

Exploring the Regulatory Mechanism of Total Alkaloids from Portulaca oleracea L. in UC Treatment Based on Network Pharmacology


Abstract
1. Introduction
2. Results
2.1. Identification of Potential Therapeutic Targets of POAs
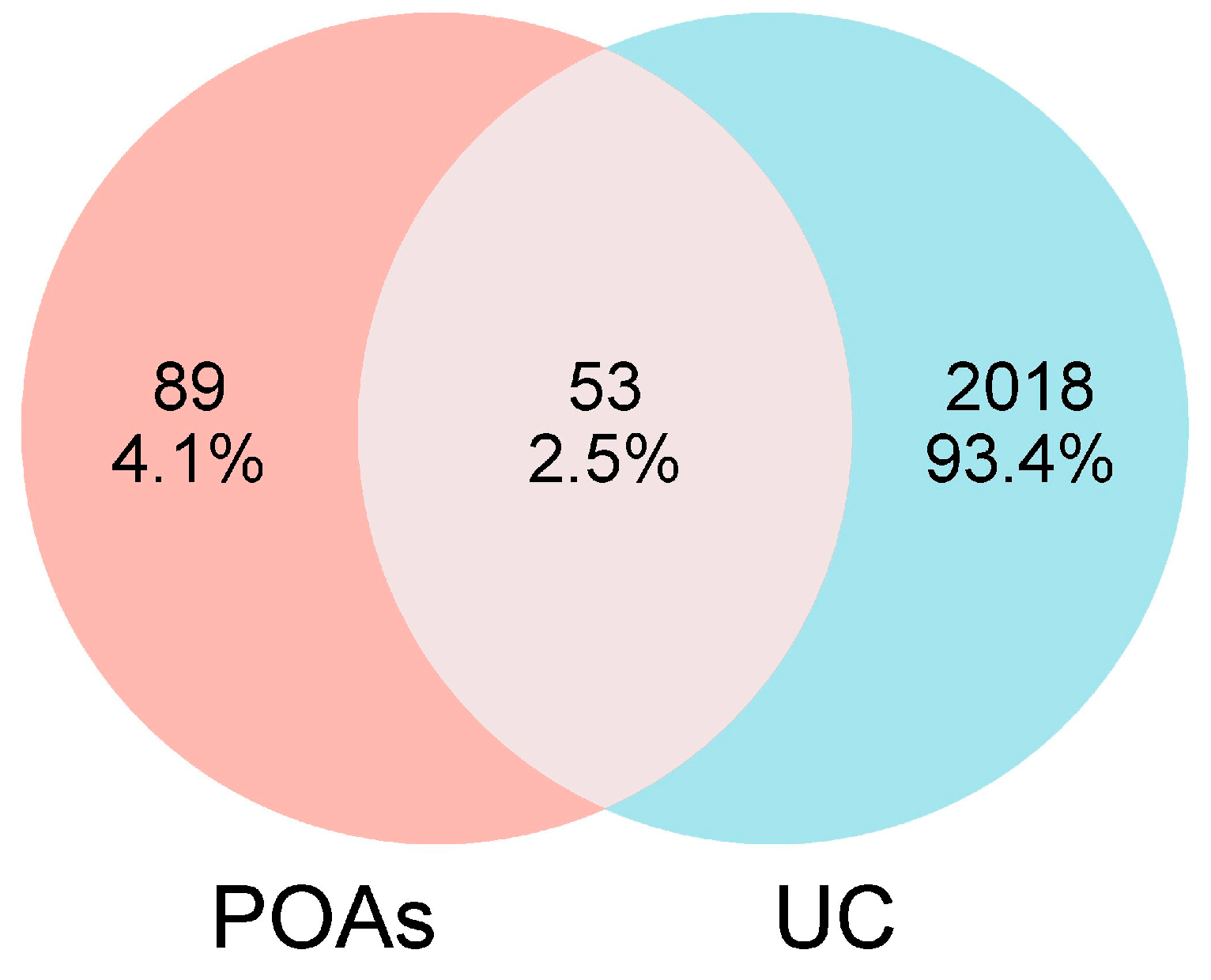
2.2. Identification of UC-Related Targets and Overlapping Targets
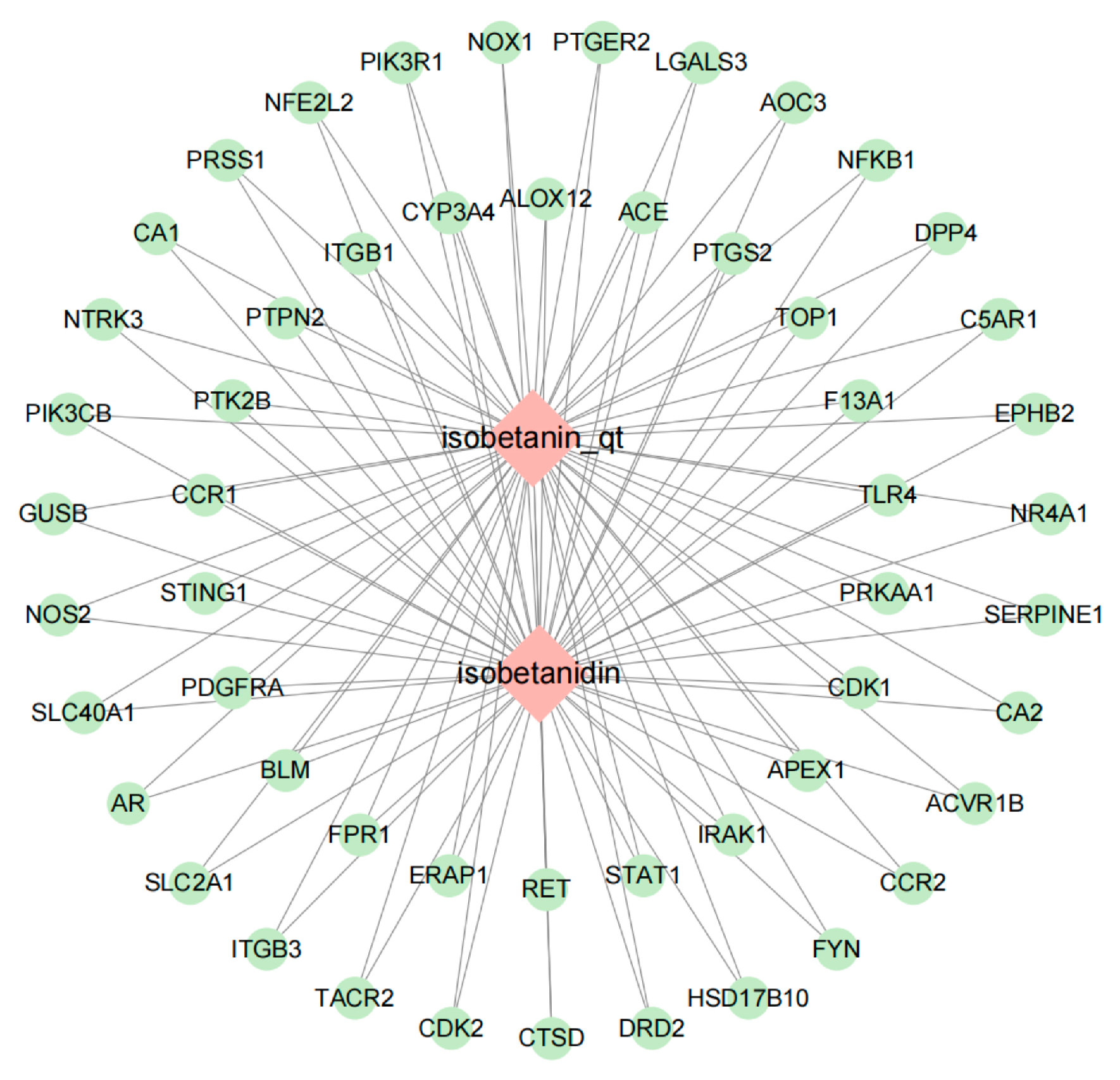
2.3. Network Construction and Analysis of “POAs—Intersection Targets”
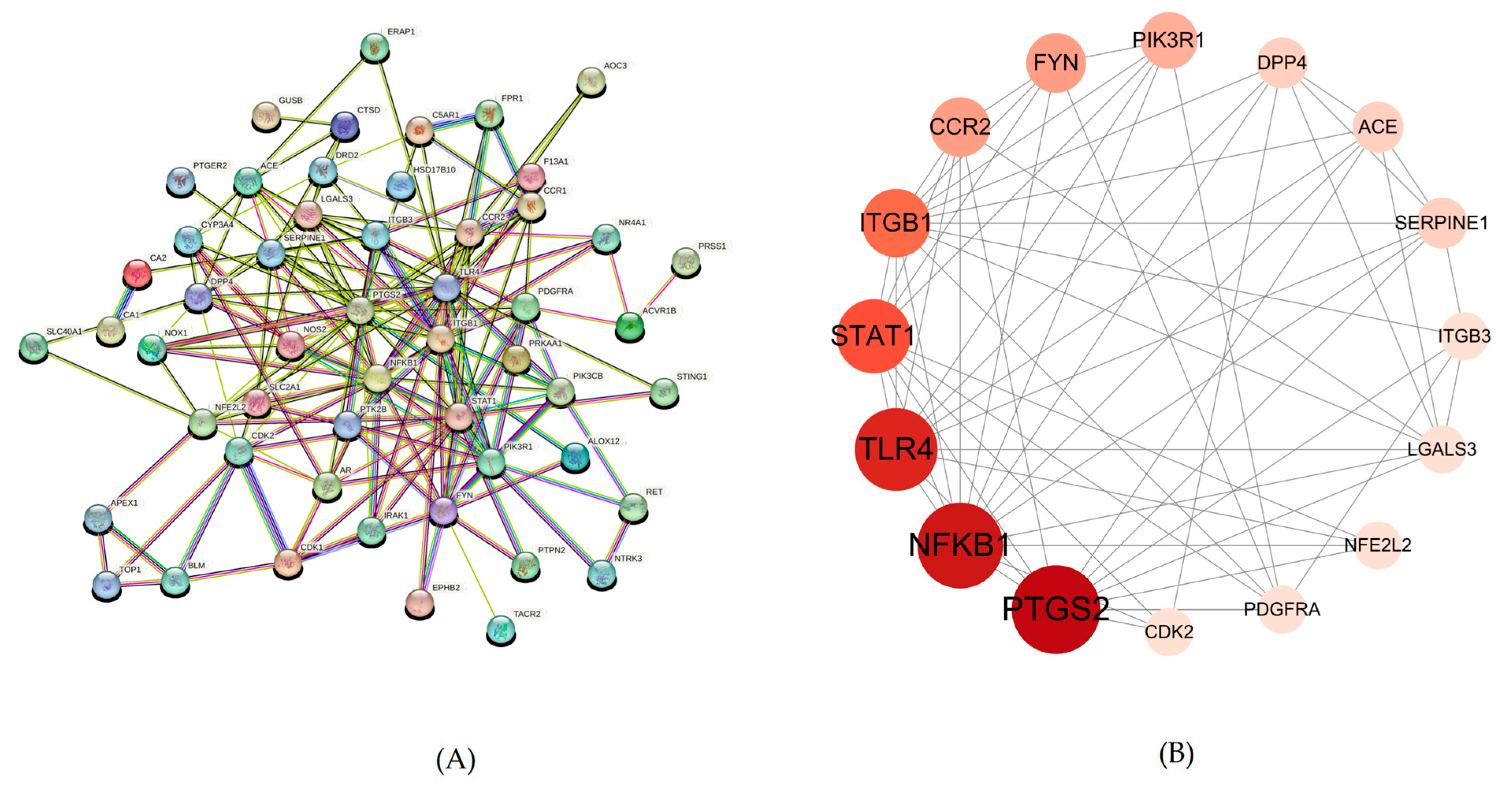
2.4. Protein–Protein Interaction (PPI) Network Analysis
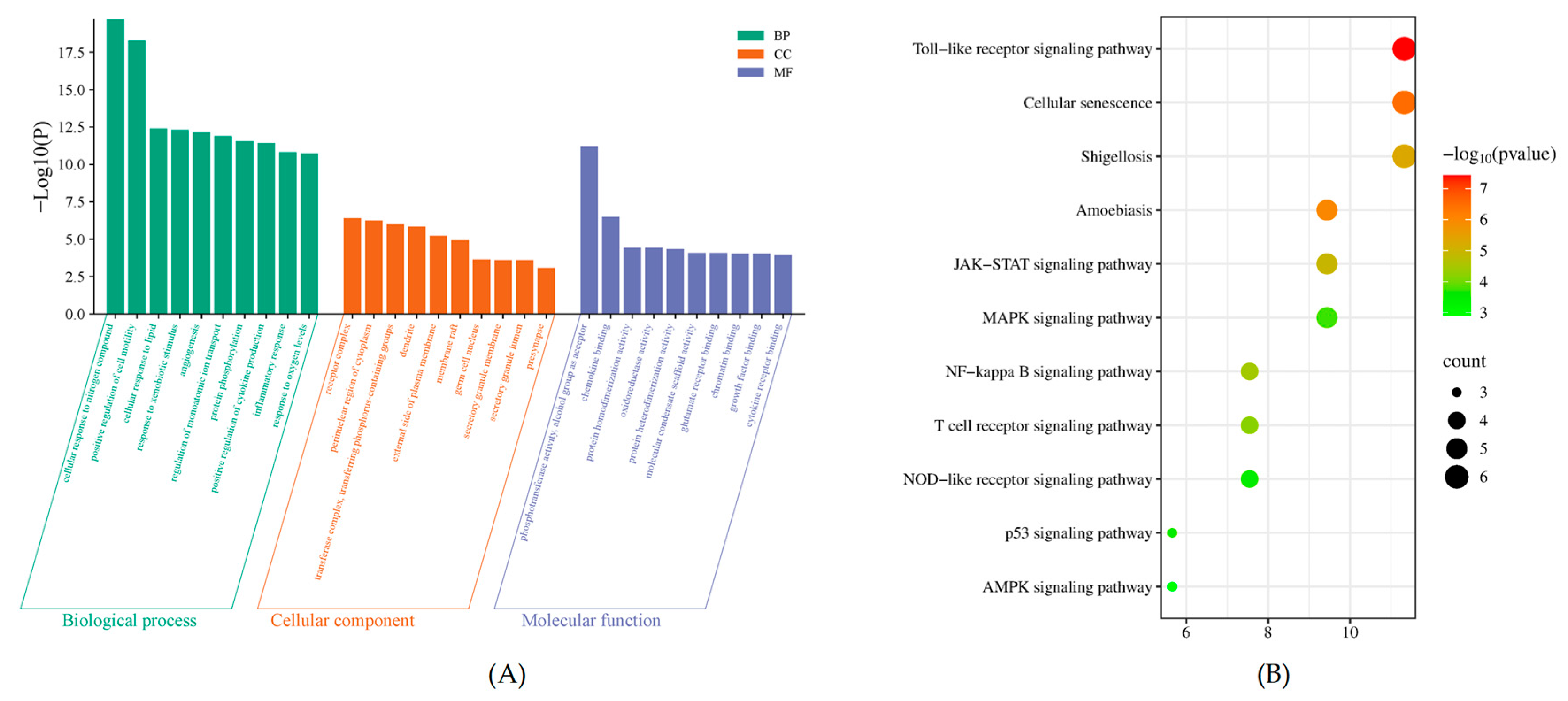
2.5. Gene Ontology (GO) Function and Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Enrichment Analysis of Intersection Targets
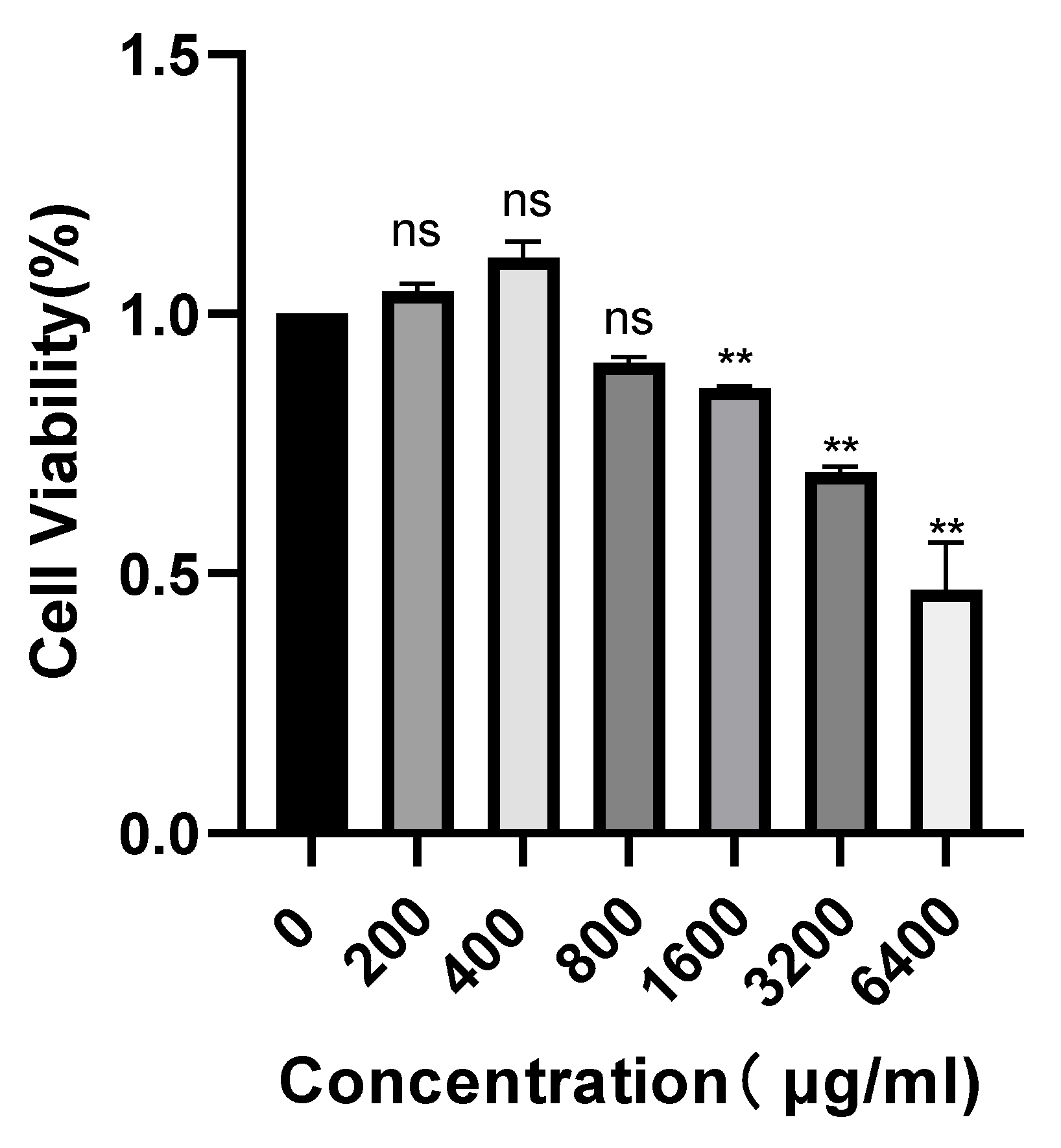
2.6. Effect of POAs on Caco-2 Cell Viability
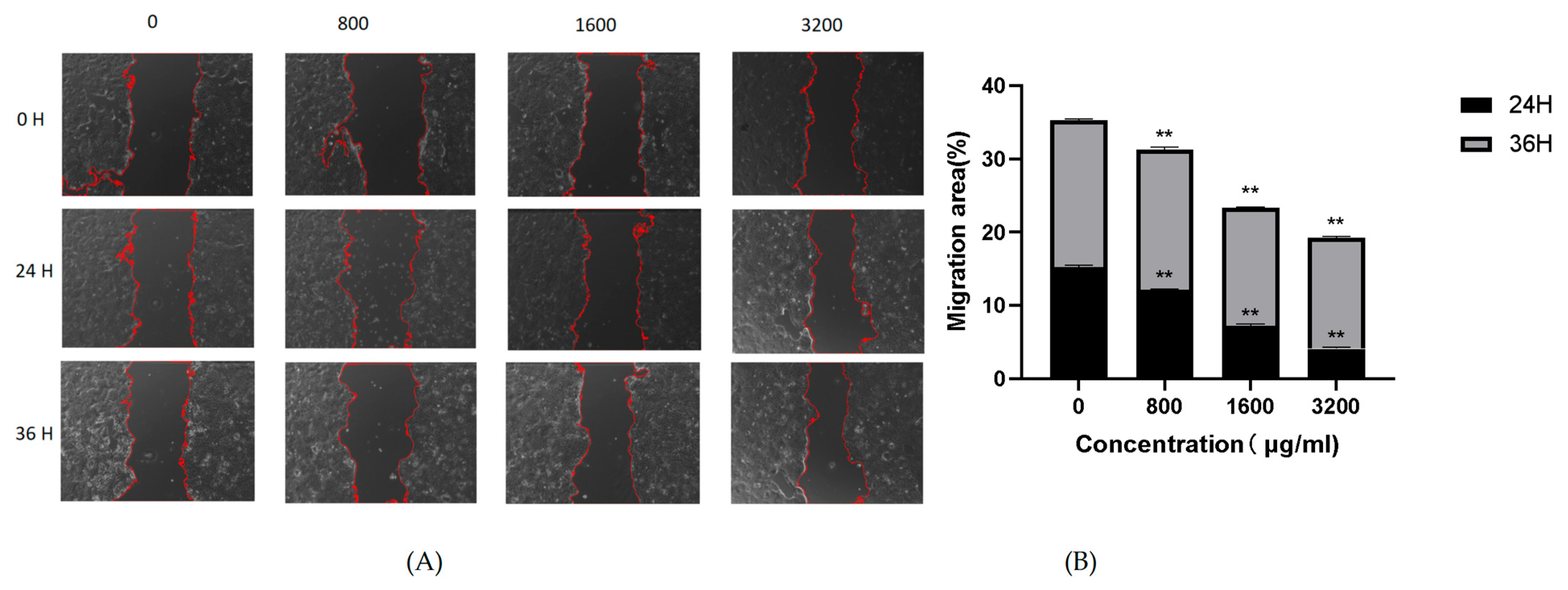
2.7. Effect of POAs on Caco-2 Cell Migration
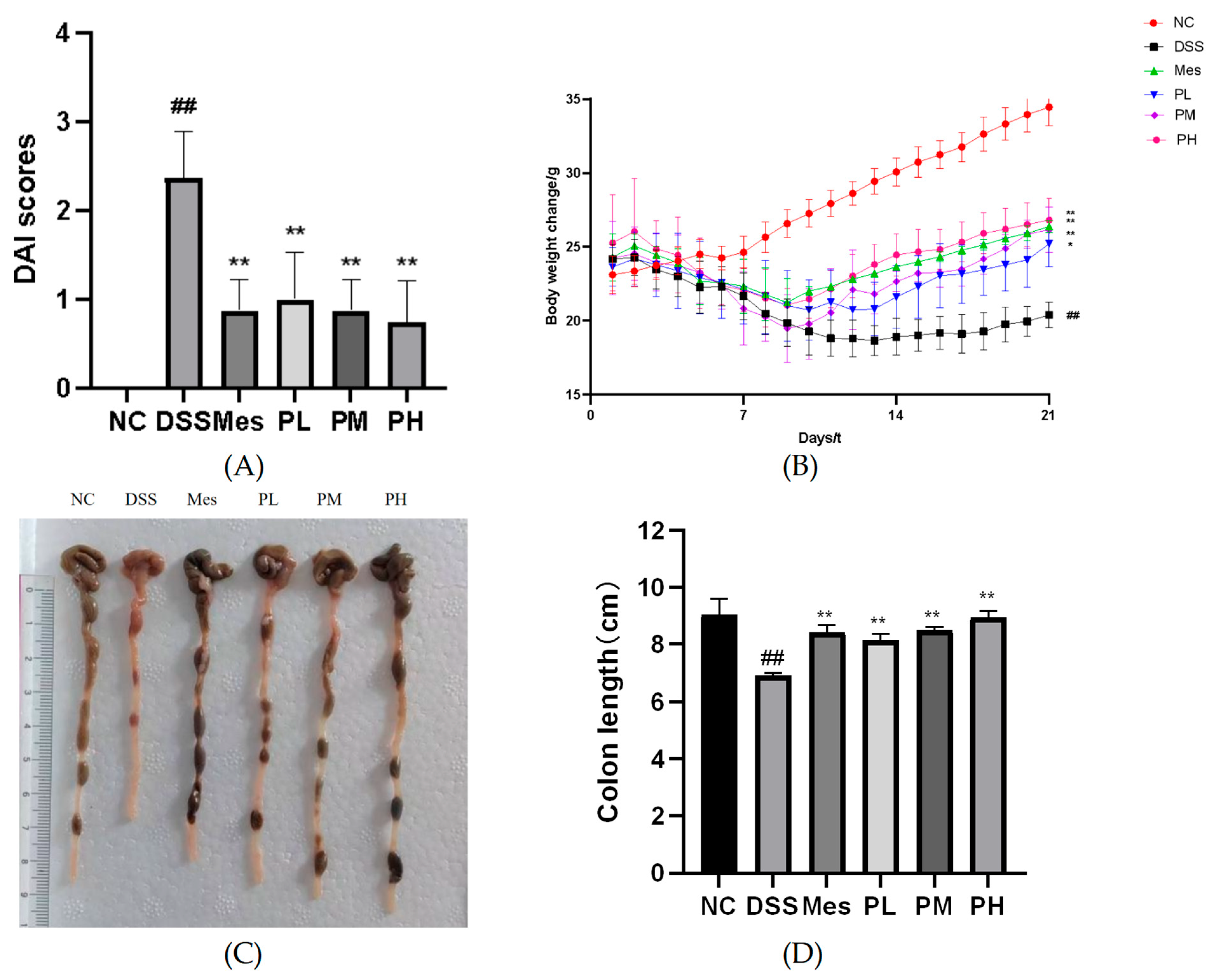
2.8. Effects of POAs on DAI Scores of UC Mice
2.9. Effects of POAs on Body Weight in UC Mice
2.10. Effect of POAs on Colon Length in UC Mice
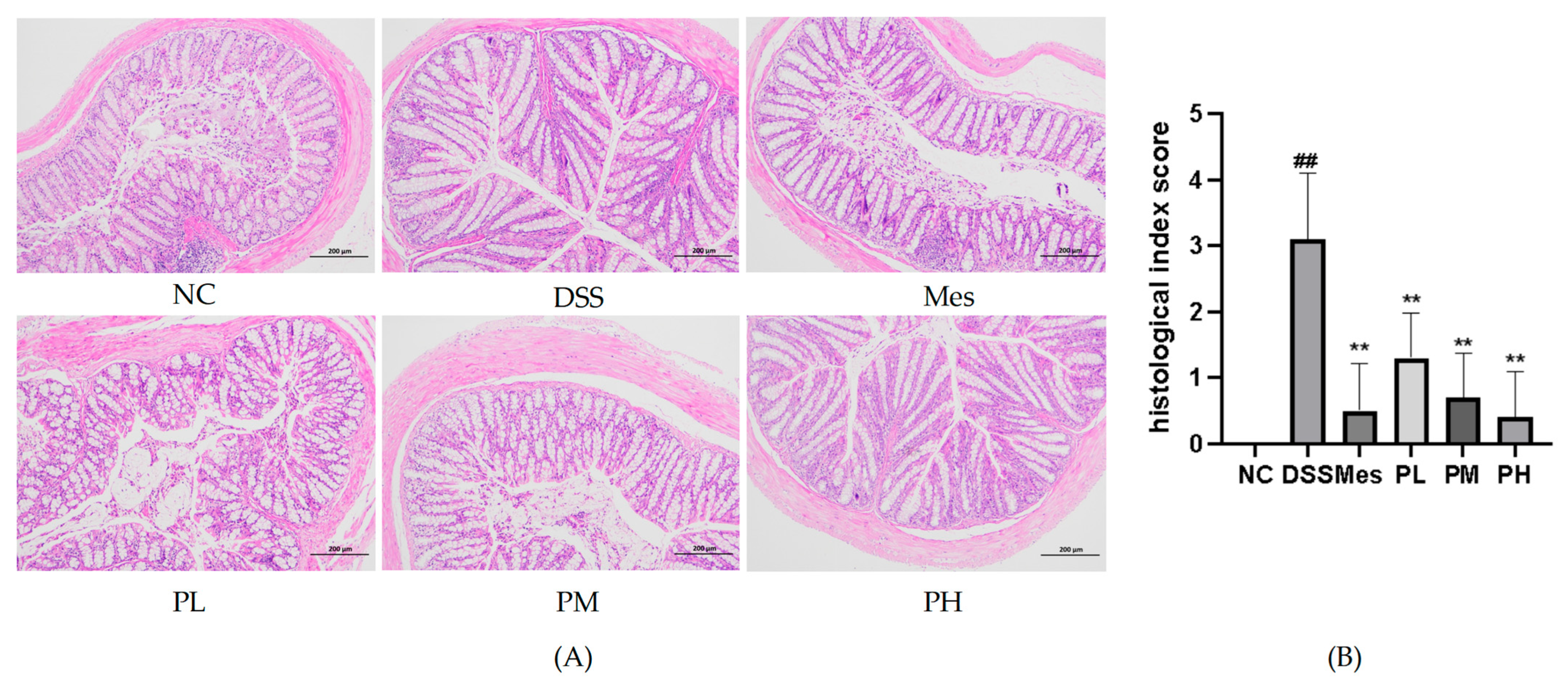
2.11. Effect of POAs on the Histopathological Changes in Colon Tissue of UC Mice
2.12. Effect of POAs on Serum Levels of Inflammatory Factors in UC Mice
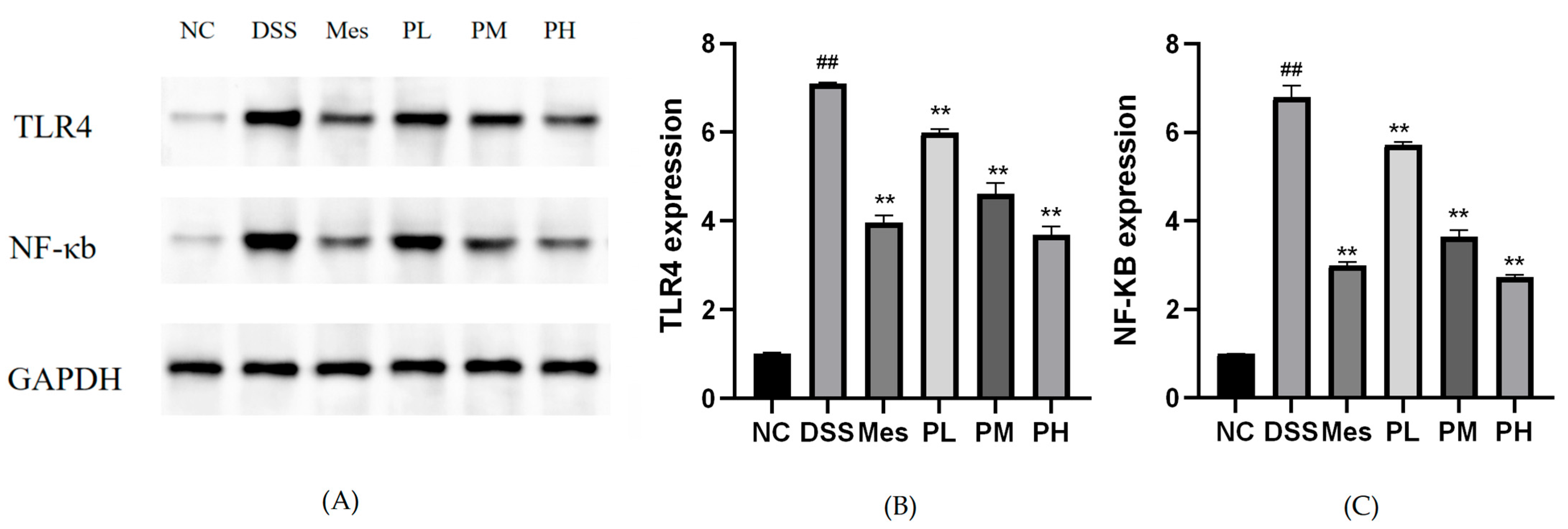
2.13. Effect of POAs on TLR4/NF-κB Protein Expression in Colon Tissues of UC Mice
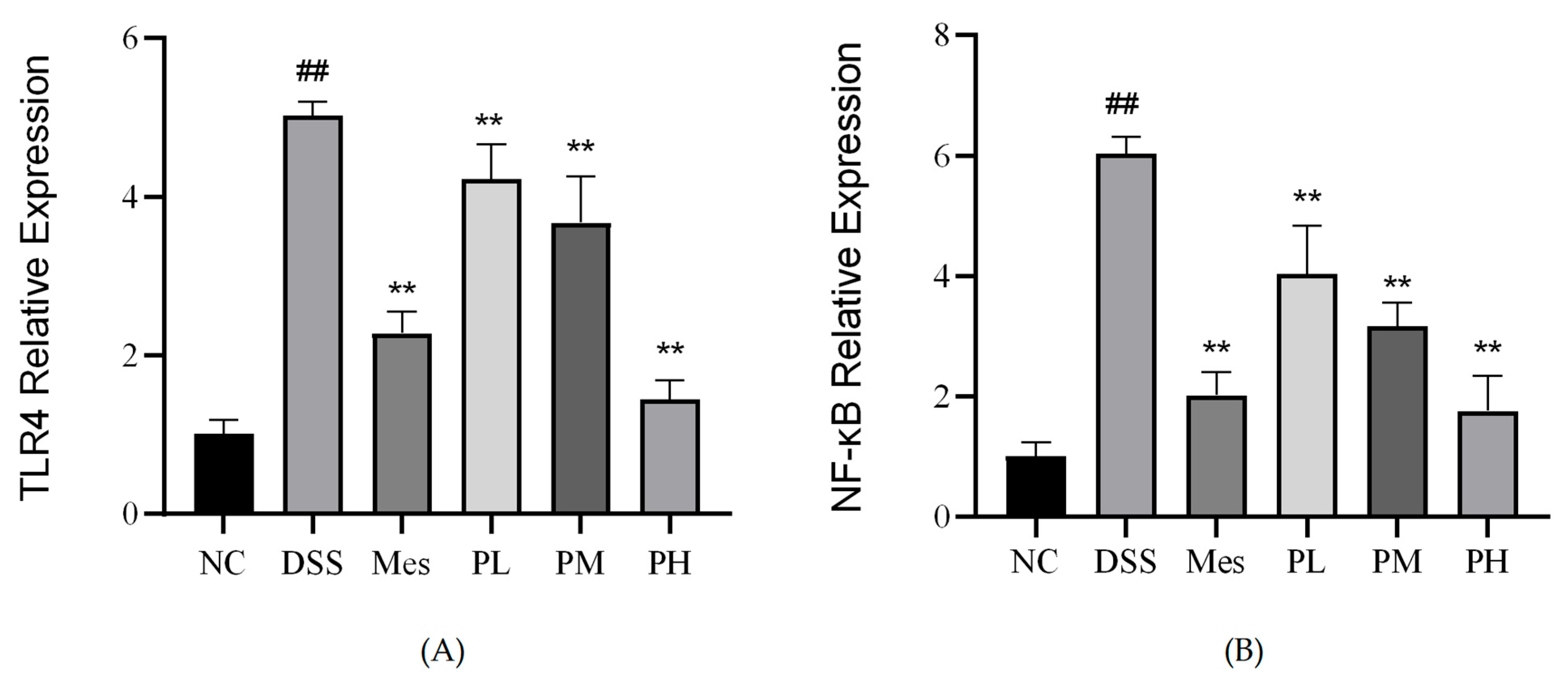
2.14. Effect of POAs on TLR4 and NF-κB mRNA Expression in Colon Tissue of UC Mice
3. Discussion
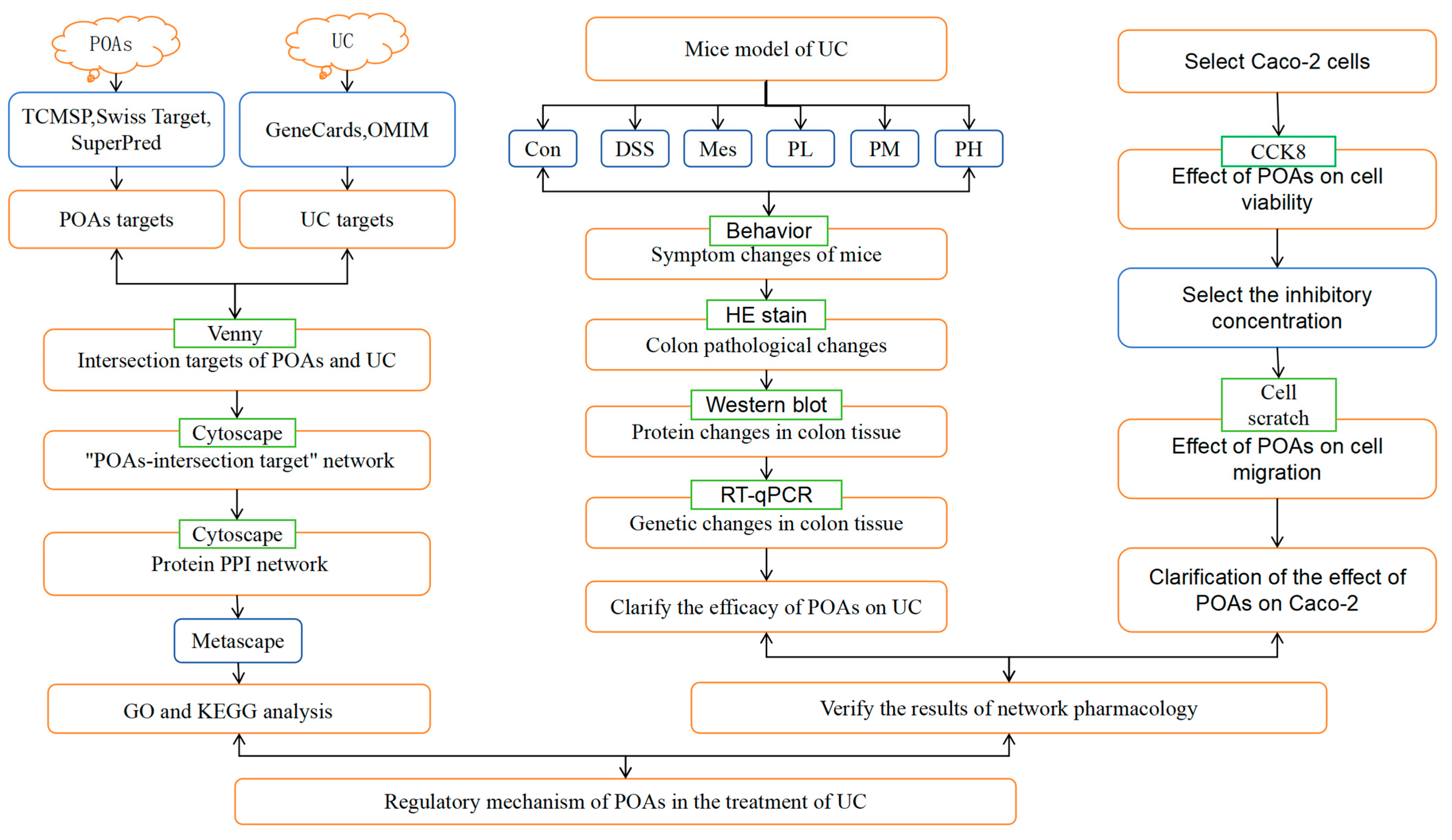
4. Materials and Methods
4.1. Network Pharmacology Analysis
4.1.1. Identification of POAs Active Compounds and Potential Targets
4.1.2. Collection of UC-Related Targets
4.1.3. Identification of Overlapping Targets Between POAs and UC
4.1.4. Construction of the POAs—Common Target Network
4.1.5. Protein–Protein Interaction (PPI) Network Construction and Key Target Identification
4.1.6. GO and KEGG Enrichment Analysis
4.2. Cell Experiments
4.2.1. Materials and Reagents
4.2.2. Tumor Cell Proliferation Assay (CCK-8)
4.2.3. Tumor Migration Wound Healing Assay
4.3. Animal Experiments
4.3.1. Animals
4.3.2. Reagents and Instruments
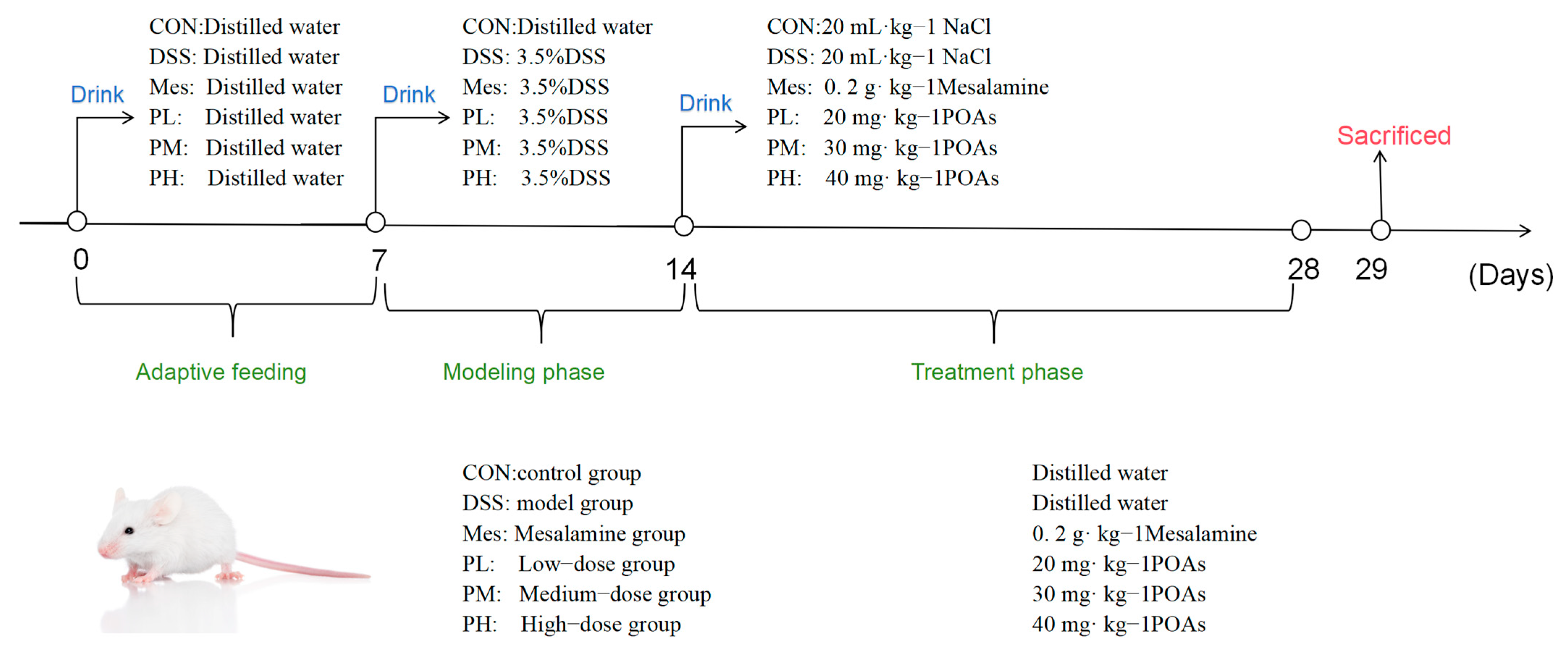
