
The Dynamic Regulation of Daxx-Mediated Transcriptional Inhibition by SUMO and PML NBs

Abstract
1. Introduction
2. SUMO and SUMOylation
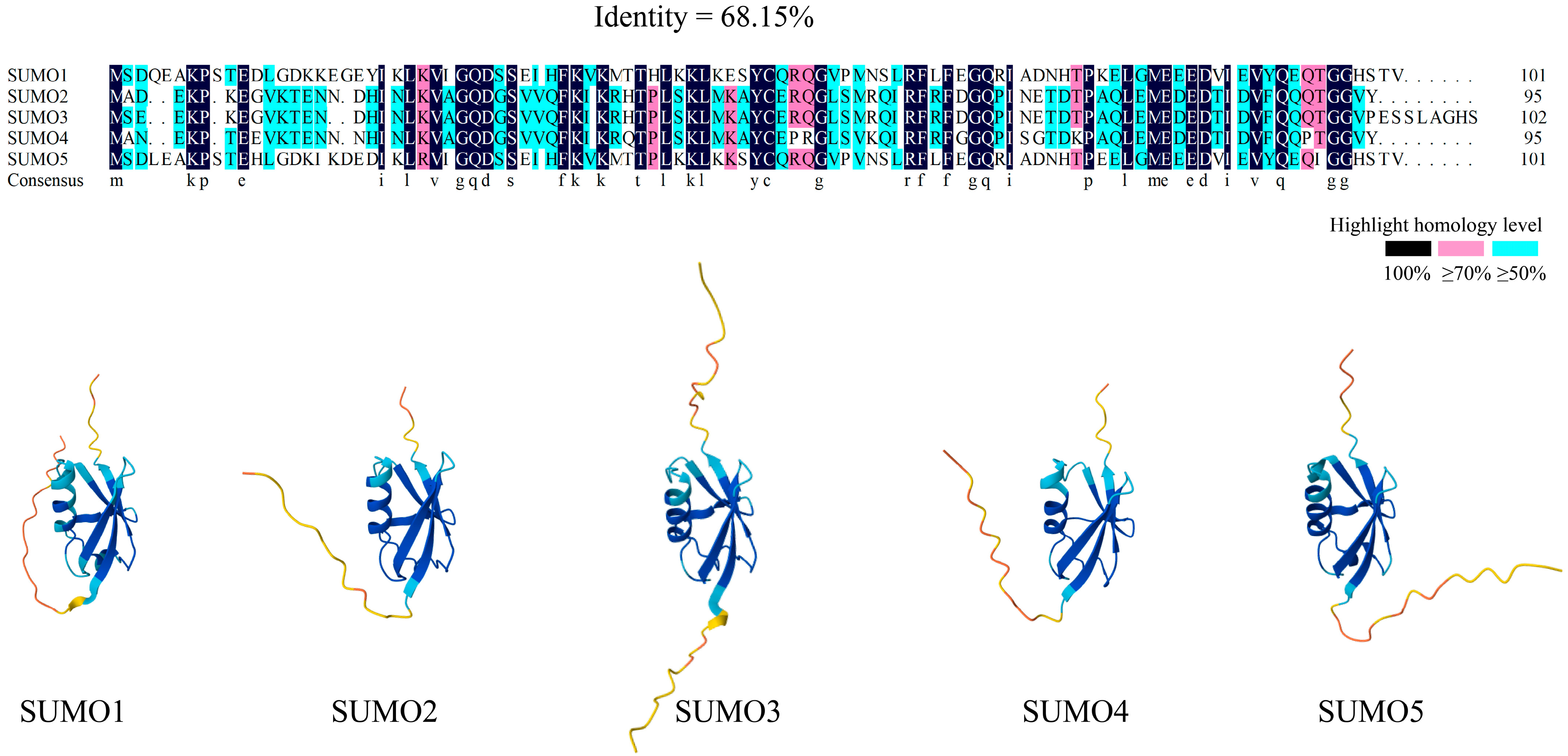
2.1. The SUMO Family
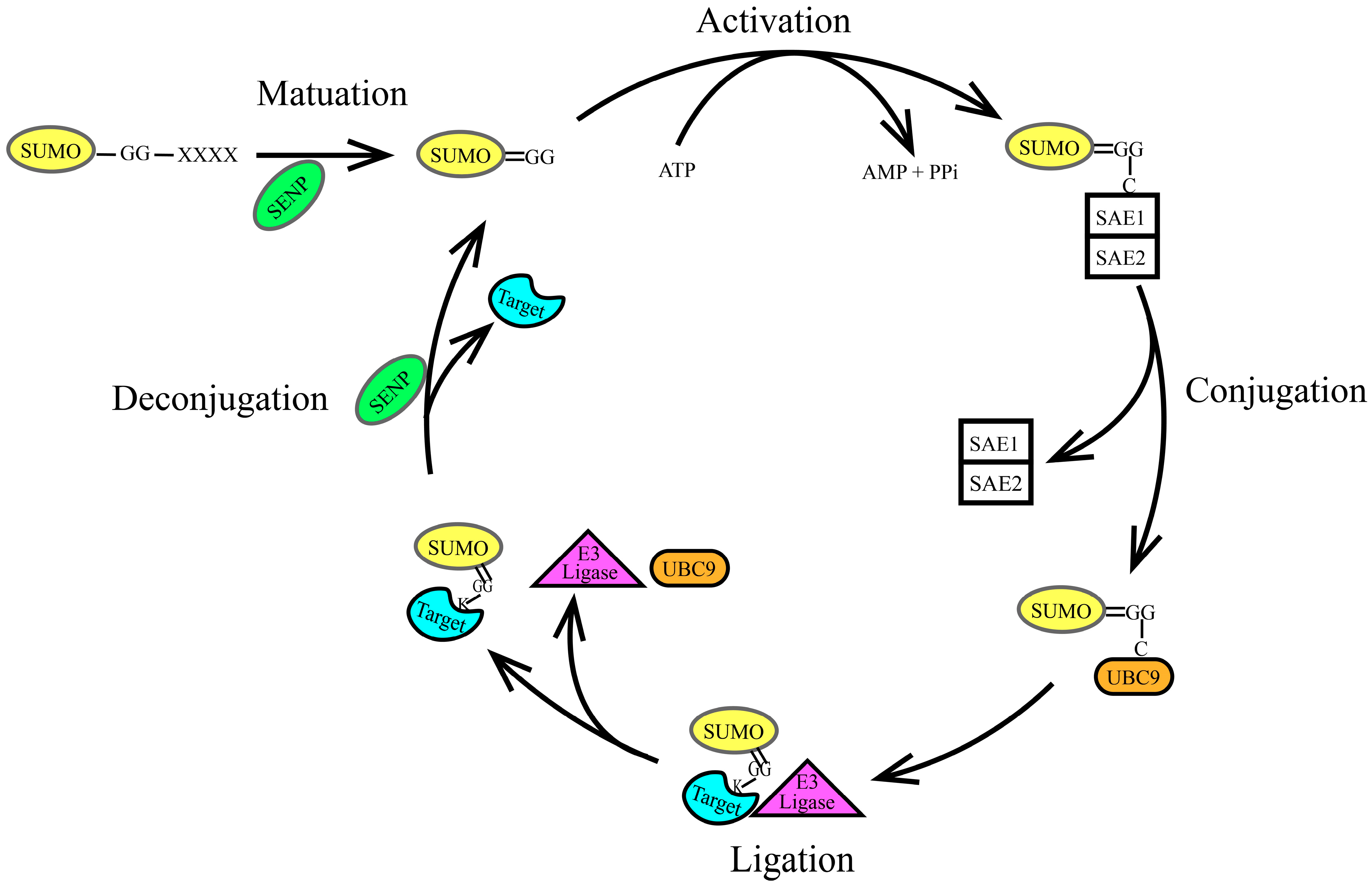
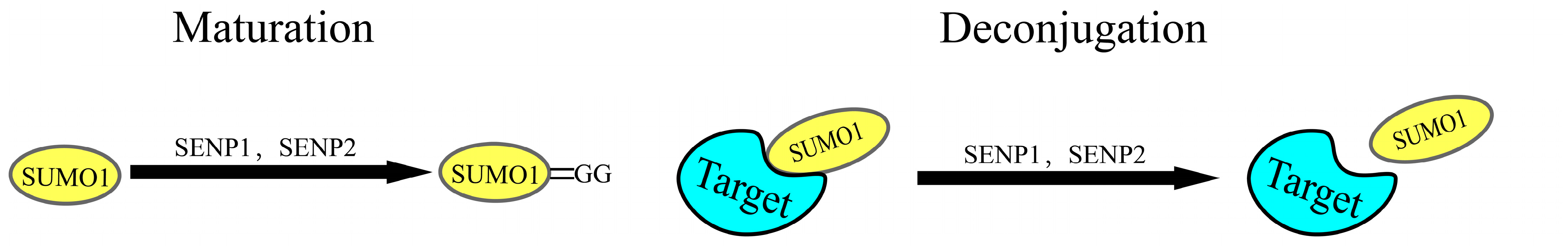
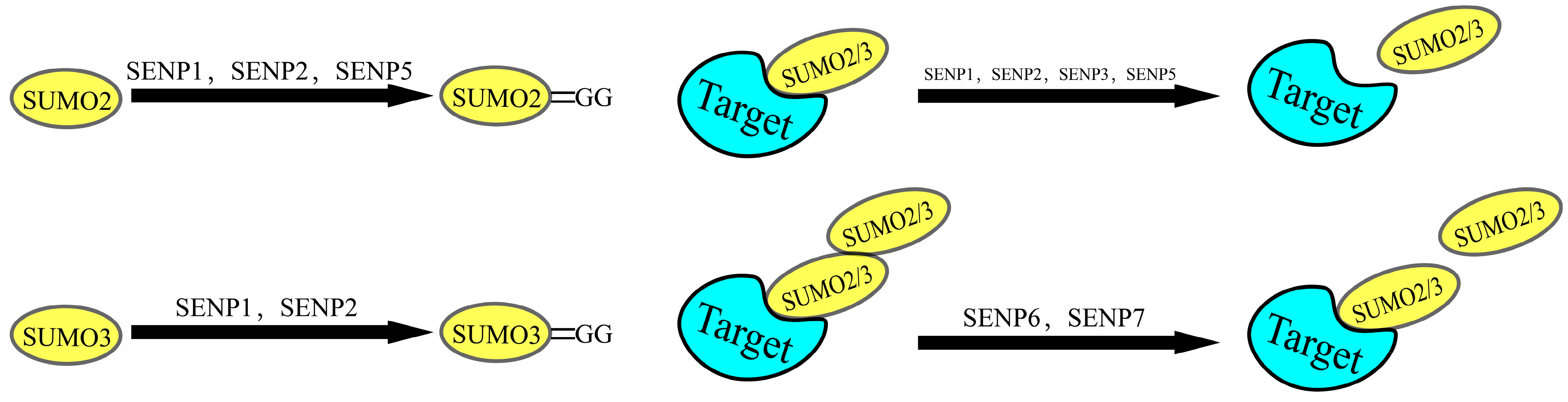
2.2. The SUMOylation Cascade
3. Interaction Between Daxx and SUMO
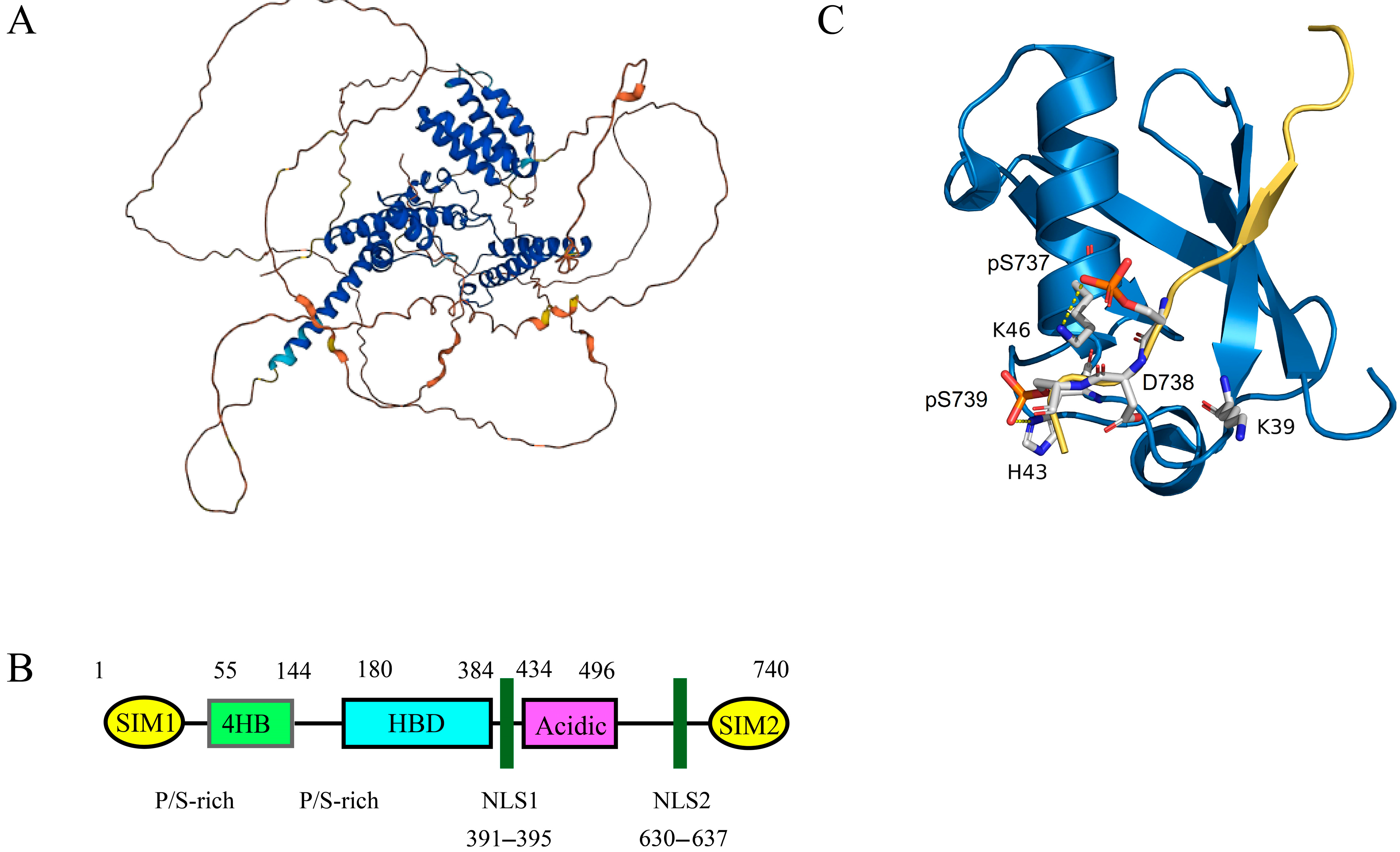
3.1. Structure and Biological Characteristics of Daxx
3.2. Structural Basis of Daxx Binding to SUMO
3.3. SUMOylation of Daxx
4. Regulation of Daxx-Mediated Transcriptional Repression by SUMO Within PML Nuclear Bodies
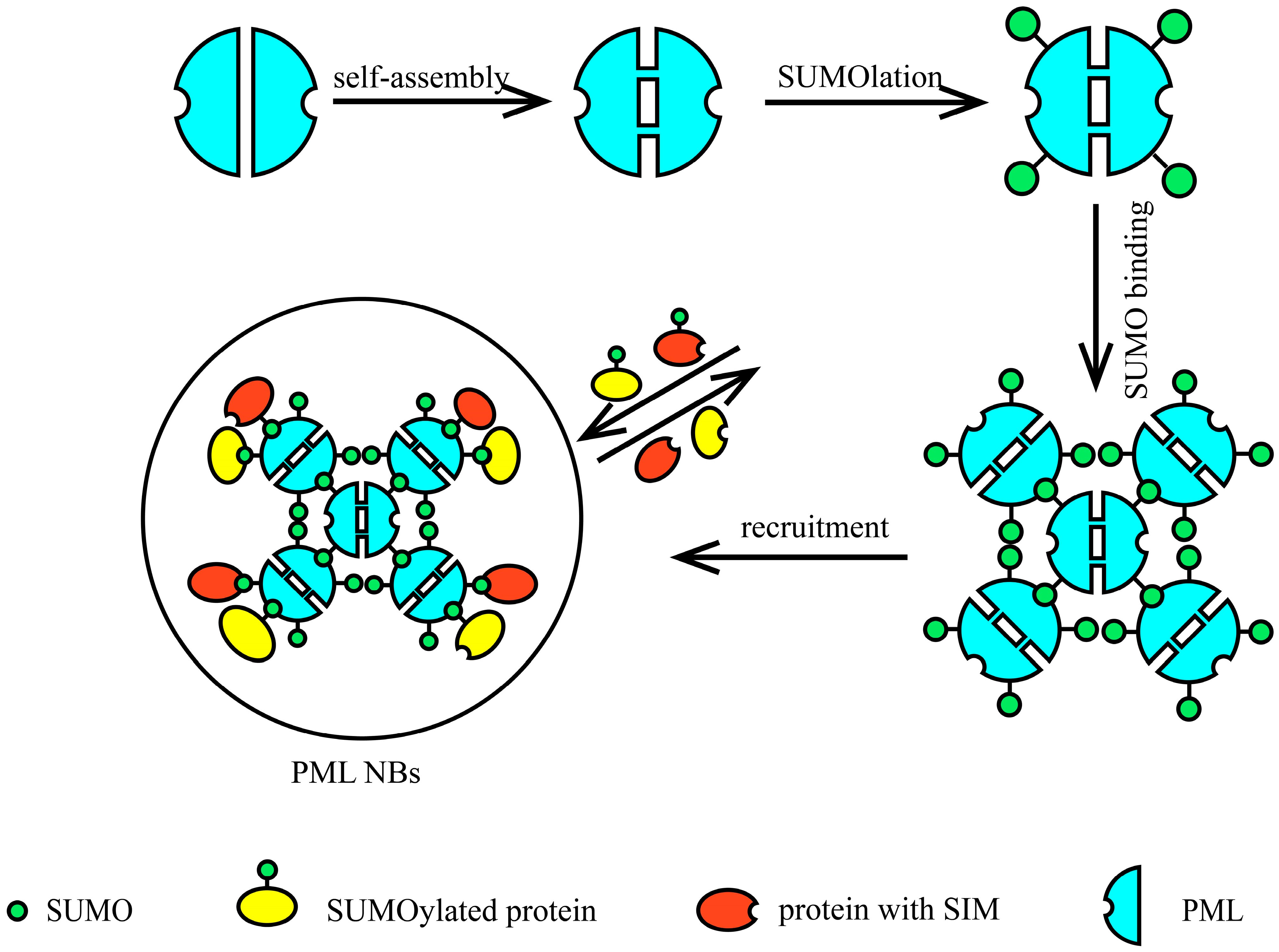
4.1. Structure of PML NBs
4.2. Role of SUMO in the Assembly of PML NBs
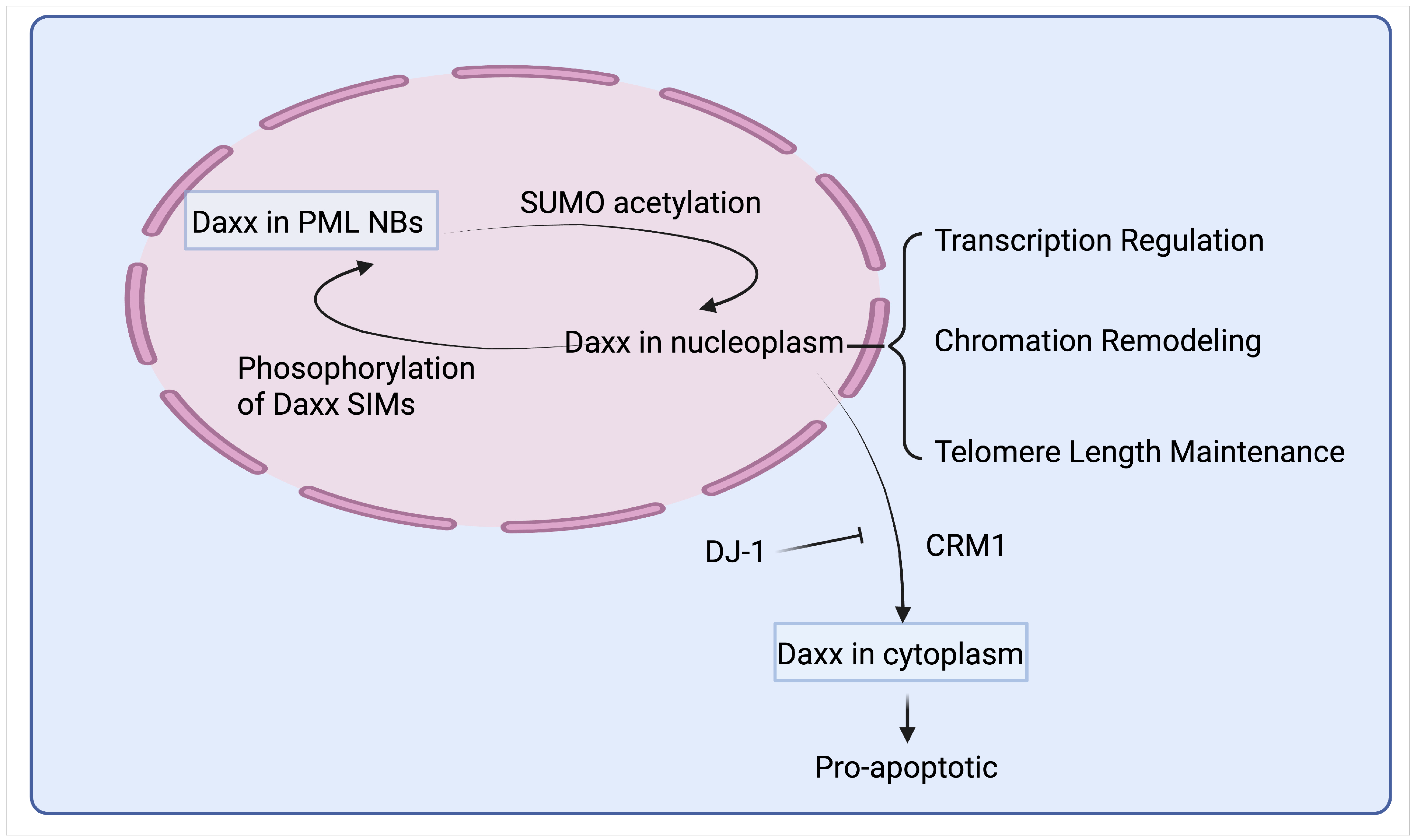
4.3. Dynamic Regulation of Daxx Localization Within PML NBs Through Post-Translational Modifications
5. The Interaction of Daxx with Specific Transcription Factors in a SUMO-Depend Manner
5.1. Daxx Inhibits the Transcriptional Activity of ETS1 in a SUMO-Dependent Manner
5.2. Daxx Inhibits the Transcriptional Activity of GR in a SUMO-Dependent Manner
5.3. Regulation of Pax3 Transcriptional Activity Through Daxx Release from PML NBs
5.4. Daxx Inhibits the Transcriptional Activity of SMAD4 in a SUMO-Dependent Manner
5.5. Involvement of SUMOylation in Daxx-Mediated Inhibition of AR Transcriptional Activity
5.6. Daxx’s Inhibition of AIRE Transcriptional Activity Via Its C-Terminal Domain
5.7. The Inhibition of SNAI2 Transcriptional Activity by Daxx May Require the Participation of SUMO
6. Potential Pathway of Daxx-Mediated Transcriptional Repression
7. Subcellular Localization of Daxx and Disease Implications
7.1. Subcellular Localization and Functions of Daxx
7.2. Pathological Consequences of Aberrant Daxx Localization
7.2.1. Enhanced Nuclear Export of Daxx May Exacerbate Parkinson’s Disease (PD)
7.2.2. The Abnormal Release of Daxx in PML NBs Exacerbates Acute Promyelocytic Leukemia (APL)
7.2.3. SUMO2/3-Mediated Increase in Nuclear DAXX Aggravates Gastric Cancer (GC)
8. Summery and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pinto, L.M.; Pailas, A.; Bondarchenko, M.; Sharma, A.B.; Neumann, K.; Rizzo, A.J.; Jeanty, C.; Nicot, N.; Racca, C.; Graham, M.K.; et al. DAXX promotes centromeric stability independently of ATRX by preventing the accumulation of R-loop-induced DNA double-stranded breaks. Nucleic Acids Res. 2024, 52, 1136–1155. [Google Scholar] [CrossRef] [PubMed]
- Groh, S.; Milton, A.V.; Marinelli, L.K.; Sickinger, C.V.; Russo, A.; Bollig, H.; de Almeida, G.P.; Schmidt, A.; Forné, I.; Imhof, A.; et al. Morc3 silences endogenous retroviruses by enabling Daxx-mediated histone H3.3 incorporation. Nat. Commun. 2021, 12, 5996. [Google Scholar] [CrossRef] [PubMed]
- Puto, L.A.; Brognard, J.; Hunter, T. Transcriptional Repressor DAXX Promotes Prostate Cancer Tumorigenicity via Suppression of Autophagy. J. Biol. Chem. 2015, 290, 15406–15420. [Google Scholar] [CrossRef] [PubMed]
- Shih, H.M.; Chang, C.C.; Kuo, H.Y.; Lin, D.Y. Daxx mediates SUMO-dependent transcriptional control and subnuclear compartmentalization. Biochem. Soc. Trans. 2007, 35, 1397–1400. [Google Scholar] [CrossRef]
- Flotho, A.; Melchior, F. Sumoylation: A regulatory protein modification in health and disease. Annu. Rev. Biochem. 2013, 82, 357–385. [Google Scholar] [CrossRef]
- Chymkowitch, P.; Nguea, P.A.; Enserink, J.M. SUMO-regulated transcription: Challenging the dogma. Bioessays 2015, 37, 1095–1105. [Google Scholar] [CrossRef]
- Lambert, S.A.; Jolma, A.; Campitelli, L.F.; Das, P.K.; Yin, Y.; Albu, M.; Chen, X.; Taipale, J.; Hughes, T.R.; Weirauch, M.T. The Human Transcription Factors. Cell 2018, 172, 650–665. [Google Scholar] [CrossRef]
- Medvedeva, Y.A.; Lennartsson, A.; Ehsani, R.; Kulakovskiy, I.V.; Vorontsov, I.E.; Panahandeh, P.; Khimulya, G.; Kasukawa, T.; Drablos, F. EpiFactors: A comprehensive database of human epigenetic factors and complexes. Database 2015, 2015, bav067. [Google Scholar] [CrossRef]
- DelRosso, N.; Tycko, J.; Suzuki, P.; Andrews, C.; Aradhana; Mukund, A.; Liongson, I.; Ludwig, C.; Spees, K.; Fordyce, P.; et al. Large-scale mapping and mutagenesis of human transcriptional effector domains. Nature 2023, 616, 365–372. [Google Scholar] [CrossRef]
- Gill, G. Something about SUMO inhibits transcription. Curr. Opin. Genet. Dev. 2005, 15, 536–541. [Google Scholar] [CrossRef]
- Ouyang, J.; Gill, G. SUMO engages multiple corepressors to regulate chromatin structure and transcription. Epigenetics 2009, 4, 440–444. [Google Scholar] [CrossRef] [PubMed]
- Rosonina, E.; Akhter, A.; Dou, Y.; Babu, J.; Sri, T.V.S. Regulation of transcription factors by sumoylation. Transcription 2017, 8, 220–231. [Google Scholar] [CrossRef] [PubMed]
- Lindsay, C.R.; Morozov, V.M.; Ishov, A.M. PML NBs (ND10) and Daxx: From nuclear structure to protein function. Front. Biosci. 2008, 13, 7132–7142. [Google Scholar] [CrossRef] [PubMed]
- Sahin, U.; de The, H.; Lallemand-Breitenbach, V. Sumoylation in Physiology, Pathology and Therapy. Cells 2022, 11, 814. [Google Scholar] [CrossRef]
- Sheng, Z.; Wang, X.; Ma, Y.; Zhang, D.; Yang, Y.; Zhang, P.; Zhu, H.; Xu, N.; Liang, S. MS-based strategies for identification of protein SUMOylation modification. Electrophoresis 2019, 40, 2877–2887. [Google Scholar] [CrossRef]
- Talamillo, A.; Barroso-Gomila, O.; Giordano, I.; Ajuria, L.; Grillo, M.; Mayor, U.; Barrio, R. The role of SUMOylation during development. Biochem. Soc. Trans. 2020, 48, 463–478. [Google Scholar] [CrossRef]
- Yang, Y.; He, Y.; Wang, X.; Liang, Z.; He, G.; Zhang, P.; Zhu, H.; Xu, N.; Liang, S. Protein SUMOylation modification and its associations with disease. Open Biol. 2017, 7, 170167. [Google Scholar] [CrossRef]
- Han, Z.J.; Feng, Y.H.; Gu, B.H.; Li, Y.M.; Chen, H. The post-translational modification, SUMOylation, and cancer (Review). Int. J. Oncol. 2018, 52, 1081–1094. [Google Scholar] [CrossRef]
