
Current Approaches to Aflatoxin B1 Control in Food and Feed Safety: Detection, Inhibition, and Mitigation
 , , , , ,
, , , , ,  , , ,
, , ,
Abstract

1. Introduction
2. Regulations on Aflatoxins in Animal Feed: An Analysis of Key Producing Countries
- Feed materials: 20 μg/kg.
- Complementary and complete feed mixtures: 10 μg/kg.
- Feed mixtures for dairy cattle and calves, dairy sheep and lambs, dairy goats and kids, piglets, and young poultry: 5 μg/kg.
- General feed mixtures for other categories not specified above: 20 μg/kg [21].
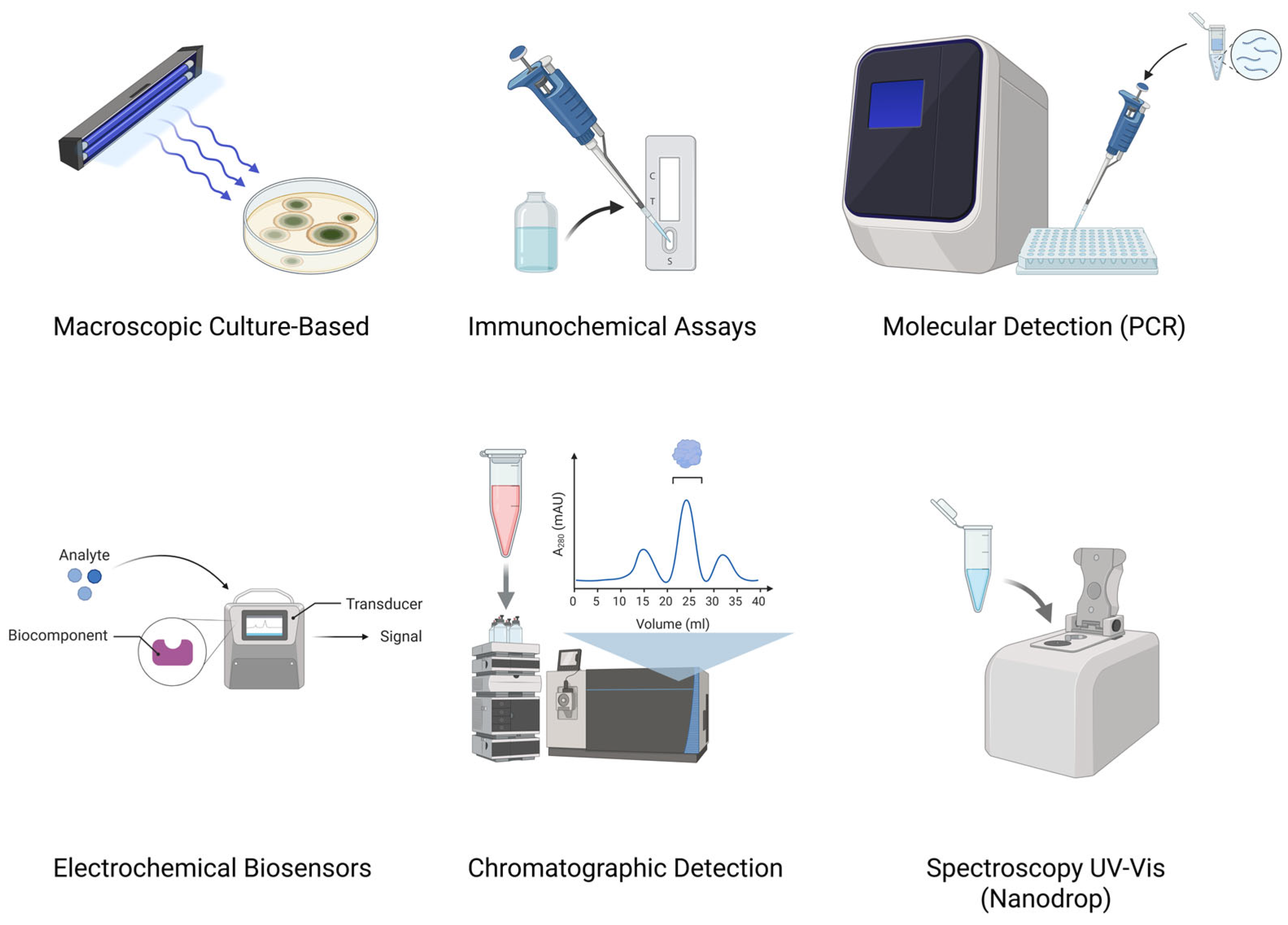
3. Aflatoxin Detection
3.1. Macroscopic Culture-Based Methods
3.2. Molecular Methods
3.3. Immunochemical Methods

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method | Detecting Matrix | Detected Aflatoxin | Limit of Detection for Aflatoxins [μg/kg] | References |
|---|---|---|---|---|
| Rapid immunochromatographic strip | Monoclonal antibody-gold nanoparticles (mAb-AuNP) | AFB1 | 1.0 | [39] |
| Monoclonal antibody-based fluorescent microsphere immunochromatographic test strip | Fluorescent microspheres–mAb | AFM1 | 4.4 | [40] |
| One-step immunochromatographic assay | mAb-AuNP | AFM1 | 0.05 (EU) 0.5 (others) | [41] |
| Quantum dot nanobead-based multiplexed immunochromatographic assay | Quantum dot nanobead with antibody | AFB1 | 0.00165 | [42] |
| Gold nanoparticle-based conjugated AFB1 antifungal strips | AuNPs conjugated with AFB1 antibody and bovine serum albumin | AFB1 | 10 | [43] |
| Nanoparticle-based competitive magnetic immunodetection | Biotinylated mAb, magnetic particles functionalized with streptavidin | AFB1 | 1.1 | [44] |
| Two-analyte immunochromatographic strip | Protein conjugates (AFM1-OVA (aflatoxin M1–ovalbumin conjugate) and chloramphenicol-ovalbumin) and goat anti-rabbit IgG (immunoglobulin G) | AFM1 | 0.1 | [45] |
| Immunochromatographic test | Antigen-modified Fe2O3 nanoprobes | AFB1 | 0.0125 | [36] |
| Pressure/colorimetric dual-readout immunochromatographic test strip | Dendritic platinum nanoparticles | AFB1 | 0.03 | [46] |
| Lateral flow immunochromatographic assay | Sprayed coupled antigens AFB1-ovalbumin (AFB1-OVA) and ochratoxin A–ovalbumin (OTA-OVA) | AFB1 | 5.0 | [47] |
| Noncompetitive immunocomplex immunoassay | Monoclonal capture antibody and a unique anti-immunocomplex antibody fragment isolated from a synthetic antibody repertoire | AFB1 | 0.07 | [48] |
| Lateral flow immunochromatographic assay | mAbs-AuNP | AFB1 | 1.0 | [49] |
| Dual immunochromatographic test strip | Double antibodies labeled with time-resolved fluorescent microspheres | AFM1 | 0.018 | [37] |
| Gold immunochromatographic test strip | mAbs-AuNP | AFB1 | 0.5 | [50] |
| Lateral flow immunochromatographic assay | Avi-tag (avidin tag)/streptavidin-oriented coupling strategy | AFB1 | 0.095 | [51] |
3.4. Electrochemical Biosensor-Based Methods
3.4.1. Ultrasensitive Devices Based on Polymer-Based Biosensors
3.4.2. Sensitive Portable Devices
3.5. Chromatographic Methods
3.6. Spectroscopic Methods
3.7. Summary of Detection Methods
3.8. Comparative Summary of Aflatoxin Detection Methods
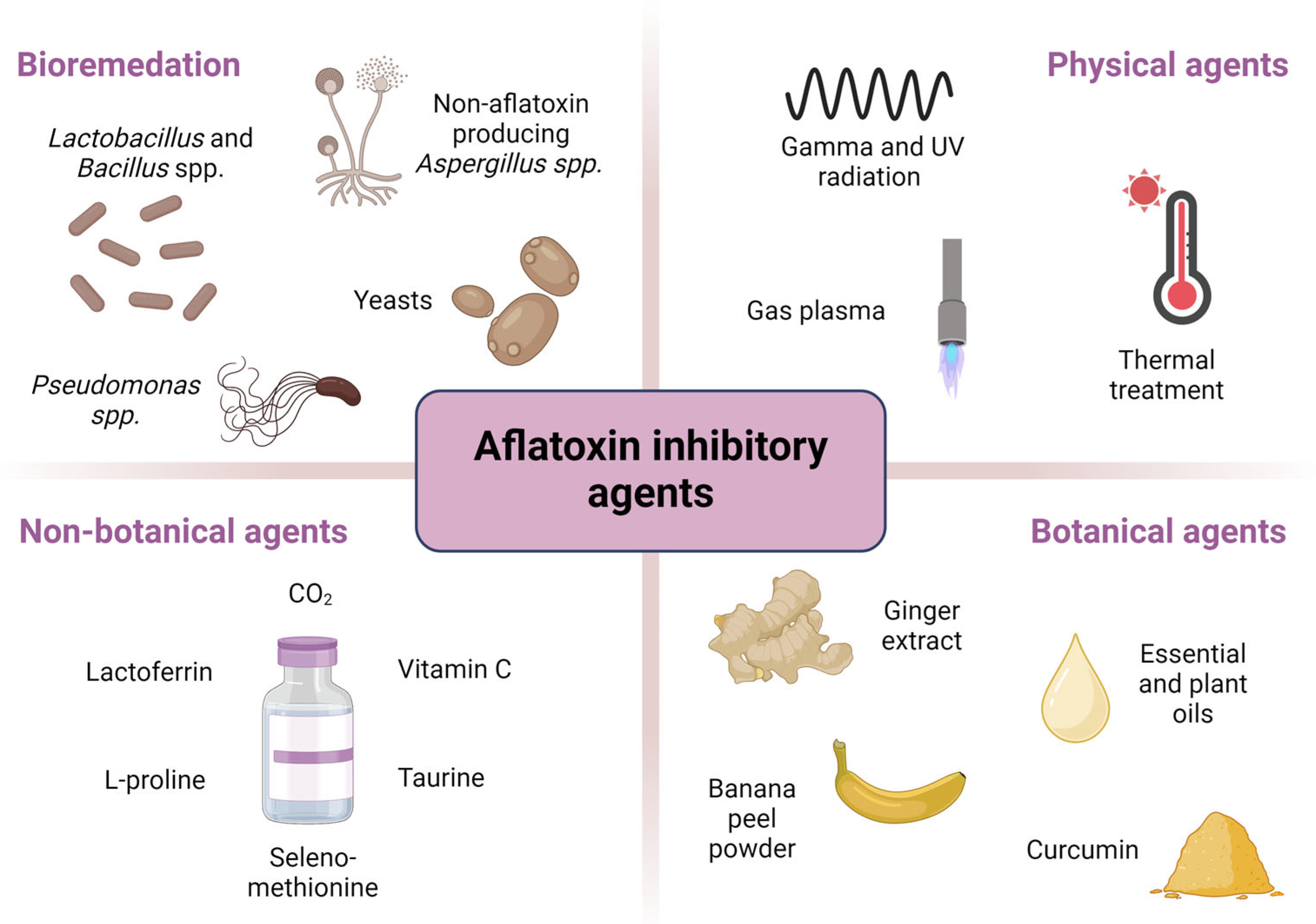
4. Inhibitory Agents
4.1. Bioremediation Agents
4.2. Physical Agents
4.3. Non-Botanical Agents
| Aflatoxin and/or Fungus | Type of Non-Botanical Agent | Model | Effects | References |
|---|---|---|---|---|
| AFB1 | Mixture of citric and phosphoric acids with arginine | in vitro |
| [132] |
| AFB1 AFM1 | Lactoferrin | Caco-2, HEK 293, Hep-G2, and SK-N-SH cells in vitro |
| [134] |
| A. parasiticus ATCC15517 AFB1 AFB2 AFG1 AFG2 | Vitamin C | in vitro |
| [135] |
| A. flavus A42 and CHAO50 | CO2 | in vitro |
| [136] |
| AFB1 AFM1 | L-proline | HEK 293 cells in vitro; mice in vivo |
| [137] |
| AFB1 | Chlorine dioxide gas | in vitro |
| [138] |
| AFB1 | Complementary feed Rhino-Hepato Forte | Chicken in vivo |
| [139] |
| AFB1 | Taurine | Rats in vivo |
| [140] |
| AFB1 | Seleno-methionine | Rabbits in vivo |
| [141] |
4.4. Botanical Agents
4.5. Integrated Decontamination Strategies
5. Occurrence of Aflatoxins in Water: Detection and Elimination Strategies
5.1. Detection of Aflatoxins in Water
5.2. Elimination of Aflatoxins from Water
5.3. Regulatory and Research Considerations
6. Perspectives on Detection and Elimination of Aflatoxins
6.1. Artificial Intelligence (AI) and Machine Learning (ML)
6.2. Intelligent and Active Packaging (AP)
6.3. Transgenic Plants
7. Future Perspectives
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 8-OHdG | 8-hydroxy-2′-deoxyguanosine |
| AChE | acetylcholinesterase |
| AFB1 | aflatoxin B1 |
| AFB2 | aflatoxin B2 |
| AFB2a-Arg | aflatoxin B2a–arginine adduct |
| AFG1 | aflatoxin G1 |
| AFG2 | aflatoxin G2 |
| AFM1 | aflatoxin M1 |
| AFM1-OVA | aflatoxin M1–ovalbumin conjugate |
| AFs | aflatoxins |
| AI | artificial intelligence |
| AKT | protein kinase B |
| ALP | alkaline phosphatase |
| AP | active packaging |
| APA | AF-producing ability |
| AST | aspartate aminotransferase |
| ATR-FTIR | attenuated total reflectance–Fourier transform infrared spectroscopy |
| AuNP | gold nanoparticles |
| Avi-tag | avidin tag |
| Bax | Bcl-2-associated X protein |
| Bcl-2 | B-cell lymphoma 2 |
| BIS | Bureau of Indian Standards |
| CAM | coconut agar medium |
| CAP | cold atmospheric plasma |
| CAT | catalase |
| CHAO50 | chao1 richness estimator |
| CK | creatine kinase |
| CK-MB | creatine kinase myocardial band |
| CMA | coconut milk agar |
| CRISPR-Cas9 | clustered regularly interspaced short palindromic repeats—CRISPR-associated protein 9 |
| CS | chitosan |
| Cyt-c | cytochrome c |
| EC | European Commission |
| ELISA | enzyme-linked immunosorbent assay |
| ERK1/2 | extracellular signal-regulated kinases 1 and 2 |
| EU | European Union |
| FAO | Food and Agriculture Organization |
| FDA | Food and Drug Administration |
| GCLC | glutamate–cysteine ligase catalytic subunit |
| GCLM | glutamate–cysteine ligase modifier subunit |
| GC-MS | gas chromatography–mass spectrometry |
| Gd-MOF/USPIO | gadolinium-based metal–organic framework/ultrasmall superparamagnetic iron oxide |
| GGT | gamma-glutamyl transferase |
| GM | genetically modified |
| GPx | glutathione peroxidase |
| GSH | glutathione |
| GSS | glutathione synthetase |
| GSTA1 | glutathione S-transferase A1 |
| H/L stress ratio | heterophil-to-lymphocyte stress ratio |
| HB | H. bacciferum |
| HIGS | host-induced gene silencing |
| HO-1 | heme oxygenase 1 |
| HPLC | high-performance liquid chromatography |
| HPLC-HRMS | high-performance liquid chromatography–high resolution mass spectrometry |
| HPLC-MS/MS | high-performance liquid chromatography–tandem mass spectrometry |
| IARC | International Agency for Research on Cancer |
| IDO | indoleamine-pyrrole 2,3-dioxygenase |
| IgG | immunoglobulin G |
| IL-1β | interleukin 1 beta |
| IoT | Internet of Things |
| IR | infrared |
| JNK | c-Jun N-terminal kinase |