4.3.3. Animal Grouping, Modeling, and Drug Administration
4.3.4. Body Weight and DAI Scoring
4.3.5. Colon Length Measurement
4.3.6. Hematoxylin–Eosin (H&E) Staining of Colon Tissues
4.3.7. ELISA for Serum Inflammatory Cytokines
4.3.8. Western Blot Analysis of Protein Expression in Colon Tissues
4.3.9. Quantitative Real-Time PCR (qRT-PCR) Analysis of Colonic Gene Expression
4.3.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bisgaard, T.H.; Allin, K.H.; Keefer, L.; Ananthakrishnan, A.N.; Bernstein, C.N.; Gearry, R.B.; Gracie, D.J.; Khan, N.; Lomer, M.C.E.; van der Woude, C.J.; et al. Depression and anxiety in inflammatory bowel disease: Epidemiology, mechanisms and treatment. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Gros, B.; Kaplan, G.G. Ulcerative Colitis in Adults: A Review. JAMA 2023, 330, 951–965. [Google Scholar] [CrossRef] [PubMed]
- Saez, A.; Herrero-Fernandez, B.; Gomez-Bris, R.; Prieto-Fernandez, J.G.; Lopez-Cruz, L.A.; Martin-Sanchez, D.; Alonso-Castro, L.; Torres-Fuentes, C.; Martinez-Perez, A.; Fernandez-Ruiz, I.; et al. Pathophysiology of Inflammatory Bowel Disease: Innate Immune System. Int. J. Mol. Sci. 2023, 24, 1526. [Google Scholar] [CrossRef] [PubMed]
- Qiu, P.; Ishimoto, T.; Fu, L.; Zhang, C.; Kimura, Y.; Kobayashi, E.; Tanaka, S.; Wang, H.; Okada, M.; Shimizu, K.; et al. The Gut Microbiota in Inflammatory Bowel Disease. Front. Cell. Infect. Microbiol. 2022, 12, 733992. [Google Scholar] [CrossRef] [PubMed]
- Ungaro, R.; Mehandru, S.; Allen, P.B.; Peyrin-Biroulet, L.; Colombel, J.F. Ulcerative colitis. Lancet 2017, 389, 1756–1770. [Google Scholar] [CrossRef] [PubMed]
- Wan, P.; Chen, H.; Guo, Y.; Bai, A.P. Advances in treatment of ulcerative colitis with herbs: From bench to bedside. World J. Gastroenterol. 2014, 20, 14099–14104. [Google Scholar] [CrossRef] [PubMed]
- Shao, G.; Liu, Y.; Lu, L.; Wu, R.; Zhang, Z.; Chen, Y.; Lin, J.; Zhao, H.; Yang, Y.; Zhou, Q.; et al. Therapeutic potential of traditional Chinese medicine in the prevention and treatment of digestive inflammatory cancer transformation: Portulaca oleracea L. as a promising drug. J. Ethnopharmacol. 2024, 327, 117999. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Huang, Y.; Santos, H.O.; Wang, Z.; Zhang, W.; Chen, M.; Li, T.; Liu, Y.; Fang, Q.; Xu, X.; et al. Effects of purslane supplementation on C-reactive protein levels and biomarkers of oxidative stress as marks for metabolic syndrome: A systematic review and meta-analysis of randomized controlled trials. Phytother. Res. 2021, 35, 5477–5486. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Lu, K.; Lin, P.; Wang, Z.; Zhang, W.; Chen, M.; Li, T.; Liu, Y.; Fang, Q.; Xu, X.; et al. Saccharina japonica Ethanol Extract Ameliorates Depression/Anxiety-Like Behavior by Inhibiting Inflammation, Oxidative Stress, and Apoptosis in Dextran Sodium Sulfate Induced Ulcerative Colitis Mice. Front. Nutr. 2021, 8, 784532. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.; Raish, M.; Ahmad, A.; Ansari, M.A.; Alkharfy, K.M.; Alqahtani, F.Y.; Almajwal, A.M.; Haque, E.; Al-Abbasi, F.A.; Ahad, A.; et al. Sinapic Acid Ameliorates Acetic Acid-Induced Ulcerative Colitis in Rats by Suppressing Inflammation, Oxidative Stress, and Apoptosis. Molecules 2022, 27, 4139. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Wu, R.; Liu, T.; Liu, Y.; Zhao, H.; Lin, J.; Zhang, Y.; Chen, Y.; Sun, Y.; Wang, C.; et al. Peroxiredoxin 6 alleviates high glucose-induced inflammation and apoptosis in HK-2 cells by inhibiting TLR4/NF-κB signaling. Ann. Transl. Med. 2023, 11, 41. [Google Scholar] [CrossRef] [PubMed]
- Yan, P.; Qin, O. Therapeutic effect and mechanisms study of Bawei Xilei Powder on ulcerative colitis patients. Chin. J. Integr. Tradit. West. Med. 2014, 34, 27–30. [Google Scholar]
- Zong, Y.; Meng, J.; Mao, T.; Zhao, W.; Liu, J.; Xu, Y.; Chen, K.; Gao, L.; Xie, Q.; Hu, Z.; et al. Repairing the intestinal mucosal barrier of traditional Chinese medicine for ulcerative colitis: A review. Front. Pharmacol. 2023, 14, 1273407. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, B.G.; Su, Y.H.; Wang, L.; Zhang, W.; He, J.; Yu, X.; Zhao, M.; Ren, J.; Chen, L.; et al. Potential activity of Traditional Chinese Medicine against Ulcerative colitis: A review. J. Ethnopharmacol. 2022, 289, 115084. