
The Role of Cytokines in Orthodontic Tooth Movement
 , , ,
, , ,
Abstract
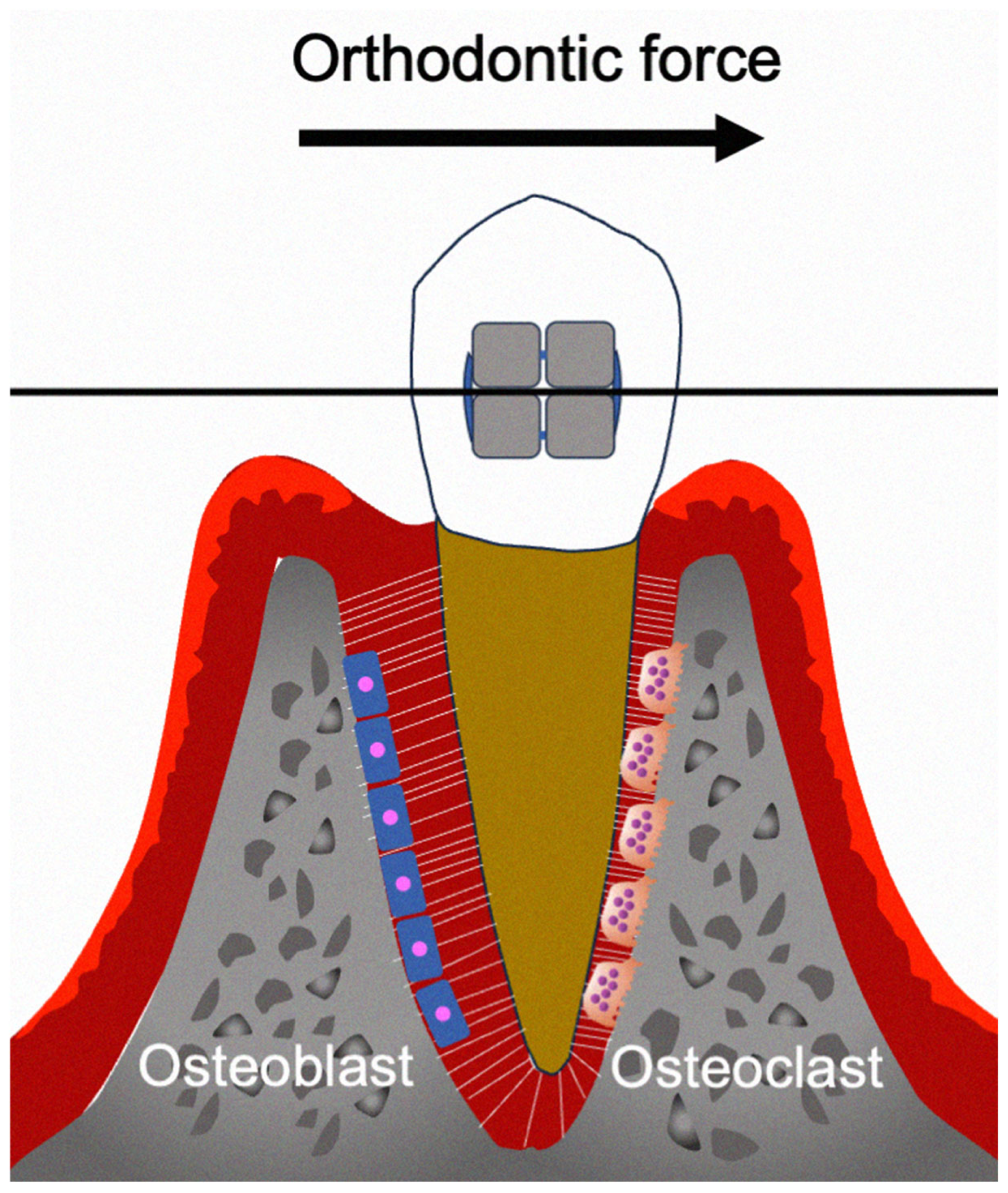
1. Introduction
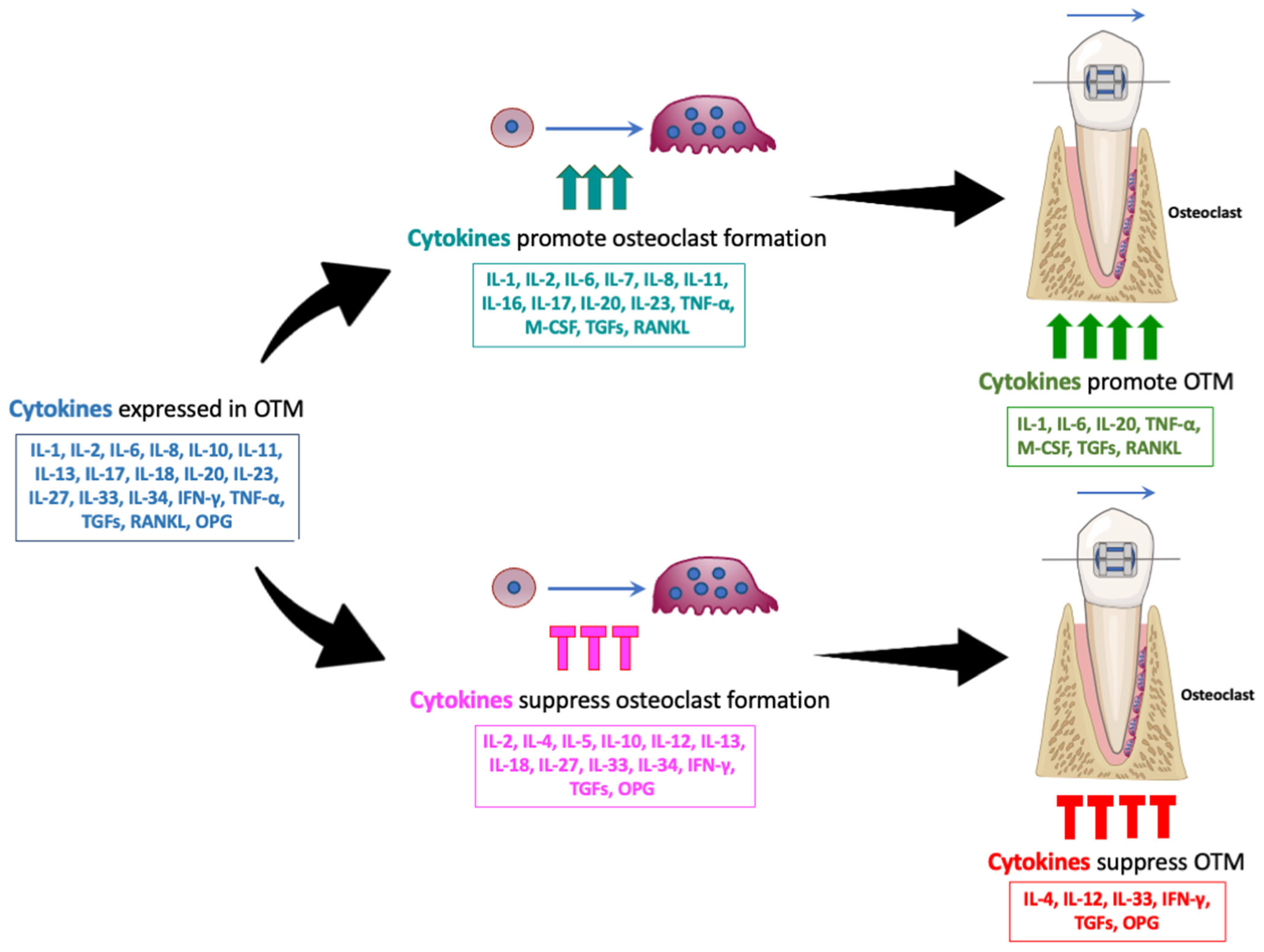
2. Cytokines Involved in Orthodontic Tooth Movement
2.1. IL-1β
2.2. IL-2
2.3. IL-4
2.4. IL-5
2.5. IL-6
2.6. IL-7
2.7. IL-8
2.8. IL-10
2.9. IL-11
2.10. IL-12
2.11. IL-13
2.12. IL-16
2.13. IL-17
2.14. IL-18
2.15. IL-20
2.16. IL-23
2.17. IL-27
2.18. IL-33
2.19. IL-34
2.20. IFN-γ
2.21. TNF-α
2.22. M-CSF
2.23. TGFs
2.24. RANKL and OPG
3. Summary and Limitations
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, Y.; Guo, W.; Li, X.; Zhang, J.; Sun, M.; Tang, Z.; Ran, W.; Yang, K.; Huang, G.; Li, L. Expert consensus on the clinical application of recombinant adenovirus human p53 for head and neck cancers. Int. J. Oral Sci. 2021, 13, 38. [Google Scholar] [CrossRef]
- Maltha, J.C.; Kuijpers-Jagtman, A.M. Mechanobiology of orthodontic tooth movement: An update. J. World Fed. Orthod. 2023, 12, 156–160. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, V.; Davidovitch, Z. Cellular, molecular, and tissue-level reactions to orthodontic force. Am. J. Orthod. Dentofac. Orthop. 2006, 129, 469.e1–469.e32. [Google Scholar] [CrossRef]
- Chaushu, S.; Klein, Y.; Mandelboim, O.; Barenholz, Y.; Fleissig, O. Immune Changes Induced by Orthodontic Forces: A Critical Review. J. Dent. Res. 2022, 101, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Cattaneo, P.M.; Dalstra, M.; Melsen, B. The finite element method: A tool to study orthodontic tooth movement. J. Dent. Res. 2005, 84, 428–433. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Fukasawa, S. Is Inflammation a Friend or Foe for Orthodontic Treatment?: Inflammation in Orthodontically Induced Inflammatory Root Resorption and Accelerating Tooth Movement. Int. J. Mol. Sci. 2021, 22, 2388. [Google Scholar] [CrossRef]
- Yu, Q.Y.; Huang, Y.P.; Li, W.R. Extracellular Matrix Remodelling of the Periodontium under Orthodontic Force. Chin. J. Dent. Res. 2024, 27, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Min, Q.; Li, X.; Liu, L.; Lv, Y.; Xu, W.; Liu, X.; Wang, H. Immune System Acts on Orthodontic Tooth Movement: Cellular and Molecular Mechanisms. BioMed Res. Int. 2022, 2022, 9668610. [Google Scholar] [CrossRef]
- Teitelbaum, S.L. Bone resorption by osteoclasts. Science 2000, 289, 1504–1508. [Google Scholar] [CrossRef]
- Kobayashi, K.; Takahashi, N.; Jimi, E.; Udagawa, N.; Takami, M.; Kotake, S.; Nakagawa, N.; Kinosaki, M.; Yamaguchi, K.; Shima, N.; et al. Tumor necrosis factor alpha stimulates osteoclast differentiation by a mechanism independent of the ODF/RANKL-RANK interaction. J. Exp. Med. 2000, 191, 275–286. [Google Scholar] [CrossRef]
- Liu, C.; Chu, D.; Kalantar-Zadeh, K.; George, J.; Young, H.A.; Liu, G. Cytokines: From Clinical Significance to Quantification. Adv. Sci. 2021, 8, e2004433. [Google Scholar] [CrossRef] [PubMed]
- Borish, L.C.; Steinke, J.W. 2. Cytokines and chemokines. J. Allergy Clin. Immunol. 2003, 111 (Suppl. S2), S460–S475. [Google Scholar] [CrossRef] [PubMed]
- Garlet, T.P.; Coelho, U.; Silva, J.S.; Garlet, G.P. Cytokine expression pattern in compression and tension sides of the periodontal ligament during orthodontic tooth movement in humans. Eur. J. Oral Sci. 2007, 115, 355–362. [Google Scholar] [CrossRef]
- Yamaguchi, M. RANK/RANKL/OPG during orthodontic tooth movement. Orthod. Craniofacial Res. 2009, 12, 113–119. [Google Scholar] [CrossRef]
- Rauner, M.; Sipos, W.; Pietschmann, P. Osteoimmunology. Int. Arch. Allergy Immunol. 2007, 143, 31–48. [Google Scholar] [CrossRef]
- Danz, J.C.; Degen, M. Selective modulation of the bone remodeling regulatory system through orthodontic tooth movement—A review. Front. Oral Health 2025, 6, 1472711. [Google Scholar] [CrossRef] [PubMed]
- Pereira, L.J.; Macari, S.; Coimbra, C.C.; Pereira, T.; Barrioni, B.R.; Gomez, R.S.; Silva, T.A.; Paiva, S.M. Aerobic and resistance training improve alveolar bone quality and interferes with bone-remodeling during orthodontic tooth movement in mice. Bone 2020, 138, 115496. [Google Scholar] [CrossRef]
- Dawalibi, A.; Alosaimi, A.A.; Mohammad, K.S. Balancing the Scales: The Dual Role of Interleukins in Bone Metastatic Microenvironments. Int. J. Mol. Sci. 2024, 25, 8163. [Google Scholar] [CrossRef]
- Han, P.; Liu, X.; He, J.; Han, L.; Li, J. Overview of mechanisms and novel therapies on rheumatoid arthritis from a cellular perspective. Front. Immunol. 2024, 15, 1461756. [Google Scholar] [CrossRef]
- Patel, J.P.; Konanur Srinivasa, N.K.; Gande, A.; Anusha, M.; Dar, H.; Baji, D.B. The Role of Biologics in Rheumatoid Arthritis: A Narrative Review. Cureus 2023, 15, e33293. [Google Scholar] [CrossRef]
- Ramadan, D.E.; Hariyani, N.; Indrawati, R.; Ridwan, R.D.; Diyatri, I. Cytokines and Chemokines in Periodontitis. Eur. J. Dent. 2020, 14, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Jin, H.M.; Kim, K.; Song, I.; Youn, B.U.; Matsuo, K.; Kim, N. The mechanism of osteoclast differentiation induced by IL-1. J. Immunol. 2009, 183, 1862–1870. [Google Scholar] [CrossRef]
- Fox, S.W.; Fuller, K.; Chambers, T.J. Activation of osteoclasts by interleukin-1: Divergent responsiveness in osteoclasts formed in vivo and in vitro. J. Cell Physiol. 2000, 184, 334–340. [Google Scholar] [CrossRef] [PubMed]
- Dipalma, G.; Inchingolo, A.D.; Fiore, A.; Balestriere, L.; Nardelli, P.; Casamassima, L.; Di Venere, D.; Palermo, A.; Inchingolo, F.; Inchingolo, A.M. The Differential Impact of Clear Aligners and Fixed Orthodontic Appliances on Periodontal Health: A Systematic Review. Children 2025, 12, 138. [Google Scholar] [CrossRef]
- Motyl, S.; Manfredini, D.; Oruba, Z.; Bugajska, J.; Sztefko, K.; Stos, W.; Osiewicz, M.; Loster, B.W.; Lobbezoo, F. Evaluation of interleukin-1 beta and the ratio of interleukin-1 beta to interleukin-1 receptor antagonist in gingival crevicular fluid during orthodontic canine retraction. Dent. Med. Probl. 2021, 58, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Soares Bonato, R.C.; Abel Mapengo, M.A.; de Azevedo-Silva, L.J.; Janson, G.; de Carvalho Sales-Peres, S.H. Tooth movement, orofacial pain, and leptin, interleukin-1beta, and tumor necrosis factor-alpha levels in obese adolescents. Angle Orthod. 2022, 92, 95–100. [Google Scholar] [CrossRef]
- Dinarello, C.A. Interleukin-1 and its biologically related cytokines. Adv. Immunol. 1989, 44, 153–205. [Google Scholar] [CrossRef]
- d’Apuzzo, F.; Cappabianca, S.; Ciavarella, D.; Monsurro, A.; Silvestrini-Biavati, A.; Perillo, L. Biomarkers of periodontal tissue remodeling during orthodontic tooth movement in mice and men: Overview and clinical relevance. Sci. World J. 2013, 2013, 105873. [Google Scholar] [CrossRef]
- Davidovitch, Z.; Nicolay, O.F.; Ngan, P.W.; Shanfeld, J.L. Neurotransmitters, cytokines, and the control of alveolar bone remodeling in orthodontics. Dent. Clin. N. Am. 1988, 32, 411–435. [Google Scholar] [CrossRef]
- Lee, K.J.; Park, Y.C.; Yu, H.S.; Choi, S.H.; Yoo, Y.J. Effects of continuous and interrupted orthodontic force on interleukin-1beta and prostaglandin E2 production in gingival crevicular fluid. Am. J. Orthod. Dentofac. Orthop. 2004, 125, 168–177. [Google Scholar] [CrossRef]
- Iwasaki, L.R.; Haack, J.E.; Nickel, J.C.; Reinhardt, R.A.; Petro, T.M. Human interleukin-1 beta and interleukin-1 receptor antagonist secretion and velocity of tooth movement. Arch. Oral Biol. 2001, 46, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Varella, A.M.; Revankar, A.V.; Patil, A.K. Low-level laser therapy increases interleukin-1beta in gingival crevicular fluid and enhances the rate of orthodontic tooth movement. Am. J. Orthod. Dentofac. Orthop. 2018, 154, 535–544.e5. [Google Scholar] [CrossRef]
- Bletsa, A.; Berggreen, E.; Brudvik, P. Interleukin-1alpha and tumor necrosis factor-alpha expression during the early phases of orthodontic tooth movement in rats. Eur. J. Oral Sci. 2006, 114, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, C.C.; Khoo, E.; Tran, J.; Chartres, I.; Liu, Y.; Thant, L.M.; Khabensky, I.; Gart, L.P.; Cisneros, G.; Alikhani, M. Cytokine expression and accelerated tooth movement. J. Dent. Res. 2010, 89, 1135–1141. [Google Scholar] [CrossRef]