| Keap1 | Kelch-like ECH-associated protein 1 |
| kGy | kilogray |
| LC-MS/MS | liquid chromatography–tandem mass spectrometry |
| LDH | lactate dehydrogenase |
| LOD | limit of detection |
| mAb | monoclonal antibody |
| MDA | malondialdehyde |
| MIC | minimum inhibitory concentration |
| ML | machine learning |
| MRS | magnetic relaxation switching |
| MWCNTs | multi-walled carbon nanotubes |
| NF-kB | nuclear factor kappa B |
| NIR | near-infrared |
| NQO1 | NAD(P)H quinone dehydrogenase 1 |
| Nrf2 | nuclear factor erythroid 2-related factor 2 |
| OD | O. dhofarense |
| p-AKT | phosphorylated AKT |
| PCR | polymerase chain reaction |
| PDA medium | potato dextrose agar |
| PDB medium | potato dextrose broth |
| PD-IPCR | proximity-dependent immuno-PCR |
| PI3K | phosphoinositide 3-kinase |
| POCT | point-of-care testing |
| POF | plastic optical fiber |
| ppb | parts per billion |
| PSO | pumpkin seed oil |
| PSO-NP | pumpkin seed oil nanoparticles |
| PTEN | phosphatase and tensin homologue |
| qPCR | quantitative PCR |
| QuEChERS | quick, easy, cheap, effective, rugged, and safe |
| RNAi | RNA interference |
| SMILE | smartphone-powered mobile microfluidic lab-on-fiber device |
| SMRs | superparamagnetic nanoparticles |
| SOD | superoxide dismutase |
| SPCE | screen-printed carbon electrode |
| TAC | total antioxidant capacity |
| TBARS | thiobarbituric acid reactive substances |
| TIMP3 | tissue inhibitor metallopeptidase 3 |
| TNF-α | tumor necrosis factor alpha |
| UHPLC-ESI-QTOF | ultrahigh-performance liquid chromatography–electrospray ionization–quadrupole time-of-flight mass spectrometry |
| UV light | ultraviolet light |
| UVC | ultraviolet light, C band |
| UV-vis–NIR | ultraviolet visible–near infrared |
| vis | visible |
| WHO | World Health Organization |
| YES | yeast extract sucrose |
| ZM | Z. multiflora |
| Zr-LMOF@Cotton | zirconium-based luminescent metal–organic framework on cotton |
References
- Kumar, P.; Mahato, D.K.; Kamle, M.; Mohanta, T.K.; Kang, S.G. Aflatoxins: A global concern for food safety, human health and their management. Front. Microbiol. 2016, 7, 2170. [Google Scholar] [CrossRef] [PubMed]
- Dai, C.; Tian, E.; Hao, Z.; Tang, S.; Wang, Z.; Sharma, G.; Jiang, H.; Shen, J. Aflatoxin B1 toxicity and protective effects of curcumin: Molecular mechanisms and clinical implications. Antioxidants 2022, 11, 2031. [Google Scholar] [CrossRef]
- Ahmad, M.M.; Qamar, F.; Saifi, M.; Abdin, M.Z. Natural inhibitors: A sustainable way to combat aflatoxins. Front. Microbiol. 2022, 13, 993834. [Google Scholar] [CrossRef]
- International Agency for Research on Cancer (IARC). Aflatoxins. In IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; IARC: Lyon, France, 2012; Volume 100, pp. 225–248. [Google Scholar]
- Ofori-Attah, E.; Hashimoto, M.; Oki, M.; Kadowaki, D. Therapeutic effect of natural products and dietary supplements on aflatoxin-induced nephropathy. Int. J. Mol. Sci. 2024, 25, 2849. [Google Scholar] [CrossRef]
- Faraji, H.; Yazdi, F.T.; Razmi, N. The influence of ultraviolet radiation on aflatoxin producing Aspergillus species isolated from Iranian rice. Toxicol. Rep. 2022, 9, 1528–1536. [Google Scholar] [CrossRef]
- Romero-Sánchez, I.; Gracia-Lor, E.; Madrid-Albarrán, Y. Aflatoxin detoxification by thermal cooking treatment and evaluation of in vitro bioaccessibility from white and brown rice. Food Chem. 2024, 436, 137738. [Google Scholar] [CrossRef]
- Nazareth, T.d.M.; Soriano Pérez, E.; Luz, C.; Meca, G.; Quiles, J.M. Comprehensive review of aflatoxin and ochratoxin A dynamics: Emergence, toxicological impact, and advanced control strategies. Foods 2024, 13, 1920. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, Q.; Chen, Z.-Y.; Liu, H.; Li, P. Investigation of Pseudomonas fluorescens strain 3JW1 on preventing and reducing aflatoxin contaminations in peanuts. PLoS ONE 2017, 12, e0178810. [Google Scholar] [CrossRef]
- Popescu, R.G.; Rădulescu, A.L.; Georgescu, S.E.; Dinischiotu, A. Aflatoxins in feed: Types, metabolism, health consequences in swine and mitigation strategies. Toxins 2022, 14, 853. [Google Scholar] [CrossRef]
- Mahmoud, Y.A.; Elkaliny, N.E.; Darwish, O.A.; Ashraf, Y.; Ebrahim, R.A.; Das, S.P.; Yahya, G. Comprehensive review for aflatoxin detoxification with special attention to cold plasma treatment. Mycotoxin Res. 2025, 41, 277–300. [Google Scholar] [CrossRef]
- Jalili, C.; Ranjbar Shamsi, R.; Amiri, B.; Kakebaraie, S.; Jalili, F.; Nasta, T.Z. Genotoxic and cytotoxic effects of aflatoxin on the reproductive system: Focus on cell cycle dynamics and apoptosis in testicular tissue. Toxicology 2024, 504, 153773. [Google Scholar] [CrossRef] [PubMed]
- FAO; WHO. Aflatoxins. In Evaluation of Certain Contaminants in Food; WHO; FAO: Geneva, Switzerland, 2017; pp. 11–28. [Google Scholar]
- Navale, V.; Vamkudoth, K.R.; Ajmera, S.; Dhuri, V. Aspergillus derived mycotoxins in food and the environment: Prevalence, detection, and toxicity. Toxicol. Rep. 2021, 8, 1008–1030. [Google Scholar] [CrossRef] [PubMed]
- Miklós, G.; Angeli, C.; Ambrus, Á.; Nagy, A.; Kardos, V.; Zentai, A.; Kerekes, K.; Farkas, Z.; Jóźwiak, Á.; Bartók, T. Detection of aflatoxins in different matrices and food-chain positions. Front. Microbiol. 2020, 11, 1916. [Google Scholar] [CrossRef] [PubMed]
- Rushing, B.R.; Selim, M.I. Adduction to arginine detoxifies aflatoxin B1 by eliminating genotoxicity and altering in vitro toxicokinetic profiles. Oncotarget 2017, 9, 4559–4570. [Google Scholar] [CrossRef]
- El-Shanshoury, A.R.; Metwally, M.A.; El-Sabbagh, S.M.; Emara, H.A.; Saba, H.E. Biocontrol of Aspergillus flavus producing aflatoxin B1 by Streptomyces exfoliates. Egypt. J. Bot. 2022, 62, 457–473. [Google Scholar]
- Farghl, A.A.M.; El-Sheekh, M.M.; El-Shahir, A.A. Seaweed extracts as biological control of aflatoxins produced by Aspergillus parasiticus and Aspergillus flavus. Egypt. J. Biol. Pest Control 2023, 33, 50. [Google Scholar] [CrossRef]
- Hojjati, M.; Shahbazi, S.; Askari, H.; Makari, M. Use of X-Irradiations in reducing the waste of aflatoxin-contaminated pistachios and evaluation of the physicochemical properties of the irradiated product. Foods 2023, 12, 3040. [Google Scholar] [CrossRef]
- García-Ramón, D.F.; Cornelio-Santiago, H.P.; Norabuena, E.; Sumarriva, L.; Álvarez-Chancasanampa, H.; Vega, M.N.; Sotelo-Méndez, A.; Espinoza-Espinoza, L.A.; Pantoja-Tirado, L.R.; Gonzales-Agama, S.H.; et al. Effective novel and conventional technologies for decontamination of aflatoxin B1 in foods: A review. Mycotoxin Res. 2025, 41, 301–321. [Google Scholar] [CrossRef]
- European Commission. Commission Regulation No 1881/2006: Setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Union 2006, L 364, 7–9, Issue 01.07.2010. [Google Scholar]
- Missouri Department of Agriculture (MDA). Aflatoxin Information. Available online: https://agriculture.mo.gov/plants/feed/aflatoxin.php (accessed on 28 October 2024).
