; Bajraktarevic, A.; Bautista Gonzalez, R.E.; Lim, M.; Dholakia, K. Optimizing image capture for low-light widefield quantitative fluorescence microscopy. APL Photonics 2025, 10, 031102. [Google Scholar] [CrossRef]
- Icha, J.; Weber, M.; Waters, J.C.; Norden, C. Phototoxicity in live fluorescence microscopy, and how to avoid it. BioEssays 2017, 39, 1700003. [Google Scholar] [CrossRef] [PubMed]
- Hoebe, R.A.; Van Der Voort, H.T.M.; Stap, J.; Van Noorden, C.J.F.; Manders, E.M.M. Quantitative determination of the reduction of phototoxicity and photobleaching by controlled light exposure microscopy. J. Microsc. 2008, 231, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Cramer, W.A.; Yamashita, E.; Baniulis, D.; Saif Hasan, S. The cytochrome b6f complex of oxygenic photosynthesis. In Encyclopedia of Inorganic and Bioinorganic Chemistry; John Wiley & Sons: Hoboken, NJ, USA, 2011. [Google Scholar]
- Guadagno, C.R.; Ewers, B.E.; Weinig, C. Circadian rhythms and redox state in plants: Till stress do us part. Front. Plant Sci. 2018, 9, 247. [Google Scholar] [CrossRef] [PubMed]
- Considine, M.J.; Foyer, C.H. Redox regulation of plant development. Antioxid. Redox Signal. 2014, 21, 1305–1326. [Google Scholar] [CrossRef] [PubMed]
- Roach, T.; Krieger-Liszkay, A. Regulation of photosynthetic electron transport and photoinhibition. Curr. Protein Pept. Sci. 2014, 15, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Fichman, Y.; Miller, G.; Mittler, R. Whole-plant live imaging of reactive oxygen species. Mol. Plant 2019, 12, 1203–1210. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Ming, Y.; Wang, H.B.; Jin, H.L. Strategies for Adaptation to High Light in Plants. Abiotech 2024, 5, 381–393. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Lu, M.; Wang, Y.; Wang, Y.; Liu, Z.; Chen, S. Response mechanism of plants to drought stress. Horticulturae 2021, 7, 50. [Google Scholar] [CrossRef]