- Kumar, B.D.; Singh, N.; Verma, S.K.; Singh, S.; Thakur, S. A Study to Evaluate Il1 And Il6 Gingival Crevicular Fluid Levels in Adolescents and Young Adults During the Early Phase of Orthodontic Tooth Movement. J. Pharm. Bioallied Sci. 2022, 14, S494–S497. [Google Scholar] [CrossRef]
- Reis, C.L.B.; de Souza Furtado, T.C.; Mendes, W.D.; Matsumoto, M.A.N.; Alves, S.Y.F.; Stuani, M.B.S.; Borsatto, M.C.; Corona, S.A.M. Photobiomodulation impacts the levels of inflammatory mediators during orthodontic tooth movement? A systematic review with meta-analysis. Lasers Med. Sci. 2022, 37, 771–787. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, A.V.B.; Marcantonio, C.C.; de Molon, R.S.; Leguizamon, N.D.P.; Silva, R.C.L.; Deschner, J.; Cerri, P.S.; Cirelli, J.A. Experimental models of orthodontic tooth movement and their effects on periodontal tissues remodelling. Arch. Oral Biol. 2021, 130, 105216. [Google Scholar] [CrossRef]
- Chen, L.; Hua, Y. Autophagy of periodontal ligament inhibits inflammation and reduces the decline of bone density during orthodontic tooth movement of mice. Arch. Oral Biol. 2021, 121, 104960. [Google Scholar] [CrossRef]
- de Mello-Neto, J.M.; Elangovan, G.; Ervolino, E.; Johnson, N.W.; Gustafsson, A.; da Silva Figueredo, C.M. Higher expression of Th1/Th2-related cytokines in the intestine of Wistar rats with ligature-induced periodontitis. J. Periodontal Res. 2023, 58, 588–595. [Google Scholar] [CrossRef]
- Sun, H.; Zhao, Y.; Wang, K.; Zhu, L.; Dong, J.; Zhao, J.; Wang, Y.; Li, H.; Sun, X.; Lu, Y. Low dose IL-2 suppress osteoclastogenesis in collagen-induced arthritis via JNK dependent pathway. Immun. Inflamm. Dis. 2020, 8, 727–735. [Google Scholar] [CrossRef]
- Wang, X.; Feng, C. The Association between IL2 Genotypes and Risk and Severity of Chronic Periodontitis in a Chinese Han Population: A Case-control Study. Immunol. Invest. 2022, 51, 924–930. [Google Scholar] [CrossRef] [PubMed]
- Basaran, G.; Ozer, T.; Kaya, F.A.; Hamamci, O. Interleukins 2, 6, and 8 levels in human gingival sulcus during orthodontic treatment. Am. J. Orthod. Dentofac. Orthop. 2006, 130, 7.e1–7.e6. [Google Scholar] [CrossRef]
- Pan, K.; Li, Q.; Guo, Z.; Li, Z. Healing action of Interleukin-4 (IL-4) in acute and chronic inflammatory conditions: Mechanisms and therapeutic strategies. Pharmacol. Ther. 2025, 265, 108760. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.A.; Hural, J. Functions of IL-4 and Control of Its Expression. Crit. Rev. Immunol. 2017, 37, 181–212. [Google Scholar] [CrossRef] [PubMed]
- Iwaszko, M.; Bialy, S.; Bogunia-Kubik, K. Significance of Interleukin (IL)-4 and IL-13 in Inflammatory Arthritis. Cells 2021, 10, 3000. [Google Scholar] [CrossRef]
- Lima Teixeira, J.F.; Henning, P.; Cintra Magalhaes, F.A.; Coletto-Nunes, G.; Floriano-Marcelino, T.; Westerlund, A.; Moverare-Skrtic, S.; Oliveira, G.; Lerner, U.H.; Souza, P.P.C. Osteoprotective effect by interleukin-4 (IL-4) on lipoprotein-induced periodontitis. Cytokine 2023, 172, 156399. [Google Scholar] [CrossRef]
- Zhou, P.; Zheng, T.; Zhao, B. Cytokine-mediated immunomodulation of osteoclastogenesis. Bone 2022, 164, 116540. [Google Scholar] [CrossRef]
- Kitaura, H.; Nagata, N.; Fujimura, Y.; Hotokezaka, H.; Tatamiya, M.; Nakao, N.; Yoshida, N.; Nakayama, K. Interleukin-4 directly inhibits tumor necrosis factor-alpha-mediated osteoclast formation in mouse bone marrow macrophages. Immunol. Lett. 2003, 88, 193–198. [Google Scholar] [CrossRef]
- Fujii, T.; Kitaura, H.; Kimura, K.; Hakami, Z.W.; Takano-Yamamoto, T. IL-4 inhibits TNF-alpha-mediated osteoclast formation by inhibition of RANKL expression in TNF-alpha-activated stromal cells and direct inhibition of TNF-alpha-activated osteoclast precursors via a T-cell-independent mechanism in vivo. Bone 2012, 51, 771–780. [Google Scholar] [CrossRef]
- Hakami, Z.; Kitaura, H.; Kimura, K.; Ishida, M.; Sugisawa, H.; Ida, H.; Jafari, S.; Takano-Yamamoto, T. Effect of interleukin-4 on orthodontic tooth movement and associated root resorption. Eur. J. Orthod. 2015, 37, 87–94. [Google Scholar] [CrossRef]
- Wu, M.; Liu, J. Inhibitory effect of exogenous IL-4 on orthodontic relapse in rats. Oral Dis. 2022, 28, 469–479. [Google Scholar] [CrossRef]
- Garcia-Lopez, S.; Villanueva, R.E.; Masso-Rojas, F.; Paez-Arenas, A.; Meikle, M.C. Micro-vibrations at 30 Hz on bone cells cultivated in vitro produce soluble factors for osteoclast inhibition and osteoblast activity. Arch. Oral Biol. 2020, 110, 104594. [Google Scholar] [CrossRef]
- Danz, J.C.; Kantarci, A.; Bornstein, M.M.; Katsaros, C.; Stavropoulos, A. Impact of Orthodontic Forces on Plasma Levels of Markers of Bone Turnover and Inflammation in a Rat Model of Buccal Expansion. Front. Physiol. 2021, 12, 637606. [Google Scholar] [CrossRef] [PubMed]
- Omata, Y.; Frech, M.; Saito, T.; Schett, G.; Zaiss, M.M.; Tanaka, S. Inflammatory Arthritis and Bone Metabolism Regulated by Type 2 Innate and Adaptive Immunity. Int. J. Mol. Sci. 2022, 23, 1104. [Google Scholar] [CrossRef] [PubMed]
- Andreev, D.; Kachler, K.; Liu, M.; Chen, Z.; Krishnacoumar, B.; Ringer, M.; Frey, S.; Kronke, G.; Voehringer, D.; Schett, G.; et al. Eosinophils preserve bone homeostasis by inhibiting excessive osteoclast formation and activity via eosinophil peroxidase. Nat. Commun. 2024, 15, 1067. [Google Scholar] [CrossRef] [PubMed]
- Grant, M.; Wilson, J.; Rock, P.; Chapple, I. Induction of cytokines, MMP9, TIMPs, RANKL and OPG during orthodontic tooth movement. Eur. J. Orthod. 2013, 35, 644–651. [Google Scholar] [CrossRef]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in inflammation, immunity, and disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef]
- Pandolfi, F.; Franza, L.; Carusi, V.; Altamura, S.; Andriollo, G.; Nucera, E. Interleukin-6 in Rheumatoid Arthritis. Int. J. Mol. Sci. 2020, 21, 5238. [Google Scholar] [CrossRef]
- Hirano, T. IL-6 in inflammation, autoimmunity and cancer. Int. Immunol. 2021, 33, 127–148. [Google Scholar] [CrossRef]
- Tyrrell, D.J.; Goldstein, D.R. Ageing and atherosclerosis: Vascular intrinsic and extrinsic factors and potential role of IL-6. Nat. Rev. Cardiol. 2021, 18, 58–68. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, F.; Hou, L. IL-6 and diabetic kidney disease. Front. Immunol. 2024, 15, 1465625. [Google Scholar] [CrossRef] [PubMed]
- Grebenciucova, E.; VanHaerents, S. Interleukin 6: At the interface of human health and disease. Front. Immunol. 2023, 14, 1255533. [Google Scholar] [CrossRef]
- Kang, S.; Narazaki, M.; Metwally, H.; Kishimoto, T. Historical overview of the interleukin-6 family cytokine. J. Exp. Med. 2020, 217, e20190347. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; He, C. TNF-alpha and IL-6: The Link between Immune and Bone System. Curr. Drug Targets 2020, 21, 213–227. [Google Scholar] [CrossRef] [PubMed]
- Udagawa, N.; Takahashi, N.; Katagiri, T.; Tamura, T.; Wada, S.; Findlay, D.M.; Martin, T.J.; Hirota, H.; Taga, T.; Kishimoto, T.; et al. Interleukin (IL)-6 induction of osteoclast differentiation depends on IL-6 receptors expressed on osteoblastic cells but not on osteoclast progenitors. J. Exp. Med. 1995, 182, 1461–1468. [Google Scholar] [CrossRef] [PubMed]
- Devlin, R.D.; Reddy, S.V.; Savino, R.; Ciliberto, G.; Roodman, G.D. IL-6 mediates the effects of IL-1 or TNF, but not PTHrP or 1,25(OH)2D3, on osteoclast-like cell formation in normal human bone marrow cultures. J. Bone Min. Res. 1998, 13, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.Y.; Wu, H.K.; Chen, Y.H.; Hsu, Y.P.; Cheng, M.T.; Yu, C.H.; Chen, S.K. Interleukin-6 transiently promotes proliferation of osteoclast precursors and stimulates the production of inflammatory mediators. Mol. Biol. Rep. 2022, 49, 3927–3937. [Google Scholar] [CrossRef]