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Ha, C. Epidemiology and Pathogenesis of Ulcerative Colitis. Gastroenterol. Clin. N. Am. 2020, 49, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Di, B.; Xu, L.L. Recent advances in the treatment of IBD: Targets, mechanisms and related therapies. Cytokine Growth Factor Rev. 2023, 71–72, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Li, C.; Li, J.; Zhang, Y.; Xue, J.; Yang, X.; Liu, Y.; Zhao, F.; Hu, Q.; Chen, X.; et al. Extraction, Purification, Structural Characteristics, Biological Activity and Application of Polysaccharides from Portulaca oleracea L. (Purslane): A Review. Molecules 2023, 28, 4813. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Y.; Liu, Z.X.; Tan, W.W.; Zhang, X.; Jiang, Y.; Huang, M.; Wei, L.; Zhao, W.; Yu, Q.; Gao, S.; et al. Portulaca oleracea L. as a Potential Therapeutic Drug Intervention in Ulcerative Colitis: Mechanisms of Action and Clinical Studies. Drug Des. Devel. Ther. 2024, 18, 5931–5946. [Google Scholar] [CrossRef] [PubMed]
- Iranshahy, M.; Javadi, B.; Iranshahi, M.; Karimi, G.; Zolfaghari, B.; Riahi-Zanjani, B.; Shakeri, A.; Askari, V.R.; Baeeri, M.; Rezaee, R.; et al. A review of traditional uses, phytochemistry and pharmacology of Portulaca oleracea L. J. Ethnopharmacol. 2017, 205, 158–172. [Google Scholar] [CrossRef] [PubMed]
- Ning, K.; Shi, C.; Chi, Y.Y.; Liu, H.; Song, D.; Zhu, M.; He, L.; Zhao, J.; Li, B.; Wang, Y.; et al. Portulaca oleracea L. polysaccharide alleviates dextran sulfate sodium-induced ulcerative colitis by regulating intestinal homeostasis. Int. J. Biol. Macromol. 2024, 256 Pt 2, 128375. [Google Scholar] [CrossRef] [PubMed]
- Su, Q.; Li, L.; Sun, Y.; Liu, J.; Wang, C.; Zhang, W.; Zhao, Y.; Chen, Z.; Luo, Y.; Zheng, W.; et al. Effects of the TLR4/Myd88/NF-κB Signaling Pathway on NLRP3 Inflammasome in Coronary Microembolization-Induced Myocardial Injury. Cell Physiol. Biochem. 2018, 47, 1497–1508. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Yao, D.; Dong, M.; Dai, C.; Wu, S. Inflammation and Inflammatory Cytokine Contribute to the Initiation and Development of Ulcerative Colitis and Its Associated Cancer. Inflamm. Bowel Dis. 2019, 25, 1595–1602. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, C.; Cao, J.; Wang, D.; Liu, Y.; Zhao, Q.; Sun, X.; Yang, J.; Li, H.; Huang, Y.; et al. Yiyi Fuzi Baijiang Decoction Alleviates Ulcerative Colitis Partly by Regulating TLR4-Mediated PI3K/Akt and NF-κB Pathways. Evid. Based Complement. Alternat. Med. 2022, 2022, 8780514. [Google Scholar] [CrossRef] [PubMed]