- Qiao, M.; Hong, C.; Jiao, Y.; Hou, S.; Gao, H. Impacts of drought on photosynthesis in major food crops and the related mechanisms of plant responses to drought. Plants 2024, 13, 1808. [Google Scholar] [CrossRef] [PubMed]
- Hald, S.; Nandha, B.; Gallois, P.; Johnson, G.N. Feedback regulation of photosynthetic electron transport by NADP (H) redox poise. Biochim. Biophys. Acta Bioenerg. 2008, 1777, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Golding, A.J.; Johnson, G.N. Down-regulation of linear and activation of cyclic electron transport during drought. Planta 2023, 218, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Shi, Q.; Liu, N.Y.; Zhang, S.B.; Huang, W. Drought stress delays photosynthetic induction and accelerates photoinhibition under short-term fluctuating light in tomato. Plant Physiol. Biochem. 2023, 196, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Bashir, N.; Athar, H.U.R.; Kalaji, H.M.; Wróbel, J.; Mahmood, S.; Zafar, Z.U.; Ashraf, M. Is photoprotection of PSII one of the key mechanisms for drought tolerance in maize? Int. J. Mol. Sci. 2021, 22, 13490. [Google Scholar] [CrossRef] [PubMed]
- Lima-Melo, Y.; Alencar, V.T.; Lobo, A.K.; Sousa, R.H.; Tikkanen, M.; Aro, E.M.; Gollan, P.J. Photoinhibition of photosystem I provides oxidative protection during imbalanced photosynthetic electron transport in Arabidopsis thaliana. Front. Plant Sci. 2019, 10, 916. [Google Scholar] [CrossRef] [PubMed]