- Hao, W.H.; Li, A.; Wang, J.; An, G.; Guan, S. Mycotoxin contamination of feeds and raw materials in China in year 2021. Front. Vet. Sci. 2022, 9, 929904. [Google Scholar] [CrossRef]
- Brasil Ministério da Saúde. Agência Nacional de Vigilância Sanitária. Resolução RDC N. 7, de 18 de fevereiro de 2011. Aprova o Regulamento Técnico sobre limites máximos tolerados (LMT) para micotoxinas em alimentos; Seção 1. D. Of. Da Repúb. Fed. Do Bras. 2011, 46, 66. [Google Scholar]
- Thakur, S.; Singh, R.K.; De, P.S.; Dey, A. Aflatoxins in feeds: Issues and concerns with safe food production. Indian J. Anim. Health 2022, 61, 1–13. [Google Scholar] [CrossRef]
- Sowmya, K.L.; Ramalingappa, B. Rapid detection of aflatoxin production by Aspergillus flavus using coconut agar medium. Food Sci. Nutr. Technol. 2024, 9, 000352. [Google Scholar] [CrossRef]
- Ronoh, P.K.; Toroitich, F.J.; Makonde, H.M.; Lelmen, E.K.; Obonyo, M.A. Reliability of the chemical, metabolic, and molecular methods in discriminating aflatoxigenic from non-aflatoxigenic Aspergillus isolates. Microbe 2024, 4, 100115. [Google Scholar] [CrossRef]
- Alameri, M.M.; Kong, S.-Y.; Aljaafari, M.N.; Ali, H.A.; Eid, K.; Sallagi, M.A.; Cheng, W.-H.; Abushelaibi, A.; Lim, S.-H.E.; Loh, J.-Y.; et al. Aflatoxin contamination: An overview on health issues, detection and management strategies. Toxins 2023, 15, 246. [Google Scholar] [CrossRef]
- Sudini, H.; Srilakshmi, P.; Kumar, K.V.K.; Njoroge, S.M.C.; Osiru, M.; Seetha, A.; Waliyar, F. Detection of aflatoxigenic Aspergillus strains by cultural and molecular methods: A critical review. Afr. J. Microbiol. Res. 2015, 9, 484–491. [Google Scholar] [CrossRef]
- Akinola, S.A.; Ateba, C.N.; Mwanza, M. Polyphasic assessment of aflatoxin production potential in selected Aspergilli. Toxins 2019, 11, 692. [Google Scholar] [CrossRef]
- Bharose, A.A.; Hajare, S.T.; Narayanrao, D.R.; Gajera, H.G.; Prajapati, H.K.; Singh, S.C.; Upadhye, V. Whole genome sequencing and annotation of Aspergillus flavus JAM-JKB-B HA-GG20. Sci. Rep. 2024, 14, 18. [Google Scholar] [CrossRef]
- Ortega, S.F.; Siciliano, I.; Prencipe, S.; Gullino, M.L.; Spadaro, D. Development of PCR, LAMP, and qPCR assays for the detection of aflatoxigenic strains of Aspergillus flavus and A. parasiticus in hazelnut. Toxins 2020, 12, 757. [Google Scholar] [CrossRef]
- Mahmoud, M.A. Detection of Aspergillus flavus in stored peanuts using real-time PCR and the expression of aflatoxin genes in toxigenic and atoxigenic A. flavus isolates. Foodborne Pathog. Dis. 2015, 12, 289–296. [Google Scholar] [CrossRef]
- Bintvihok, A.; Treebonmuang, S.; Srisakwattana, K.; Nuanchun, W.; Patthanachai, K.; Usawang, S. A rapid and sensitive detection of aflatoxin-producing fungus using an optimized polymerase chain reaction (PCR). Toxicol. Res. 2016, 32, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Qu, W.; Hao, X.; Fang, M.; Yang, Q.; Li, Y.; Gong, Z.; Li, P. Immunochromatographic strip based on tetrahedral DNA immunoprobe for the detection of aflatoxin B1 in rice bran oil. Foods 2024, 13, 2410. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Bu, T.; Zhao, S.; Bai, F.; Zhang, M.; Tian, Y.; Sun, X.; Dong, M.; Wang, L. Well-orientation strategy for direct binding of antibodies: Development of the immunochromatographic test using the antigen modified Fe2O3 nanoprobes for sensitive detection of aflatoxin B1. Food Chem. 2021, 364, 129583. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Li, M.; Xing, F.; Wang, H.; Zhang, Y.; Sun, X. Novel dual immunochromatographic test strip based on double antibodies and biotin-streptavidin system for simultaneous sensitive detection of aflatoxin M1 and ochratoxin A in milk. Food Chem. 2022, 375, 131682. [Google Scholar] [CrossRef]
- Zhuo, Y.; Xu, W.; Chen, Y.; Long, F. Rapid and sensitive point-of-need aflatoxin B1 testing in feedstuffs using a smartphone-powered mobile microfluidic lab-on-fiber device. J. Hazard. Mater. 2023, 460, 132406. [Google Scholar] [CrossRef]
- Liu, J.-W.; Lu, C.C.; Liu, B.-H.; Yu, F.-Y. Development of novel monoclonal antibodies-based ultrasensitive enzyme-linked immunosorbent assay and rapid immunochromatographic strip for aflatoxin B1 detection. Food Control 2016, 59, 700–707. [Google Scholar] [CrossRef]
- Zhang, X.; Wen, K.; Wang, Z.; Jiang, H.; Beier, R.C.; Shen, J. An ultra-sensitive monoclonal antibody-based fluorescent microsphere immunochromatographic test strip assay for detecting aflatoxin M1 in milk. Food Control 2016, 60, 588–595. [Google Scholar] [CrossRef]
- Wu, C.; Liu, D.; Peng, T.; Shan, S.; Zhang, G.; Xiong, Y.; Lai, W. Development of a one-step immunochromatographic assay with two cutoff values of aflatoxin M1. Food Control 2016, 63, 11–14. [Google Scholar] [CrossRef]
- Shao, Y.; Duan, H.; Guo, L.; Leng, Y.; Lai, W.; Xiong, Y. Quantum dot nanobead-based multiplexed immunochromatographic assay for simultaneous detection of aflatoxin B1 and zearalenone. Anal. Chim. Acta 2018, 1025, 163–171. [Google Scholar] [CrossRef]
- Sojinrin, T.; Liu, K.; Wang, K.; Cui, D.; Byrne, H.J.; Curtin, J.F.; Tian, F. Developing gold nanoparticles-conjugated aflatoxin B1 antifungal strips. Int. J. Mol. Sci. 2019, 20, 6260. [Google Scholar] [CrossRef]
- Pietschmann, J.; Spiegel, H.; Krause, H.-J.; Schillberg, S.; Schröper, F. Sensitive aflatoxin B1 detection using nanoparticle-based competitive magnetic immunodetection. Toxins 2020, 12, 337. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.-W.; Ko, J.-L.; Liu, B.-H.; Yu, F.-Y. A sensitive two-analyte immunochromatographic strip for simultaneously detecting aflatoxin M1 and chloramphenicol in milk. Toxins 2020, 12, 637. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Zhang, L.; Li, J.; Ouyang, H.; Fu, Z. Pressure/colorimetric dual-readout immunochromatographic test strip for point-of-care testing of aflatoxin B1. Talanta 2021, 227, 122203. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Jin, X.; Lin, Z.; Guo, Q.; Liu, B.; Yuan, Y.; Yue, T.; Zhao, X. Simultaneous rapid detection of aflatoxin B1 and ochratoxin A in spices using lateral flow immuno-chromatographic assay. Foods 2021, 10, 2738. [Google Scholar] [CrossRef]
- Peltomaa, R.; Abbas, A.; Yli-Mattila, T.; Lamminmäki, U. Single-step noncompetitive immunocomplex immunoassay for rapid aflatoxin detection. Food Chem. 2022, 392, 133287. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, F.; Zhou, X.; Liu, M.; Zang, H.; Liu, X.; Shan, A.; Feng, X. Alleviation of oral exposure to aflatoxin B1-induced renal dysfunction, oxidative stress, and cell apoptosis in mice kidney by curcumin. Antioxidants 2022, 11, 1082. [Google Scholar] [CrossRef]
- Zhang, J.; Li, X.; Xie, J.; Huang, Z. Rapid and simultaneous detection of aflatoxin B1, zearalenone, and T-2 toxin in medicinal and edible food using gold immunochromatographic test strip. Foods 2023, 12, 633. [Google Scholar] [CrossRef]
- Wang, X.; Sun, T.; Shen, W.; Liu, M.; Liu, W.; Zuo, H.; Zhang, Y.; Geng, L.; Wang, W.; Shao, C.; et al. A lateral flow immunochromatographic assay based on nanobody-oriented coupling strategy for aflatoxin B1 detection. Sens. Actuators B Chem. 2023, 394, 134419. [Google Scholar] [CrossRef]
- Liu, S.; Jiang, S.; Yao, Z.; Liu, M. Aflatoxin detection technologies: Recent advances and future prospects. Environ. Sci. Pollut. Res. Int. 2023, 30, 79627–79653. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Y.; Han, J.; Chu, P.K.; Feng, J.; Dong, Y. A sensitive non-enzymatic immunosensor composed of silver nanoflowers for squamous cell carcinoma antigen. RSC Adv. 2017, 4, 2242–2248. [Google Scholar] [CrossRef]
- Pal, T.; Aditya, S.; Mathai, T.; Mukherji, S. Polyaniline-coated plastic optic fiber biosensor for detection of aflatoxin B1 in nuts, cereals, beverages, and body fluids. Sens. Actuators B Chem. 2023, 389, 133897. [Google Scholar] [CrossRef]
- Lu, Y.; Chen, R.; Dong, Y.; Zhao, W.; Ruan, S.; Yang, W.; Chen, Y.; Wang, C. Magnetic relaxation switching immunoassay based on “limited-magnitude” particles for sensitive quantification of aflatoxin B1. Anal. Chim. Acta 2023, 1266, 341329. [Google Scholar] [CrossRef] [PubMed]
- Guo, G.; Wang, T.; Liu, Z.; Liu, X.; Li, T.; Chen, Y.; Fan, J.; Bukye, E.; Huang, X.; Song, L. A self-assembled DNA double-crossover-based fluorescent aptasensor for highly sensitivity and selectivity in the simultaneous detection of aflatoxin M1 and aflatoxin B1. Talanta 2023, 265, 124908. [Google Scholar] [CrossRef]