- Hu, M.M.; Yang, Q.; Xie, X.Q.; Liao, C.Y.; Lin, H.; Liu, T.T.; Yin, L.; Shu, H.B. Sumoylation Promotes the Stability of the DNA Sensor cGAS and the Adaptor STING to Regulate the Kinetics of Response to DNA Virus. Immunity 2016, 45, 555–569. [Google Scholar] [CrossRef]
- Liang, Y.C.; Lee, C.C.; Yao, Y.L.; Lai, C.C.; Schmitz, M.L.; Yang, W.M. SUMO5, a Novel Poly-SUMO Isoform, Regulates PML Nuclear Bodies. Sci. Rep. 2016, 6, 26509. [Google Scholar] [CrossRef]
- Hay, R.T. SUMO: A history of modification. Mol. Cell 2005, 18, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Bernstock, J.D.; Yang, W.; Ye, D.G.; Shen, Y.; Pluchino, S.; Lee, Y.J.; Hallenbeck, J.M.; Paschen, W. SUMOylation in brain ischemia: Patterns, targets, and translational implications. J. Cereb. Blood Flow Metab. 2018, 38, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Enserink, J.M. Sumo and the cellular stress response. Cell Div. 2015, 10, 4. [Google Scholar] [CrossRef]
- Niskanen, E.A.; Palvimo, J.J. Chromatin SUMOylation in heat stress: To protect, pause and organise? SUMO stress response on chromatin. Bioessays 2017, 39, 1600263. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, Z.; Zhou, Z.; Yang, P.; Wang, C.Y. Sumoylation Modulates the Susceptibility to Type 1 Diabetes. Adv. Exp. Med. Biol. 2017, 963, 299–322. [Google Scholar]
- Chang, H.M.; Yeh, E.T.H. SUMO: From Bench to Bedside. Physiol. Rev. 2020, 100, 1599–1619. [Google Scholar] [CrossRef]
- Hickey, C.M.; Wilson, N.R.; Hochstrasser, M. Function and regulation of SUMO proteases. Nat. Rev. Mol. Cell Biol. 2012, 13, 755–766. [Google Scholar] [CrossRef]
- Bialik, P.; Wozniak, K. SUMO proteases as potential targets for cancer therapy. Postepy Hig. Med. Dosw. (Online) 2017, 71, 997–1004. [Google Scholar] [CrossRef]
- Kunz, K.; Piller, T.; Muller, S. SUMO-specific proteases and isopeptidases of the SENP family at a glance. J. Cell Sci. 2018, 131, jcs211904. [Google Scholar] [CrossRef]
- Varejao, N.; Lascorz, J.; Li, Y.; Reverter, D. Molecular mechanisms in SUMO conjugation. Biochem. Soc. Trans. 2020, 48, 123–135. [Google Scholar] [CrossRef]
- Kunadis, E.; Lakiotaki, E.; Korkolopoulou, P.; Piperi, C. Targeting post-translational histone modifying enzymes in glioblastoma. Pharmacol. Ther. 2021, 220, 107721. [Google Scholar] [CrossRef] [PubMed]
- Lara-Urena, N.; Jafari, V.; Garcia-Dominguez, M. Cancer-Associated Dysregulation of Sumo Regulators: Proteases and Ligases. Int. J. Mol. Sci. 2022, 23, 8012. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Khosravi-Far, R.; Chang, H.Y.; Baltimore, D. Daxx, a novel Fas-binding protein that activates JNK and apoptosis. Cell 1997, 89, 1067–1076. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.Y.; Nishitoh, H.; Yang, X.; Ichijo, H.; Baltimore, D. Activation of apoptosis signal-regulating kinase 1 (ASK1) by the adapter protein Daxx. Science 1998, 281, 1860–1863. [Google Scholar] [CrossRef]
- Santiago, A.; Godsey, A.C.; Hossain, J.; Zhao, L.Y.; Liao, D. Identification of two independent SUMO-interacting motifs in Daxx: Evolutionary conservation from Drosophila to humans and their biochemical functions. Cell Cycle 2009, 8, 76–87. [Google Scholar] [CrossRef]
- Michaelson, J.S.; Bader, D.; Kuo, F.; Kozak, C.; Leder, P. Loss of Daxx, a promiscuously interacting protein, results in extensive apoptosis in early mouse development. Genes Dev. 1999, 13, 1918–1923. [Google Scholar] [CrossRef]
- Wasylishen, A.R.; Estrella, J.S.; Pant, V.; Chau, G.P.; Lozano, G. Daxx Functions Are p53-Independent In Vivo. Mol. Cancer Res. 2018, 16, 1523–1529. [Google Scholar] [CrossRef]
- Escobar-Cabrera, E.; Lau, D.K.; Giovinazzi, S.; Ishov, A.M.; McIntosh, L.P. Structural characterization of the DAXX N-terminal helical bundle domain and its complex with Rassf1C. Structure 2010, 18, 1642–1653. [Google Scholar] [CrossRef]
- Mahmud, I.; Liao, D. DAXX in cancer: Phenomena, processes, mechanisms and regulation. Nucleic Acids Res. 2019, 47, 7734–7752. [Google Scholar] [CrossRef]
- Hollenbach, A.D.; Sublett, J.E.; McPherson, C.J.; Grosveld, G. The Pax3-FKHR oncoprotein is unresponsive to the Pax3-associated repressor hDaxx. EMBO J. 1999, 18, 3702–3711. [Google Scholar] [CrossRef]
- Ishov, A.M.; Sotnikov, A.G.; Negorev, D.; Vladimirova, O.V.; Neff, N.; Kamitani, T.; Yeh, E.T.; Strauss, J.F.R.; Maul, G.G. PML is critical for ND10 formation and recruits the PML-interacting protein daxx to this nuclear structure when modified by SUMO-1. J. Cell Biol. 1999, 147, 221–234. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Leo, C.; Zhu, J.; Wu, X.; O’Neil, J.; Park, E.J.; Chen, J.D. Sequestration and inhibition of Daxx-mediated transcriptional repression by PML. Mol. Cell. Biol. 2000, 20, 1784–1796. [Google Scholar] [CrossRef]
- Lehembre, F.; Müller, S.; Pandolfi, P.P.; Dejean, A. Regulation of Pax3 transcriptional activity by SUMO-1-modified PML. Oncogene 2001, 20, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hollenbach, A.D.; McPherson, C.J.; Mientjes, E.J.; Iyengar, R.; Grosveld, G. Daxx and histone deacetylase II associate with chromatin through an interaction with core histones and the chromatin-associated protein Dek. J. Cell Sci. 2002, 115, 3319–3330. [Google Scholar] [CrossRef]
- Lin, D.Y.; Huang, Y.S.; Jeng, J.C.; Kuo, H.Y.; Chang, C.C.; Chao, T.T.; Ho, C.C.; Chen, Y.C.; Lin, T.P.; Fang, H.I.; et al. Role of SUMO-interacting motif in Daxx SUMO modification, subnuclear localization, and repression of sumoylated transcription factors. Mol. Cell 2006, 24, 341–354. [Google Scholar] [CrossRef]
- Lewis, P.W.; Elsaesser, S.J.; Noh, K.M.; Stadler, S.C.; Allis, C.D. Daxx is an H3.3-specific histone chaperone and cooperates with ATRX in replication-independent chromatin assembly at telomeres. Proc. Natl. Acad. Sci. USA 2010, 107, 14075–14080. [Google Scholar] [CrossRef]
- Drane, P.; Ouararhni, K.; Depaux, A.; Shuaib, M.; Hamiche, A. The death-associated protein DAXX is a novel histone chaperone involved in the replication-independent deposition of H3.3. Genes Dev. 2010, 24, 1253–1265. [Google Scholar] [CrossRef]