- Shao, S.; Jia, R.; Zhao, L.; Wu, X.; Zhang, Y.; Zhou, Q.; Li, J.; Chen, Y.; Zeng, H.; Zhang, R.; et al. Xiao-Chai-Hu-Tang ameliorates tumor growth in cancer comorbid depressive symptoms via modulating gut microbiota-mediated TLR4/MyD88/NF-κB signaling pathway. Phytomedicine 2021, 88, 153606. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Li, Y.; Wu, Z.; Zhang, X.; Liu, H.; Wang, S.; Yang, J.; Zhu, H.; Lin, Y.; Guo, W.; et al. Heparin alleviates LPS-induced endothelial injury by regulating the TLR4/MyD88 signaling pathway. Exp. Ther. Med. 2021, 22, 1397. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Li, X.; Qu, L.; Tang, C.; Liu, Y.; Wang, Y.; Zhao, Z.; Wang, F.; Chen, Y.; Zhang, W.; et al. DXXK exerts anti-inflammatory effects by inhibiting the lipopolysaccharide-induced NF-κB/COX-2 signalling pathway and the expression of inflammatory mediators. J. Ethnopharmacol. 2016, 178, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhu, S.; Gu, Y.; Wang, X.; Fang, C.; Liu, R.; Han, Y.; Liu, Y.; Chen, H.; Wang, L.; et al. Network Pharmacology Combined with Experimental Validation to Investigate the Mechanism of the Anti-Hyperuricemia Action of Portulaca oleracea Extract. Nutrients 2024, 16, 3549. [Google Scholar] [CrossRef] [PubMed]
- Gazim, Z.C.; Valle, J.S.; Carvalho Dos Santos, I.; Souza, F.; Cardoso, G.; Oliveira, B.; Pereira, R.; Lopes, F.; Almeida, L.; Gonçalves, L.; et al. Ethnomedicinal, phytochemical and pharmacological investigations of Baccharis dracunculifolia DC. (ASTERACEAE). Front. Pharmacol. 2022, 13, 1048688. [Google Scholar] [CrossRef] [PubMed]
- Yikui, L. Methodology of Pharmacological Experiments in Chinese Medicine; Shanghai Scientific and Technical Publishers: Shanghai, China, 2006. [Google Scholar]
- Cordes, F.; Foell, D.; Ding, J.N.; Kesselring, R.; Trautwein, C.; Ludwig, D.; Weigmann, B.; Zeitz, M.; Galle, P.R.; Schreiber, S.; et al. Differential regulation of JAK/STAT-signaling in patients with ulcerative colitis and Crohn’s disease. World J. Gastroenterol. 2020, 26, 4055–4075. [Google Scholar] [CrossRef] [PubMed]