- Uludag, Y.; Esen, E.; Kokturk, G.; Ozer, H.; Muhammad, T.; Olcer, Z.; Basegmez, H.I.O.; Simsek, S.; Barut, S.; Gok, M.Y.; et al. Lab-on-a-chip based biosensor for the real-time detection of aflatoxin. Talanta 2016, 160, 381–388. [Google Scholar] [CrossRef]
- Azri, F.; Sukor, R.; Selamat, J.; Bakar, F.A.; Yusof, N.; Hajian, R. Electrochemical immunosensor for detection of aflatoxin B1 based on indirect competitive ELISA. Toxins 2018, 10, 196. [Google Scholar] [CrossRef]
- Abera, B.D.; Falco, A.; Ibba, P.; Cantarella, G.; Petti, L.; Lugli, P. Development of flexible dispense-printed electrochemical immunosensor for aflatoxin M1 detection in milk. Sensors 2019, 19, 3912. [Google Scholar] [CrossRef]
- Ren, X.; Zhang, Q.; Wu, W.; Yan, T.; Tang, X.; Zhang, W.; Yu, L.; Li, P. Anti-idiotypic nanobody-phage display-mediated real-time immuno-PCR for sensitive, simultaneous and quantitative detection of total aflatoxins and zearalenone in grains. Food Chem. 2019, 297, 124912. [Google Scholar] [CrossRef]
- Hu, D.; Xiao, S.; Guo, Q.; Yue, R.; Geng, D.; Ji, D. Luminescence method for detection of aflatoxin B1 using ATP-releasing nucleotides. RSC Adv. 2021, 11, 24027–24031. [Google Scholar] [CrossRef]
- Zhang, S.; Li, Z.; An, J.; Yang, Y.; Tang, X. Identification of aflatoxin B1 in peanut using near-infrared spectroscopy combined with naive bayes classifier. Spectrosc. Lett. 2021, 54, 340–351. [Google Scholar] [CrossRef]
- Zhao, L.; Mao, J.; Hu, L.; Zhang, S.; Yang, X. Self-replicating catalyzed hairpin assembly for rapid aflatoxin B1 detection. Anal. Methods 2021, 13, 222–226. [Google Scholar] [CrossRef]
- Dai, S.; Xing, K.; Jiao, Y.; Yu, S.; Yang, X.; Yao, L.; Jia, P.; Cheng, Y.; Xu, Z. A novel magnetic resonance tuning-magnetic relaxation switching sensor based on Gd-MOF/USPIO assembly for sensitive and convenient aflatoxin B1 detection. Food Chem. 2024, 443, 138537. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Song, X.; Wang, S.; Liu, H.; Xiong, C.; Wang, S.; Zhang, X.; Chen, M.M. Portable dual-mode paper chips for highly sensitive and rapid determination of aflatoxin B1 via an aptamer-gated MOFs. Food Chem. 2024, 457, 140182. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Bai, Y.; Zhao, T.; Liang, M.; Hu, X.; Wang, D.; Tang, X.; Yu, L.; Zhang, Q.; Li, P.; et al. Intelligent electrochemical point-of-care test method with interface control based on DNA pyramids: Aflatoxin B1 detection in food and the environment. Foods 2023, 12, 4447. [Google Scholar] [CrossRef]
- Li, Y.; Li, Z.; Jia, B.; Tu, Z.; Zeng, J.; Pang, J.; Ren, W.; Huang, Z.; He, B.; Wang, Z. Detection of AFB1 by immunochromatographic test strips based on double-probe signal amplification with nanobody and biotin–streptavidin system. Foods 2024, 13, 3396. [Google Scholar] [CrossRef]
- Zhao, H.; Chen, X.; Shen, C.; Qu, B. Determination of 16 mycotoxins in vegetable oils using a QuEChERS method combined with high-performance liquid chromatography-tandem mass spectrometry. Food Addit. Contam. A 2016, 34, 255–264. [Google Scholar] [CrossRef]
- Sharmili, K.; Jinap, S.; Sukor, R. Development, optimization and validation of QuEChERS based liquid chromatography tandem mass spectrometry method for determination of multimycotoxin in vegetable oil. Food Control 2016, 70, 152–160. [Google Scholar] [CrossRef]
- Lijalem, Y.G.; Gab-Allah, M.A.; Yu, H.; Choi, K.; Kim, B. Development of isotope dilution-ultrahigh-performance liquid chromatography-tandem mass spectrometry for the accurate determination of aflatoxins in grains. J. Food Compost. Anal. 2024, 126, 105896. [Google Scholar] [CrossRef]
- Zhao, Y.; Huang, J.; Ma, L.; Wang, F. Development and validation of a simple and fast method for simultaneous determination of aflatoxin B1 and sterigmatocystin in grains. Food Chem. 2017, 221, 11–17. [Google Scholar] [CrossRef]
- Shuib, N.S.; Makahleh, A.; Salhimi, S.M.; Saad, B. Determination of aflatoxin M1 in milk and dairy products using high performance liquid chromatography-fluorescence with post column photochemical derivatization. J. Chromatogr. A 2017, 1510, 51–56. [Google Scholar] [CrossRef]
- Campone, L.; Piccinelli, A.L.; Celano, R.; Pagano, I.; Di Sanzo, R.; Carabetta, S.; Russo, M.; Rastrelli, L. Occurrence of aflatoxin M1 in milk samples from Italy analysed by online-SPE UHPLC-MS/MS. Nat. Prod. Res. 2018, 32, 1803–1808. [Google Scholar] [CrossRef]
- Paschoal, F.N.; De Azevedo Silva, D.; Von Sperling De Souza, R.; De Oliveira, M.S.; Pereira, D.A.A.; De Souza, S.V.C. A rapid single-extraction method for the simultaneous determination of aflatoxins B1, B2, G1, G2, fumonisin B1, and zearalenone in corn meal by ultra-performance liquid chromatography tandem mass spectrometry. Food Anal. Methods 2017, 10, 1631–1644. [Google Scholar] [CrossRef]
- Mao, J.; Zheng, N.; Wen, F.; Guo, L.; Fu, C.; Ouyang, H.; Zhong, L.; Wang, J.; Lei, S. Multi-mycotoxins analysis in raw milk by ultra-high performance liquid chromatography coupled to quadrupole orbitrap mass spectrometry. Food Control 2018, 84, 305–311. [Google Scholar] [CrossRef]
- Flores-Flores, M.E.; González-Peñas, E. An LC–MS/MS method for multi-mycotoxin quantification in cow milk. Food Chem. 2017, 218, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Sartori, A.V.; De Moraes, M.H.P.; Dos Santos, R.P.; Souza, Y.P.; Da Nóbrega, A.W. Determination of mycotoxins in cereal-based porridge destined for infant consumption by ultra-high performance liquid chromatography-tandem mass spectrometry. Food Anal. Methods 2017, 10, 4049–4061. [Google Scholar] [CrossRef]
- Huertas-Pérez, J.F.; Arroyo-Manzanares, N.; Hitzler, D.; Castro-Guerrero, F.G.; Gámiz-Gracia, L.; García-Campaña, A.M. Simple determination of aflatoxins in rice by ultra-high performance liquid chromatography coupled to chemical post-column derivatization and fluorescence detection. Food Chem. 2018, 245, 189–195. [Google Scholar] [CrossRef]
- Zareshahrabadi, Z.; Bahmyari, R.; Nouraei, H.; Khodadadi, H.; Mehryar, P.; Asadian, F.; Zomorodian, K. Detection of aflatoxin and ochratoxin A in spices by high-performance liquid chromatography. J. Food Qual. 2020, 2020, 8858889. [Google Scholar] [CrossRef]
- Algammal, A.M.; Elsayed, M.E.; Hashem, H.R.; Ramadan, H.; Sheraba, N.S.; El-Diasty, E.M.; Abbas, S.M.; Hetta, H.F. Molecular and HPLC-based approaches for detection of aflatoxin B1 and ochratoxin A released from toxigenic Aspergillus species in processed meat. BMC Microbiol. 2021, 21, 82. [Google Scholar] [CrossRef]
- Salisu, B.; Anua, S.; Isha, W.; Mazlan, N. Development and validation of quantitative thin layer chromatographic technique for determination of total aflatoxins in poultry feed and food grains without sample clean-up. J. Adv. Vet. Anim. Res. 2021, 8, 656–670. [Google Scholar] [CrossRef]
- Al-Ghouti, M.A.; AlHusaini, A.; Abu-Dieyeh, M.H.; Abd Elkhabeer, M.; Alam, M.M. Determination of aflatoxins in coffee by means of ultra-high performance liquid chromatography-fluorescence detector and fungi isolation. Int. J. Environ. Anal. Chem. 2022, 102, 6999–7014. [Google Scholar] [CrossRef]
- Smeesters, L. Fluorescence spectroscopy enhancing aflatoxin detection in solid food products: From laboratory setup towards handheld sensing units. Biophotonics Point Care III 2024, 13008, 1300802. [Google Scholar] [CrossRef]
- Jiang, H.; Zhao, Y.; Li, J.; Zhao, M.; Deng, J.; Bai, X. Quantitative detection of aflatoxin B1 in peanuts using Raman spectra and multivariate analysis methods. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2024, 316, 124322. [Google Scholar] [CrossRef] [PubMed]
- Yao, S.; Miyagusuku-Cruzado, G.; West, M.; Nwosu, V.; Dowd, E.; Fountain, J.; Giusti, M.M.; Rodriguez-Saona, L.E. Nondestructive and rapid screening of aflatoxin-contaminated single peanut kernels using field-portable spectroscopy instruments (FT-IR and Raman). Foods 2024, 13, 157. [Google Scholar] [CrossRef] [PubMed]
- Smeesters, L.; Meulebroeck, W.; Raeymaekers, S.; Thienpont, H. Optical detection of aflatoxins in maize using one- and two-photon induced fluorescence spectroscopy. Food Control 2015, 51, 408–416. [Google Scholar] [CrossRef]
- Magnus, I.; Abbasi, F.; Thienpont, H.; Smeesters, L. Laser-induced fluorescence spectroscopy enhancing pistachio nut quality screening. Food Control 2024, 158, 110192. [Google Scholar] [CrossRef]