- Falcinelli, M.; Dell’Omo, G.; Grassi, E.; Mariella, E.; Leto, S.M.; Scardellato, S.; Lorenzato, A.; Arena, S.; Bertotti, A.; Trusolino, L.; et al. Colorectal cancer patient-derived organoids and cell lines harboring ATRX and/or DAXX mutations lack Alternative Lengthening of Telomeres (ALT). Cell Death Dis. 2023, 14, 96. [Google Scholar] [CrossRef]
- Lovejoy, C.A.; Li, W.; Reisenweber, S.; Thongthip, S.; Bruno, J.; de Lange, T.; De, S.; Petrini, J.H.; Sung, P.A.; Jasin, M.; et al. Loss of ATRX, genome instability, and an altered DNA damage response are hallmarks of the alternative lengthening of telomeres pathway. PLoS Genet. 2012, 8, e1002772. [Google Scholar] [CrossRef]
- Huang, L.; Agrawal, T.; Zhu, G.; Yu, S.; Tao, L.; Lin, J.; Marmorstein, R.; Shorter, J.; Yang, X. DAXX represents a new type of protein-folding enabler. Nature 2021, 597, 132–137. [Google Scholar] [CrossRef]
- Chang, C.C.; Naik, M.T.; Huang, Y.S.; Jeng, J.C.; Liao, P.H.; Kuo, H.Y.; Ho, C.C.; Hsieh, Y.L.; Lin, C.H.; Huang, N.J.; et al. Structural and functional roles of Daxx SIM phosphorylation in SUMO paralog-selective binding and apoptosis modulation. Mol. Cell 2011, 42, 62–74. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wen, J.; Tuo, Q.H.; Liao, D.F. [Effects of SUMOylation on the subcellular localization and function of DAXX]. Sheng Li Xue Bao 2013, 65, 89–95. [Google Scholar] [PubMed]
- Escobar-Cabrera, E.; Okon, M.; Lau, D.K.; Dart, C.F.; Bonvin, A.M.; McIntosh, L.P. Characterizing the N- and C-terminal Small ubiquitin-like modifier (SUMO)-interacting motifs of the scaffold protein DAXX. J. Biol. Chem. 2011, 286, 19816–19829. [Google Scholar] [CrossRef] [PubMed]
- Hecker, C.M.; Rabiller, M.; Haglund, K.; Bayer, P.; Dikic, I. Specification of SUMO1- and SUMO2-interacting motifs. J. Biol. Chem. 2006, 281, 16117–16127. [Google Scholar] [CrossRef]
- Stehmeier, P.; Muller, S. Phospho-regulated SUMO interaction modules connect the SUMO system to CK2 signaling. Mol. Cell 2009, 33, 400–409. [Google Scholar] [CrossRef]
- Cappadocia, L.; Pichler, A.; Lima, C.D. Structural basis for catalytic activation by the human ZNF451 SUMO E3 ligase. Nat. Struct. Mol. Biol. 2015, 22, 968–975. [Google Scholar] [CrossRef]
- Ishov, A.M.; Vladimirova, O.V.; Maul, G.G. Daxx-mediated accumulation of human cytomegalovirus tegument protein pp71 at ND10 facilitates initiation of viral infection at these nuclear domains. J. Virol. 2002, 76, 7705–7712. [Google Scholar] [CrossRef]
- Lalioti, V.S.; Vergarajauregui, S.; Pulido, D.; Sandoval, I.V. The insulin-sensitive glucose transporter, GLUT4, interacts physically with Daxx. Two proteins with capacity to bind Ubc9 and conjugated to SUMO1. J. Biol. Chem. 2002, 277, 19783–19791. [Google Scholar] [CrossRef]
- Lalioti, V.S.; Vergarajauregui, S.; Tsuchiya, Y.; Hernandez-Tiedra, S.; Sandoval, I.V. Daxx functions as a scaffold of a protein assembly constituted by GLUT4, JNK1 and KIF5B. J. Cell. Physiol. 2009, 218, 416–426. [Google Scholar] [CrossRef]
- Rubin, B.R.; Bogan, J.S. Intracellular retention and insulin-stimulated mobilization of GLUT4 glucose transporters. Vitam. Horm. 2009, 80, 155–192. [Google Scholar]
- Hu, J.; Wang, S.; Xiong, Z.; Cheng, Z.; Yang, Z.; Lin, J.; Wang, T.; Feng, X.; Gao, E.; Wang, H.; et al. Exosomal Mst1 transfer from cardiac microvascular endothelial cells to cardiomyocytes deteriorates diabetic cardiomyopathy. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 3639–3649. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Pei, H.; Watson, D.K.; Papas, T.S. EAP1/Daxx interacts with ETS1 and represses transcriptional activation of ETS1 target genes. Oncogene 2000, 19, 745–753. [Google Scholar] [CrossRef] [PubMed]
- Mahmud, I.; Tian, G.; Wang, J.; Hutchinson, T.E.; Kim, B.J.; Awasthee, N.; Hale, S.; Meng, C.; Moore, A.; Zhao, L.; et al. DAXX drives de novo lipogenesis and contributes to tumorigenesis. Nat. Commun. 2023, 14, 1927. [Google Scholar] [CrossRef]
- Gurer, C.; Berthoux, L.; Luban, J. Covalent modification of human immunodeficiency virus type 1 p6 by SUMO-1. J. Virol. 2005, 79, 910–917. [Google Scholar] [CrossRef]
- Li, X.D.; Makela, T.P.; Guo, D.; Soliymani, R.; Koistinen, V.; Vapalahti, O.; Vaheri, A.; Lankinen, H. Hantavirus nucleocapsid protein interacts with the Fas-mediated apoptosis enhancer Daxx. J. Gen. Virol. 2002, 83, 759–766. [Google Scholar] [CrossRef]
- Kuo, H.Y.; Chang, C.C.; Jeng, J.C.; Hu, H.M.; Lin, D.Y.; Maul, G.G.; Kwok, R.P.; Shih, H.M. SUMO modification negatively modulates the transcriptional activity of CREB-binding protein via the recruitment of Daxx. Proc. Natl. Acad. Sci. USA 2005, 102, 16973–16978. [Google Scholar] [CrossRef]
- Geiss-Friedlander, R.; Melchior, F. Concepts in sumoylation: A decade on. Nat. Rev. Mol. Cell Biol. 2007, 8, 947–956. [Google Scholar] [CrossRef]
- Spector, D.L. Nuclear domains. J. Cell Sci. 2001, 114, 2891–2893. [Google Scholar] [CrossRef]
- Wang, I.F.; Reddy, N.M.; Shen, C.K. Higher order arrangement of the eukaryotic nuclear bodies. Proc. Natl. Acad. Sci. USA 2002, 99, 13583–13588. [Google Scholar] [CrossRef]
- Lang, M.; Jegou, T.; Chung, I.; Richter, K.; Munch, S.; Udvarhelyi, A.; Cremer, C.; Hemmerich, P.; Engelhardt, J.; Hell, S.W.; et al. Three-dimensional organization of promyelocytic leukemia nuclear bodies. J. Cell Sci. 2010, 123, 392–400. [Google Scholar] [CrossRef]
- Kleijwegt, C.; Bressac, F.; Seurre, C.; Bouchereau, W.; Cohen, C.; Texier, P.; Simonet, T.; Schaeffer, L.; Lomonte, P.; Corpet, A. Interplay between PML NBs and HIRA for H3.3 dynamics following type I interferon stimulus. eLife 2023, 12, e80156. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Kao, H.Y. Post-translational modifications of PML: Consequences and implications. Front. Oncol. 2012, 2, 210. [Google Scholar] [CrossRef] [PubMed]
- Boddy, M.N.; Howe, K.; Etkin, L.D.; Solomon, E.; Freemont, P.S. PIC 1, a novel ubiquitin-like protein which interacts with the PML component of a multiprotein complex that is disrupted in acute promyelocytic leukaemia. Oncogene 1996, 13, 971–982. [Google Scholar] [PubMed]
