
Lipodystrophy in HIV: Evolving Challenges and Unresolved Questions

 ,
,  , and
, and {kind=link}
{kind=link}
Abstract
1. Introduction
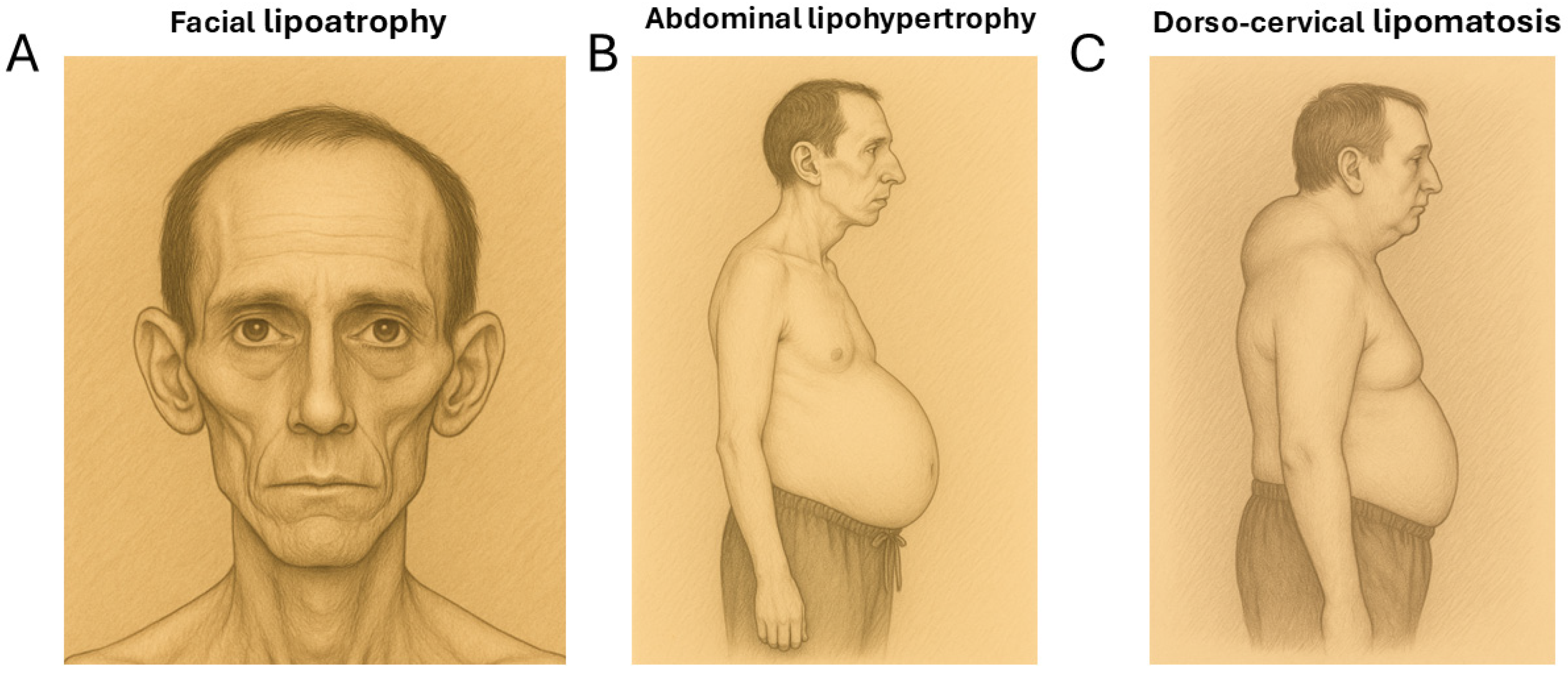
2. Causal Agents of Lipodystrophy in PLWH
2.1. Lipoatrophy
2.2. Lipohypertrophy
2.3. Lipomatosis
2.4. Systemic Alterations in HIV-Associated Lipodystrophy
3. Mechanisms Underlying HIV-Associated Lipodystrophy
3.1. Antiretroviral Treatment-Driven Effects
3.2. HIV Infection-Related Effects
4. Adipose Tissue Changes Under Modern Antiretroviral Regimens in PLWH
5. Conclusions: Current Challenges for Management of Healthy Adipose Tissue in PLWH
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AIDS | acquired immunodeficiency syndrome |
| cART | combined antiretroviral therapy |
| GLP-1 | glucagon-like peptide-1 |
| HIV | human immunodeficiency virus |
| INSTI | integrase strand transfer inhibitor |
| MASLD | metabolic dysfunction-associated steatotic liver disease |
| MASH | metabolic dysfunction-associated steatohepatitis |
| MC4R | melanocortin-4 receptor |
| mtDNA | mitochondrial DNA |
| NRTI | nucleotide analog reverse transcriptase inhibitor |
| NNRTI | non-nucleoside analog reverse transcriptase inhibitor |
| PCNA | proliferating cell nuclear antigen |
| PI | protease inhibitor |
| PLWH | people living with HIV |
| PPARγ | peroxisome proliferator-activated-γ |
| TAF | tenofovir alafenamide |
| TDF | tenofovir difumarate |
| UCP1 | uncoupling protein-1 |
References
- Giralt, M.; Domingo, P.; Villarroya, F. Adipose tissue biology and HIV-infection. Best Pract. Res. Clin. Endocrinol. Metab. 2011, 25, 487–499. [Google Scholar] [CrossRef] [PubMed]
- Koethe, J.R.; Lagathu, C.; Lake, J.E.; Domingo, P.; Calmy, A.; Falutz, J.; Brown, T.T.; Capeau, J. HIV and antiretroviral therapy-related fat alterations. Nat. Rev. Dis. Primers 2020, 6, 48. [Google Scholar] [CrossRef]
- Ramirez Bustamante, C.E.; Agarwal, N.; Cox, A.R.; Hartig, S.M.; Lake, J.E.; Balasubramanyam, A. Adipose Tissue Dysfunction and Energy Balance Paradigms in People Living with HIV. Endocr. Rev. 2024, 45, 190–209. [Google Scholar] [CrossRef]
- Capeau, J.; Lagathu, C.; Béréziat, V.; Fève, B. Recent data on adipose tissue, insulin resistance, diabetes and dyslipidaemia in antiretroviral therapy controlled HIV-infected persons. Curr. Opin. HIV AIDS 2021, 16, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Tilhet, L.; Rouanet, M.; Henry, A.S.; Pop, A.; Claudic, Y.; Saraoui, W.; Nachaoui, H.; Hu, W. Current status and perspectives in the treatment of facial lipoatrophy in HIV-positive patients in 2024. Ann. Chir. Plast. Esthet. 2025, 70, 192–205. [Google Scholar] [CrossRef]
- Domingo, P.; Estrada, V.; López-Aldeguer, J.; Villaroya, F.; Martínez, E. Fat redistribution syndromes associated with HIV-1 infection and combination antiretroviral therapy. AIDS Rev. 2012, 14, 112–123. [Google Scholar] [PubMed]
- Carr, A.; Law, M. HIV Lipodystrophy Case Definition Study Group. An objective lipodystrophy severity grading scale derived from the lipodystrophy case definition score. J. Acquir. Immune Defic. Syndr. 2003, 33, 571–576. [Google Scholar] [CrossRef]
- Mallon, P.W.; Miller, J.; Cooper, D.A.; Carr, A. Prospective evaluation of the effects of antiretroviral therapy on body composition in HIV-1-infected men starting therapy. AIDS 2003, 17, 971–979. [Google Scholar] [CrossRef]
- Martin, A.; Mallon, P.W. Therapeutic approaches to combating lipoatrophy: Do they work? J. Antimicrob. Chemother. 2005, 55, 612–615. [Google Scholar] [CrossRef]
- Glidden, D.V.; Mulligan, K.; McMahan, V.; Anderson, P.L.; Guanira, J.; Chariyalertsak, S.; Buchbinder, S.P.; Bekker, L.G.; Schechter, M.; Grinsztejn, B.; et al. Metabolic Effects of Preexposure Prophylaxis with Coformulated Tenofovir Disoproxil Fumarate and Emtricitabine. Clin. Infect. Dis. 2018, 67, 411–419. [Google Scholar] [CrossRef]