- Jayaprakash, P.K.; Basavanna, J.M.; Grewal, H.; Modi, P.; Sapawat, P.; Bohara, P.D. Elevated levels of Interleukin (IL)-1beta, IL-6, tumor necrosis factor-alpha, epidermal growth factor, and beta2-microglobulin levels in gingival crevicular fluid during human Orthodontic tooth movement (OTM). J. Fam. Med. Prim. Care 2019, 8, 1602–1606. [Google Scholar] [CrossRef]
- Rath-Deschner, B.; Nogueira, A.V.B.; Beisel-Memmert, S.; Nokhbehsaim, M.; Eick, S.; Cirelli, J.A.; Deschner, J.; Jager, A.; Damanaki, A. Interaction of periodontitis and orthodontic tooth movement—An in vitro and in vivo study. Clin. Oral Investig. 2022, 26, 171–181. [Google Scholar] [CrossRef]
- Uzun, M.; Cesur, M.G.; Erdogan, O. Evaluation of the effects of obesity on orthodontic tooth movement. Korean J. Orthod. 2025, 55, 3–14. [Google Scholar] [CrossRef]
- Altindal, D.; Tunca, Y.; Tunca, M. Evaluation of IL-8 and IL-6 levels in gingival crevicular fluid of individuals undergoing clear aligner therapy. Angle Orthod. 2025, 95, 212–218. [Google Scholar] [CrossRef]
- Toyama, N.; Ono, T.; Ono, T.; Nakashima, T. The interleukin-6 signal regulates orthodontic tooth movement and pain. Biochem. Biophys. Res. Commun. 2023, 684, 149068. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.; Li, Z.; Liu, L.; Zhao, J.; Ge, W.; Zhang, K.; Zhou, Z.; Liu, Y. Static magnetic field-induced IL-6 secretion in periodontal ligament stem cells accelerates orthodontic tooth movement. Sci. Rep. 2024, 14, 9851. [Google Scholar] [CrossRef]
- Dwivedi, P.; Agrawal, A.; Gupta, S.C.; Chou, T. The Effect of Varied Time Interval and Micro-Current (Direct) on the Level of Biomarker (IL-6) and Rate of Tooth Movement: An Animal Study. Indian. J. Dent. Res. 2023, 34, 375–380. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.Y.; Li, S.Q.; Jia, X.; Wu, M.Y.; Song, B.; Li, J.Y.; Guo, Y.; Gao, R. Expression patterns of interleukin-6 and microRNA-146A during orthodontic relapse in a rat model. J. Physiol. Pharmacol. 2024, 75, 447–456. [Google Scholar]
- Winer, H.; Rodrigues, G.O.L.; Hixon, J.A.; Aiello, F.B.; Hsu, T.C.; Wachter, B.T.; Li, W.; Durum, S.K. IL-7: Comprehensive review. Cytokine 2022, 160, 156049. [Google Scholar] [CrossRef]
- Toraldo, G.; Roggia, C.; Qian, W.P.; Pacifici, R.; Weitzmann, M.N. IL-7 induces bone loss in vivo by induction of receptor activator of nuclear factor kappa B ligand and tumor necrosis factor alpha from T cells. Proc. Natl. Acad. Sci. USA 2003, 100, 125–130. [Google Scholar] [CrossRef]
- Xu, H.; Cai, L.; Li, Z.; Zhang, L.; Wang, G.; Xie, R.; Jiang, Y.; Yuan, Y.; Nie, H. Dual effect of IL-7/IL-7R signalling on the osteoimmunological system: A potential therapeutic target for rheumatoid arthritis. Immunology 2021, 164, 161–172. [Google Scholar] [CrossRef]
- Kim, S.J.; Chang, H.J.; Volin, M.V.; Umar, S.; Van Raemdonck, K.; Chevalier, A.; Palasiewicz, K.; Christman, J.W.; Volkov, S.; Arami, S.; et al. Macrophages are the primary effector cells in IL-7-induced arthritis. Cell Mol. Immunol. 2020, 17, 728–740. [Google Scholar] [CrossRef]
- Sato, T.; Watanabe, K.; Masuhara, M.; Hada, N.; Hakeda, Y. Production of IL-7 is increased in ovariectomized mice, but not RANKL mRNA expression by osteoblasts/stromal cells in bone, and IL-7 enhances generation of osteoclast precursors in vitro. J. Bone Min. Metab. 2007, 25, 19–27. [Google Scholar] [CrossRef]
- Chami, V.O.; Nunes, L.; Capelli Junior, J. Expression of cytokines in gingival crevicular fluid associated with tooth movement induced by aligners: A pilot study. Dent. Press. J. Orthod. 2018, 23, 41–46. [Google Scholar] [CrossRef]
- Matsushima, K.; Yang, D.; Oppenheim, J.J. Interleukin-8: An evolving chemokine. Cytokine 2022, 153, 155828. [Google Scholar] [CrossRef] [PubMed]
- Park, S.E.; Park, K.; Kim, E.; Kim, C.Y.; Hwang, S.M.; Lee, J.M.; Suh, J.Y.; Lee, Y.; Kim, M.O.; Kim, Y.G. CXCL5/CXCL8 induces neutrophilic inflammation in peri-implantitis. J. Periodontal Res. 2024, 59, 698–711. [Google Scholar] [CrossRef]
- Cambier, S.; Gouwy, M.; Proost, P. The chemokines CXCL8 and CXCL12: Molecular and functional properties, role in disease and efforts towards pharmacological intervention. Cell Mol. Immunol. 2023, 20, 217–251. [Google Scholar] [CrossRef] [PubMed]
- Fousek, K.; Horn, L.A.; Palena, C. Interleukin-8: A chemokine at the intersection of cancer plasticity, angiogenesis, and immune suppression. Pharmacol. Ther. 2021, 219, 107692. [Google Scholar] [CrossRef]
- Malhotra, H.; Garg, V.; Singh, G. Biomarker Approach Towards Rheumatoid Arthritis Treatment. Curr. Rheumatol. Rev. 2021, 17, 162–175. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chen, Z.; Lan, T.; Liang, P.; Tao, Q. Upregulation of interleukin-8 and activin A induces osteoclastogenesis in ameloblastoma. Int. J. Mol. Med. 2019, 43, 2329–2340. [Google Scholar] [CrossRef]
- Nassar, E.A.; Almasoud, N.N.; Al-Qurashi, M.S.; Alsulaiman, A.A.; Hassan, K.S. An Evaluation of Microbial Flora, Alkaline Phosphatase and IL-8 Levels in GCF of Orthodontic Patients with Self-Ligating and Conventional Brackets. Clin. Cosmet. Investig. Dent. 2021, 13, 343–352. [Google Scholar] [CrossRef]
- Asano, M.; Yamaguchi, M.; Nakajima, R.; Fujita, S.; Utsunomiya, T.; Yamamoto, H.; Kasai, K. IL-8 and MCP-1 induced by excessive orthodontic force mediates odontoclastogenesis in periodontal tissues. Oral Dis. 2011, 17, 489–498. [Google Scholar] [CrossRef]
- Yang, J.H.; Li, Z.C.; Kong, W.D.; Zhang, W.; Jia, Y.P.; Zhang, Y.L.; Liu, L.B.; Han, X.P. Effect of orthodontic force on inflammatory periodontal tissue remodeling and expression of IL-6 and IL-8 in rats. Asian Pac. J. Trop. Med. 2013, 6, 757–761. [Google Scholar] [CrossRef]
- Fernandes, M.R.U.; Suzuki, S.S.; Suzuki, H.; Martinez, E.F.; Garcez, A.S. Photobiomodulation increases intrusion tooth movement and modulates IL-6, IL-8 and IL-1beta expression during orthodontically bone remodeling. J. Biophotonics 2019, 12, e201800311. [Google Scholar] [CrossRef] [PubMed]
- Tuncer, B.B.; Ozmeric, N.; Tuncer, C.; Teoman, I.; Cakilci, B.; Yucel, A.; Alpar, R.; Balos, K. Levels of interleukin-8 during tooth movement. Angle Orthod. 2005, 75, 631–636. [Google Scholar]
- Saraiva, M.; Vieira, P.; O’Garra, A. Biology and therapeutic potential of interleukin-10. J. Exp. Med. 2020, 217, e20190418. [Google Scholar] [CrossRef]
- Nagata, K.; Nishiyama, C. IL-10 in Mast Cell-Mediated Immune Responses: Anti-Inflammatory and Proinflammatory Roles. Int. J. Mol. Sci. 2021, 22, 4972. [Google Scholar] [CrossRef] [PubMed]
- Rasquinha, M.T.; Sur, M.; Lasrado, N.; Reddy, J. IL-10 as a Th2 Cytokine: Differences Between Mice and Humans. J. Immunol. 2021, 207, 2205–2215. [Google Scholar] [CrossRef]
- Gao, X.; Ge, J.; Zhou, W.; Xu, L.; Geng, D. IL-10 inhibits osteoclast differentiation and osteolysis through MEG3/IRF8 pathway. Cell. Signal. 2022, 95, 110353. [Google Scholar] [CrossRef]
- Rios-Arce, N.D.; Dagenais, A.; Feenstra, D.; Coughlin, B.; Kang, H.J.; Mohr, S.; McCabe, L.R.; Parameswaran, N. Loss of interleukin-10 exacerbates early Type-1 diabetes-induced bone loss. J. Cell Physiol. 2020, 235, 2350–2365. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wan, Z.; Yang, L.; Song, S.; Fu, Z.; Tang, K.; Chen, L.; Song, Y. Exosomes derived from reparative M2-like macrophages prevent bone loss in murine periodontitis models via IL-10 mRNA. J. Nanobiotechnology 2022, 20, 110. [Google Scholar] [CrossRef]
- Zhu, Z.; Chen, G.; Yu, S.; Huang, X.; Lu, X.; Feng, G.; Yi, M.; Wang, J.; Liu, Y.; Chen, L. Circadian clock disruption stimulates bone loss via regulatory T cell-Mediated regulation of IL-10 expression. Int. Immunopharmacol. 2024, 139, 112589. [Google Scholar] [CrossRef]
- Zhang, Q.; Chen, B.; Yan, F.; Guo, J.; Zhu, X.; Ma, S.; Yang, W. Interleukin-10 inhibits bone resorption: A potential therapeutic strategy in periodontitis and other bone loss diseases. Biomed. Res. Int. 2014, 2014, 284836. [Google Scholar] [CrossRef]
- Tzaneti, A.; Athanasopoulou, E.; Fessatou, S.; Fotis, L. Chronic Nonbacterial Osteomyelitis in Inflammatory Bowel Disease. Life 2023, 13, 2347. [Google Scholar] [CrossRef] [PubMed]