- Van Gennep, S.; Fung, I.C.N.; Jong, D.C.; van der Meulen-de Jong, A.E.; Meijer, S.L.; van der Woude, C.J.; Oldenburg, B.; D’Haens, G.R.; Wildenberg, M.E.; de Boer, N.K.; et al. Histological Outcomes and JAK-STAT Signalling in Ulcerative Colitis Patients Treated with Tofacitinib. J. Crohns Colitis 2024, 18, 1283–1291. [Google Scholar] [CrossRef] [PubMed]
- Scalavino, V.; Piccinno, E.; Giannelli, G.; Serino, G. miR-369-3p Ameliorates Inflammation and Apoptosis in Intestinal Epithelial Cells via the MEK/ERK Signaling Pathway. Int. J. Mol. Sci. 2025, 26, 4288. [Google Scholar] [CrossRef] [PubMed]
- Ru, J.; Li, P.; Wang, J.; Zhou, W.; Li, B.; Huang, C.; Li, P.; Guo, Z.; Tao, W.; Yang, Y.; et al. TCMSP: A database of systems pharmacology for drug discovery from herbal medicines. J. Cheminform. 2014, 6, 13. [Google Scholar] [CrossRef] [PubMed]
- Kim, S. Getting the most out of PubChem for virtual screening. Expert. Opin. Drug Discov. 2016, 11, 843–855. [Google Scholar] [CrossRef] [PubMed]
- Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: Updated data and new features for efficient prediction of protein targets of small molecules. Nucleic Acids Res. 2019, 47, W357–W364. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.X.; Qin, Y.S.; Shi, B.H.; Zhang, L.J.; Xu, Y.; Zhou, Y.L.; Sun, L.; Fang, Y.; Wang, Y.; Guo, L.; et al. Effects of Curcumin on Radiation/Chemotherapy-Induced Oral Mucositis: Combined Meta-Analysis, Network Pharmacology, Molecular Docking, and Molecular Dynamics Simulation. Curr. Issues Mol. Biol. 2024, 46, 10545–10569. [Google Scholar] [CrossRef] [PubMed]
- Safran, M.; Dalah, I.; Alexander, J.; Rosen, N.; Iny Stein, T.; Shmoish, M.; Nativ, N.; Bahir, I.; Doniger, T.; Krug, H.; et al. GeneCards Version 3: The human gene integrator. Database 2010, 2010, baq020. [Google Scholar] [CrossRef] [PubMed]
- Amberger, J.S.; Bocchini, C.A.; Schiettecatte, F.; Scott, A.F.; Hamosh, A. OMIM.org: Online Mendelian Inheritance in Man (OMIM®), an online catalog of human genes and genetic disorders. Nucleic Acids Res. 2015, 43, D789–D798. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Morris, J.H.; Cook, H.; Kuhn, M.; Wyder, S.; Simonovic, M.; Santos, A.; Doncheva, N.T.; Roth, A.; Bork, P.; et al. The STRING database in 2017: Quality-controlled protein-protein association networks, made broadly accessible. Nucleic Acids Res. 2017, 45, D362–D368. [Google Scholar] [CrossRef] [PubMed]