- Martínez, E.; Ribera, E.; Clotet, B.; Estrada, V.; Sanz, J.; Berenguer, J.; Rubio, R.; Pulido, F.; Larrousse, M.; Curran, A.; et al. Switching from zidovudine/lamivudine to tenofovir/emtricitabine improves fat distribution as measured by fat mass ratio. HIV Med. 2015, 16, 370–374. [Google Scholar] [CrossRef]
- Capeau, J.; Lagathu, C.; Béréziat, V. Recent data on the role of antiretroviral therapy in weight gain and obesity in persons living with HIV. Curr. Opin. HIV AIDS 2024, 19, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, E.; Navarro, A.; Curto, J.; Medina, P.; Rozas, N.; Barrera, G.; Saumoy, M.; Tiraboschi, J.M.; Gomez, C.; Podzamczer, D. Long-term fat redistribution in ARV-naïve HIV+ patients initiating a non-thymidine containing regimen in clinical practice. J. Int. AIDS Soc. 2014, 17 (Suppl. 3), 19553. [Google Scholar] [CrossRef]
- Grant, P.M.; Kitch, D.; McComsey, G.A.; Collier, A.C.; Bartali, B.; Koletar, S.L.; Erlandson, K.M.; Lake, J.E.; Yin, M.T.; Melbourne, K.; et al. Long-term body composition changes in antiretroviral-treated HIV-infected individuals. AIDS 2016, 30, 2805–2813. [Google Scholar] [CrossRef] [PubMed]
- Lo, J.C.; Mulligan, K.; Tai, V.W.; Algren, H.; Schambelan, M. “Buffalo hump” in men with HIV-1 infection. Lancet 1998, 351, 867–870. [Google Scholar] [CrossRef]
- Guaraldi, G.; Orlando, G.; Squillace, N.; Roverato, A.; De Fazio, D.; Vandelli, M.; Nardini, G.; Beghetto, B.; De Paola, M.; Esposito, R.; et al. Prevalence of and risk factors for pubic lipoma development in HIV-infected persons. J. Acquir. Immune Defic. Syndr. 2007, 45, 72–76. [Google Scholar] [CrossRef]
- Cereijo, R.; Gallego-Escuredo, J.M.; Moure, R.; Villarroya, J.; Domingo, J.C.; Fontdevila, J.; Martínez, E.; Gutiérrez, M.M.; Mateo, M.G.; Giralt, M.; et al. The Molecular Signature of HIV-1-Associated Lipomatosis Reveals Differential Involvement of Brown and Beige/Brite Adipocyte Cell Lineages. PLoS ONE 2015, 10, e0136571. [Google Scholar] [CrossRef] [PubMed]
- Guaraldi, G.; De Fazio, D.; Orlando, G.; Murri, R.; Wu, A.; Guaraldi, P.; Esposito, R. Facial lipohypertrophy in HIV-infected subjects who underwent autologous fat tissue transplantation. Clin. Infect. Dis. 2005, 40, e13–e15. [Google Scholar] [CrossRef]
- Guallar, J.P.; Gallego-Escuredo, J.M.; Domingo, J.C.; Alegre, M.; Fontdevila, J.; Martínez, E.; Hammond, E.L.; Domingo, P.; Giralt, M.; Villarroya, F. Differential gene expression indicates that ‘buffalo hump’ is a distinct adipose tissue disturbance in HIV-1-associated lipodystrophy. AIDS 2008, 22, 575–584. [Google Scholar] [CrossRef]
- Béréziat, V.; Cervera, P.; Le Dour, C.; Verpont, M.C.; Dumont, S.; Vantyghem, M.C.; Capeau, J.; Vigouroux, C.; Lipodystrophy Study Group. LMNA mutations induce a non-inflammatory fibrosis and a brown fat-like dystrophy of enlarged cervical adipose tissue. Am. J. Pathol. 2011, 179, 2443–2453. [Google Scholar] [CrossRef]
- Bourgeois, C.; Gorwood, J.; Olivo, A.; Le Pelletier, L.; Capeau, J.; Lambotte, O.; Béréziat, V.; Lagathu, C. Contribution of Adipose Tissue to the Chronic Immune Activation and Inflammation Associated with HIV Infection and Its Treatment. Front. Immunol. 2021, 12, 670566. [Google Scholar] [CrossRef] [PubMed]
- Ghaben, A.L.; Scherer, P.E. Adipogenesis and metabolic health. Nat. Rev. Mol. Cell. Biol. 2019, 20, 242–258. [Google Scholar] [CrossRef] [PubMed]
- Kadowaki, T.; Yamauchi, T.; Kubota, N.; Hara, K.; Ueki, K.; Tobe, K. Adiponectin and adiponectin receptors in insulin resistance, diabetes, and the metabolic syndrome. J. Clin. Investig. 2006, 116, 1784–1792. [Google Scholar] [CrossRef] [PubMed]
- Tong, Q.; Sankal, J.L.; Hadigan, C.M.; Tan, G.; Rosenberg, E.S.; Kanki, P.J.; Grinspoon, S.K.; Hotamisligil, G.S. Regulation of adiponectin in human immunodeficiency virus-infected patients: Relationship to body composition and metabolic in-dices. J. Clin. Endocrinol. Metab. 2003, 88, 1559–1564. [Google Scholar] [CrossRef]