- Muller, S.; Matunis, M.J.; Dejean, A. Conjugation with the ubiquitin-related modifier SUMO-1 regulates the partitioning of PML within the nucleus. EMBO J. 1998, 17, 61–70. [Google Scholar] [CrossRef]
- Zhong, S.; Müller, S.; Ronchetti, S.; Freemont, P.S.; Dejean, A.; Pandolfi, P.P. Role of SUMO-1-modified PML in nuclear body formation. Blood 2000, 95, 2748–2752. [Google Scholar] [CrossRef]
- Geng, Y.; Monajembashi, S.; Shao, A.; Cui, D.; He, W.; Chen, Z.; Hemmerich, P.; Tang, J. Contribution of the C-terminal regions of promyelocytic leukemia protein (PML) isoforms II and V to PML nuclear body formation. J. Biol. Chem. 2012, 287, 30729–30742. [Google Scholar] [CrossRef]
- Shen, T.H.; Lin, H.K.; Scaglioni, P.P.; Yung, T.M.; Pandolfi, P.P. The mechanisms of PML-nuclear body formation. Mol. Cell 2006, 24, 331–339. [Google Scholar] [CrossRef]
- Gao, C.; Ho, C.C.; Reineke, E.; Lam, M.; Cheng, X.; Stanya, K.J.; Liu, Y.; Chakraborty, S.; Shih, H.M.; Kao, H.Y. Histone deacetylase 7 promotes PML sumoylation and is essential for PML nuclear body formation. Mol. Cell. Biol. 2008, 28, 5658–5667. [Google Scholar] [CrossRef]
- Cappadocia, L.; Mascle, X.H.; Bourdeau, V.; Tremblay-Belzile, S.; Chaker-Margot, M.; Lussier-Price, M.; Wada, J.; Sakaguchi, K.; Aubry, M.; Ferbeyre, G.; et al. Structural and functional characterization of the phosphorylation-dependent interaction between PML and SUMO1. Structure 2015, 23, 126–138. [Google Scholar] [CrossRef]
- Matunis, M.J.; Zhang, X.D.; Ellis, N.A. SUMO: The glue that binds. Dev. Cell 2006, 11, 596–597. [Google Scholar] [CrossRef]
- Jeanne, M.; Lallemand-Breitenbach, V.; Ferhi, O.; Koken, M.; Le Bras, M.; Duffort, S.; Peres, L.; Berthier, C.; Soilihi, H.; Raught, B.; et al. PML/RARA oxidation and arsenic binding initiate the antileukemia response of As2O3. Cancer Cell 2010, 18, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Sahin, U.; Ferhi, O.; Jeanne, M.; Benhenda, S.; Berthier, C.; Jollivet, F.; Niwa-Kawakita, M.; Faklaris, O.; Setterblad, N.; de The, H.; et al. Oxidative stress-induced assembly of PML nuclear bodies controls sumoylation of partner proteins. J. Cell Biol. 2014, 204, 931–945. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Benhenda, S.; Wu, H.; Lallemand-Breitenbach, V.; Zhen, T.; Jollivet, F.; Peres, L.; Li, Y.; Chen, S.J.; Chen, Z.; et al. RING tetramerization is required for nuclear body biogenesis and PML sumoylation. Nat. Commun. 2018, 9, 1277. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ma, X.; Chen, Z.; Wu, H.; Wang, P.; Wu, W.; Cheng, N.; Zeng, L.; Zhang, H.; Cai, X.; et al. B1 oligomerization regulates PML nuclear body biogenesis and leukemogenesis. Nat. Commun. 2019, 10, 3789. [Google Scholar] [CrossRef]
- Ching, R.W.; Dellaire, G.; Eskiw, C.H.; Bazett-Jones, D.P. PML bodies: A meeting place for genomic loci? J. Cell Sci. 2005, 118, 847–854. [Google Scholar] [CrossRef]
- El, B.J.; Dianoux, L.; Chelbi-Alix, M.K. PML positively regulates interferon gamma signaling. Biochimie 2011, 93, 389–398. [Google Scholar]
- Ulbricht, T.; Alzrigat, M.; Horch, A.; Reuter, N.; von Mikecz, A.; Steimle, V.; Schmitt, E.; Kramer, O.H.; Stamminger, T.; Hemmerich, P. PML promotes MHC class II gene expression by stabilizing the class II transactivator. J. Cell Biol. 2012, 199, 49–63. [Google Scholar] [CrossRef]
- Chen, Y.; Wright, J.; Meng, X.; Leppard, K.N. Promyelocytic Leukemia Protein Isoform II Promotes Transcription Factor Recruitment To Activate Interferon Beta and Interferon-Responsive Gene Expression. Mol. Cell. Biol. 2015, 35, 1660–1672. [Google Scholar] [CrossRef]
- Dellaire, G.; Bazett-Jones, D.P. PML nuclear bodies: Dynamic sensors of DNA damage and cellular stress. Bioessays 2004, 26, 963–977. [Google Scholar] [CrossRef]
- Bernardi, R.; Pandolfi, P.P. Structure, dynamics and functions of promyelocytic leukaemia nuclear bodies. Nat. Rev. Mol. Cell Biol. 2007, 8, 1006–1016. [Google Scholar] [CrossRef]
- Chang, H.R.; Munkhjargal, A.; Kim, M.J.; Park, S.Y.; Jung, E.; Ryu, J.H.; Yang, Y.; Lim, J.S.; Kim, Y. The functional roles of PML nuclear bodies in genome maintenance. Mutat. Res. 2018, 809, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Corpet, A.; Kleijwegt, C.; Roubille, S.; Juillard, F.; Jacquet, K.; Texier, P.; Lomonte, P. PML nuclear bodies and chromatin dynamics: Catch me if you can! Nucleic Acids Res. 2020, 48, 11890–11912. [Google Scholar] [CrossRef] [PubMed]
- Spegg, V.; Altmeyer, M. Genome maintenance meets mechanobiology. Chromosoma 2024, 133, 15–36. [Google Scholar] [CrossRef] [PubMed]
- Chung, I.; Osterwald, S.; Deeg, K.I.; Rippe, K. PML body meets telomere: The beginning of an ALTernate ending? Nucleus 2012, 3, 263–275. [Google Scholar] [CrossRef]
- Ryabchenko, B.; Sroller, V.; Hornikova, L.; Lovtsov, A.; Forstova, J.; Huerfano, S. The interactions between PML nuclear bodies and small and medium size DNA viruses. Virol. J. 2023, 20, 82. [Google Scholar] [CrossRef]
- Pichler, A.; Fatouros, C.; Lee, H.; Eisenhardt, N. SUMO conjugation—A mechanistic view. Biomol. Concepts 2017, 8, 13–36. [Google Scholar] [CrossRef]
- Dellaire, G.; Bazett-Jones, D.P. Beyond repair foci: Subnuclear domains and the cellular response to DNA damage. Cell Cycle 2007, 6, 1864–1872. [Google Scholar] [CrossRef]
- Palvimo, J.J. PIAS proteins as regulators of small ubiquitin-related modifier (SUMO) modifications and transcription. Biochem. Soc. Trans. 2007, 35, 1405–1408. [Google Scholar] [CrossRef]
- Schmidt, D.; Muller, S. PIAS/SUMO: New partners in transcriptional regulation. Cell. Mol. Life Sci. 2003, 60, 2561–2574. [Google Scholar] [CrossRef]
- Ullmann, R.; Chien, C.D.; Avantaggiati, M.L.; Muller, S. An acetylation switch regulates SUMO-dependent protein interaction networks. Mol. Cell 2012, 46, 759–770. [Google Scholar] [CrossRef]
- Deribe, Y.L.; Pawson, T.; Dikic, I. Post-translational modifications in signal integration. Nat. Struct. Mol. Biol. 2010, 17, 666–672. [Google Scholar] [CrossRef] [PubMed]