- Lima-Melo, Y.; Gollan, P.J.; Tikanen, M.; Silveira, J.A.; Aro, E.M. Consequences of photosystem-I damage and repair on photosynthesis and carbon use in Arabidopsis thaliana. Plant J. 2019, 97, 1061–1072. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Li, G.; Sun, H.; Ma, L.; Guo, Y.; Zhao, Z.; Mei, L. Effects of drought stress on photosynthesis and photosynthetic electron transport chain in young apple tree leaves. Biol. Open 2018, 7, 035279. [Google Scholar] [CrossRef] [PubMed]
- Fulgosi, H.; Vojta, L. Tweaking photosynthesis: FNR-TROL interaction as potential target for crop fortification. Front. Plant Sci. 2020, 11, 318. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.F. Cyclic, pseudocyclic and noncyclic photophosphorylation: New links in the chain. Trends Plant Sci. 2003, 8, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Ullah, F.; Zhou, D.X.; Yi, M.; Zhao, Y. Mechanisms of ROS regulation of plant development and stress responses. Front. Plant Sci. 2019, 10, 800. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Quesada, M.J.; Traverso, J.Á.; Alché, J.D.D. NADPH oxidase-dependent superoxide production in plant reproductive tissues. Front. Plant Sci. 2016, 7, 359. [Google Scholar] [CrossRef] [PubMed]
- Bowler, C.; Van Camp, W.; Van Montagu, M.; Inzé, D.; Asada, K. Superoxide dismutase in plants. CRC Crit. Rev. Plant Sci. 1994, 13, 99–218. [Google Scholar] [CrossRef]
- Zandalinas, S.I.; Mittler, R. ROS-induced ROS release in plant and animal cells. Free Radic. Biol. Med. 2018, 122, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Palatnik, J.F.; Valle, E.M.; Carillo, N. Oxidative stress causes ferredoxin-NADP+ reductase solubilization from the thylakoid membranes in methyl viologen-treated plants. Plant Physiol. 1997, 115, 1721–1727. [Google Scholar] [CrossRef] [PubMed]