- Nunes, L.; Quintanilha, L.; Perinetti, G.; Capelli, J.J. Effect of orthodontic force on expression levels of ten cytokines in gingival crevicular fluid. Arch. Oral Biol. 2017, 76, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Alnazeh, A.A.; Kamran, M.A.; Aseeri, Y.; Alrwuili, M.R.; Aljabab, M.A.; Baig, E.A.; Hameed, M.S. Levels of Inflammatory and Bone Metabolic Markers in the Gingival Crevicular Fluid of Individuals Undergoing Fixed Orthodontic Treatment in Comparison to Those Utilizing Invisalign. Medicina 2023, 59, 2107. [Google Scholar] [CrossRef]
- Salla, J.T.; Taddei, S.R.; Queiroz-Junior, C.M.; Andrade Junior, I.; Teixeira, M.M.; Silva, T.A. The effect of IL-1 receptor antagonist on orthodontic tooth movement in mice. Arch. Oral Biol. 2012, 57, 519–524. [Google Scholar] [CrossRef]
- Wang, Y.; Groeger, S.; Yong, J.; Ruf, S. Orthodontic Compression Enhances Macrophage M2 Polarization via Histone H3 Hyperacetylation. Int. J. Mol. Sci. 2023, 24, 3117. [Google Scholar] [CrossRef] [PubMed]
- Fung, K.Y.; Louis, C.; Metcalfe, R.D.; Kosasih, C.C.; Wicks, I.P.; Griffin, M.D.W.; Putoczki, T.L. Emerging roles for IL-11 in inflammatory diseases. Cytokine 2022, 149, 155750. [Google Scholar] [CrossRef]
- Cook, S.A. Understanding interleukin 11 as a disease gene and therapeutic target. Biochem. J. 2023, 480, 1987–2008. [Google Scholar] [CrossRef]
- Han, Y.; Gao, H.; Gan, X.; Liu, J.; Bao, C.; He, C. Roles of IL-11 in the regulation of bone metabolism. Front. Endocrinol. 2023, 14, 1290130. [Google Scholar] [CrossRef]
- Dong, B.; Zhu, J.; Chen, X.; Jiang, H.; Deng, Y.; Xu, L.; Wang, Y.; Li, S. The Emerging Role of Interleukin-(IL)-11/IL-11R in Bone Metabolism and Homeostasis: From Cytokine to Osteokine. Aging Dis. 2023, 14, 2113–2126. [Google Scholar] [CrossRef]
- Zou, R.; Huang, X.; Xu, P. The study of gp130/the inflammatory factors regulating osteoclast differentiation in rheumatoid arthritis. Biochem. Biophys. Rep. 2021, 26, 100934. [Google Scholar] [CrossRef]
- Tuerlings, M.; van Hoolwerff, M.; Houtman, E.; Suchiman, E.; Lakenberg, N.; Mei, H.; van der Linden, E.; Nelissen, R.; Ramos, Y.; Coutinho de Almeida, R.; et al. RNA Sequencing Reveals Interacting Key Determinants of Osteoarthritis Acting in Subchondral Bone and Articular Cartilage: Identification of IL11 and CHADL as Attractive Treatment Targets. Arthritis Rheumatol. 2021, 73, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zheng, W.; Liu, J.S.; Wang, J.; Yang, P.; Li, M.L.; Zhao, Z.H. Expression of osteoclastogenesis inducers in a tissue model of periodontal ligament under compression. J. Dent. Res. 2011, 90, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Xu, C.; Li, Y.; Hao, C.; Zheng, J.; Jin, X.; Yu, J.; Zhu, Y.; Guan, Z.; Yin, Q. The gingival crevicular fluid biomarkers with micropulse vibration device: A pilot study. Heliyon 2024, 10, e31982. [Google Scholar] [CrossRef]
- Ullrich, K.A.; Schulze, L.L.; Paap, E.M.; Muller, T.M.; Neurath, M.F.; Zundler, S. Immunology of IL-12: An update on functional activities and implications for disease. EXCLI J. 2020, 19, 1563–1589. [Google Scholar] [CrossRef]
- Nguyen, K.G.; Vrabel, M.R.; Mantooth, S.M.; Hopkins, J.J.; Wagner, E.S.; Gabaldon, T.A.; Zaharoff, D.A. Localized Interleukin-12 for Cancer Immunotherapy. Front. Immunol. 2020, 11, 575597. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, E.; Carson, W.E., 3rd. Analysis of potential biomarkers of response to IL-12 therapy. J. Leukoc. Biol. 2022, 112, 557–567. [Google Scholar] [CrossRef]
- Floss, D.M.; Moll, J.M.; Scheller, J. IL-12 and IL-23-Close Relatives with Structural Homologies but Distinct Immunological Functions. Cells 2020, 9, 2184. [Google Scholar] [CrossRef]
- Nagata, N.; Kitaura, H.; Yoshida, N.; Nakayama, K. Inhibition of RANKL-induced osteoclast formation in mouse bone marrow cells by IL-12: Involvement of IFN-gamma possibly induced from non-T cell population. Bone 2003, 33, 721–732. [Google Scholar] [CrossRef]
- Kitaura, H.; Nagata, N.; Fujimura, Y.; Hotokezaka, H.; Yoshida, N.; Nakayama, K. Effect of IL-12 on TNF-alpha-mediated osteoclast formation in bone marrow cells: Apoptosis mediated by Fas/Fas ligand interaction. J. Immunol. 2002, 169, 4732–4738. [Google Scholar] [CrossRef]
- Horwood, N.J.; Udagawa, N.; Elliott, J.; Grail, D.; Okamura, H.; Kurimoto, M.; Dunn, A.R.; Martin, T.; Gillespie, M.T. Interleukin 18 inhibits osteoclast formation via T cell production of granulocyte macrophage colony-stimulating factor. J. Clin. Invest. 1998, 101, 595–603. [Google Scholar] [CrossRef]
- Yoshimatsu, M.; Kitaura, H.; Fujimura, Y.; Kohara, H.; Morita, Y.; Eguchi, T.; Yoshida, N. Inhibitory effects of IL-12 on experimental tooth movement and root resorption in mice. Arch. Oral Biol. 2012, 57, 36–43. [Google Scholar] [CrossRef]
- Bernstein, Z.J.; Shenoy, A.; Chen, A.; Heller, N.M.; Spangler, J.B. Engineering the IL-4/IL-13 axis for targeted immune modulation. Immunol. Rev. 2023, 320, 29–57. [Google Scholar] [CrossRef] [PubMed]
- Tubau, C.; Puig, L. Therapeutic targeting of the IL-13 pathway in skin inflammation. Expert. Rev. Clin. Immunol. 2021, 17, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Roeb, E. Interleukin-13 (IL-13)-A Pleiotropic Cytokine Involved in Wound Healing and Fibrosis. Int. J. Mol. Sci. 2023, 24, 12884. [Google Scholar] [CrossRef]
- Palmqvist, P.; Lundberg, P.; Persson, E.; Johansson, A.; Lundgren, I.; Lie, A.; Conaway, H.H.; Lerner, U.H. Inhibition of hormone and cytokine-stimulated osteoclastogenesis and bone resorption by interleukin-4 and interleukin-13 is associated with increased osteoprotegerin and decreased RANKL and RANK in a STAT6-dependent pathway. J. Biol. Chem. 2006, 281, 2414–2429. [Google Scholar] [CrossRef]
- Chen, Z.; Cheng, Z.; Tang, Z.; Li, M.; Tang, X.; Zhang, H.; Sun, L. Interleukin-13 reduces bone erosion in rheumatoid arthritis by up-regulating osteoprotegerin expression in fibroblast-like synoviocytes: An in vitro and in vivo study. Clin. Exp. Rheumatol. 2023, 41, 2151–2161. [Google Scholar] [CrossRef] [PubMed]
- Cruikshank, W.W.; Kornfeld, H.; Center, D.M. Interleukin-16. J. Leukoc. Biol. 2000, 67, 757–766. [Google Scholar] [CrossRef]
- Chang, Y.; Hsiao, Y.M.; Hu, C.C.; Chang, C.H.; Li, C.Y.; Ueng, S.W.N.; Chen, M.F. Synovial Fluid Interleukin-16 Contributes to Osteoclast Activation and Bone Loss through the JNK/NFATc1 Signaling Cascade in Patients with Periprosthetic Joint Infection. Int. J. Mol. Sci. 2020, 21, 2904. [Google Scholar] [CrossRef]
- Luo, H.; Fang, S.; Liu, Q.; Dang, W.; Wang, Y. Comparison of interleukin expression in gingival crevicular fluid between patients with invisible orthodontics treat-ment and fixed orthodontics treatment. Hua Xi Kou Qiang Yi Xue Za Zhi 2022, 40, 293–296. [Google Scholar] [CrossRef]
- Huangfu, L.; Li, R.; Huang, Y.; Wang, S. The IL-17 family in diseases: From bench to bedside. Signal Transduct. Target. Ther. 2023, 8, 402. [Google Scholar] [CrossRef]
- Mills, K.H.G. IL-17 and IL-17-producing cells in protection versus pathology. Nat. Rev. Immunol. 2023, 23, 38–54. [Google Scholar] [CrossRef] [PubMed]
- McGeachy, M.J.; Cua, D.J.; Gaffen, S.L. The IL-17 Family of Cytokines in Health and Disease. Immunity 2019, 50, 892–906. [Google Scholar] [CrossRef]
- Adamopoulos, I.E.; Chao, C.C.; Geissler, R.; Laface, D.; Blumenschein, W.; Iwakura, Y.; McClanahan, T.; Bowman, E.P. Interleukin-17A upregulates receptor activator of NF-kappaB on osteoclast precursors. Arthritis Res. Ther. 2010, 12, R29. [Google Scholar] [CrossRef]
- Yago, T.; Nanke, Y.; Ichikawa, N.; Kobashigawa, T.; Mogi, M.; Kamatani, N.; Kotake, S. IL-17 induces osteoclastogenesis from human monocytes alone in the absence of osteoblasts, which is potently inhibited by anti-TNF-alpha antibody: A novel mechanism of osteoclastogenesis by IL-17. J. Cell Biochem. 2009, 108, 947–955. [Google Scholar] [CrossRef]
- Ueda, M.; Hikida, T.; Shimizu, M.; Kikuta, J.; Takagi, K.; Tsukada, M.; Yamaguchi, M. Involvement of interleukins-17 and -34 in exacerbated orthodontic root resorption by jiggling force during rat experimental tooth movement. J. World Fed. Orthod. 2020, 9, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, N.; Yamaguchi, M.; Nakajima, R.; Utsunomiya, T.; Yamamoto, H.; Kasai, K. T-helper 17 cells mediate the osteo/odontoclastogenesis induced by excessive orthodontic forces. Oral Dis. 2012, 18, 375–388. [Google Scholar] [CrossRef]