- Durmuş, E.; Güneş, A.; Kalkan, H. Detection of aflatoxin and surface mould contaminated figs by using Fourier transform Near-Infrared Reflectance spectroscopy. J. Sci. Food Agric. 2017, 97, 317–323. [Google Scholar] [CrossRef]
- Lai, W.; Zeng, Q.; Tang, J.; Zhang, M.; Tang, D. A conventional chemical reaction for use in an unconventional assay: A colorimetric immunoassay for aflatoxin B1 by using enzyme-responsive just-in-time generation of a MnO2 based nanocatalyst. Microchim. Acta 2018, 185, 92. [Google Scholar] [CrossRef]
- Jaiswal, P.; Jha, S.N.; Kaur, J.; Borah, A.; Ramya, H.G. Detection of aflatoxin M1 in milk using spectroscopy and multivariate analyses. Food Chem. 2018, 238, 209–214. [Google Scholar] [CrossRef]
- Cheng, X.; Vella, A.; Stasiewicz, M.J. Classification of aflatoxin contaminated single corn kernels by ultraviolet to near infrared spectroscopy. Food Control 2019, 98, 253–261. [Google Scholar] [CrossRef]
- Shen, F.; Wu, Q.; Shao, X.; Zhang, Q. Non-destructive and rapid evaluation of aflatoxins in brown rice by using near-infrared and mid-infrared spectroscopic techniques. J. Food Sci. Technol. 2018, 55, 1175–1184. [Google Scholar] [CrossRef]
- Wu, Q.; Xu, H. Application of multiplexing fiber optic laser induced fluorescence spectroscopy for detection of aflatoxin B1 contaminated pistachio kernels. Food Chem. 2019, 290, 24–31. [Google Scholar] [CrossRef]
- Bertani, F.R.; Businaro, L.; Gambacorta, L.; Mencattini, A.; Brenda, D.; Di Giuseppe, D.; De Ninno, A.; Solfrizzo, M.; Martinelli, E.; Gerardino, A. Optical detection of aflatoxins B in grained almonds using fluorescence spectroscopy and machine learning algorithms. Food Control 2020, 112, 107073. [Google Scholar] [CrossRef]
- Putthang, R.; Sirisomboon, P.; Sirisomboon, C.D. Shortwave near-infrared spectroscopy for rapid detection of aflatoxin B1 contamination in polished rice. J. Food Prot. 2019, 82, 796–803. [Google Scholar] [CrossRef] [PubMed]
- Tao, F.; Yao, H.; Zhu, F.; Hruska, Z.; Liu, Y.; Rajasekaran, K.; Bhatnagar, D. A rapid and nondestructive method for simultaneous determination of aflatoxigenic fungus and aflatoxin contamination on corn kernels. J. Agric. Food Chem. 2019, 67, 5230–5239. [Google Scholar] [CrossRef]
- Liu, W.; Zhao, P.; Wu, C.; Liu, C.; Yang, J.; Zheng, L. Rapid determination of aflatoxin B1 concentration in soybean oil using terahertz spectroscopy with chemometric methods. Food Chem. 2019, 293, 213–219. [Google Scholar] [CrossRef]
- Wu, Q.; Xu, H. Design and development of an on-line fluorescence spectroscopy system for detection of aflatoxin in pistachio nuts. Postharvest Biol. Technol. 2020, 159, 111016. [Google Scholar] [CrossRef]
- Zhongzhi, H.; Limiao, D. Aflatoxin contaminated degree detection by hyperspectral data using band index. Food Chem. Toxicol. 2020, 137, 111159. [Google Scholar] [CrossRef]
- Zheng, S.-Y.; Wei, Z.-S.; Li, S.; Zhang, S.-J.; Xie, C.-F.; Yao, D.-S.; Liu, D.-L. Near-infrared reflectance spectroscopy-based fast versicolorin A detection in maize for early aflatoxin warning and safety sorting. Food Chem. 2020, 332, 127419. [Google Scholar] [CrossRef]
- Chen, M.; He, X.; Pang, Y.; Shen, F.; Fang, Y.; Hu, Q. Laser induced fluorescence spectroscopy for detection of aflatoxin B1 contamination in peanut oil. J. Food Meas. Charact. 2021, 15, 2231–2239. [Google Scholar] [CrossRef]
- Jha, S.N.; Jaiswal, P.; Kaur, J.; Ramya, H.G. Rapid detection and quantification of aflatoxin B1 in milk using Fourier transform infrared spectroscopy. J. Inst. Eng. A 2021, 102, 259–265. [Google Scholar] [CrossRef]
- Tang, W.; Qi, Y.; Li, Z. A portable, cost-effective and user-friendly instrument for colorimetric enzyme-linked immunosorbent assay and rapid detection of aflatoxin B1. Foods 2021, 10, 2483. [Google Scholar] [CrossRef]
- Chavez, R.A.; Cheng, X.; Herrman, T.J.; Stasiewicz, M.J. Single kernel aflatoxin and fumonisin contamination distribution and spectral classification in commercial corn. Food Control 2022, 131, 108393. [Google Scholar] [CrossRef]
- Bartolić, D.; Mutavdžić, D.; Carstensen, J.M.; Stanković, S.; Nikolić, M.; Krstović, S.; Radotić, K. Fluorescence spectroscopy and multispectral imaging for fingerprinting of aflatoxin-B1 contaminated (Zea mays L.) seeds: A preliminary study. Sci. Rep. 2022, 12, 4849. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; He, J.; Yao, W.; Jiang, H.; Chen, Q. Determination of aflatoxin B1 value in corn based on Fourier transform near-infrared spectroscopy: Comparison of optimization effect of characteristic wavelengths. LWT 2022, 164, 113657. [Google Scholar] [CrossRef]
- Smeesters, L.; Kuntzel, T.; Thienpont, H.; Guilbert, L. Handheld fluorescence spectrometer enabling sensitive aflatoxin detection in maize. Toxins 2023, 15, 361. [Google Scholar] [CrossRef]
- Lutz, É.; Carteri Coradi, P. Applications of new technologies for monitoring and predicting grains quality stored: Sensors, Internet of Things, and Artificial Intelligence. Measurement 2022, 188, 110609. [Google Scholar] [CrossRef]
- Zhang, W.; Dou, J.; Wu, Z.; Li, Q.; Wang, S.; Xu, H.; Wu, W.; Sun, C. Application of non-aflatoxigenic Aspergillus flavus for the biological control of aflatoxin contamination in China. Toxins 2022, 14, 681. [Google Scholar] [CrossRef]
- Nguyen, T.; Chen, X.; Ma, L.; Feng, Y. Mycotoxin biodegradation by Bacillus bacteria—A review. Toxins 2024, 16, 478. [Google Scholar] [CrossRef]
- Shu, X.; Wang, Y.; Zhou, Q.; Li, M.; Hu, H.; Ma, Y.; Chen, X.; Ni, J.; Zhao, W.; Huang, S.; et al. Biological degradation of aflatoxin B1 by cell-free extracts of Bacillus velezensis DY3108 with broad pH stability and excellent thermostability. Toxins 2018, 10, 330. [Google Scholar] [CrossRef]
- Xu, L.; Ahmed, M.F.E.; Sangare, L.; Zhao, Y.; Selvaraj, J.N.; Xing, F.; Wang, Y.; Yang, H.; Liu, Y. Novel aflatoxin-degrading enzyme from Bacillus shackletonii L7. Toxins 2017, 9, 36. [Google Scholar] [CrossRef]
- Zhang, L.Y.; Liu, S.; Zhao, X.J.; Wang, N.; Jiang, X.; Xin, H.S.; Zhang, Y.G. Lactobacillus rhamnosus GG modulates gastrointestinal absorption, excretion patterns, and toxicity in Holstein calves fed a single dose of aflatoxin B1. J. Dairy Sci. 2019, 102, 1330–1340. [Google Scholar] [CrossRef]
- Intanoo, M.; Kongkeitkajorn, M.B.; Suriyasathaporn, W.; Phasuk, Y.; Bernard, J.K.; Pattarajinda, V. Effect of supplemental Kluyveromyces marxianus and Pichia kudriavzevii on aflatoxin M1 excretion in milk of lactating dairy cows. Animals 2020, 10, 709. [Google Scholar] [CrossRef] [PubMed]
- Lopes, D.C.; Martins, J.H.; Lacerda Filho, A.F.; Melo, E.C.; Monteiro, P.M.B.; Queiroz, D.M. Aeration strategy for controlling grain storage based on simulation and on real data acquisition. Comput. Electron. Agric. 2008, 63, 140–146. [Google Scholar] [CrossRef]
- Ghanbari, R.; Aghaee, E.M.; Rezaie, S.; Khaniki, G.J.; Alimohammadi, M.; Soleimani, M.; Noorbakhsh, F. The inhibitory effect of lactic acid bacteria on aflatoxin production and expression of aflR gene in Aspergillus parasiticus. J. Food Saf. 2017, 38, e12413. [Google Scholar] [CrossRef]
- Siahmoshteh, F.; Hamidi-Esfahani, Z.; Spadaro, D.; Shams-Ghahfarokhi, M.; Razzaghi-Abyaneh, M. Unraveling the mode of antifungal action of Bacillus subtilis and Bacillus amyloliquefaciens as potential biocontrol agents against aflatoxigenic Aspergillus parasiticus. Food Control 2018, 89, 300–307. [Google Scholar] [CrossRef]
- Quattrini, M.; Bernardi, C.; Stuknytė, M.; Masotti, F.; Passera, A.; Ricci, G.; Vallone, L.; De Noni, I.; Brasca, M.; Fortina, M.G. Functional characterization of Lactobacillus plantarum ITEM 17215: A potential biocontrol agent of fungi with plant growth promoting traits, able to enhance the nutritional value of cereal products. Food Res. Int. 2018, 106, 936–944. [Google Scholar] [CrossRef]
- González Pereyra, M.L.; Martínez, M.P.; Petroselli, G.; Erra Balsells, R.; Cavaglieri, L.R. Antifungal and aflatoxin-reducing activity of extracellular compounds produced by soil Bacillus strains with potential application in agriculture. Food Control 2018, 85, 392–399. [Google Scholar] [CrossRef]
- Yang, M.; Lu, L.; Pang, J.; Hu, Y.; Guo, Q.; Li, Z.; Wu, S.; Liu, H.; Wang, C. Biocontrol activity of volatile organic compounds from Streptomyces alboflavus TD-1 against Aspergillus flavus growth and aflatoxin production. J. Microbiol. 2019, 57, 396–404. [Google Scholar] [CrossRef]
- Liu, Y.; Mao, H.; Hu, C.; Tron, T.; Lin, J.; Wang, J.; Sun, B. Molecular docking studies and in vitro degradation of four aflatoxins (AFB1, AFB2, AFG1, and AFG2) by a recombinant laccase from Saccharomyces cerevisiae. J. Food Sci. 2020, 85, 1353–1360. [Google Scholar] [CrossRef]