- Lagathu, C.; Bastard, J.P.; Auclair, M.; Maachi, M.; Kornprobst, M.; Capeau, J.; Caron, M. Antiretroviral drugs with adverse effects on adipocyte lipid metabolism and survival alter the expression and secretion of proinflammatory cytokines and adiponectin in vitro. Antivir. Ther. 2004, 9, 911–920. [Google Scholar] [CrossRef]
- Lagathu, C.; Eustace, B.; Prot, M.; Frantz, D.; Gu, Y.; Bastard, J.P.; Maachi, M.; Azoulay, S.; Briggs, M.; Caron, M.; et al. Some HIV antiretrovirals increase oxidative stress and alter chemokine, cytokine or adiponectin production in human adipocytes and macrophages. Antivir. Ther. 2007, 12, 489–500. [Google Scholar] [CrossRef]
- Jones, S.P.; Janneh, O.; Back, D.J.; Pirmohamed, M. Altered adipokine response in murine 3T3-F442A adipocytes treated with protease inhibitors and nucleoside reverse transcriptase inhibitors. Antivir. Ther. 2005, 10, 207–213. [Google Scholar] [CrossRef]
- Gallego-Escuredo, J.M.; Del Mar Gutierrez, M.; Diaz-Delfin, J.; Domingo, J.C.; Mateo, M.G.; Domingo, P.; Giralt, M.; Villarroya, F. Differential effects of efavirenz and lopinavir/ritonavir on human adipocyte differentiation, gene expression and release of adipokines and pro-inflammatory cytokines. Curr. HIV Res. 2010, 8, 545–553. [Google Scholar] [CrossRef]
- Tiliscan, C.; Aramă, V.; Mihăilescu, R.; Munteanu, D.I.; Streinu-Cercel, A.; Ion, D.A.; Rădulescu, M.A.; Popescu, C.; Lobodan, A.E.; Negru, A.R.; et al. Leptin expression in HIV-infected patients during antiretroviral therapy. Germs 2015, 5, 92–98. [Google Scholar] [CrossRef]
- Giralt, M.; Díaz-Delfín, J.; Gallego-Escuredo, J.M.; Villarroya, J.; Domingo, P.; Villarroya, F. Lipotoxicity on the basis of metabolic syndrome and lipodystrophy in HIV-1-infected patients under antiretroviral treatment. Curr. Pharm. Des. 2010, 16, 3371–3378. [Google Scholar] [CrossRef]
- Villarroya, F.; Domingo, P.; Giralt, M. Drug-induced lipotoxicity: Lipodystrophy associated with HIV-1 infection and antiretroviral treatment. Biochim. Biophys. Acta 2010, 1801, 392–399. [Google Scholar] [CrossRef]
- Torriani, M.; Thomas, B.J.; Barlow, R.B.; Librizzi, J.; Dolan, S.; Grinspoon, S. Increased intramyocellular lipid accumulation in HIV-infected women with fat redistribution. J. Appl. Physiol. 2006, 100, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Sutinen, J.; Häkkinen, A.M.; Westerbacka, J.; Seppälä-Lindroos, A.; Vehkavaara, S.; Halavaara, J.; Järvinen, A.; Ristola, M.; Yki-Järvinen, H. Increased fat accumulation in the liver in HIV-infected patients with antiretroviral therapy-associated lipodystrophy. AIDS 2002, 16, 2183–2193. [Google Scholar] [CrossRef] [PubMed]
- Neilan, T.G.; Nguyen, K.L.; Zaha, V.G.; Chew, K.W.; Morrison, L.; Ntusi, N.A.B.; Toribio, M.; Awadalla, M.; Drobni, Z.D.; Nelson, M.D.; et al. Myocardial Steatosis Among Antiretroviral Therapy-Treated People with Human Immunodeficiency Virus Participating in the REPRIEVE Trial. J. Infect. Dis. 2020, 222 (Suppl. 1), S63–S69. [Google Scholar] [CrossRef]
- Tuddenham, S.A.; Koay, W.L.A.; Zhao, N.; White, J.R.; Ghanem, K.G.; Sears, C.L.; HIV Microbiome Re-analysis Consortium. The Impact of Human Immunodeficiency Virus Infection on Gut Microbiota α-Diversity: An Individual-level Meta-analysis. Clin. Infect. Dis. 2020, 70, 615–627. [Google Scholar] [CrossRef] [PubMed]
- Brenchley, J.M.; Price, D.A.; Schacker, T.W.; Asher, T.E.; Silvestri, G.; Rao, S.; Kazzaz, Z.; Bornstein, E.; Lambotte, O.; Altmann, D.; et al. Microbial translocation is a cause of systemic immune activation in chronic HIV infection. Nat. Med. 2006, 12, 1365–1371. [Google Scholar] [CrossRef]
- Gogokhia, L.; Taur, Y.; Juluru, K.; Yagan, N.; Zhu, Y.S.; Pamer, E.; Glesby, M.J. Intestinal Dysbiosis and Markers of Systemic Inflammation in Viscerally and Generally Obese Persons Living with HIV. J. Acquir. Immune Defic. Syndr. 2020, 83, 81–89. [Google Scholar] [CrossRef]