- Sagar, S.; Ramani, P.; Moses, S.; Gheena, S.; Selvaraj, J. Correlation of salivary cytokine IL-17A and 1,25 dihydroxycholecalciferol in patients undergoing orthodontic treatment. Odontology 2024, 112, 966–975. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.; Yang, L.; Zheng, W.; Zhang, B. Matrix metalloproteinases and Th17 cytokines in the gingival crevicular fluid during orthodontic tooth movement. Eur. J. Paediatr. Dent. 2021, 22, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.; Yang, L.; Zheng, W.; Zhang, B. Th17 Cytokines and its Correlation with Receptor Activator of Nuclear Factor kappa B Ligand During Orthodontic Tooth Movement. Iran. J. Immunol. 2020, 17, 137–143. [Google Scholar] [CrossRef]
- Landy, E.; Carol, H.; Ring, A.; Canna, S. Biological and clinical roles of IL-18 in inflammatory diseases. Nat. Rev. Rheumatol. 2024, 20, 33–47. [Google Scholar] [CrossRef]
- Vecchie, A.; Bonaventura, A.; Toldo, S.; Dagna, L.; Dinarello, C.A.; Abbate, A. IL-18 and infections: Is there a role for targeted therapies? J. Cell Physiol. 2021, 236, 1638–1657. [Google Scholar] [CrossRef] [PubMed]
- Ihim, S.A.; Abubakar, S.D.; Zian, Z.; Sasaki, T.; Saffarioun, M.; Maleknia, S.; Azizi, G. Interleukin-18 cytokine in immunity, inflammation, and autoimmunity: Biological role in induction, regulation, and treatment. Front. Immunol. 2022, 13, 919973. [Google Scholar] [CrossRef]
- Novick, D. IL-18 and IL-18BP: A Unique Dyad in Health and Disease. Int. J. Mol. Sci. 2024, 25, 13505. [Google Scholar] [CrossRef]
- Rex, D.A.B.; Agarwal, N.; Prasad, T.S.K.; Kandasamy, R.K.; Subbannayya, Y.; Pinto, S.M. A comprehensive pathway map of IL-18-mediated signalling. J. Cell Commun. Signal 2020, 14, 257–266. [Google Scholar] [CrossRef]
- Kitaura, H.; Tatamiya, M.; Nagata, N.; Fujimura, Y.; Eguchi, T.; Yoshida, N.; Nakayama, K. IL-18 induces apoptosis of adherent bone marrow cells in TNF-alpha mediated osteoclast formation in synergy with IL-12. Immunol. Lett. 2006, 107, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Horwood, N.J.; Elliott, J.; Martin, T.J.; Gillespie, M.T. IL-12 alone and in synergy with IL-18 inhibits osteoclast formation in vitro. J. Immunol. 2001, 166, 4915–4921. [Google Scholar] [CrossRef] [PubMed]
- Morita, Y.; Kitaura, H.; Yoshimatsu, M.; Fujimura, Y.; Kohara, H.; Eguchi, T.; Yoshida, N. IL-18 inhibits TNF-alpha-induced osteoclastogenesis possibly via a T cell-independent mechanism in synergy with IL-12 in vivo. Calcif. Tissue Int. 2010, 86, 242–248. [Google Scholar] [CrossRef]
- Nogueira, A.V.; de Molon, R.S.; Nokhbehsaim, M.; Deschner, J.; Cirelli, J.A. Contribution of biomechanical forces to inflammation-induced bone resorption. J. Clin. Periodontol. 2017, 44, 31–41. [Google Scholar] [CrossRef]
- Blumberg, H.; Conklin, D.; Xu, W.F.; Grossmann, A.; Brender, T.; Carollo, S.; Eagan, M.; Foster, D.; Haldeman, B.A.; Hammond, A.; et al. Interleukin 20: Discovery, receptor identification, and role in epidermal function. Cell 2001, 104, 9–19. [Google Scholar] [CrossRef]
- Meng, B.; Wu, D.; Cheng, Y.; Huang, P.; Liu, Y.; Gan, L.; Liu, C.; Cao, Y. Interleukin-20 differentially regulates bone mesenchymal stem cell activities in RANKL-induced osteoclastogenesis through the OPG/RANKL/RANK axis and the NF-kappaB, MAPK and AKT signalling pathways. Scand. J. Immunol. 2020, 91, e12874. [Google Scholar] [CrossRef]
- Yemenoglu, H.; Senkal, R.; Kose, O.; Yilmaz, A.; Mataraci Karakas, S.; Akyildiz, K. The effect of interleukin-20 on periodontal tissue destruction in individuals with periodontitis. J. Periodontal Res. 2024, 59, 480–490. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ai, Y.; Sun, X.; Meng, B.; Chen, X.; Wu, D.; Gan, L.; Yang, B.; Fu, C.; Wu, Y.; et al. Interleukin-20 Acts as a Promotor of Osteoclastogenesis and Orthodontic Tooth Movement. Stem Cells Int. 2021, 2021, 5539962. [Google Scholar] [CrossRef]
- Meng, B.; Yang, B.; Qu, Y.; Liu, Y.; Wu, D.; Fu, C.; He, Y.; Chen, X.; Liu, C.; Kou, X.; et al. Dual Role of Interleukin-20 in Different Stages of Osteoclast Differentiation and Its Osteoimmune Regulation during Alveolar Bone Remodeling. Int. J. Mol. Sci. 2023, 24, 3810. [Google Scholar] [CrossRef]
- Sewell, G.W.; Kaser, A. Interleukin-23 in the Pathogenesis of Inflammatory Bowel Disease and Implications for Therapeutic Intervention. J. Crohns Colitis 2022, 16, ii3–ii19. [Google Scholar] [CrossRef] [PubMed]
- Furuya, H.; Nguyen, C.T.; Gu, R.; Hsieh, S.L.; Maverakis, E.; Adamopoulos, I.E. Interleukin-23 Regulates Inflammatory Osteoclastogenesis via Activation of CLEC5A(+) Osteoclast Precursors. Arthritis Rheumatol. 2023, 75, 1477–1489. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Pang, D.D.; Dai, S.M. Expression Profile of Osteoclasts Following the Stimulation With Interleukin-23 in Mice. Arch. Rheumatol. 2020, 35, 533–544. [Google Scholar] [CrossRef]
- Chen, S.Y.; Tsai, T.C.; Li, Y.T.; Ding, Y.C.; Wang, C.T.; Hsieh, J.L.; Wu, C.L.; Wu, P.T.; Shiau, A.L. Interleukin-23 Mediates Osteoclastogenesis in Collagen-Induced Arthritis by Modulating MicroRNA-223. Int. J. Mol. Sci. 2022, 23, 9718. [Google Scholar] [CrossRef]
- Xu, W.D.; Wang, D.C.; Zhao, M.; Huang, A.F. An updated advancement of bifunctional IL-27 in inflammatory autoimmune diseases. Front. Immunol. 2024, 15, 1366377. [Google Scholar] [CrossRef]
- Yoshida, H.; Hunter, C.A. The immunobiology of interleukin-27. Annu. Rev. Immunol. 2015, 33, 417–443. [Google Scholar] [CrossRef]
- Li, X.; Luo, W.; Hu, J.; Chen, Y.; Yu, T.; Yang, J.; Dong, S.; Tian, X.; Sun, L. Interleukin-27 prevents LPS-induced inflammatory osteolysis by inhibiting osteoclast formation and function. Am. J. Transl. Res. 2019, 11, 1154–1169. [Google Scholar]
- Morita, Y.; Saito, M.; Rangel-Moreno, J.; Franchini, A.M.; Owen, J.R.; Martinez, J.C.; Daiss, J.L.; de Mesy Bentley, K.L.; Kates, S.L.; Schwarz, E.M.; et al. Systemic IL-27 administration prevents abscess formation and osteolysis via local neutrophil recruitment and activation. Bone Res. 2022, 10, 56. [Google Scholar] [CrossRef] [PubMed]
- Kamiya, S.; Okumura, M.; Chiba, Y.; Fukawa, T.; Nakamura, C.; Nimura, N.; Mizuguchi, J.; Wada, S.; Yoshimoto, T. IL-27 suppresses RANKL expression in CD4+ T cells in part through STAT3. Immunol. Lett. 2011, 138, 47–53. [Google Scholar] [CrossRef]
- Dwyer, G.K.; D’Cruz, L.M.; Turnquist, H.R. Emerging Functions of IL-33 in Homeostasis and Immunity. Annu. Rev. Immunol. 2022, 40, 15–43. [Google Scholar] [CrossRef]
- Alarcon-Sanchez, M.A.; Romero-Castro, N.S.; Reyes-Fernandez, S.; Sanchez-Tecolapa, E.U.; Heboyan, A. Expression of IL-33 in subjects with periodontitis: A systematic review and meta-analysis. Eur. J. Med. Res. 2024, 29, 440. [Google Scholar] [CrossRef] [PubMed]
- Kiyomiya, H.; Ariyoshi, W.; Okinaga, T.; Kaneuji, T.; Mitsugi, S.; Sakurai, T.; Habu, M.; Yoshioka, I.; Tominaga, K.; Nishihara, T. IL-33 inhibits RANKL-induced osteoclast formation through the regulation of Blimp-1 and IRF-8 expression. Biochem. Biophys. Res. Commun. 2015, 460, 320–326. [Google Scholar] [CrossRef]
- Ohori, F.; Kitaura, H.; Ogawa, S.; Shen, W.R.; Qi, J.; Noguchi, T.; Marahleh, A.; Nara, Y.; Pramusita, A.; Mizoguchi, I. IL-33 Inhibits TNF-alpha-Induced Osteoclastogenesis and Bone Resorption. Int. J. Mol. Sci. 2020, 21, 1130. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Feng, J.; Li, B.; Bai, D.; Xu, H. Inhibition of osteoclastogenesis by interleukin-33 administration in the periodontal ligament under mechanical loading. J. Periodontal Res. 2022, 57, 1003–1013. [Google Scholar] [CrossRef]
- Dong, X.; Feng, J.; Wen, J.; Bai, D.; Xu, H. Effect of interleukin-33 on cementoblast-mediated cementum repair during orthodontic tooth movement. Arch. Oral Biol. 2020, 112, 104663. [Google Scholar] [CrossRef]