- Scaglioni, P.P.; Yung, T.M.; Cai, L.F.; Erdjument-Bromage, H.; Kaufman, A.J.; Singh, B.; Teruya-Feldstein, J.; Tempst, P.; Pandolfi, P.P. A CK2-dependent mechanism for degradation of the PML tumor suppressor. Cell 2006, 126, 269–283. [Google Scholar] [CrossRef] [PubMed]
- Rabellino, A.; Carter, B.; Konstantinidou, G.; Wu, S.Y.; Rimessi, A.; Byers, L.A.; Heymach, J.V.; Girard, L.; Chiang, C.M.; Teruya-Feldstein, J.; et al. The SUMO E3-ligase PIAS1 regulates the tumor suppressor PML and its oncogenic counterpart PML-RARA. Cancer Res. 2012, 72, 2275–2284. [Google Scholar] [CrossRef]
- Cho, G.; Lim, Y.; Golden, J.A. SUMO interaction motifs in Sizn1 are required for promyelocytic leukemia protein nuclear body localization and for transcriptional activation. J. Biol. Chem. 2009, 284, 19592–19600. [Google Scholar] [CrossRef]
- Negorev, D.; Ishov, A.M.; Maul, G.G. Evidence for separate ND10-binding and homo-oligomerization domains of Sp100. J. Cell Sci. 2001, 114, 59–68. [Google Scholar] [CrossRef]
- Rasheed, Z.A.; Saleem, A.; Ravee, Y.; Pandolfi, P.P.; Rubin, E.H. The topoisomerase I-binding RING protein, topors, is associated with promyelocytic leukemia nuclear bodies. Exp. Cell Res. 2002, 277, 152–160. [Google Scholar] [CrossRef]
- Sung, K.S.; Lee, Y.A.; Kim, E.T.; Lee, S.R.; Ahn, J.H.; Choi, C.Y. Role of the SUMO-interacting motif in HIPK2 targeting to the PML nuclear bodies and regulation of p53. Exp. Cell. Res. 2011, 317, 1060–1070. [Google Scholar] [CrossRef]
- Mukhopadhyay, D.; Matunis, M.J. SUMmOning Daxx-mediated repression. Mol. Cell 2011, 42, 4–5. [Google Scholar] [CrossRef]
- Cheema, A.; Knights, C.D.; Rao, M.; Catania, J.; Perez, R.; Simons, B.; Dakshanamurthy, S.; Kolukula, V.K.; Tilli, M.; Furth, P.A.; et al. Functional mimicry of the acetylated C-terminal tail of p53 by a SUMO-1 acetylated domain, SAD. J. Cell. Physiol. 2010, 225, 371–384. [Google Scholar] [CrossRef]
- Mascle, X.H.; Gagnon, C.; Wahba, H.M.; Lussier-Price, M.; Cappadocia, L.; Sakaguchi, K.; Omichinski, J.G. Acetylation of SUMO1 Alters Interactions with the SIMs of PML and Daxx in a Protein-Specific Manner. Structure 2020, 28, 157–168.e155. [Google Scholar] [CrossRef]
- Ji, Z.; Degerny, C.; Vintonenko, N.; Deheuninck, J.; Foveau, B.; Leroy, C.; Coll, J.; Tulasne, D.; Baert, J.L.; Fafeur, V. Regulation of the Ets-1 transcription factor by sumoylation and ubiquitinylation. Oncogene 2007, 26, 395–406. [Google Scholar] [CrossRef] [PubMed]
- Hua, G.; Ganti, K.P.; Chambon, P. Glucocorticoid-induced tethered transrepression requires SUMOylation of GR and formation of a SUMO-SMRT/NCoR1-HDAC3 repressing complex. Proc. Natl. Acad. Sci. USA 2016, 113, E635–E643. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.Y.; Lai, M.Z.; Ann, D.K.; Shih, H.M. Promyelocytic leukemia protein (PML) functions as a glucocorticoid receptor co-activator by sequestering Daxx to the PML oncogenic domains (PODs) to enhance its transactivation potential. J. Biol. Chem. 2003, 278, 15958–15965. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.S.; Chang, C.; Liu, D.; Derynck, R. Sumoylation of Smad4, the common Smad mediator of transforming growth factor-β family signaling. J. Biol. Chem. 2003, 278, 27853–27863. [Google Scholar] [CrossRef]
- Chang, C.C.; Lin, D.Y.; Fang, H.I.; Chen, R.H.; Shih, H.M. Daxx mediates the small ubiquitin-like modifier-dependent transcriptional repression of Smad4. J. Biol. Chem. 2005, 280, 10164–10173. [Google Scholar] [CrossRef]
- Lin, D.Y.; Fang, H.I.; Ma, A.H.; Huang, Y.S.; Pu, Y.S.; Jenster, G.; Kung, H.J.; Shih, H.M. Negative modulation of androgen receptor transcriptional activity by Daxx. Mol. Cell. Biol. 2004, 24, 10529–10541. [Google Scholar] [CrossRef]
- Poukka, H.; Karvonen, U.; Janne, O.A.; Palvimo, J.J. Covalent modification of the androgen receptor by small ubiquitin-like modifier 1 (SUMO-1). Proc. Natl. Acad. Sci. USA 2000, 97, 14145–14150. [Google Scholar] [CrossRef]
- Meloni, A.; Fiorillo, E.; Corda, D.; Incani, F.; Serra, M.L.; Contini, A.; Cao, A.; Rosatelli, M.C. DAXX is a new AIRE-interacting protein. J. Biol. Chem. 2010, 285, 13012–13021. [Google Scholar] [CrossRef]
- Lin, C.W.; Wang, L.K.; Wang, S.P.; Chang, Y.L.; Wu, Y.Y.; Chen, H.Y.; Hsiao, T.H.; Lai, W.Y.; Lu, H.H.; Chang, Y.H.; et al. Daxx inhibits hypoxia-induced lung cancer cell metastasis by suppressing the HIF-1alpha/HDAC1/Slug axis. Nat. Commun. 2016, 7, 13867. [Google Scholar] [CrossRef]
- Xie, Y.; Liu, S.; Lu, W.; Yang, Q.; Williams, K.D.; Binhazim, A.A.; Carver, B.S.; Matusik, R.J.; Chen, Z. Slug regulates E-cadherin repression via p19Arf in prostate tumorigenesis. Mol. Oncol. 2014, 8, 1355–1364. [Google Scholar] [CrossRef]
- Yamamoto, H.; Ihara, M.; Matsuura, Y.; Kikuchi, A. Sumoylation is involved in β-catenin-dependent activation of Tcf-4. EMBO J. 2003, 22, 2047–2059. [Google Scholar] [CrossRef] [PubMed]
- Tzeng, S.L.; Cheng, Y.W.; Li, C.H.; Lin, Y.S.; Hsu, H.C.; Kang, J.J. Physiological and functional interactions between Tcf4 and Daxx in colon cancer cells. J. Biol. Chem. 2006, 281, 15405–15411. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Wang, M.; Li, J.; Xiao, M.; Chin, Y.E.; Cheng, J.; Yeh, E.T.; Yang, J.; Yi, J. SUMOylation and SENP3 regulate STAT3 activation in head and neck cancer. Oncogene 2016, 35, 5826–5838. [Google Scholar] [CrossRef] [PubMed]
- Muromoto, R.; Nakao, K.; Watanabe, T.; Sato, N.; Sekine, Y.; Sugiyama, K.; Oritani, K.; Shimoda, K.; Matsuda, T. Physical and functional interactions between Daxx and STAT3. Oncogene 2006, 25, 2131–2136. [Google Scholar] [CrossRef]
- Feng, Z.J.; Gurung, B.; Jin, G.H.; Yang, X.L.; Hua, X.X. SUMO modification of menin. Am. J. Cancer Res. 2013, 3, 96–106. [Google Scholar]
- Feng, Z.; Wang, L.; Sun, Y.; Jiang, Z.; Domsic, J.; An, C.; Xing, B.; Tian, J.; Liu, X.; Metz, D.C.; et al. Menin and Daxx Interact to Suppress Neuroendocrine Tumors through Epigenetic Control of the Membrane Metallo-Endopeptidase. Cancer Res. 2017, 77, 401–411. [Google Scholar] [CrossRef]
- Liu, Y.; Bridges, R.; Wortham, A.; Kulesz-Martin, M. NF-kappaB repression by PIAS3 mediated RelA SUMOylation. PLoS ONE 2012, 7, e37636. [Google Scholar]