- Vojta, L.; Fulgosi, H. Topology of TROL protein in thylakoid membranes of Arabidopsis thaliana. Physiol. Plant. 2019, 166, 300–308. [Google Scholar] [CrossRef] [PubMed]
- Vojta, L.; Fulgosi, H. Energy conductance from thylakoid complexes to stromal reducing equivalents. In Advances in Photosynthesis-Fundamental Aspects; In Tech: London, UK, 2012; pp. 175–190. [Google Scholar]
- Dumančić, E.; Fulgosi, H. The Role of TROL-FNR Protein Interaction in Arabidopsis thaliana (L.) Heynh. In Response to Drought Stress. Faculty of Science, University of Zagreb: Zagreb, Croatia, 2026; manuscript in preparation. [Google Scholar]
- Smirnoff, N.; Arnaud, D. Hydrogen peroxide metabolism and functions in plants. New Phytol. 2019, 221, 1197–1214. [Google Scholar] [CrossRef] [PubMed]
- Černý, M.; Habanova, H.; Berka, M.; Luklova, M.; Brzobohatý, B. Hydrogen peroxide: Its role in plant biology and crosstalk with signalling networks. Int. J. Mol. Sci. 2018, 19, 2812. [Google Scholar] [CrossRef] [PubMed]
- Niu, L.; Liao, W. Hydrogen peroxide signaling in plant development and abiotic responses: Crosstalk with nitric oxide and calcium. Front. Plant Sci. 2016, 7, 230. [Google Scholar] [CrossRef] [PubMed]
- Rac, A. Molecular Analysis of the FNR-TROL Interaction Elements. Ph.D. Thesis, University of Zagreb, Faculty of Science, Department of Biology, Zagreb, Croatia, 2021. [Google Scholar]
- Dogra, V.; Kim, C. Singlet oxygen metabolism: From genesis to signaling. Front. Plant Sci. 2020, 10, 1640. [Google Scholar] [CrossRef] [PubMed]
- Laloi, C.; Havaux, M. Key players of singlet oxygen-induced cell death in plants. Front. Plant Sci. 2015, 6, 39. [Google Scholar] [CrossRef] [PubMed]