- Dillon, S.M.; Kibbie, J.; Lee, E.J.; Guo, K.; Santiago, M.L.; Austin, G.L.; Gianella, S.; Landay, A.L.; Donovan, A.M.; Frank, D.N.; et al. Low abundance of colonic butyrate-producing bacteria in HIV infection is associated with microbial translocation and immune activation. AIDS 2017, 31, 511–521. [Google Scholar] [CrossRef]
- Lloreta, J.; Domingo, P.; Pujol, R.M.; Arroyo, J.A.; Baixeras, N.; Matias-Guiu, X.; Gilaberte, M.; Sambeat, M.A.; Serrano, S. Ultra-structural features of highly active antiretroviral therapy-associated partial lipodystrophy. Virchows Arch. 2002, 441, 599–604. [Google Scholar] [CrossRef]
- Jan, V.; Cervera, P.; Maachi, M.; Baudrimont, M.; Kim, M.; Vidal, H.; Girard, P.M.; Levan, P.; Rozenbaum, W.; Lombès, A.; et al. Altered fat differentiation and adipocytokine expression are interrelated and linked to morphological changes and insulin resistance in HIV-1-infected lipodystrophic patients. Antivir. Ther. 2004, 9, 555–564. [Google Scholar] [CrossRef] [PubMed]
- Guaraldi, G.; Luzi, K.; Bellistrì, G.M.; Zona, S.; Domingues da Silva, A.R.; Bai, F.; Garlassi, E.; Marchetti, G.; Capeau, J.; Monforte, A.D. CD8 T-cell activation is associated with lipodystrophy and visceral fat accumulation in antiretroviral therapy-treated virologically suppressed HIV-infected patients. J. Acquir. Immune Defic. Syndr. 2013, 64, 360–366. [Google Scholar] [CrossRef]
- Giralt, M.; Villarroya, F.; Araujo-Vilar, D. Lipodystrophies. In Encyclopedia of Endocrine Diseases, 2nd ed.; Huhyaniemi, I., Martini, L., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 482–495. [Google Scholar]
- Villarroya, F.; Domingo, P.; Giralt, M. Lipodystrophy associated with highly active anti-retroviral therapy for HIV infection: The adipocyte as a target of anti-retroviral-induced mitochondrial toxicity. Trends Pharmacol. Sci. 2005, 26, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Caron, M.; Auclair, M.; Lagathu, C.; Lombès, A.; Walker, U.A.; Kornprobst, M.; Capeau, J. The HIV-1 nucleoside reverse transcriptase inhibitors stavudine and zidovudine alter adipocyte functions in vitro. AIDS 2004, 18, 2127–2136. [Google Scholar] [CrossRef]
- Stankov, M.V.; Lücke, T.; Das, A.M.; Schmidt, R.E.; Behrens, G.M. Mitochondrial DNA depletion and respiratory chain activity in primary human subcutaneous adipocytes treated with nucleoside analogue reverse transcriptase inhibitors. Antimicrob. Agents Chemother. 2010, 54, 280–287. [Google Scholar] [CrossRef]
- De Luca, A.; Nasi, M.; Di Giambenedetto, S.; Cozzi-Lepri, A.; Pinti, M.; Marzocchetti, A.; Mussini, C.; Fabbiani, M.; Bracciale, L.; Cauda, R.; et al. Mitochondrial DNA haplogroups and incidence of lipodystrophy in HIV-infected patients on long-term antiretroviral therapy. J. Acquir. Immune Defic. Syndr. 2012, 59, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Gallego-Escuredo, J.M.; Villarroya, J.; Domingo, P.; Targarona, E.M.; Alegre, M.; Domingo, J.C.; Villarroya, F.; Giralt, M. Differentially altered molecular signature of visceral adipose tissue in HIV-1-associated lipodystrophy. J. Acquir. Immune Defic. Syndr. 2013, 64, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Kovsan, J.; Osnis, A.; Maissel, A.; Mazor, L.; Tarnovscki, T.; Hollander, L.; Ovadia, S.; Meier, B.; Klein, J.; Bashan, N.; et al. Depot-specific adipocyte cell lines reveal differential drug-induced responses of white adipocytes--relevance for partial lipodystrophy. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E315–E322. [Google Scholar] [CrossRef]
- Villarroya, F.; Domingo, P.; Giralt, M. Lipodystrophy in HIV 1-infected patients: Lessons for obesity research. Int. J. Obes. 2007, 31, 1763–1776. [Google Scholar] [CrossRef]
- Srinivasa, S.; Grinspoon, S.K. Metabolic and body composition effects of newer antiretrovirals in HIV-infected patients. Eur. J. Endocrinol. 2014, 170, R185–R202. [Google Scholar] [CrossRef]