- Si, Y.; Zhang, J.; Bao, S.; Wise, S.G.; Wang, Y.; Zhang, Y.; Tang, Y. IL-32 and IL-34 in hepatocellular carcinoma. Front. Med. 2022, 9, 1051113. [Google Scholar] [CrossRef]
- Lelios, I.; Cansever, D.; Utz, S.G.; Mildenberger, W.; Stifter, S.A.; Greter, M. Emerging roles of IL-34 in health and disease. J. Exp. Med. 2020, 217, e20190290. [Google Scholar] [CrossRef]
- Duarte, C.; Yamada, C.; Ngala, B.; Garcia, C.; Akkaoui, J.; Birsa, M.; Ho, A.; Nusbaum, A.; AlQallaf, H.; John, V.; et al. Effects of IL-34 and anti-IL-34 neutralizing mAb on alveolar bone loss in a ligature-induced model of periodontitis. Mol. Oral Microbiol. 2024, 39, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Van Raemdonck, K.; Umar, S.; Palasiewicz, K.; Volin, M.V.; Elshabrawy, H.A.; Romay, B.; Tetali, C.; Ahmed, A.; Amin, M.A.; Zomorrodi, R.K.; et al. Interleukin-34 Reprograms Glycolytic and Osteoclastic Rheumatoid Arthritis Macrophages via Syndecan 1 and Macrophage Colony-Stimulating Factor Receptor. Arthritis Rheumatol. 2021, 73, 2003–2014. [Google Scholar] [CrossRef]
- Ng, C.T.; Fong, L.Y.; Abdullah, M.N.H. Interferon-gamma (IFN-gamma): Reviewing its mechanisms and signaling pathways on the regulation of endothelial barrier function. Cytokine 2023, 166, 156208. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Wu, M.; Liu, Z. Dysregulation in IFN-gamma signaling and response: The barricade to tumor immunotherapy. Front. Immunol. 2023, 14, 1190333. [Google Scholar] [CrossRef]
- Casanova, J.L.; MacMicking, J.D.; Nathan, C.F. Interferon-gamma and infectious diseases: Lessons and prospects. Science 2024, 384, eadl2016. [Google Scholar] [CrossRef] [PubMed]
- Mezouar, S.; Mege, J.L. Changing the paradigm of IFN-gamma at the interface between innate and adaptive immunity: Macrophage-derived IFN-gamma. J. Leukoc. Biol. 2020, 108, 419–426. [Google Scholar] [CrossRef]
- Tang, M.; Tian, L.; Luo, G.; Yu, X. Interferon-Gamma-Mediated Osteoimmunology. Front. Immunol. 2018, 9, 1508. [Google Scholar] [CrossRef]
- Tan, J.; Dai, A.; Pan, L.; Zhang, L.; Wang, Z.; Ke, T.; Sun, W.; Wu, Y.; Ding, P.H.; Chen, L. Inflamm-Aging-Related Cytokines of IL-17 and IFN-gamma Accelerate Osteoclastogenesis and Periodontal Destruction. J. Immunol. Res. 2021, 2021, 9919024. [Google Scholar] [CrossRef]
- Biros, E.; Malabu, U.H.; Vangaveti, V.N.; Birosova, E.; Moran, C.S. The IFN-gamma/miniTrpRS signaling axis: An insight into the pathophysiology of osteoporosis and therapeutic potential. Cytokine Growth Factor Rev. 2022, 64, 7–11. [Google Scholar] [CrossRef]
- Kohara, H.; Kitaura, H.; Fujimura, Y.; Yoshimatsu, M.; Morita, Y.; Eguchi, T.; Masuyama, R.; Yoshida, N. IFN-gamma directly inhibits TNF-alpha-induced osteoclastogenesis in vitro and in vivo and induces apoptosis mediated by Fas/Fas ligand interactions. Immunol. Lett. 2011, 137, 53–61. [Google Scholar] [CrossRef]
- Kohara, H.; Kitaura, H.; Yoshimatsu, M.; Fujimura, Y.; Morita, Y.; Eguchi, T.; Yoshida, N. Inhibitory effect of interferon-gamma on experimental tooth movement in mice. J. Interferon Cytokine Res. 2012, 32, 426–431. [Google Scholar] [CrossRef] [PubMed]
- Xin, T.Y.; Zhu, Z.L.; Chen, L.J.; Han, B.; Yang, R.L.; Shi, J. Profile of Inflammatory Cytokines in Gingival Crevicular Fluid and Plasma in Patients With Grade C Periodontitis During Orthodontic Treatment: A Longitudinal Case Series Report. Orthod. Craniofacial Res. 2025, 28, 324–335. [Google Scholar] [CrossRef] [PubMed]
- Mermut, S.; Bengi, A.O.; Akin, E.; Kurkcu, M.; Karacay, S. Effects of interferon-gamma on bone remodeling during experimental tooth movement. Angle Orthod. 2007, 77, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Liu, F.; Kou, X.; Liu, D.; Yang, R.; Wang, X.; Song, Y.; He, D.; Gan, Y.; Zhou, Y. T Cells Are Required for Orthodontic Tooth Movement. J. Dent. Res. 2015, 94, 1463–1470. [Google Scholar] [CrossRef]
- An, S.; Zhang, Y.; Chen, Q.; Xiong, B.; Hao, J.; Zheng, Y.; Zhou, X.; Wang, J. Effect of systemic delivery of Substance P on experimental tooth movement in rats. Am. J. Orthod. Dentofac. Orthop. 2019, 155, 642–649. [Google Scholar] [CrossRef]
- Jang, D.I.; Lee, A.H.; Shin, H.Y.; Song, H.R.; Park, J.H.; Kang, T.B.; Lee, S.R.; Yang, S.H. The Role of Tumor Necrosis Factor Alpha (TNF-alpha) in Autoimmune Disease and Current TNF-alpha Inhibitors in Therapeutics. Int. J. Mol. Sci. 2021, 22, 2719. [Google Scholar] [CrossRef]
- van Loo, G.; Bertrand, M.J.M. Death by TNF: A road to inflammation. Nat. Rev. Immunol. 2023, 23, 289–303. [Google Scholar] [CrossRef]
- Berkhout, L.C.; I’Ami, M.J.; Kruithof, S.; Vogelzang, E.H.; Hooijberg, F.; Hart, M.H.L.; Bentlage, A.E.H.; Thomas, D.; Vermeire, S.; Vidarsson, G.; et al. Formation and clearance of TNF-TNF inhibitor complexes during TNF inhibitor treatment. Br. J. Pharmacol. 2024, 181, 1165–1181. [Google Scholar] [CrossRef]
- Idriss, H.T.; Naismith, J.H. TNF alpha and the TNF receptor superfamily: Structure-function relationship(s). Microsc. Res. Tech. 2000, 50, 184–195. [Google Scholar] [CrossRef]
- Huyghe, J.; Priem, D.; Bertrand, M.J.M. Cell death checkpoints in the TNF pathway. Trends Immunol. 2023, 44, 628–643. [Google Scholar] [CrossRef]
- Veerasubramanian, P.K.; Wynn, T.A.; Quan, J.; Karlsson, F.J. Targeting TNF/TNFR superfamilies in immune-mediated inflammatory diseases. J. Exp. Med. 2024, 221, e20240806. [Google Scholar] [CrossRef] [PubMed]
- Horiuchi, T.; Mitoma, H.; Harashima, S.; Tsukamoto, H.; Shimoda, T. Transmembrane TNF-alpha: Structure, function and interaction with anti-TNF agents. Rheumatology 2010, 49, 1215–1228. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Getting, S.J.; Locke, I.C. Regulation of TNF-Induced Osteoclast Differentiation. Cells 2021, 11, 132. [Google Scholar] [CrossRef] [PubMed]
- Guberna, L.; Nyssen, O.P.; Chaparro, M.; Gisbert, J.P. Frequency and Effectiveness of Empirical Anti-TNF Dose Intensification in Inflammatory Bowel Disease: Systematic Review with Meta-Analysis. J. Clin. Med. 2021, 10, 2132. [Google Scholar] [CrossRef]
- Yao, Q.; He, L.; Bao, C.; Yan, X.; Ao, J. The role of TNF-alpha in osteoporosis, bone repair and inflammatory bone diseases: A review. Tissue Cell 2024, 89, 102422. [Google Scholar] [CrossRef]
- Neurath, N.; Kesting, M. Cytokines in gingivitis and periodontitis: From pathogenesis to therapeutic targets. Front. Immunol. 2024, 15, 1435054. [Google Scholar] [CrossRef]
- Siegmund, D.; Wajant, H. TNF and TNF receptors as therapeutic targets for rheumatic diseases and beyond. Nat. Rev. Rheumatol. 2023, 19, 576–591. [Google Scholar] [CrossRef]
- Kitaura, H.; Marahleh, A.; Ohori, F.; Noguchi, T.; Nara, Y.; Pramusita, A.; Kinjo, R.; Ma, J.; Kanou, K.; Mizoguchi, I. Role of the Interaction of Tumor Necrosis Factor-alpha and Tumor Necrosis Factor Receptors 1 and 2 in Bone-Related Cells. Int. J. Mol. Sci. 2022, 23, 1481. [Google Scholar] [CrossRef]
- Abu-Amer, Y.; Erdmann, J.; Alexopoulou, L.; Kollias, G.; Ross, F.P.; Teitelbaum, S.L. Tumor necrosis factor receptors types 1 and 2 differentially regulate osteoclastogenesis. J. Biol. Chem. 2000, 275, 27307–27310. [Google Scholar] [CrossRef]
- Lowney, J.J.; Norton, L.A.; Shafer, D.M.; Rossomando, E.F. Orthodontic forces increase tumor necrosis factor alpha in the human gingival sulcus. Am. J. Orthod. Dentofac. Orthop. 1995, 108, 519–524. [Google Scholar] [CrossRef]
- Uematsu, S.; Mogi, M.; Deguchi, T. Interleukin (IL)-1 beta, IL-6, tumor necrosis factor-alpha, epidermal growth factor, and beta 2-microglobulin levels are elevated in gingival crevicular fluid during human orthodontic tooth movement. J. Dent. Res. 1996, 75, 562–567. [Google Scholar] [CrossRef]
- Li, H.; Chen, Y.; Sun, W. Effects of Kangfuxinye on NF-kappaB and Inflammatory Cytokines in Gingival Crevicular Fluid of Patients with Orthodontic Gingivitis Caused by Orthodontic Treatment. Cell Mol. Biol. 2023, 69, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, P.; Kharbanda, O.P.; Monga, N.; Miglani, R.; Kapila, S. Effect of orthodontic forces on cytokine and receptor levels in gingival crevicular fluid: A systematic review. Prog. Orthod. 2014, 15, 65. [Google Scholar] [CrossRef]