- Triantaphylidès, C.; Havaux, M. Singlet oxygen in plants: Production, detoxification and signaling. Trends Plant Sci. 2009, 14, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Neill, S.J.; Hancock, J.T.; Wilson, I.D. Reactive Oxygen Species, Nitric Oxide, and Signal Crosstalk. In Signal Crosstalk in Plant Stress Responses; Wiley-Blackwell: Ames, IA, USA, 2009; pp. 136–160. [Google Scholar]
- Neill, S.; Barros, R.; Bright, J.; Desikan, R.; Hancock, J.; Harrison, J.; Wilson, I. Nitric oxide, stomatal closure, and abiotic stress. J. Exp. Bot. 2008, 59, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Jiang, M.; Zhang, J.; Ding, H.; Xu, S.; Hu, X.; Tan, M. Nitric oxide induced by hydrogen peroxide mediates abscisic acid-induced activation of the mitogen-activated protein kinase cascade involved in antioxidant defense in maize leaves. New Phytol. 2007, 175, 36–50. [Google Scholar] [CrossRef] [PubMed]
- De Pinto, M.C.; Paradiso, A.; Leonetti, P.; De Gara, L. Hydrogen peroxide, nitric oxide and cytosolic ascorbate peroxidase at the crossroad between defence and cell death. Plant J. 2006, 48, 784–795. [Google Scholar] [CrossRef] [PubMed]
- Klotz, L.O. Oxidant-induced signaling: Effects of peroxynitrite and singlet oxygen. Biol. Chem. 2002, 383, 443–456. [Google Scholar] [CrossRef] [PubMed]
- Schafer, F.Q.; Buettner, G.R. Redox environment of the cell as viewed through the redox state of the glutathione disulfide/glutathione couple. Free Radic. Biol. Med. 2001, 30, 1191–1212. [Google Scholar] [CrossRef] [PubMed]
- Gadjev, I.; Vanderauwera, S.; Gechev, T.S.; Laloi, C.; Minkov, I.N.; Shulaev, V.; Van Breusegem, F. Transcriptomic footprints disclose specificity of reactive oxygen species signaling in Arabidopsis. Plant Physiol. 2006, 141, 436–445. [Google Scholar] [CrossRef] [PubMed]