- Shackleton, S.; Lloyd, D.J.; Jackson, S.N.; Evans, R.; Niermeijer, M.F.; Singh, B.M.; Schmidt, H.; Brabant, G.; Kumar, S.; Durrington, P.N.; et al. LMNA, encoding lamin A/C, is mutated in partial lipodystrophy. Nat. Genet. 2000, 24, 153–156. [Google Scholar] [CrossRef]
- Torriani, M.; Srinivasa, S.; Fitch, K.V.; Thomou, T.; Wong, K.; Petrow, E.; Kahn, C.R.; Cypess, A.M.; Grinspoon, S.K. Dysfunctional Subcutaneous Fat with Reduced Dicer and Brown Adipose Tissue Gene Expression in HIV-Infected Patients. J. Clin. Endocrinol. Metab. 2016, 10, 1225–1234. [Google Scholar] [CrossRef]
- Srinivasa, S.; Garcia-Martin, R.; Torriani, M.; Fitch, K.V.; Carlson, A.R.; Kahn, C.R.; Grinspoon, S.K. Altered pattern of circulating miRNAs in HIV lipodystrophy perturbs key adipose differentiation and inflammation pathways. JCI Insight 2021, 6, e150399. [Google Scholar] [CrossRef]
- Munier, S.; Borjabad, A.; Lemaire, M.; Mariot, V.; Hazan, U. In vitro infection of human primary adipose cells with HIV-1: A reassessment. AIDS 2003, 17, 2537–2539. [Google Scholar] [CrossRef] [PubMed]
- Couturier, J.; Suliburk, J.W.; Brown, J.M.; Luke, D.J.; Agarwal, N.; Yu, X.; Nguyen, C.; Iyer, D.; Kozinetz, C.A.; Overbeek, P.A.; et al. Human adipose tissue as a reservoir for memory CD4+ T cells and HIV. AIDS 2015, 29, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Damouche, A.; Lazure, T.; Avettand-Fènoël, V.; Huot, N.; Dejucq-Rainsford, N.; Satie, A.P.; Mélard, A.; David, L.; Gommet, C.; Ghosn, J.; et al. Adipose Tissue Is a Neglected Viral Reservoir and an Inflammatory Site during Chronic HIV and SIV Infection. PLoS Pathog. 2015, 11, e1005153. [Google Scholar] [CrossRef] [PubMed]
- Couturier, J.; Lewis, D.E. HIV Persistence in Adipose Tissue Reservoirs. Curr. HIV/AIDS Rep. 2018, 15, 60–71. [Google Scholar] [CrossRef]
- Gorwood, J.; Bourgeois, C.; Mantecon, M.; Atlan, M.; Pourcher, V.; Pourcher, G.; Le Grand, R.; Desjardins, D.; Fève, B.; Lambotte, O.; et al. Impact of HIV/simian immunodeficiency virus infection and viral proteins on adipose tissue fibrosis and adipogenesis. AIDS 2019, 33, 953–964. [Google Scholar] [CrossRef]
- Gorwood, J.; Ejlalmanesh, T.; Bourgeois, C.; Mantecon, M.; Rose, C.; Atlan, M.; Desjardins, D.; Le Grand, R.; Fève, B.; Lambotte, O.; et al. SIV Infection and the HIV Proteins Tat and Nef Induce Senescence in Adipose Tissue and Human Adipose Stem Cells, Resulting in Adipocyte Dysfunction. Cells 2020, 9, 854. [Google Scholar] [CrossRef]
- Agarwal, N.; Balasubramanyam, A. Viral mechanisms of adipose dysfunction: Lessons from HIV-1 Vpr. Adipocyte 2014, 4, 55–59. [Google Scholar] [CrossRef]
- Díaz-Delfín, J.; Domingo, P.; Wabitsch, M.; Giralt, M.; Villarroya, F. HIV-1 Tat protein impairs adipogenesis and induces the expression and secretion of proinflammatory cytokines in human SGBS adipocytes. Antivir. Ther. 2012, 17, 529–540. [Google Scholar] [CrossRef]
- Giralt, M.; Domingo, P.; Guallar, J.P.; Rodriguez de la Concepción, M.L.; Alegre, M.; Domingo, J.C.; Villarroya, F. HIV-1 infection alters gene expression in adipose tissue, which contributes to HIV- 1/HAART-associated lipodystrophy. Antivir. Ther. 2006, 11, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Sax, P.E.; Erlandson, K.M.; Lake, J.E.; Mccomsey, G.A.; Orkin, C.; Esser, S.; Brown, T.T.; Rockstroh, J.K.; Wei, X.; Carter, C.C.; et al. Weight Gain Following Initiation of Antiretroviral Therapy: Risk Factors in Randomized Comparative Clinical Trials. Clin. Infect. Dis. 2020, 71, 1379–1389. [Google Scholar] [CrossRef] [PubMed]
- Venter, W.D.F.; Moorhouse, M.; Sokhela, S.; Fairlie, L.; Mashabane, N.; Masenya, M.; Serenata, C.; Akpomiemie, G.; Qavi, A.; Chandiwana, N.; et al. Dolutegravir plus Two Different Prodrugs of Tenofovir to Treat HIV. N. Engl. J. Med. 2019, 381, 803–815. [Google Scholar] [CrossRef]