- Debnath, P.; Bangi, S.L.; Hussain, M.F.; Rafiq, S.; Tousifulla, S.; Abdu, M.; Gupta, S. The Evaluation of Gingival Crevicular Fluid Biomarkers as Predictors of Gingival Enlargement in Patients Undergoing Fixed Orthodontic Treatment: A Prospective Study. Cureus 2024, 16, e74281. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Mi, D.; Wu, H.; Fu, Y.; Liu, L.; Chen, X.; Dong, Y.; Zhang, W. Changes in Matrix Metalloproteinase-8, Interleukin-6 and Tumor Necrosis Factor-A in Gingival Crevicular Fluid during Rapid Maxillary Expansion in Adolescent Patients. Iran. J. Public. Health 2021, 50, 1944–1952. [Google Scholar] [CrossRef]
- Basaran, G.; Ozer, T.; Kaya, F.A.; Kaplan, A.; Hamamci, O. Interleukine-1beta and tumor necrosis factor-alpha levels in the human gingival sulcus during orthodontic treatment. Angle Orthod. 2006, 76, 830–836. [Google Scholar] [PubMed]
- Ogasawara, T.; Yoshimine, Y.; Kiyoshima, T.; Kobayashi, I.; Matsuo, K.; Akamine, A.; Sakai, H. In situ expression of RANKL, RANK, osteoprotegerin and cytokines in osteoclasts of rat periodontal tissue. J. Periodontal Res. 2004, 39, 42–49. [Google Scholar] [CrossRef]
- Yoshimatsu, M.; Shibata, Y.; Kitaura, H.; Chang, X.; Moriishi, T.; Hashimoto, F.; Yoshida, N.; Yamaguchi, A. Experimental model of tooth movement by orthodontic force in mice and its application to tumor necrosis factor receptor-deficient mice. J. Bone Min. Metab. 2006, 24, 20–27. [Google Scholar] [CrossRef]
- Andrade, I., Jr.; Silva, T.A.; Silva, G.A.; Teixeira, A.L.; Teixeira, M.M. The role of tumor necrosis factor receptor type 1 in orthodontic tooth movement. J. Dent. Res. 2007, 86, 1089–1094. [Google Scholar] [CrossRef]
- Kitaura, H.; Yoshimatsu, M.; Fujimura, Y.; Eguchi, T.; Kohara, H.; Yamaguchi, A.; Yoshida, N. An anti-c-Fms antibody inhibits orthodontic tooth movement. J. Dent. Res. 2008, 87, 396–400. [Google Scholar] [CrossRef]
- Ma, J.; Kitaura, H.; Ogawa, S.; Ohori, F.; Noguchi, T.; Marahleh, A.; Nara, Y.; Pramusita, A.; Kinjo, R.; Kanou, K.; et al. Docosahexaenoic acid inhibits TNF-alpha-induced osteoclast formation and orthodontic tooth movement through GPR120. Front. Immunol. 2022, 13, 929690. [Google Scholar] [CrossRef]
- Marahleh, A.; Kitaura, H.; Ohori, F.; Kishikawa, A.; Ogawa, S.; Shen, W.R.; Qi, J.; Noguchi, T.; Nara, Y.; Mizoguchi, I. TNF-alpha Directly Enhances Osteocyte RANKL Expression and Promotes Osteoclast Formation. Front. Immunol. 2019, 10, 2925. [Google Scholar] [CrossRef] [PubMed]
- Marahleh, A.; Kitaura, H.; Ohori, F.; Noguchi, T.; Mizoguchi, I. The osteocyte and its osteoclastogenic potential. Front. Endocrinol. 2023, 14, 1121727. [Google Scholar] [CrossRef]
- Noguchi, T.; Kitaura, H.; Ogawa, S.; Qi, J.; Shen, W.R.; Ohori, F.; Marahleh, A.; Nara, Y.; Pramusita, A.; Mizoguchi, I. TNF-alpha stimulates the expression of RANK during orthodontic tooth movement. Arch. Oral Biol. 2020, 117, 104796. [Google Scholar] [CrossRef] [PubMed]
- Marahleh, A.; Kitaura, H.; Ohori, F.; Noguchi, T.; Nara, Y.; Pramusita, A.; Kinjo, R.; Ma, J.; Kanou, K.; Mizoguchi, I. Effect of TNF-alpha on osteocyte RANKL expression during orthodontic tooth movement. J. Dent. Sci. 2021, 16, 1191–1197. [Google Scholar] [CrossRef]
- Kanou, K.; Kitaura, H.; Noguchi, T.; Ohori, F.; Marahleh, A.; Kinjo, R.; Ma, J.; Ren, J.; Ogasawara, K.; Mizoguchi, I. Effect of age on orthodontic tooth movement in mice. J. Dent. Sci. 2024, 19, 828–836. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, T.; Kitaura, H.; Marahleh, A.; Ohori, F.; Nara, Y.; Pramusita, A.; Kinjo, R.; Ma, J.; Kanou, K.; Mizoguchi, I. Tumor necrosis factor-alpha enhances the expression of vascular endothelial growth factor in a mouse orthodontic tooth movement model. J. Dent. Sci. 2022, 17, 415–420. [Google Scholar] [CrossRef]
- Ohori, F.; Kitaura, H.; Marahleh, A.; Ma, J.; Miura, M.; Ren, J.; Narita, K.; Fan, Z.; Lin, A.; Mizoguchi, I. Osteocyte necroptosis drives osteoclastogenesis and alveolar bone resorption during orthodontic tooth movement. Sci. Rep. 2025, 15, 19413. [Google Scholar] [CrossRef]
- Sehgal, A.; Irvine, K.M.; Hume, D.A. Functions of macrophage colony-stimulating factor (CSF1) in development, homeostasis, and tissue repair. Semin. Immunol. 2021, 54, 101509. [Google Scholar] [CrossRef]
- Mun, S.H.; Park, P.S.U.; Park-Min, K.H. The M-CSF receptor in osteoclasts and beyond. Exp. Mol. Med. 2020, 52, 1239–1254. [Google Scholar] [CrossRef]
- Begg, S.K.; Radley, J.M.; Pollard, J.W.; Chisholm, O.T.; Stanley, E.R.; Bertoncello, I. Delayed hematopoietic development in osteopetrotic (op/op) mice. J. Exp. Med. 1993, 177, 237–242. [Google Scholar] [CrossRef]
- Udagawa, N.; Koide, M.; Nakamura, M.; Nakamichi, Y.; Yamashita, T.; Uehara, S.; Kobayashi, Y.; Furuya, Y.; Yasuda, H.; Fukuda, C.; et al. Osteoclast differentiation by RANKL and OPG signaling pathways. J. Bone Min. Metab. 2021, 39, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Kitaura, H.; Zhou, P.; Kim, H.J.; Novack, D.V.; Ross, F.P.; Teitelbaum, S.L. M-CSF mediates TNF-induced inflammatory osteolysis. J. Clin. Investig. 2005, 115, 3418–3427. [Google Scholar] [CrossRef]
- Nara, Y.; Kitaura, H.; Ogawa, S.; Shen, W.R.; Qi, J.; Ohori, F.; Noguchi, T.; Marahleh, A.; Pramusita, A.; Kinjo, R.; et al. Anti-c-fms Antibody Prevents Osteoclast Formation and Bone Resorption in Co-Culture of Osteoblasts and Osteoclast Precursors In Vitro and in Ovariectomized Mice. Int. J. Mol. Sci. 2020, 21, 6120. [Google Scholar] [CrossRef]
- Brooks, P.J.; Heckler, A.F.; Wei, K.; Gong, S.G. M-CSF accelerates orthodontic tooth movement by targeting preosteoclasts in mice. Angle Orthod. 2011, 81, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Qi, J.; Kitaura, H.; Shen, W.R.; Kishikawa, A.; Ogawa, S.; Ohori, F.; Noguchi, T.; Marahleh, A.; Nara, Y.; Mizoguchi, I. Establishment of an orthodontic retention mouse model and the effect of anti-c-Fms antibody on orthodontic relapse. PLoS ONE 2019, 14, e0214260. [Google Scholar] [CrossRef] [PubMed]
- Massague, J.; Sheppard, D. TGF-beta signaling in health and disease. Cell 2023, 186, 4007–4037. [Google Scholar] [CrossRef]
- Larson, C.; Oronsky, B.; Carter, C.A.; Oronsky, A.; Knox, S.J.; Sher, D.; Reid, T.R. TGF-beta: A master immune regulator. Expert. Opin. Ther. Targets 2020, 24, 427–438. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, X.; Wan, Y.Y. Intricacies of TGF-beta signaling in Treg and Th17 cell biology. Cell Mol. Immunol. 2023, 20, 1002–1022. [Google Scholar] [CrossRef]
- Takai, H.; Kanematsu, M.; Yano, K.; Tsuda, E.; Higashio, K.; Ikeda, K.; Watanabe, K.; Yamada, Y. Transforming growth factor-beta stimulates the production of osteoprotegerin/osteoclastogenesis inhibitory factor by bone marrow stromal cells. J. Biol. Chem. 1998, 273, 27091–27096. [Google Scholar] [CrossRef]
- Karst, M.; Gorny, G.; Galvin, R.J.; Oursler, M.J. Roles of stromal cell RANKL, OPG, and M-CSF expression in biphasic TGF-beta regulation of osteoclast differentiation. J. Cell Physiol. 2004, 200, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Weitzmann, M.N.; Cenci, S.; Haug, J.; Brown, C.; DiPersio, J.; Pacifici, R. B lymphocytes inhibit human osteoclastogenesis by secretion of TGFbeta. J. Cell Biochem. 2000, 78, 318–324. [Google Scholar] [CrossRef]
- Quinn, J.M.; Itoh, K.; Udagawa, N.; Hausler, K.; Yasuda, H.; Shima, N.; Mizuno, A.; Higashio, K.; Takahashi, N.; Suda, T.; et al. Transforming growth factor beta affects osteoclast differentiation via direct and indirect actions. J. Bone Min. Res. 2001, 16, 1787–1794. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Macdonald, B.R.; Hon, J.; Winkler, M.E.; Derynck, R.; Mundy, G.R.; Roodman, G.D. Recombinant Human Transforming Growth-Factor-Alpha-Stimulates the Formation of Osteoclast-Like Cells in Long-Term Human Marrow Cultures. J. Clin. Investig. 1986, 78, 894–898. [Google Scholar] [CrossRef]
- Heubel, B.; Nohe, A. The Role of BMP Signaling in Osteoclast Regulation. J. Dev. Biol. 2021, 9, 24. [Google Scholar] [CrossRef]