- Fouad, M.; El-desouky, T.A. Anti-toxigenic effect of lactic acid bacteria against Aspergillus spp. isolated from wheat grains. Open Microbiol. J. 2020, 14, 252–259. [Google Scholar] [CrossRef]
- da Cruz, P.O.; de Matos, C.J.; Nascimento, Y.M.; Tavares, J.F.; de Souza, E.L.; Magalhães, H.I.F. Efficacy of potentially probiotic fruit-derived Lactobacillus fermentum, L. paracasei and L. plantarum to remove aflatoxin M1 in vitro. Toxins 2021, 13, 4. [Google Scholar] [CrossRef]
- Rämö, S.; Kahal, M.; Joutsjoki, V. Aflatoxin B1 binding by lactic acid bacteria in protein-rich plant material fermentation. Appl. Sci. 2022, 12, 12769. [Google Scholar] [CrossRef]
- Kumar, V.; Bahuguna, A.; Ramalingam, S.; Lee, J.S.; Han, S.S.; Chun, H.S.; Kim, M. Aflatoxin reduction and retardation of aflatoxin production by microorganisms in doenjang during a one-year fermentation. J. Fungi 2022, 8, 190. [Google Scholar] [CrossRef]
- Chen, G.; Fang, Q.; Liao, Z.; Xu, C.; Liang, Z.; Liu, T.; Zhong, Q.; Wang, L.; Fang, X.; Wang, J. Detoxification of aflatoxin B1 by a potential probiotic Bacillus amyloliquefaciens WF2020. Front. Microbiol. 2022, 13, 891091. [Google Scholar] [CrossRef]
- Papp, D.A.; Kocsubé, S.; Farkas, Z.; Szekeres, A.; Vágvölgyi, C.; Hamari, Z.; Varga, M. Aflatoxin B1 control by various Pseudomonas isolates. Toxins 2024, 16, 367. [Google Scholar] [CrossRef]
- Patras, A.; Julakanti, S.; Yannam, S.R.R.; Bansode, M.R.; Burns, M.; Vergne, M.J. Effect of UV irradiation on aflatoxin reduction: A cytotoxicity evaluation study using human hepatoma cell line. Mycotoxin Res. 2017, 33, 343–350. [Google Scholar] [CrossRef]
- Domijan, A.M.; Čermak, A.M.M.; Vulić, A.; Bujak, I.T.; Pavičić, I.; Pleadin, J.; Markov, K.; Mihaljević, B. Cytotoxicity of gamma irradiated aflatoxin B1 and ochratoxin A. J. Environ. Sci. Health B 2019, 54, 155–162. [Google Scholar] [CrossRef]
- Ferreira, C.D.; Lang, G.H.; Da Silva Lindemann, I.; Da Silva Timm, N.; Hoffmann, J.F.; Ziegler, V.; De Oliveira, M. Postharvest UV-C irradiation for fungal control and reduction of mycotoxins in brown, black, and red rice during long-term storage. Food Chem. 2021, 339, 127810. [Google Scholar] [CrossRef]
- Hojnik, N.; Modic, M.; Walsh, J.L.; Žigon, D.; Javornik, U.; Plavec, J.; Žegura, B.; Filipič, M.; Cvelbar, U. Unravelling the pathways of air plasma induced aflatoxin B1 degradation and detoxification. J. Hazard. Mater. 2021, 403, 123593. [Google Scholar] [CrossRef]
- Rushing, B.R.; Selim, M.I. Aflatoxin B1: A review on metabolism, toxicity, occurrence in food, occupational exposure, and detoxification methods. Food Chem. Toxicol. 2019, 124, 81–100. [Google Scholar] [CrossRef]
- Lorán, S.; Carramiñana, J.J.; Juan, T.; Ariño, A.; Herrera, M. Inhibition of Aspergillus parasiticus growth and aflatoxins production by natural essential oils and phenolic acids. Toxins 2022, 14, 384. [Google Scholar] [CrossRef]
- Zheng, N.; Zhang, H.; Li, S.; Wang, J.; Liu, J.; Ren, H.; Gao, Y. Lactoferrin inhibits aflatoxin B1- and aflatoxin M1-induced cytotoxicity and DNA damage in Caco-2, HEK, Hep-G2, and SK-N-SH cells. Toxicon 2018, 150, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Akbari, D.M.; Kordbacheh, P.; Daei Ghazvini, R.; Moazeni, M.; Nazemi, L.; Rezaie, S. Inhibitory effect of vitamin C on Aspergillus parasiticus growth and aflatoxin gene expression. Curr. Med. Mycol. 2018, 4, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Mahbobinejhad, Z.; Aminian, H.; Ebrahimi, L.; Vahdati, K. Reduction of aflatoxin production by exposing Aspergillus flavus to CO2. J. Crop Prot. 2019, 8, 441–448. [Google Scholar]
- Li, H.; Li, S.; Yang, H.; Wang, Y.; Wang, J.; Zheng, N. L-proline alleviates kidney injury caused by AFB1 and AFM1 through regulating excessive apoptosis of kidney cells. Toxins 2019, 11, 226. [Google Scholar] [CrossRef]
- Yu, Y.; Shi, J.; Xie, B.; He, Y.; Qin, Y.; Wang, D.; Shi, H.; Ke, Y.; Sun, Q. Detoxification of aflatoxin B1 in corn by chlorine dioxide gas. Food Chem. 2020, 328, 127121. [Google Scholar] [CrossRef]
- Al-Kuhla, A.A.M.; Ibraheem, A.M. Protective effects of rhino-hepato forte in broiler chickens during aflatoxicosis. Egypt. J. Vet. Sci. 2021, 52, 203–212. [Google Scholar] [CrossRef]
- Wu, G.; San, J.; Pang, H.; Du, Y.; Li, W.; Zhou, X.; Yang, X.; Hu, J.; Yang, J. Taurine attenuates AFB1-induced liver injury by alleviating oxidative stress and regulating mitochondria-mediated apoptosis. Toxicon 2022, 215, 17–27. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, J.; Chang, Z.; Li, S.; Zhang, Z.; Liu, S.; Wang, S.; Wei, L.; Lv, Q.; Ding, K.; et al. SeMet alleviates AFB1-induced oxidative stress and apoptosis in rabbit kidney by regulating Nrf2/Keap1/NQO1 and PI3K/AKT signaling pathways. Ecotoxicol. Environ. Saf. 2024, 269, 115742. [Google Scholar] [CrossRef]
- Yang, D.; Zhang, S.; Cao, H.; Wu, H.; Liang, Y.; Teng, C.B.; Yu, H.F. Detoxification of aflatoxin B1 by phytochemicals in agriculture and food science. J. Agric. Food Chem. 2024, 72, 14481–14497. [Google Scholar] [CrossRef]
- Haque, M.A.; Wang, Y.; Shen, Z.; Li, X.; Saleemi, M.K.; He, C. Mycotoxin contamination and control strategy in human, domestic animal and poultry: A review. Microb. Pathog. 2020, 142, 104095. [Google Scholar] [CrossRef]
- Silva, B.A.; Ferreres, F.; Malva, J.O.; Dias, A.C.P. Phytochemical and antioxidant characterization of Hypericum perforatum alcoholic extracts. Food Chem. 2005, 90, 157–167. [Google Scholar] [CrossRef]
- Brborić, J.; Klisić, A.; Kotur-Stevuljević, J.; Delogu, G.; Gjorgieva Ackova, D.; Kostić, K.; Dettori, M.A.; Fabbri, D.; Carta, P.; Saso, L. Natural and natural-like polyphenol compounds: In vitro antioxidant activity and potential for therapeutic application. Arch. Med. Sci. 2021, 19, 651–671. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Razek, A.G.; Badr, A.N.; Alharthi, S.S.; Selim, K.A. Efficacy of bottle gourd seeds’ extracts in chemical hazard reduction secreted as toxigenic fungi metabolites. Toxins 2021, 13, 789. [Google Scholar] [CrossRef] [PubMed]
- Ali, D.; Abdel-Rahman, T.; Abo-Hagger, A.; Ahmed, M. In vitro and in vivo assessment of banana peel powder as an aflatoxins biosorbent. Egypt. J. Bot. 2019, 59, 399–411. [Google Scholar] [CrossRef]
- Badr, A.N.; Shehata, M.G.; Abdel-Razek, A.G. Antioxidant activities and potential impacts to reduce aflatoxins utilizing jojoba and jatropha oils and extracts. Int. J. Pharmacol. 2017, 13, 1103–1114. [Google Scholar] [CrossRef]
- Abdel-Latif, M.S.; Elmeleigy, K.M.; Aly, T.A.A.; Khattab, M.S.; Mohamed, S.M. Pathological and biochemical evaluation of coumarin and chlorophyllin against aflatoxicosis in rat. Exp. Toxicol. Pathol. 2017, 69, 285–291. [Google Scholar] [CrossRef]
- Vipin, A.V.; Raksha Rao, K.; Kurrey, N.K.; Appaiah, K.A.; Venkateswaran, G. Protective effects of phenolics-rich extract of ginger against aflatoxin B1-induced oxidative stress and hepatotoxicity. Biomed. Pharmacother. 2017, 91, 415–424. [Google Scholar] [CrossRef]
- Badr, A.N.; Naeem, M.A. Protective efficacy using cape-golden berry against pre-carcinogenic aflatoxins induced in rats. Toxicol. Rep. 2019, 6, 607–615. [Google Scholar] [CrossRef]
- Abdel-Sattar, W.M.; Sadek, K.M.; Elbestawy, A.R.; Mourad, D.M. The protective role of date palm (Phoenix dactylifera seeds) against aflatoxicosis in broiler chickens regarding carcass characteristics, hepatic and renal biochemical function tests and histopathology. J. World’s Poult. Res. 2019, 9, 59–69. [Google Scholar] [CrossRef]
- El-Mekkawy, H.I.; Al-Kahtani, M.A.; Shati, A.A.; Alshehri, M.A.; Al-Doaiss, A.A.; Elmansi, A.A.; Ahmed, A.E. Black tea and curcumin synergistically mitigate the hepatotoxicity and nephropathic changes induced by chronic exposure to aflatoxin-B1 in Sprague-Dawley rats. J. Food Biochem. 2020, 44, e13346. [Google Scholar] [CrossRef]
- Singto, T.; Tassaneeyakul, W.; Porasuphatana, S. Protective effects of purple waxy corn on aflatoxin B1-induced oxidative stress and micronucleus in HepG2 cells. Indian J. Pharm. Sci. 2020, 82, 506–513. [Google Scholar] [CrossRef]
- Jin, S.; Yang, H.; Jiao, Y.; Pang, Q.; Wang, Y.; Wang, M.; Shan, A.; Feng, X. Dietary curcumin alleviated acute ileum damage of ducks (Anas platyrhynchos) induced by AFB1 through regulating Nrf2-ARE and NF-κB signaling pathways. Foods 2021, 10, 1370. [Google Scholar] [CrossRef] [PubMed]