- Zaninotto, F.; Camera, S.L.; Polverari, A.; Delledonne, M. Cross talk between reactive nitrogen and oxygen species during the hypersensitive disease resistance response. Plant Physiol. 2006, 141, 379–383. [Google Scholar] [CrossRef] [PubMed]
- Jurić, S. The Role of the Gene Product At4g01050 in the Regulation of Photosynthesis in Arabidopsis thaliana (L.) Heynh. Ph.D. Thesis, University of Zagreb, Faculty of Science, Department of Biology, Zagreb, Croatia, 2010. [Google Scholar]
- Maynard, D.N.; Hochmuth, G.J. Knott’s Handbook for Vegetable Growers; John Wiley & Sons: Hoboken, NJ, USA, 2006; Volume 5, pp. 143–248. [Google Scholar]
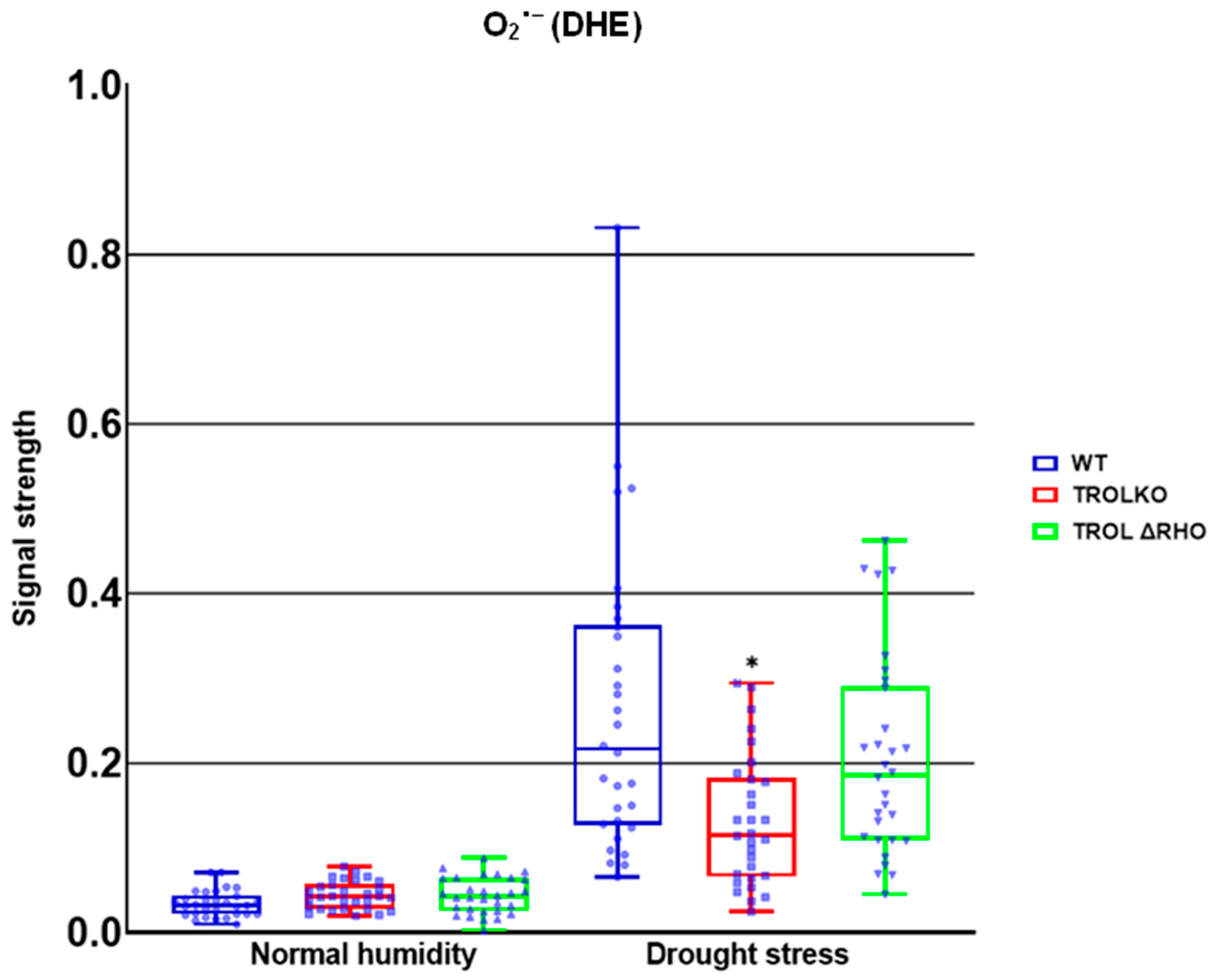
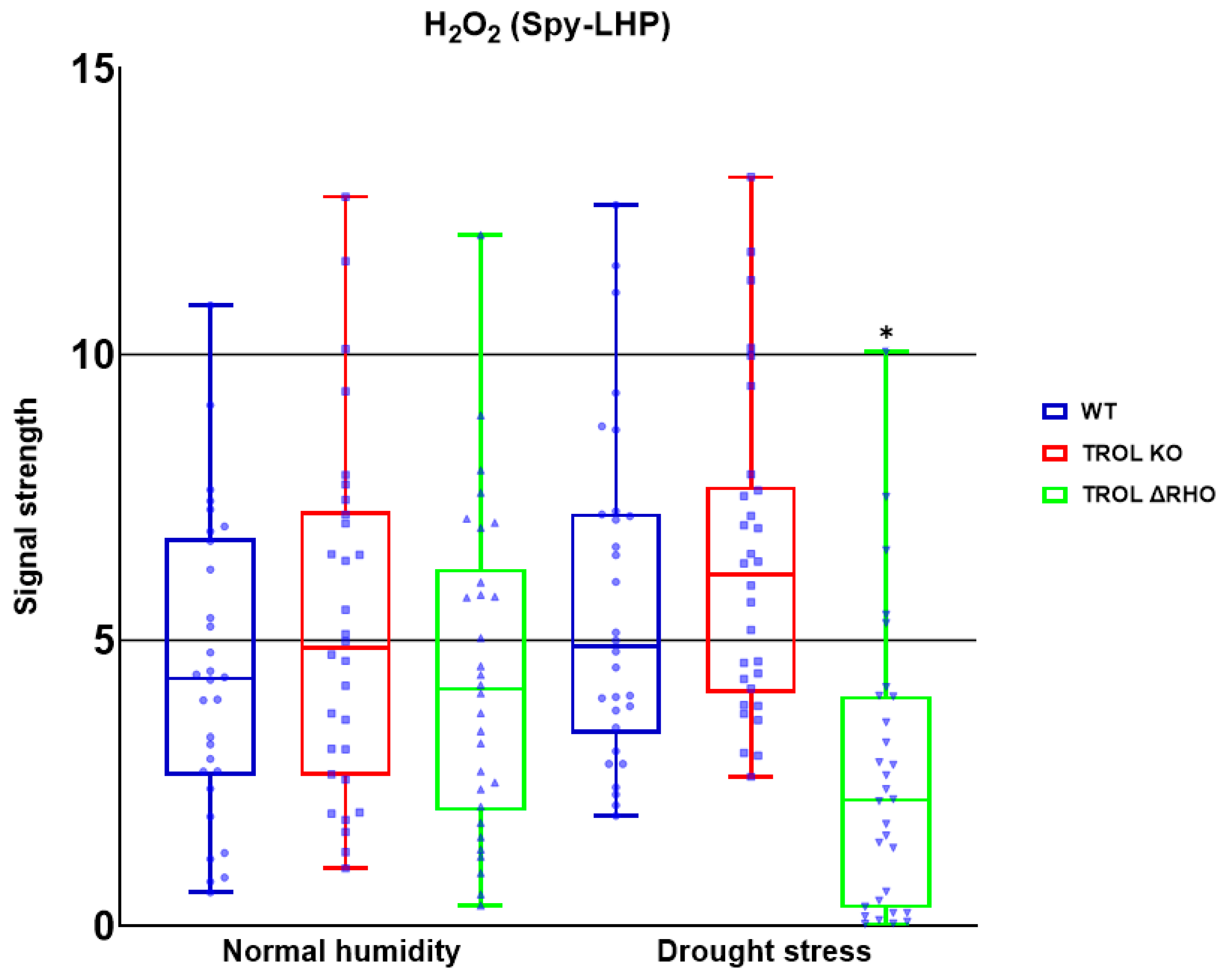
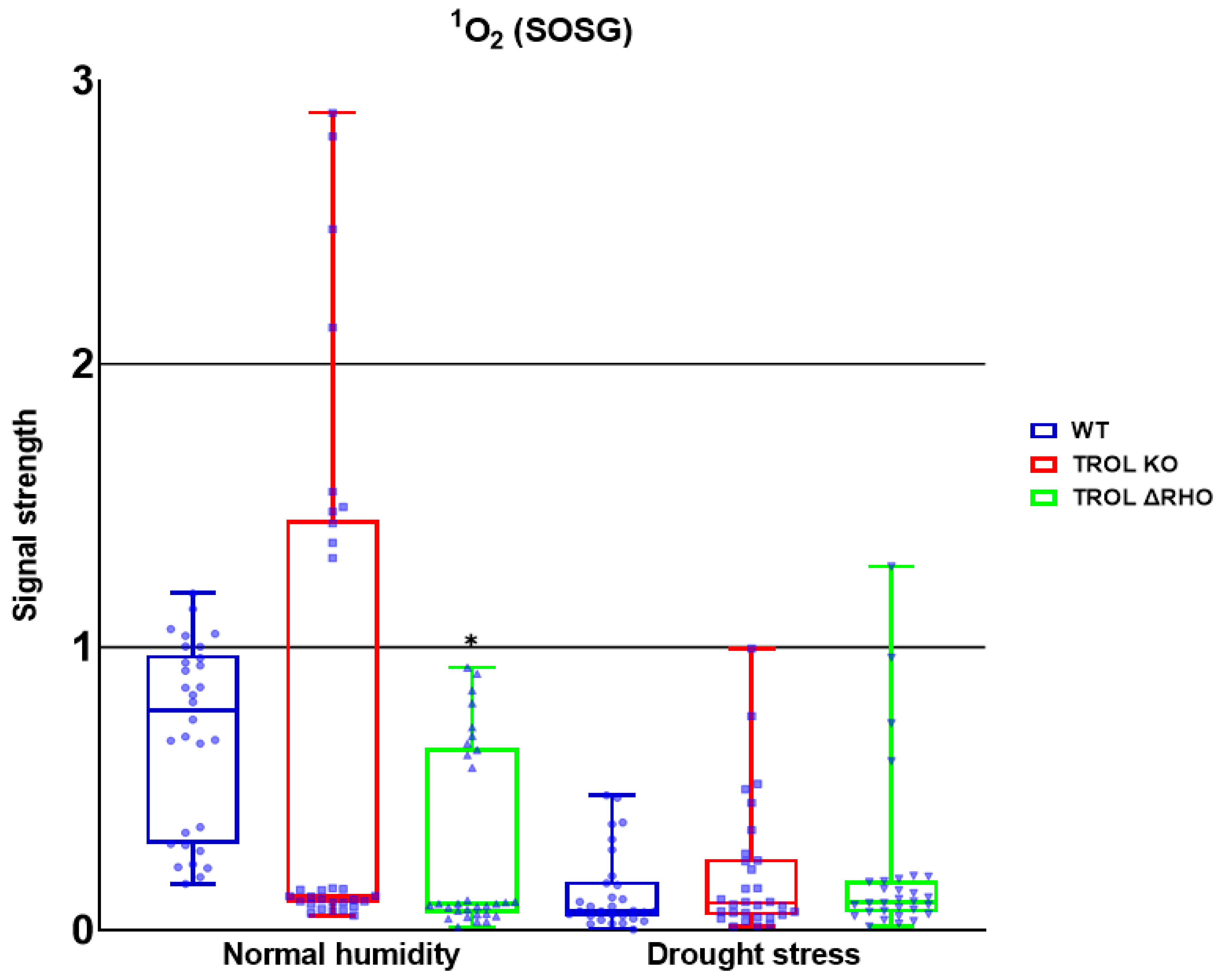
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dumančić, E.; Vojta, L.; Fulgosi, H. Confocal Laser Scanning Microscopy of Light-Independent ROS in Arabidopsis thaliana (L.) Heynh. TROL-FNR Mutants. Int. J. Mol. Sci. 2025, 26, 7000. https://doi.org/10.3390/ijms26147000
Dumančić E, Vojta L, Fulgosi H. Confocal Laser Scanning Microscopy of Light-Independent ROS in Arabidopsis thaliana (L.) Heynh. TROL-FNR Mutants. International Journal of Molecular Sciences. 2025; 26(14):7000. https://doi.org/10.3390/ijms26147000
Chicago/Turabian StyleDumančić, Ena, Lea Vojta, and Hrvoje Fulgosi. 2025. "Confocal Laser Scanning Microscopy of Light-Independent ROS in Arabidopsis thaliana (L.) Heynh. TROL-FNR Mutants" International Journal of Molecular Sciences 26, no. 14: 7000. https://doi.org/10.3390/ijms26147000
APA StyleDumančić, E., Vojta, L., & Fulgosi, H. (2025). Confocal Laser Scanning Microscopy of Light-Independent ROS in Arabidopsis thaliana (L.) Heynh. TROL-FNR Mutants. International Journal of Molecular Sciences, 26(14), 7000. https://doi.org/10.3390/ijms26147000