- Maroulakou, I.G.; Bowe, D.B. Expression and function of Ets transcription factors in mammalian development: A regulatory network. Oncogene 2000, 19, 6432–6442. [Google Scholar] [CrossRef]
- Zhang, C.; Kavurma, M.M.; Lai, A.; Khachigian, L.M. Ets-1 protects vascular smooth muscle cells from undergoing apoptosis by activating p21WAF1/Cip1: ETS-1 regulates basal and and inducible p21WAF1/Cip1: ETS-1 regulates basal and inducible p21WAF1/Cip1 transcription via distinct cis-acting elements in the p21WAF/Cip1 promoter. J. Biol. Chem. 2003, 278, 27903–27909. [Google Scholar]
- Marziali, G.; Perrotti, E.; Ilari, R.; Lulli, V.; Coccia, E.M.; Moret, R.; Kuhn, L.C.; Testa, U.; Battistini, A. Role of Ets-1 in transcriptional regulation of transferrin receptor and erythroid differentiation. Oncogene 2002, 21, 7933–7944. [Google Scholar] [CrossRef]
- Naito, S.; Shimizu, S.; Matsuu, M.; Nakashima, M.; Nakayama, T.; Yamashita, S.; Sekine, I. Ets-1 upregulates matrix metalloproteinase-1 expression through extracellular matrix adhesion in vascular endothelial cells. Biochem. Biophys. Res. Commun. 2002, 291, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Iwasaka, C.; Tanaka, K.; Abe, M.; Sato, Y. Ets-1 regulates angiogenesis by inducing the expression of urokinase-type plasminogen activator and matrix metalloproteinase-1 and the migration of vascular endothelial cells. J. Cell. Physiol. 1996, 169, 522–531. [Google Scholar] [CrossRef]
- Chen, Z.; Fisher, R.J.; Riggs, C.W.; Rhim, J.S.; Lautenberger, J.A. Inhibition of vascular endothelial growth factor-induced endothelial cell migration by ETS1 antisense oligonucleotides. Cancer Res. 1997, 57, 2013–2019. [Google Scholar]
- Oda, N.; Abe, M.; Sato, Y. ETS-1 converts endothelial cells to the angiogenic phenotype by inducing the expression of matrix metalloproteinases and integrin β3. J. Cell. Physiol. 1999, 178, 121–132. [Google Scholar] [CrossRef]
- Xu, D.; Wilson, T.J.; Chan, D.; De Luca, E.; Zhou, J.; Hertzog, P.J.; Kola, I. Ets1 is required for p53 transcriptional activity in UV-induced apoptosis in embryonic stem cells. EMBO J. 2002, 21, 4081–4093. [Google Scholar] [CrossRef]
- Timmermans, S.; Souffriau, J.; Libert, C. A General Introduction to Glucocorticoid Biology. Front. Immunol. 2019, 10, 1545. [Google Scholar] [CrossRef]
- Cruz-Topete, D.; Cidlowski, J.A. One hormone, two actions: Anti- and pro-inflammatory effects of glucocorticoids. Neuroimmunomodulation 2015, 22, 20–32. [Google Scholar] [CrossRef]
- Wang, J.C.; Gray, N.E.; Kuo, T.; Harris, C.A. Regulation of triglyceride metabolism by glucocorticoid receptor. Cell Biosci. 2012, 2, 19. [Google Scholar] [CrossRef]
- Tassabehji, M.; Read, A.P.; Newton, V.E.; Harris, R.; Balling, R.; Gruss, P.; Strachan, T. Waardenburg’s syndrome patients have mutations in the human homologue of the Pax-3 paired box gene. Nature 1992, 355, 635–636. [Google Scholar] [CrossRef]
- Qi, J.; Yan, L.; Sun, J.; Huang, C.; Su, B.; Cheng, J.; Shen, L. SUMO-specific protease 1 regulates germinal center B cell response through deSUMOylation of PAX5. Proc. Natl. Acad. Sci. USA 2024, 121, e2314619121. [Google Scholar] [CrossRef]
- Yan, Q.; Gong, L.; Deng, M.; Zhang, L.; Sun, S.; Liu, J.; Ma, H.; Yuan, D.; Chen, P.C.; Hu, X.; et al. Sumoylation activates the transcriptional activity of Pax-6, an important transcription factor for eye and brain development. Proc. Natl. Acad. Sci. USA 2010, 107, 21034–21039. [Google Scholar] [CrossRef] [PubMed]
- Luan, Z.; Liu, Y.; Stuhlmiller, T.J.; Marquez, J.; Garcia-Castro, M.I. SUMOylation of Pax7 is essential for neural crest and muscle development. Cell. Mol. Life Sci. 2013, 70, 1793–1806. [Google Scholar] [CrossRef] [PubMed]
- Emelyanov, A.V.; Kovac, C.R.; Sepulveda, M.A.; Birshtein, B.K. The interaction of Pax5 (BSAP) with Daxx can result in transcriptional activation in B cells. J. Biol. Chem. 2002, 277, 11156–11164. [Google Scholar] [CrossRef] [PubMed]
- Igalouzene, R.; Hernandez-Vargas, H.; Benech, N.; Guyennon, A.; Bauche, D.; Barrachina, C.; Dubois, E.; Marie, J.C.; Soudja, S.M. SMAD4 TGF-β-independent function preconditions naive CD8+ T cells to prevent severe chronic intestinal inflammation. J. Clin. Invest. 2022, 132, e151020. [Google Scholar] [CrossRef]
- Li, T.P.; Sun, S.W.; Xiong, G.Z.; Qiu, F.; Yang, D.M.; Sun, S.Y.; Xie, X.J.; Liao, D.F.; Chen, J.X.; Tuo, Q.H. Direct Interaction of Daxx and Androgen Receptor Is Required for Their Regulatory Activity in Cholesterol Biosynthesis. Pharmacology 2021, 106, 29–36. [Google Scholar] [CrossRef]
- Sun, S.; Wen, J.; Qiu, F.; Yin, Y.; Xu, G.; Li, T.; Nie, J.; Xiong, G.; Zhang, C.; Liao, D.; et al. Identification of the C-terminal domain of Daxx acts as a potential regulator of intracellular cholesterol synthesis in HepG2 cells. Biochem. Biophys. Res. Commun. 2016, 480, 139–145. [Google Scholar] [CrossRef]
- Anderson, M.S.; Venanzi, E.S.; Klein, L.; Chen, Z.; Berzins, S.P.; Turley, S.J.; von Boehmer, H.; Bronson, R.; Dierich, A.; Benoist, C.; et al. Projection of an immunological self shadow within the thymus by the aire protein. Science 2002, 298, 1395–1401. [Google Scholar] [CrossRef]
- Sansom, S.N.; Shikama-Dorn, N.; Zhanybekova, S.; Nusspaumer, G.; Macaulay, I.C.; Deadman, M.E.; Heger, A.; Ponting, C.P.; Hollander, G.A. Population and single-cell genomics reveal the Aire dependency, relief from Polycomb silencing, and distribution of self-antigen expression in thymic epithelia. Genome Res. 2014, 24, 1918–1931. [Google Scholar] [CrossRef]
- Meredith, M.; Zemmour, D.; Mathis, D.; Benoist, C. Aire controls gene expression in the thymic epithelium with ordered stochasticity. Nat. Immunol. 2015, 16, 942–949. [Google Scholar] [CrossRef]
- Barrallo-Gimeno, A.; Nieto, M.A. The Snail genes as inducers of cell movement and survival: Implications in development and cancer. Development 2005, 132, 3151–3161. [Google Scholar] [CrossRef]
- Nieto, M.A. The snail superfamily of zinc-finger transcription factors. Nat. Rev. Mol. Cell Biol. 2002, 3, 155–166. [Google Scholar] [CrossRef]