- Uematsu, S.; Mogi, M.; Deguchi, T. Increase of transforming growth factor-beta 1 in gingival crevicular fluid during human orthodontic tooth movement. Arch. Oral Biol. 1996, 41, 1091–1095. [Google Scholar] [CrossRef]
- Chen, Y.; Mei, L.; Qian, Y.; Zhou, X.; Zhao, Z.; Zheng, W.; Li, Y. Integrated bioinformatic analysis of protein landscape in gingival crevicular fluid unveils sequential bioprocess in orthodontic tooth movement. Prog. Orthod. 2024, 25, 37. [Google Scholar] [CrossRef]
- Sasaki, K.; Takeshita, N.; Fukunaga, T.; Seiryu, M.; Sakamoto, M.; Oyanagi, T.; Maeda, T.; Takano-Yamamoto, T. Vibration accelerates orthodontic tooth movement by inducing osteoclastogenesis via transforming growth factor-beta signalling in osteocytes. Eur. J. Orthod. 2022, 44, 698–704. [Google Scholar] [CrossRef] [PubMed]
- Akbulut, S.; Yagci, A.; Yay, A.H.; Yalcin, B. Experimental investigation of effects of platelet-rich plasma on early phases of orthodontic tooth movement. Am. J. Orthod. Dentofac. Orthop. 2019, 155, 71–79. [Google Scholar] [CrossRef]
- Jeon, H.H.; Teixeira, H.; Tsai, A.N. Mechanistic Insight into Orthodontic Tooth Movement Based on Animal Studies: A Critical Review. J. Clin. Med. 2021, 10, 1733. [Google Scholar] [CrossRef]
- Gao, Y.; Zhang, M.; Tian, X.; Wang, M.; Zhang, F. Experimental animal study on BMP-3 expression in periodontal tissues in the process of orthodontic tooth movement. Exp. Ther. Med. 2019, 17, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.L.; Fan, J.D.; Wang, A.A.; Jin, X.; Zhang, Z.B.; Hu, X.T.; Liu, L.; Zhao, Y.T.; Li, Y.F. Effect of local application of bone morphogenetic protein-2 on experimental tooth movement and biological remodeling in rats. Front. Physiol. 2023, 14, 1111857. [Google Scholar] [CrossRef]
- Yasuda, H.; Shima, N.; Nakagawa, N.; Yamaguchi, K.; Kinosaki, M.; Mochizuki, S.; Tomoyasu, A.; Yano, K.; Goto, M.; Murakami, A.; et al. Osteoclast differentiation factor is a ligand for osteoprotegerin/osteoclastogenesis-inhibitory factor and is identical to TRANCE/RANKL. Proc. Natl. Acad. Sci. USA 1998, 95, 3597–3602. [Google Scholar] [CrossRef]
- Kim, J.M.; Lin, C.; Stavre, Z.; Greenblatt, M.B.; Shim, J.H. Osteoblast-Osteoclast Communication and Bone Homeostasis. Cells 2020, 9, 2073. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, T.; Hayashi, M.; Takayanagi, H. New insights into osteoclastogenic signaling mechanisms. Trends Endocrinol. Metab. 2012, 23, 582–590. [Google Scholar] [CrossRef]
- Ono, T.; Hayashi, M.; Sasaki, F.; Nakashima, T. RANKL biology: Bone metabolism, the immune system, and beyond. Inflamm. Regen. 2020, 40, 2. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, T.; Hayashi, M.; Fukunaga, T.; Kurata, K.; Oh-Hora, M.; Feng, J.Q.; Bonewald, L.F.; Kodama, T.; Wutz, A.; Wagner, E.F.; et al. Evidence for osteocyte regulation of bone homeostasis through RANKL expression. Nat. Med. 2011, 17, 1231–1234. [Google Scholar] [CrossRef]
- Kramer, I.; Halleux, C.; Keller, H.; Pegurri, M.; Gooi, J.H.; Weber, P.B.; Feng, J.Q.; Bonewald, L.F.; Kneissel, M. Osteocyte Wnt/beta-catenin signaling is required for normal bone homeostasis. Mol. Cell Biol. 2010, 30, 3071–3085. [Google Scholar] [CrossRef]
- Chang, J.H.; Chen, P.J.; Arul, M.R.; Dutra, E.H.; Nanda, R.; Kumbar, S.G.; Yadav, S. Injectable RANKL sustained release formulations to accelerate orthodontic tooth movement. Eur. J. Orthod. 2020, 42, 317–325. [Google Scholar] [CrossRef]
- Li, C.; Chung, C.J.; Hwang, C.J.; Lee, K.J. Local injection of RANKL facilitates tooth movement and alveolar bone remodelling. Oral Dis. 2019, 25, 550–560. [Google Scholar] [CrossRef]
- Nishijima, Y.; Yamaguchi, M.; Kojima, T.; Aihara, N.; Nakajima, R.; Kasai, K. Levels of RANKL and OPG in gingival crevicular fluid during orthodontic tooth movement and effect of compression force on releases from periodontal ligament cells in vitro. Orthod. Craniofacial Res. 2006, 9, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Miyazawa, K.; Tabuchi, M.; Yabumoto, T.; Kadota, M.; Yoshizako, M.; Yamane, C.; Kawatani, M.; Osada, H.; Maeda, H.; et al. Effect of Reveromycin A on experimental tooth movement in OPG-/- mice. J. Dent. Res. 2012, 91, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Shoji-Matsunaga, A.; Ono, T.; Hayashi, M.; Takayanagi, H.; Moriyama, K.; Nakashima, T. Osteocyte regulation of orthodontic force-mediated tooth movement via RANKL expression. Sci. Rep. 2017, 7, 8753. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Mi, S.C.; Li, X.Y.; Liu, Y.T.; Han, N.N.; Xu, J.J.; Liu, Y.; Li, S.; Guo, L.J. MicroRNA-155 targets SOCS1 to inhibit osteoclast differentiation during orthodontic tooth movement. BMC Oral Health 2023, 23, 63–70. [Google Scholar] [CrossRef]
- Cultrera, G.; Lo Giudice, A.; Santonocito, S.; Ronsivalle, V.; Conforte, C.; Reitano, G.; Leonardi, R.; Isola, G. MicroRNA Modulation during Orthodontic Tooth Movement: A Promising Strategy for Novel Diagnostic and Personalized Therapeutic Interventions. Int. J. Mol. Sci. 2022, 23, 15501. [Google Scholar] [CrossRef]
- Han, Y.N.; Yang, Q.L.; Huang, Y.P.; Li, X.B.; Zhu, Y.Y.; Jia, L.F.; Zheng, Y.F.; Li, W.R. Mechanical force inhibited hPDLSCs proliferation with the downregulation of via DNA methylation. Oral Dis. 2021, 27, 1268–1282. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Cytokines | Effect on Osteoclast Formation | Source (Osteoclast) | Detection of Cytokines During OTM | Effect on OTM | Source (OTM) |
|---|---|---|---|---|---|
| IL-1β | + | [6,19,20,21,22,23] | detect | + | [25,26,28,29,30,31,32,33,34,35,36,37,38] |
| IL-2 | + or − | [39] (+), [40] (−) | detect | [42] | |
| IL-4 | − | [46,47,48,49] | − | [50,51,53] | |
| IL-5 | − | [55] | undetectable | [56] | |
| IL-6 | + | [21,64,65,66,67] | detect | + | [35,68,69,70,71,72,73,74,75] |
| IL-7 | + | [77,78,79,80] | no different | [81] | |
| IL-8 | + | [87] | detect | [36,71,88,89,90,91,92] | |
| IL-10 | − | [96,97,98,99,100,101] | detect | [102,103,104] | |
| IL-11 | + | [108,109,110,111] | detect | [112,113] | |
| IL-12 | − | [118,119,120] | − | [121] | |
| IL-13 | − | [125,126] | detect | [102] | |
| IL-16 | + | [128] | no different | [129] | |
| IL-17 | + | [133,134] | detect | [135,136,137,138,139] | |
| IL-18 | − | [120,145,146,147] | detect | [34,113,148] | |
| IL-20 | + | [150,151] | detect | + | [152,153] |
| IL-23 | + | [157] | detect | [139] | |
| IL-27 | − | [160] | detect | [139] | |
| IL-33 | − | [165,166] | detect | − | [167,168] |
| IL-34 | − | [171,172] | detect | [135] | |
| IFN-γ | − | [177,178,179,180] | detect | − | [181,182,183,184,185] |
| TNF-α | + | [191,193,194,198,199] | detect | + | [102,200,201,202,203,204,205,206,207,208,209,210,212,213,214,215,216,217,218] |
| M-CSF | + | [219,220,221,222,223] | + | [210,224,225] | |
| TGFs | + or − | [223] (+), [230,231,232,234,235] (+) | detect | + or − | [13,236,237,238,239,240,241,242] |
| RANKL | + | [221,242,243,244,245,246] | detect | + | [53,215,248,249,250,252] |
| OPG | − | [221,247] | detect | − | [53,250,251] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kitaura, H.; Ohori, F.; Marahleh, A.; Ma, J.; Lin, A.; Fan, Z.; Narita, K.; Murakami, K.; Kanetaka, H. The Role of Cytokines in Orthodontic Tooth Movement. Int. J. Mol. Sci. 2025, 26, 6688. https://doi.org/10.3390/ijms26146688
Kitaura H, Ohori F, Marahleh A, Ma J, Lin A, Fan Z, Narita K, Murakami K, Kanetaka H. The Role of Cytokines in Orthodontic Tooth Movement. International Journal of Molecular Sciences. 2025; 26(14):6688. https://doi.org/10.3390/ijms26146688
Chicago/Turabian StyleKitaura, Hideki, Fumitoshi Ohori, Aseel Marahleh, Jinghan Ma, Angyi Lin, Ziqiu Fan, Kohei Narita, Kou Murakami, and Hiroyasu Kanetaka. 2025. "The Role of Cytokines in Orthodontic Tooth Movement" International Journal of Molecular Sciences 26, no. 14: 6688. https://doi.org/10.3390/ijms26146688
APA StyleKitaura, H., Ohori, F., Marahleh, A., Ma, J., Lin, A., Fan, Z., Narita, K., Murakami, K., & Kanetaka, H. (2025). The Role of Cytokines in Orthodontic Tooth Movement. International Journal of Molecular Sciences, 26(14), 6688. https://doi.org/10.3390/ijms26146688