- Grabar, S.; Potard, V.; Piroth, L.; Abgrall, S.; Bernard, L.; Allavena, C.; Caby, F.; de Truchis, P.; Duvivier, C.; Enel, P.; et al. Striking differences in weight gain after cART initiation depending on early or advanced presentation: Results from the ANRS CO4 FHDH cohort. J. Antimicrob. Chemother. 2023, 78, 757–768. [Google Scholar] [CrossRef]
- Lake, J.E.; Wu, K.; Bares, S.H.; Debroy, P.; Godfrey, C.; Koethe, J.R.; McComsey, G.A.; Palella, F.J.; Tassiopoulos, K.; Erlandson, K.M. Risk Factors for Weight Gain Following Switch to Integrase Inhibitor-Based Antiretroviral Therapy. Clin. Infect. Dis. 2020, 71, e471–e477. [Google Scholar] [CrossRef]
- Bailin, S.S.; Koethe, J.R.; Rebeiro, P.F. The pathogenesis of obesity in people living with HIV. Curr. Opin. HIV AIDS 2024, 19, 6–13. [Google Scholar] [CrossRef]
- Domingo, P.; Villarroya, F.; Giralt, M.; Domingo, J.C. Potential role of the melanocortin signaling system interference in the excess weight gain associated to some antiretroviral drugs in people living with HIV. Int. J. Obes. 2020, 44, 1970–1973. [Google Scholar] [CrossRef] [PubMed]
- Chandiwana, N.C.; Siedner, M.J.; Marconi, V.C.; Hill, A.; Ali, M.K.; Batterham, R.L.; Venter, W.D.F. Weight Gain After HIV Therapy Initiation: Pathophysiology and Implications. J. Clin. Endocrinol. Metab. 2024, 109, e478–e487. [Google Scholar] [CrossRef]
- Ngono Ayissi, K.; Gorwood, J.; Le Pelletier, L.; Bourgeois, C.; Beaupère, C.; Auclair, M.; Foresti, R.; Motterlini, R.; Atlan, M.; Bar-rail-Tran, A.; et al. Inhibition of Adipose Tissue Beiging by HIV Integrase Inhibitors, Dolutegravir and Bictegravir, Is Associated with Adipocyte Hypertrophy, Hypoxia, Elevated Fibrosis, and Insulin Resistance in Simian Adipose Tissue and Human Adipocytes. Cells 2022, 11, 1841. [Google Scholar] [CrossRef]
- Gorwood, J.; Bourgeois, C.; Pourcher, V.; Pourcher, G.; Charlotte, F.; Mantecon, M.; Rose, C.; Morichon, R.; Atlan, M.; Le Grand, R.; et al. The Integrase Inhibitors Dolutegravir and Raltegravir Exert Proadipogenic and Profibrotic Effects and Induce Insulin Resistance in Human/Simian Adipose Tissue and Human Adipocytes. Clin. Infect. Dis. 2020, 71, e549–e560. [Google Scholar] [CrossRef]
- Domingo, P.; Quesada-López, T.; Villarroya, J.; Cairó, M.; Gutierrez, M.D.M.; Mateo, M.G.; Mur, I.; Corbacho, N.; Domingo, J.C.; Villarroya, F.; et al. Differential effects of dolutegravir, bictegravir and raltegravir in adipokines and inflammation markers on human adipocytes. Life Sci. 2022, 308, 120948. [Google Scholar] [CrossRef]
- Quesada-López, T.; Cereijo, R.; Blasco-Roset, A.; Mestres-Arenas, A.; Prieto, P.; Domingo, J.C.; Villarroya, F.; Domingo, P.; Giralt, M. Divergent effects of the antiretroviral drugs, dolutegravir, tenofovir alafenamide, and tenofovir disoproxil fumarate, on human adipocyte function. Biochem. Pharmacol. 2024, 220, 116010. [Google Scholar] [CrossRef] [PubMed]
- Perna, A.; Carleo, M.A.; Mascolo, S.; Guida, A.; Contieri, M.; Sellitto, C.; Hay, E.; De Blasiis, P.; Lucariello, A.; Guerra, G.; et al. Adipocyte differentiation of 3T3-L1 cells under tenofovir alafenamide, tenofovir disoproxil fumarate, and integrase strand transfer inhibitors selective challenge: An in-vitro model. AIDS 2023, 37, 561–570. [Google Scholar] [CrossRef]
- Hocqueloux, L.; Menard, A.; Arvieux, C.; Joly, V.; Becker, A.; Chéret, A.; Duvivier, C.; Cabié, A.; Delpierre, C.; Allavena, C.; et al. Weight gain following the single substitution of tenofovir disoproxil fumarate by tenofovir alafenamide in HIV-infected people from the French Dat’AIDS cohort: A propensity score-matched analysis. HIV Med. 2023, 24, 925–932. [Google Scholar] [CrossRef] [PubMed]
- Dulion, B.; Olali, A.Z.; Patel, N.; Virdi, A.K.; Naqib, A.; Wallace, J.; Ross, R.D. Tenofovir alafenamide promotes weight gain and impairs fatty acid metabolism-related signaling pathways in visceral fat tissue compared to tenofovir disoproxil fumarate. Antivir. Res. 2025, 237, 106151. [Google Scholar] [CrossRef] [PubMed]