- Hung, P.F.; Hong, T.M.; Chang, C.C.; Hung, C.L.; Hsu, Y.L.; Chang, Y.L.; Wu, C.T.; Chang, G.C.; Chan, N.L.; Yu, S.L.; et al. Hypoxia-induced Slug SUMOylation enhances lung cancer metastasis. J. Exp. Clin. Cancer Res. 2019, 38, 5. [Google Scholar] [CrossRef] [PubMed]
- Obradovic, D.; Tirard, M.; Nemethy, Z.; Hirsch, O.; Gronemeyer, H.; Almeida, O.F. DAXX, FLASH, and FAF-1 modulate mineralocorticoid and glucocorticoid receptor-mediated transcription in hippocampal cells—Toward a basis for the opposite actions elicited by two nuclear receptors? Mol. Pharmacol. 2004, 65, 761–769. [Google Scholar] [CrossRef] [PubMed]
- Muromoto, R.; Kuroda, M.; Togi, S.; Sekine, Y.; Nanbo, A.; Shimoda, K.; Oritani, K.; Matsuda, T. Functional involvement of Daxx in gp130-mediated cell growth and survival in BaF3 cells. Eur. J. Immunol. 2010, 40, 3570–3580. [Google Scholar] [CrossRef] [PubMed]
- Chao, W.; Shen, Y.; Li, L.; Zhao, H.; Meiler, S.E.; Cook, S.A.; Rosenzweig, A. Fas-associated death-domain protein inhibits TNF-alpha mediated NF-kappaB activation in cardiomyocytes. Am. J. Physiol. Heart Circ. Physiol. 2005, 289, H2073–H2080. [Google Scholar] [CrossRef]
- Park, J.; Lee, J.H.; La, M.; Jang, M.J.; Chae, G.W.; Kim, S.B.; Tak, H.; Jung, Y.; Byun, B.; Ahn, J.K.; et al. Inhibition of NF-kappaB acetylation and its transcriptional activity by Daxx. J. Mol. Biol. 2007, 368, 388–397. [Google Scholar] [CrossRef]
- Puto, L.A.; Reed, J.C. Daxx represses RelB target promoters via DNA methyltransferase recruitment and DNA hypermethylation. Genes Dev. 2008, 22, 998–1010. [Google Scholar] [CrossRef]
- Zhao, Y.Q.; Jin, H.R.; Kim, D.; Jung, S.H.; Liu, S.; Wan, J.; Lo, H.Y.; Fu, X.Q.; Wang, Q.; Hao, C.; et al. SUMO1 degrader induces ER stress and ROS accumulation through deSUMOylation of TCF4 and inhibition of its transcription of StarD7 in colon cancer. Mol. Carcinog. 2023, 62, 1249–1262. [Google Scholar] [CrossRef]
- Ihara, M.; Yamamoto, H.; Kikuchi, A. SUMO-1 modification of PIASy, an E3 ligase, is necessary for PIASy-dependent activation of Tcf-4. Mol. Cell. Biol. 2005, 25, 3506–3518. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, J.; Xue, Z.; Chang, M.; Feng, X.; Cai, Y.; Bai, L.; Wang, W.; Liu, E.; Zhao, S.; et al. Deficiency of protein inhibitor of activated STAT3 exacerbates atherosclerosis by modulating VSMC phenotypic switching. Atherosclerosis 2023, 380, 117195. [Google Scholar] [CrossRef]
- Nayak, A.; Viale-Bouroncle, S.; Morsczeck, C.; Muller, S. The SUMO-specific isopeptidase SENP3 regulates MLL1/MLL2 methyltransferase complexes and controls osteogenic differentiation. Mol. Cell 2014, 55, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wu, Z.; Han, D.; Wei, C.; Liang, Y.; Jiang, T.; Chen, L.; Sha, M.; Cao, Y.; Huang, F.; et al. Mesencephalic Astrocyte-Derived Neurotrophic Factor Inhibits Liver Cancer Through Small Ubiquitin-Related Modifier (SUMO)ylation-Related Suppression of NF-kappaB/Snail Signaling Pathway and Epithelial-Mesenchymal Transition. Hepatology 2020, 71, 1262–1278. [Google Scholar] [CrossRef] [PubMed]
- Kracklauer, M.P.; Schmidt, C. At the crossroads of SUMO and NF-kappaB. Mol. Cancer 2003, 2, 39. [Google Scholar] [CrossRef]
- Chen, S.; Fu, X.; Wang, R.; Li, M.; Yan, X.; Yue, Z.; Chen, S.W.; Dong, M.; Xu, A.; Huang, S. SUMO and PIAS repress NF-kappaB activation in a basal chordate. Fish Shellfish Immunol. 2023, 137, 108754. [Google Scholar] [CrossRef]
- Wang, C.H.; Hung, P.W.; Chiang, C.W.; Lombes, M.; Chen, C.H.; Lee, K.H.; Lo, Y.C.; Wu, M.H.; Chang, W.C.; Lin, D.Y. Identification of two independent SUMO-interacting motifs in Fas-associated factor 1 (FAF1): Implications for mineralocorticoid receptor (MR)-mediated transcriptional regulation. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 1282–1297. [Google Scholar] [CrossRef]
- Wu, C.; Ding, H.; Wang, S.; Li, Y.; Liu, S.B.; Wang, X.; Zheng, J.; Xue, T.; Amin, H.M.; Song, Y.H.; et al. DAXX inhibits cancer stemness and epithelial-mesenchymal transition in gastric cancer. Br. J. Cancer 2020, 122, 1477–1485. [Google Scholar] [CrossRef]
- Peiffer, D.S.; Wyatt, D.; Zlobin, A.; Piracha, A.; Ng, J.; Dingwall, A.K.; Albain, K.S.; Osipo, C. DAXX Suppresses Tumor-Initiating Cells in Estrogen Receptor-Positive Breast Cancer Following Endocrine Therapy. Cancer Res. 2019, 79, 4965–4977. [Google Scholar] [CrossRef]
- Muromoto, R.; Sugiyama, K.; Takachi, A.; Imoto, S.; Sato, N.; Yamamoto, T.; Oritani, K.; Shimoda, K.; Matsuda, T. Physical and functional interactions between Daxx and DNA methyltransferase 1-associated protein, DMAP1. J. Immunol. 2004, 172, 2985–2993. [Google Scholar] [CrossRef]
- Ecsedy, J.A.; Michaelson, J.S.; Leder, P. Homeodomain-interacting protein kinase 1 modulates Daxx localization, phosphorylation, and transcriptional activity. Mol. Cell. Biol. 2003, 23, 950–960. [Google Scholar] [CrossRef]
- Bogolyubova, I.O.; Sailau, Z.K.; Bogolyubov, D.S. The dynamics of DAXX protein distribution in the nucleus of mouse early embryos. Acta Histochem. 2019, 121, 522–529. [Google Scholar] [CrossRef]
- Muromoto, R. [Death domain-associated protein (DAXX)-mediated regulation of transcription and cell death]. Yakugaku Zasshi 2012, 132, 979–984. [Google Scholar] [CrossRef] [PubMed]
- Meinecke, I.; Cinski, A.; Baier, A.; Peters, M.A.; Dankbar, B.; Wille, A.; Drynda, A.; Mendoza, H.; Gay, R.E.; Hay, R.T.; et al. Modification of nuclear PML protein by SUMO-1 regulates Fas-induced apoptosis in rheumatoid arthritis synovial fibroblasts. Proc. Natl. Acad. Sci. USA 2007, 104, 5073–5078. [Google Scholar] [CrossRef] [PubMed]
- Song, J.J.; Lee, Y.J. Tryptophan 621 and serine 667 residues of Daxx regulate its nuclear export during glucose deprivation. J. Biol. Chem. 2004, 279, 30573–30578. [Google Scholar] [CrossRef] [PubMed]
- Junn, E.; Taniguchi, H.; Jeong, B.S.; Zhao, X.; Ichijo, H.; Mouradian, M.M. Interaction of DJ-1 with Daxx inhibits apoptosis signal-regulating kinase 1 activity and cell death. Proc. Natl. Acad. Sci. USA 2005, 102, 9691–9696. [Google Scholar] [CrossRef]