- Otasek, D.; Morris, J.H.; Bouças, J.; Pico, A.R.; Demchak, B.; Bader, G.D. Cytoscape Automation: Empowering workflow-based network analysis. Genome Biol. 2019, 20, 185. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tao, X.; Gao, Y.; Wang, Y.; Huang, W.; Zhu, Y.; Sun, G.; Wu, J.; Li, D.; Chen, X.; et al. Study on the mechanism of Shujin Tongluo granules in treating cervical spondylosis based on network pharmacology and molecular docking. Medicine 2023, 102, e34030. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.X.Y.; Gao, S.; Zhang, Y.; Zhang, X.; Zhu, J.; Jiang, D. Determination of Total Alkaloids Content in Portulaca oleracea L. from Different Habitats. Appl. Chem. Ind. 2014, 12, 2310–2312. [Google Scholar]
- Yang, C.; Merlin, D. Unveiling Colitis: A Journey through the Dextran Sodium Sulfate-induced Model. Inflamm. Bowel Dis. 2024, 30, 844–853. [Google Scholar] [CrossRef] [PubMed]
- Sehgal, P.; Colombel, J.F.; Aboubakr, A.; Narula, N. Systematic review: Safety of mesalazine in ulcerative colitis. Aliment. Pharmacol. Ther. 2018, 47, 1597–1609. [Google Scholar] [CrossRef] [PubMed]
- Commission, C.P. Pharmacopoeia of the People’s Republic of China, 10th ed.; China Medical Science and Technology Press: Beijing, China, 2020. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component Name | Mol ID | Molecular Formula | Category |
|---|---|---|---|
| arachidonic acid | MOL001439 | C20H32O2 | Polyunsaturated fatty acid |
| cycloartenol | MOL003578 | C30H50O | Triterpenoid |
| beta-carotene | MOL002773 | C40H56 | Carotenoid |
| beta-sitosterol | MOL000358 | C29H50O | Phytosterol |
| kaempferol | MOL000422 | C15H10O6 | Flavonol |
| 5,7-dihydroxy-2-(3-hydroxy-4-methoxyphenyl)chroman-4-one | MOL005100 | C16H14O6 | Flavonoids |
| luteolin | MOL000006 | C15H10O6 | Flavonoids |
| isobetanidin | MOL006657 | C18H16N2O8 | Betalains |
| isobetanin_qt | MOL006662 | C24H26N2O13 | Betalains |
| quercetin | MOL000098 | C15H10O7 | Flavonoids |
| Score | Weight Loss/% | Diarrhea | Hematochezia |