- Milic, J.; Renzetti, S.; Ferrari, D.; Barbieri, S.; Menozzi, M.; Carli, F.; Dolci, G.; Ciusa, G.; Mussini, C.; Calza, S.; et al. Relationship between weight gain and insulin resistance in people living with HIV switching to integrase strand transfer inhibitors-based regimens. AIDS 2022, 36, 1643–1653. [Google Scholar] [CrossRef]
- González-Cordón, A.; Assoumou, L.; Moyle, G.; Waters, L.; Johnson, M.; Domingo, P.; Fox, J.; Stellbrink, H.J.; Guaraldi, G.; Masiá, M.; et al. Switching from boosted PIs to dolutegravir decreases soluble CD14 and adiponectin in high cardiovascular risk people living with HIV. J. Antimicrob. Chemother. 2021, 76, 2380–2393. [Google Scholar] [CrossRef]
- Summers, N.A.; Lahiri, C.D.; Angert, C.D.; Aldredge, A.; Mehta, C.C.; Ofotokun, I.; Kerchberger, A.M.; Gustafson, D.; Weiser, S.D.; Kassaye, S.; et al. Metabolic Changes Associated with the Use of Integrase Strand Transfer Inhibitors Among Virally Controlled Women. J. Acquir. Immune Defic. Syndr. 2020, 85, 355–362. [Google Scholar] [CrossRef]
- Neesgaard, B.; Greenberg, L.; Miró, J.M.; Grabmeier-Pfistershammer, K.; Wandeler, G.; Smith, C.; De Wit, S.; Wit, F.; Pelchen-Matthews, A.; Mussini, C.; et al. Associations between integrase strand-transfer inhibitors and cardiovascular disease in people living with HIV: A multicentre prospective study from the RESPOND cohort consortium. Lancet HIV 2022, 9, e474–e485. [Google Scholar] [CrossRef]
- Rebeiro, P.F.; Emond, B.; Rossi, C.; Bookhart, B.K.; Shah, A.; Caron-Lapointe, G.; Lafeuille, M.H.; Donga, P. Incidence of cardiometabolic outcomes among people living with HIV-1 initiated on integrase strand transfer inhibitor versus non-integrase strand transfer inhibitor antiretroviral therapies: A retrospective analysis of insurance claims in the United States. J. Int. AIDS Soc. 2023, 26, e26123. [Google Scholar] [CrossRef]
- Bastard, J.P.; Couffignal, C.; Fellahi, S.; Bard, J.M.; Mentre, F.; Salmon, D.; Katlama, C.; Raffi, F.; Leport, C.; Capeau, J.; et al. Diabetes and dyslipidaemia are associated with oxidative stress independently of inflammation in long-term antiretroviral-treated HIV-infected patients. Diabetes Metab. 2019, 45, 573–581. [Google Scholar] [CrossRef]
- Gelpi, M.; Afzal, S.; Lundgren, J.; Ronit, A.; Roen, A.; Mocroft, A.; Gerstoft, J.; Lebech, A.M.; Lindegaard, B.; Kofoed, K.F.; et al. Higher Risk of Abdominal Obesity, Elevated Low-Density Lipoprotein Cholesterol, and Hypertriglyceridemia, but not of Hypertension, in People Living with Human Immunodeficiency Virus (HIV): Results From the Copenhagen Comorbidity in HIV Infection Study. Clin. Infect. Dis. 2018, 67, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Arrive, E.; Viard, J.P.; Salanave, B.; Dollfus, C.; Matheron, S.; Reliquet, V.; Arezes, E.; Nailler, L.; Vigouroux, C.; Warszawski, J.; et al. Metabolic risk factors in young adults infected with HIV since childhood compared with the general population. PLoS ONE 2018, 13, e0206745. [Google Scholar] [CrossRef]
- Bailin, S.S.; Gabriel, C.L.; Gangula, R.D.; Hannah, L.; Nair, S.; Carr, J.J.; Terry, J.G.; Silver, H.J.; Simmons, J.D.; Mashayekhi, M.; et al. Single-Cell Analysis of Subcutaneous Fat Reveals Profibrotic Cells That Correlate with Visceral Adiposity in HIV. J. Clin. Endocrinol. Metab. 2024, 110, 238–253. [Google Scholar] [CrossRef] [PubMed]
- Janssen-Telders, C.; Eringa, E.C.; de Groot, J.R.; de Man, F.S.; Handoko, M.L. The role of epicardial adipose tissue remodelling in heart failure with preserved ejection fraction. Cardiovasc. Res. 2025, 16, cvaf056. [Google Scholar] [CrossRef] [PubMed]
- Han, W.M.; Apornpong, T.; Tumkosit, M.; Avihingsanon, A.; Chattranukulchai, P. Epicardial fat tissue and diastolic dysfunction in both men and women with HIV. AIDS 2024, 38, 1893–1894. [Google Scholar] [CrossRef]