- El-Sheshtawy, S.M.; El-Zoghby, A.F.; Shawky, N.A.; Samak, D.H. Aflatoxicosis in Pekin duckling and the effects of treatments with lycopene and silymarin. Vet. World 2021, 14, 788–793. [Google Scholar] [CrossRef]
- Ibrahim, E.A.; Yousef, M.I.; Ghareeb, D.A.; Augustyniak, M.; Giesy, J.P.; Aboul-Soud, M.A.M.; El Wakil, A. Artichoke leaf extract-mediated neuroprotection against effects of aflatoxin in male rats. BioMed Res. Int. 2022, 2022, 4421828. [Google Scholar] [CrossRef]
- Wan, F.; Tang, L.; Rao, G.; Zhong, G.; Jiang, X.; Wu, S.; Huang, R.; Tang, Z.; Ruan, Z.; Chen, Z.; et al. Curcumin activates the Nrf2 Pathway to alleviate AFB1-induced immunosuppression in the spleen of ducklings. Toxicon 2022, 209, 18–27. [Google Scholar] [CrossRef]
- Khattab, M.S.; Aly, T.A.A.; Mohamed, S.M.; Naguib, A.M.M.; Al-Farga, A.; Abdel-Rahim, E.A. Hordeum vulgare L. microgreen mitigates reproductive dysfunction and oxidative stress in streptozotocin-induced diabetes and aflatoxicosis in male rats. Food Sci. Nutr. 2022, 10, 3355–3367. [Google Scholar] [CrossRef]
- Wang, L.; He, K.; Wang, X.; Wang, Q.; Quan, H.; Wang, P.; Xu, X. Recent progress in visual methods for aflatoxin detection. Crit. Rev. Food Sci. Nutr. 2022, 62, 7849–7865. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, F.; Liu, M.; Zhou, X.; Wang, M.; Cao, K.; Jin, S.; Shan, A.; Feng, X. Curcumin mitigates aflatoxin B1-induced liver injury via regulating the NLRP3 inflammasome and Nrf2 signaling pathway. Food Chem. Toxicol. 2022, 161, 112823. [Google Scholar] [CrossRef]
- Pauletto, M.; Giantin, M.; Tolosi, R.; Bassan, I.; Bardhi, A.; Barbarossa, A.; Montanucci, L.; Zaghini, A.; Dacasto, M. Discovering the protective effects of quercetin on aflatoxin B1-induced toxicity in bovine foetal hepatocyte-derived cells (BFH12). Toxins 2023, 15, 555. [Google Scholar] [CrossRef]
- Dazuk, V.; Tarasconi, L.; Molosse, V.L.; Cécere, B.G.O.; Deolindo, G.L.; Strapazzon, J.V.; Bottari, N.B.; Bissacotti, B.F.; Schetinger, M.R.C.; Sareta, L.; et al. Can the inclusion of a vegetable biocholine additive in pig feed contaminated with aflatoxin reduce toxicological impacts on animal health and performance? Animals 2023, 13, 3010. [Google Scholar] [CrossRef]
- Hassan, M.A.; Abo-Elmaaty, A.M.A.; Zaglool, A.W.; Mohamed, S.A.M.; Abou-Zeid, S.M.; Farag, M.R.; Alagawany, M.; Di Cerbo, A.; Azzam, M.M.; Alhotan, R.; et al. Origanum vulgare essential oil modulates the AFB1-induced oxidative damages, nephropathy, and altered inflammatory responses in growing rabbits. Toxins 2023, 15, 69. [Google Scholar] [CrossRef] [PubMed]
- Altyar, A.E.; Kensara, O.A.; Sayed, A.A.; Aleya, L.; Almutairi, M.H.; Zaazouee, M.S.; Elshanbary, A.A.; El-Demerdash, F.M.; Abdel-Daim, M.M. Acute aflatoxin B1-induced hepatic and cardiac oxidative damage in rats: Ameliorative effects of morin. Heliyon 2023, 9, e21837. [Google Scholar] [CrossRef] [PubMed]
- Velazhahan, R.; Al-Sadi, A.M.; Waly, M.I.; Soundra Pandian, S.B.; Al-Sabahi, J.; Al-Farsi, K. Aflatoxin B1 detoxification and antioxidant effect of selected Omani medicinal plants against aflatoxin B1-induced oxidative stress pathogenesis in the mouse liver. Appl. Sci. 2024, 14, 5378. [Google Scholar] [CrossRef]
- Khalid, S.A.; Elokle, A.A. Evaluation of pumpkin seed oil and its chitosan-Arabic gum nanoparticles on the viability of Aspergillus flavus and inhibition of total aflatoxin in beef sausage. Food Control 2025, 168, 110854. [Google Scholar] [CrossRef]
- Pascale, M.; Logrieco, A.F.; Graeber, M.; Hirschberger, M.; Reichel, M.; Lippolis, V.; De Girolamo, A.; Lattanzio, V.M.T.; Slettengren, K. Aflatoxin reduction in maize by industrial-scale cleaning solutions. Toxins 2020, 12, 331. [Google Scholar] [CrossRef]
- Sipos, P.; Peles, F.; Brassó, D.L.; Béri, B.; Pusztahelyi, T.; Pócsi, I.; Győri, Z. Physical and chemical methods for reduction in aflatoxin content of feed and food. Toxins 2021, 13, 204. [Google Scholar] [CrossRef]
- Rastegar, H.; Shoeibi, S.; Yazdanpanah, H.; Amirahmadi, M.; Khaneghah, A.M.; Campagnollo, F.B.; Sant’Ana, A.S. Removal of aflatoxin B1 by roasting with lemon juice and/or citric acid in contaminated pistachio nuts. Food Control 2017, 71, 279–284. [Google Scholar] [CrossRef]
- Shen, M.-H.; Singh, R.K. Effect of rotating peanuts on aflatoxin detoxification by ultraviolet C light and irradiation uniformity evaluated by AgCl-based bosimeter. Food Control 2021, 120, 107533. [Google Scholar] [CrossRef]
- Jubeen, F.; Sher, F.; Hazafa, A.; Zafar, F.; Ameen, M.; Rasheed, T. Evaluation and detoxification of aflatoxins in ground and tree nuts using food grade organic acids. Biocatal. Agric. Biotechnol. 2020, 29, 101749. [Google Scholar] [CrossRef]
- Pereyra, C.M.; Gil, S.; Cristofolini, A.; Bonci, M.; Makita, M.; Monge, M.P.; Montenegro, M.A.; Cavaglieri, L.R. The production of yeast cell wall using an agroindustrial waste influences the wall thickness and is implicated on the aflatoxin B1 adsorption process. Food Res. Int. 2018, 111, 306–313. [Google Scholar] [CrossRef]
- Yiannikouris, A.; Apajalahti, J.; Siikanen, O.; Dillon, G.P.; Moran, C.A. Saccharomyces cerevisiae cell wall-based adsorbent reduces aflatoxin B1 absorption in rats. Toxins 2021, 13, 209. [Google Scholar] [CrossRef] [PubMed]
- Čolović, R.; Puvača, N.; Cheli, F.; Avantaggiato, G.; Greco, D.; Đuragić, O.; Kos, J.; Pinotti, L. Decontamination of mycotoxin-contaminated feedstuffs and compound feed. Toxins 2019, 11, 617. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-Y.; Herrera-Balandrano, D.D.; Shi, X.-C.; Chen, X.; Liu, F.-Q.; Laborda, P. Occurrence of aflatoxins in water and decontamination strategies: A review. Water Res. 2023, 232, 119703. [Google Scholar] [CrossRef] [PubMed]
- Mata, A.T.; Ferreira, J.P.; Oliveira, B.R.; Batoréu, M.C.; Barreto Crespo, M.T.; Pereira, V.J.; Bronze, M.R. Bottled water: Analysis of mycotoxins by LC–MS/MS. Food Chem. 2015, 176, 455–464. [Google Scholar] [CrossRef]
- Oliveira, B.R.; Mata, A.T.; Ferreira, J.P.; Barreto Crespo, M.T.; Pereira, V.J.; Bronze, M.R. Production of mycotoxins by filamentous fungi in untreated surface water. Environ. Sci. Pollut. Res. 2018, 25, 17519–17528. [Google Scholar] [CrossRef]
- Picardo, M.; Sanchís, J.; Núñez, O.; Farré, M. Suspect screening of natural toxins in surface and drinking water by high performance liquid chromatography and high-resolution mass spectrometry. Chemosphere 2020, 261, 127888. [Google Scholar] [CrossRef]
- He, K.; Quan, H.; Wang, L.; Zhang, J.; Wang, H.; Zhu, X.; Xu, X. Engineering cotton fibers with zirconium metal-organic frameworks as a recyclable material for sensing and removing aflatoxins in water. Sens. Actuators B Chem. 2023, 397, 134673. [Google Scholar] [CrossRef]
- Stanley, J.; Patras, A.; Pendyala, B.; Bansode, R.R. Performance of a UV-A LED system for degradation of aflatoxins B1 and M1 in pure water: Kinetics and cytotoxicity study. Sci. Rep. 2020, 10, 13473. [Google Scholar] [CrossRef]
- Chen, J.; Shi, H.; Gong, M.; Chen, H.; Teng, L.; Xu, P.; Wang, Y.; Hu, Z.; Zeng, Z. β-Lactoglobulin-based aerogels: Facile preparation and sustainable removal of organic contaminants from water. Int. J. Biol. Macromol. 2024, 272, 132856. [Google Scholar] [CrossRef]
- Siri-Anusornsak, W.; Kolawole, O.; Soiklom, S.; Petchpoung, K.; Keawkim, K.; Chuaysrinule, C.; Maneeboon, T. Innovative use of Spirogyra spp. biomass for the sustainable adsorption of aflatoxin B1 and ochratoxin A in aqueous solutions. Molecules 2024, 29, 5038. [Google Scholar] [CrossRef]
- FAO; OIE; WHO. Taking a Multisectoral One Health Approach: A Tripartite Guide to Addressing Zoonotic Diseases in Countries; Food and Agriculture Organization of the United Nations; World Organization for Animal Health; World Health Organization: Rome, Italy, 2019. [Google Scholar]
- Humboldt-Dachroeden, S.; Mantovani, A. Assessing Environmental Factors within the One Health Approach. Medicina 2021, 57, 240. [Google Scholar] [CrossRef] [PubMed]
- Pettan-Brewer, C.; Martins, A.F.; de Abreu, D.P.B.; Brandão, A.P.D.; Barbosa, D.S.; Figueroa, D.P.; Cediel, N.; Kahn, L.H.; Brandespim, D.F.; Velásquez, J.C.C.; et al. From the Approach to the Concept: One Health in Latin America—Experiences and Perspectives in Brazil, Chile, and Colombia. Front. Public Health 2021, 9, 687110. [Google Scholar] [CrossRef] [PubMed]
- Mor, N. Organising for One Health in a developing country. One Health 2023, 17, 100611. [Google Scholar] [CrossRef] [PubMed]