|---|---|---|---|
| 0 | <1 | Normal | No bleeding |
| 1 | 1~5 | Loose stools | Occult bleeding |
| 2 | 6~10 | Pasty stools | Visual bleeding |
| 3 | >10 | Diarrhea | Gross bleeding |
| Score | Epithelial Damage | Inflammatory Infiltration | Crypt Architecture | Ulceration |
|---|---|---|---|---|
| 0 | Intact epithelium | No infiltration | Normal crypts | None |
| 1 | Focal erosion | Minimal damage | <10% crypt distortion | No ulcers |
| 2 | Multifocal erosion | Moderate | 10–30% crypt loss | <0.5 mm |
| 3 | Extensive erosion | Severe inflammation with microabscesses | 30–70% crypt destruction | 0.5–2 mm |
| 4 | Complete loss | Transmural inflammation with necrosis | >70% crypt loss | >2 mm |
| Gene Name | Primer Sequence (5′→3′) |
|---|---|
| TLR4 | F:TCCCTGCATAGAGGTAGTTCC |
| R:TCAAGGGGTTGAAGCTCAGA | |
| Nf-κb | F:CTCTGGCACAGAAGTTGGGT |
| R:TCCCGGAGTTCATCTCATAGT | |
| GAPDH | F:GGAGAGTGTTTCCTCGTCCC |
| R:ATGAAGGGGTCGTTGATGGC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, T.; Gao, L.; Wang, Q.; Zheng, J.; Wang, X.; Jiang, M.; Wu, K.; Ai, J. Exploring the Regulatory Mechanism of Total Alkaloids from Portulaca oleracea L. in UC Treatment Based on Network Pharmacology. Int. J. Mol. Sci. 2025, 26, 6978. https://doi.org/10.3390/ijms26146978
Zhang T, Gao L, Wang Q, Zheng J, Wang X, Jiang M, Wu K, Ai J. Exploring the Regulatory Mechanism of Total Alkaloids from Portulaca oleracea L. in UC Treatment Based on Network Pharmacology. International Journal of Molecular Sciences. 2025; 26(14):6978. https://doi.org/10.3390/ijms26146978
Chicago/Turabian StyleZhang, Tianci, Linran Gao, Qianying Wang, Jiahui Zheng, Xinyu Wang, Meng Jiang, Kaixin Wu, and Jinxia Ai. 2025. "Exploring the Regulatory Mechanism of Total Alkaloids from Portulaca oleracea L. in UC Treatment Based on Network Pharmacology" International Journal of Molecular Sciences 26, no. 14: 6978. https://doi.org/10.3390/ijms26146978
APA StyleZhang, T., Gao, L., Wang, Q., Zheng, J., Wang, X., Jiang, M., Wu, K., & Ai, J. (2025). Exploring the Regulatory Mechanism of Total Alkaloids from Portulaca oleracea L. in UC Treatment Based on Network Pharmacology. International Journal of Molecular Sciences, 26(14), 6978. https://doi.org/10.3390/ijms26146978