- Goldberg, R.L.; Peterson, T.E.; Haberlen, S.A.; Witt, M.D.; Palella, F.J.; Magnani, J.W.; Brown, T.T.; Lake, J.E.; Lima, J.A.C.; Budoff, M.J.; et al. Response to “Epicardial fat tissue and diastolic dysfunction in both men and women with HIV”. AIDS 2024, 38, 1895. [Google Scholar] [CrossRef]
- Foldyna, B.; Mayrhofer, T.; Zanni, M.V.; Lyass, A.; Barve, R.; Karady, J.; McCallum, S.; Burdo, T.H.; Fitch, K.V.; Paradis, K.; et al. Pericoronary Adipose Tissue Density, Inflammation, and Subclinical Coronary Artery Disease Among People with HIV in the REPRIEVE Cohort. Clin. Infect. Dis. 2023, 77, 1676–1686. [Google Scholar] [CrossRef]
- Wilcox, C.S.; Herbert, C.; Wang, C.; Ma, Y.; Sun, P.; Li, T.; Verbesey, J.; Kumar, P.; Kassaye, S.; Welch, W.J.; et al. Signals From Inflamed Perivascular Adipose Tissue Contribute to Small-Vessel Dysfunction in Women with Human Immunodeficiency Virus. J. Infect. Dis. 2024, 230, 67–77. [Google Scholar] [CrossRef]
- Basta, D.; Latinovic, O.S.; Tagaya, Y.; Silvestri, G. Potential Advantages of a Well-balanced Nutrition Regimen for People Living with Human Immunodeficiency Virus Type-1. J. AIDS HIV Treat. 2024, 6, 11–27. [Google Scholar] [CrossRef]
- Domingo, P.; Fernández, I.; Gallego-Escuredo, J.M.; Torres, F.; Gutierrez, M.D.M.; Mateo, M.G.; Villarroya, J.; Giralt, M.; Vidal, F.; Villarroya, F.; et al. Effects of docosahexanoic acid on metabolic and fat parameters in HIV-infected patients on cART: A randomized, double-blind, placebo-controlled study. Clin. Nutr. 2018, 37, 1340–1347. [Google Scholar] [CrossRef] [PubMed]
- Pastor-Ibáñez, R.; Blanco-Heredia, J.; Etcheverry, F.; Sánchez-Palomino, S.; Díez-Fuertes, F.; Casas, R.; Navarrete-Muñoz, M.Á.; Castro-Barquero, S.; Lucero, C.; Fernández, I.; et al. Adherence to a Supplemented Mediterranean Diet Drives Changes in the Gut Microbiota of HIV-1-Infected Individuals. Nutrients 2021, 13, 1141. [Google Scholar] [CrossRef] [PubMed]
- Haidar, L.; Crane, H.M.; Nance, R.M.; Webel, A.; Ruderman, S.A.; Whitney, B.M.; Willig, A.L.; Napravnik, S.; Mixson, L.S.; Leong, C.; et al. Weight loss associated with semaglutide treatment among people with HIV. AIDS 2024, 38, 531–535. [Google Scholar] [CrossRef]
- Lee, D.; Capeau, J. Is the GLP-1 receptor agonist, semaglutide, a good option for weight loss in persons with HIV? AIDS 2024, 38, 603–605. [Google Scholar] [CrossRef] [PubMed]
- Eckard, A.R.; Wu, Q.; Sattar, A.; Ansari-Gilani, K.; Labbato, D.; Foster, T.; Fletcher, A.A.; Adekunle, R.O.; McComsey, G.A. Once-weekly semaglutide in people with HIV-associated lipohypertrophy: A randomised, double-blind, placebo-controlled phase 2b single-centre clinical trial. Lancet Diabetes Endocrinol. 2024, 12, 523–534. [Google Scholar] [CrossRef]
- Thomas, T.S.; Srinivasa, S. Weighing in: Glucagon-like Peptide-1 Receptor Agonism for Persons with HIV. Top. Antivir. Med. 2024, 32, 579–588. [Google Scholar]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giralt, M.; Domingo, P.; Quesada-López, T.; Cereijo, R.; Villarroya, F. Lipodystrophy in HIV: Evolving Challenges and Unresolved Questions. Int. J. Mol. Sci. 2025, 26, 6546. https://doi.org/10.3390/ijms26146546
Giralt M, Domingo P, Quesada-López T, Cereijo R, Villarroya F. Lipodystrophy in HIV: Evolving Challenges and Unresolved Questions. International Journal of Molecular Sciences. 2025; 26(14):6546. https://doi.org/10.3390/ijms26146546
Chicago/Turabian StyleGiralt, Marta, Pere Domingo, Tania Quesada-López, Rubén Cereijo, and Francesc Villarroya. 2025. "Lipodystrophy in HIV: Evolving Challenges and Unresolved Questions" International Journal of Molecular Sciences 26, no. 14: 6546. https://doi.org/10.3390/ijms26146546
APA StyleGiralt, M., Domingo, P., Quesada-López, T., Cereijo, R., & Villarroya, F. (2025). Lipodystrophy in HIV: Evolving Challenges and Unresolved Questions. International Journal of Molecular Sciences, 26(14), 6546. https://doi.org/10.3390/ijms26146546