- Saeed, U.; Ray, A.; Valli, R.K.; Kumar, A.M.; Ravindranath, V. DJ-1 loss by glutaredoxin but not glutathione depletion triggers Daxx translocation and cell death. Antioxid. Redox Signal. 2010, 13, 127–144. [Google Scholar] [CrossRef]
- Lind-Holm Mogensen, F.; Sousa, C.; Ameli, C.; Badanjak, K.; Pereira, S.L.; Muller, A.; Kaoma, T.; Coowar, D.; Scafidi, A.; Poovathingal, S.K.; et al. PARK7/DJ-1 deficiency impairs microglial activation in response to LPS-induced inflammation. J. Neuroinflammation 2024, 21, 174. [Google Scholar] [CrossRef]
- Guo, T.; Zhou, L.; Xiong, M.; Xiong, J.; Huang, J.; Li, Y.; Zhang, G.; Chen, G.; Wang, Z.H.; Xiao, T.; et al. N-homocysteinylation of DJ-1 promotes neurodegeneration in Parkinson’s disease. Aging Cell 2024, 23, e14124. [Google Scholar] [CrossRef]
- Karunakaran, S.; Diwakar, L.; Saeed, U.; Agarwal, V.; Ramakrishnan, S.; Iyengar, S.; Ravindranath, V. Activation of apoptosis signal regulating kinase 1 (ASK1) and translocation of death-associated protein, Daxx, in substantia nigra pars compacta in a mouse model of Parkinson’s disease: Protection by alpha-lipoic acid. FASEB J. 2007, 21, 2226–2236. [Google Scholar] [CrossRef]
- Hwang, S.; Song, S.; Hong, Y.K.; Choi, G.; Suh, Y.S.; Han, S.Y.; Lee, M.; Park, S.H.; Lee, J.H.; Lee, S.; et al. Drosophila DJ-1 decreases neural sensitivity to stress by negatively regulating Daxx-like protein through dFOXO. PLoS Genet. 2013, 9, e1003412. [Google Scholar] [CrossRef]
- Grimwade, D.; Lo Coco, F. Acute promyelocytic leukemia: A model for the role of molecular diagnosis and residual disease monitoring in directing treatment approach in acute myeloid leukemia. Leukemia 2002, 16, 1959–1973. [Google Scholar] [CrossRef]
- Nowak, D.; Stewart, D.; Koeffler, H.P. Differentiation therapy of leukemia: 3 decades of development. Blood 2009, 113, 3655–3665. [Google Scholar] [CrossRef] [PubMed]
- Sirulnik, A.; Melnick, A.; Zelent, A.; Licht, J.D. Molecular pathogenesis of acute promyelocytic leukaemia and APL variants. Best. Pract. Res. Clin. Haematol. 2003, 16, 387–408. [Google Scholar] [CrossRef]
- Salsman, J.; Rapkin, L.M.; Margam, N.N.; Duncan, R.; Bazett-Jones, D.P.; Dellaire, G. Myogenic differentiation triggers PML nuclear body loss and DAXX relocalization to chromocentres. Cell Death Dis. 2017, 8, e2724. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.; Salomoni, P.; Ronchetti, S.; Guo, A.; Ruggero, D.; Pandolfi, P.P. Promyelocytic leukemia protein (PML) and Daxx participate in a novel nuclear pathway for apoptosis. J. Exp. Med. 2000, 191, 631–640. [Google Scholar] [CrossRef]
- Torii, S.; Egan, D.A.; Evans, R.A.; Reed, J.C. Human Daxx regulates Fas-induced apoptosis from nuclear PML oncogenic domains (PODs). EMBO J. 1999, 18, 6037–6049. [Google Scholar] [CrossRef]
- Wethkamp, N.; Klempnauer, K.H. Daxx is a transcriptional repressor of CCAAT/enhancer-binding protein β. J. Biol. Chem. 2009, 284, 28783–28794. [Google Scholar] [CrossRef]
- Duprez, E.; Wagner, K.; Koch, H.; Tenen, D.G. C/EBPβ: A major PML-RARA-responsive gene in retinoic acid-induced differentiation of APL cells. EMBO J. 2003, 22, 5806–5816. [Google Scholar] [CrossRef]
- Lin, R.J.; Egan, D.A.; Evans, R.M. Molecular genetics of acute promyelocytic leukemia. Trends Genet. 1999, 15, 179–184. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Joshi, S.S.; Badgwell, B.D. Current treatment and recent progress in gastric cancer. CA Cancer J. Clin. 2021, 71, 264–279. [Google Scholar] [CrossRef]
- Xu, J.F.; Zhao, Z.G.; Ye, L.L.; Zhuge, W.; Han, Z.; Zhang, T.M.; Ye, S.S.; Chen, W.J.; Zhu, S.; Shi, L.; et al. Prognostic significance of Daxx NCR (Nuclear/Cytoplasmic Ratio) in gastric cancer. Cancer Med. 2017, 6, 2063–2075. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Sun, X.; Xie, W.; Chen, S.; Hu, Y.; Xing, D.; Xu, J.; Chen, X.; Zhao, Z.; Han, Z.; et al. Opposing biological functions of the cytoplasm and nucleus DAXX modified by SUMO-2/3 in gastric cancer. Cell Death Dis. 2020, 11, 514. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.S.; Shih, H.M. Daxx positively modulates β-catenin/TCF4-mediated transcriptional potential. Biochem. Biophys. Res. Commun. 2009, 386, 762–768. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Daxx Interacting Transcription Factor | Whether It Is SUMOylated or Not | SUMOylation Site | Effect | References |
|---|---|---|---|---|
| ETS1 | Yes | Lys15, Lys227 | transcriptional repression | [62,111] |
| GR | Yes | Lys277, Lys293 | transcriptional repression | [112,113] |
| Pax3 | unclear | transcriptional repression | [43] | |
| SMAD4 | Yes | Lys159, Lys113 | transcriptional repression | [114,115] |
| AR | Yes | Lys388, Lys521 | transcriptional repression | [116,117] |
| AIRE | unclear | transcriptional repression | [118] | |
| SNAI2 (Slug) | yes | Lys192 | transcriptional repression | [119,120] |
| TCF7L2 (TCF4) | Yes | Lys297 | transcriptional repression | [121,122] |
| STAT3 | Yes | Lys451 | transcriptional repression | [123,124] |
| MENIN | yes | Lys591 | transcriptional repression | [125,126] |
| RELB (NF-kappaB) | Yes | unclear | transcriptional repression | [127] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, J.; Liu, T.; Yang, D.; Tuo, Q. The Dynamic Regulation of Daxx-Mediated Transcriptional Inhibition by SUMO and PML NBs. Int. J. Mol. Sci. 2025, 26, 6703. https://doi.org/10.3390/ijms26146703
Gao J, Liu T, Yang D, Tuo Q. The Dynamic Regulation of Daxx-Mediated Transcriptional Inhibition by SUMO and PML NBs. International Journal of Molecular Sciences. 2025; 26(14):6703. https://doi.org/10.3390/ijms26146703
Chicago/Turabian StyleGao, Jiatao, Tingting Liu, Dongmei Yang, and Qinhui Tuo. 2025. "The Dynamic Regulation of Daxx-Mediated Transcriptional Inhibition by SUMO and PML NBs" International Journal of Molecular Sciences 26, no. 14: 6703. https://doi.org/10.3390/ijms26146703
APA StyleGao, J., Liu, T., Yang, D., & Tuo, Q. (2025). The Dynamic Regulation of Daxx-Mediated Transcriptional Inhibition by SUMO and PML NBs. International Journal of Molecular Sciences, 26(14), 6703. https://doi.org/10.3390/ijms26146703