- Kumar, I.; Rawat, J.; Mohd, N.; Husain, S. Opportunities of Artificial Intelligence and Machine Learning in the Food Industry. J. Food Qual. 2021, 2021, 4535567. [Google Scholar] [CrossRef]
- Kim, Y.K.; Qin, J.; Baek, I.; Lee, K.M.; Kim, S.Y.; Kim, S.; Chan, D.; Herrman, T.J.; Kim, N.; Kim, M.S. Detection of aflatoxins in ground maize using a compact and automated Raman spectroscopy system with machine learning. Curr. Res. Food Sci. 2023, 7, 100647. [Google Scholar] [CrossRef]
- Sandlin, N.; Russell Kish, D.; Kim, J.; Zaccaria, M.; Momeni, B. Current and emerging tools of computational biology to improve the detoxification of mycotoxins. Appl. Environ. Microbiol. 2022, 88, e0210221. [Google Scholar] [CrossRef]
- Opoku, B.; Osekre, E.A.; Opit, G.; Bosomtwe, A.; Bingham, G.V. Evaluation of Hermetic Storage Bags for the Preservation of Yellow Maize in Poultry Farms in Dormaa Ahenkro, Ghana. Insects 2023, 14, 141. [Google Scholar] [CrossRef]
- Kumar, D.; Kalita, P. Reducing postharvest losses during storage of grain crops to strengthen food security in developing countries. Foods 2017, 6, 8. [Google Scholar] [CrossRef]
- Drago, E.; Campardelli, R.; Pettinato, M.; Perego, P. Innovations in smart packaging concepts for food: An extensive review. Foods 2020, 9, 1628. [Google Scholar] [CrossRef]
- Moreno, M.A.; Vallejo, A.M.; Ballester, A.R.; Zampini, C.; Isla, M.I.; López-Rubio, A.; Fabra, M.J. Antifungal edible coatings containing Argentinian propolis extract and their application in raspberries. Food Hydrocoll. 2020, 107, 105973. [Google Scholar] [CrossRef]
- Escamilla-García, M.; Calderón-Domínguez, G.; Chanona-Pérez, J.J.; Mendoza-Madrigal, A.G.; Di Pierro, P.; García-Almendárez, B.E.; Amaro-Reyes, A.; Regalado-González, C. Physical, structural, barrier, and antifungal characterization of chitosan-zein edible films with added essential oils. Int. J. Mol. Sci. 2017, 18, 12370. [Google Scholar] [CrossRef] [PubMed]
- Luz, C.; Calpe, J.; Saladino, F.; Luciano, F.B.; Fernandez-Franzon, M.; Manes, J.; Meca, G. Antimicrobial packaging based on E-polylysine bioactive film for the control of mycotoxigenic fungi in vitro and in bread. J. Food Process. Preserv. 2018, 42, e13370. [Google Scholar] [CrossRef] [PubMed]
- Kazemian-Bazkiaee, F.; Ebrahimi, A.; Hosseini, S.M.; Shojaee-Aliabadi, S.; Farhoodi, M.; Rahmatzadeh, B.; Sheikhi, Z. Evaluating the protective effect of edible coatings on lipid oxidation, fatty acid composition, and aflatoxins levels of roasted peanut kernels. Food Meas. 2020, 14, 1025–1038. [Google Scholar] [CrossRef]
- El-Sayed, H.S.; El-Sayed, S.M.; Mabrouk, A.M.M.; Nawwar, G.A.; Youssef, A.M. Development of eco-friendly probiotic edible coatings based on chitosan, alginate and carboxymethyl cellulose for improving the shelf life of UF soft cheese. J. Polym. Environ. 2021, 29, 1941–1953. [Google Scholar] [CrossRef]
- Gomes, A.S.d.L.P.B.; Weber, S.H.; Luciano, F.B. Resistance of transgenic maize cultivars to mycotoxin production—systematic review and meta-analysis. Toxins 2024, 16, 373. [Google Scholar] [CrossRef]
- Rajasekaran, K.; Sayler, R.J.; Sickler, C.M.; Majumdar, R.; Jaynes, J.M.; Cary, J.W. Control of Aspergillus flavus growth and aflatoxin production in transgenic maize kernels expressing a tachyplesin-derived synthetic peptide, AGM182. Plant Sci. 2018, 270, 150–156. [Google Scholar] [CrossRef]
- Thakare, D.; Zhang, J.; Wing, R.A.; Cotty, P.J.; Schmidt, M.A. Aflatoxin-free transgenic maize using host-induced gene silencing. Sci. Adv. 2017, 3, e1602382. [Google Scholar] [CrossRef]
- Raruang, Y.; Omolehin, O.; Hu, D.; Wei, Q.; Promyou, S.; Parekattil, L.J.; Rajasekaran, K.; Cary, J.W.; Wang, K.; Chen, Z.-Y. Targeting the Aspergillus flavus p2c gene through host-induced gene silencing reduces A. flavus infection and aflatoxin contamination in transgenic maize. Front. Plant Sci. 2023, 14, 1150086. [Google Scholar] [CrossRef]


| AF Level [μg/kg] | Class of Animal | Commodities |
|---|---|---|
| 20 | Dairy animals, animals not specified in other categories, or animals with unknown use | For corn, peanut products, cottonseed meal, and other animal feeds and feed ingredients |
| 20 | Immature animals | For corn, peanut products, and other animal feeds and feed ingredients, excluding cottonseed meal |
| 20 | Pets of all ages (e.g., dogs, cats, rabbits) | For corn, peanut products, cottonseed meal, other food ingredients, and complete pet food |
| 100 | Breeding cattle, breeding swine, and mature poultry (e.g., laying hens) | For corn and peanut products |
| 200 | Finishing swine (weighing 100 pounds or more) | For corn and peanut products |
| 300 | Beef cattle, swine, and poultry (regardless of age or breeding status) | For cotton seed meal |
| 300 | Finishing beef cattle (e.g., feedlot cattle) | For corn and peanut products |
| Feedstuff | AB1 Level [μg/kg] | |
|---|---|---|
| Raw materials | Maize processing products, peanut meal | ≤50 |
| Vegetable oil | ≤10 | |
| Maize oil, peanut oil | ≤20 | |
| Other plant-based feed materials | ≤30 | |
| Products | Concentrated feed for piglets and young birds | ≤10 |
| Concentrated feed for meat ducks, growing ducks, and ducks for egg production | ≤15 | |
| Other concentrated feed | ≤20 | |
| Calf and lamb concentrate supplement | ≤20 | |
| Concentrate supplement for lactation | ≤10 | |
| Other concentrate supplements | ≤30 | |
| Compound feed for piglets and young birds | ≤10 | |
| Compound feed for meat ducks, growing ducks, and laying ducks | ≤15 | |
| Other formula feed | ≤20 | |
| Method | Amplified Gene | Detected Strain | Limit of Detection | References |
|---|---|---|---|---|
| Polymerase chain reaction | aflP | Aflatoxigenic strain | Not calculated | [31] |
| Real-time polymerase chain reaction | aflD, aflM (ver), aflP, aflQ | Aflatoxigenic strain | Not calculated | [33] |
| Polymerase chain reaction and real-time polymerase chain reaction | aflR, aflM (ver), aflD (nor) | Aflatoxigenic strain | Not calculated | [34] |
| Polymerase chain reaction | afM (ver), aflJ, afIR, afID (nor) | Aflatoxigenic strain | Not calculated | [30] |
| Loop-mediated isothermal amplification and real-time polymerase chain reaction | aflT | Aflatoxigenic strain | 100-999 picograms of DNA (loop-mediated isothermal amplification) 160 femtograms of DNA (qPCR) | [32] |
| Method | Detection Matrix | Detection Method | Detected Aflatoxin | Limit of Detection for AFs [μg/kg] | References |
|---|---|---|---|---|---|
| Fluorescent aptamer | Fluorescently labeled aptamers bind aflatoxins, blocking DNA nanostructures and quenching fluorescence | Fluorescence | AFB1, AFM1 | 0.009 0.00624 | [56] |
| MiSens biosensor chip | MiSens silicone dioxide biochip consisting of 2 sets of Au electrode arrays with 3 working electrodes each, surface modified with protein A–aflatoxin antibody conjugate | MiSens and HPLC (high-performance liquid chromatography) | AFB1 | 390 | [57] |
| Electrochemical immunosensor using modified MWCNTs/CS/SPCE | Multi-walled carbon nanotubes/chitosan/screen-printed carbon electrode | Differential pulse voltammetry | AFB1 | 0.0003 | [58] |
| Flexible, dispense-printed electrochemical biosensors | Dispense-printed electrodes, which are functionalized with single-walled carbon nanotubes and subsequently coated with specific antibodies | Chronoamperometric | AFM1 | 0.02 | [59] |
| Anti-idiotypic nanobody proximity-dependent immuno-polymerase chain reaction (PD-IPCR) | Two phages displaying the variable domain of the heavy chain anti-idiotypic nanobody that binds to an aflatoxin- or zearalenone-specific monoclonal antibody | Real-time quantitative polymerase chain reaction | Total AFs | 0.03 | [60] |
| Luminescence method using ATP-releasing nucleotides | Magnetic bead aptamer complex | Luminescence | AFB1 | 0.000009 | [61] |
| Hyperbranched gold plasmonic blackbody- enhanced immunochromatographic test strip | Hyperbranched gold plasmonic blackbody | Optical density of test and control line | AFB1 | 0.016 | [62] |
| Self-replicating catalyzed hairpin assembly | Hairpin auxiliary probes, H1 and H2 | Fluorescence | AFB1 | 0.13 | [63] |
| Gd-MOF/USPIO magnetic field sensor | Gadolinium-based metal–organic framework (Gd-MOF) and ultrasmall superparamagnetic iron oxide (USPIO) | Magnetic resonance | AFB1 | 0.00054 | [64] |
| Method | Detector | Detected Aflatoxins | Limit of Detection for AFs [μg/kg] | References |
|---|---|---|---|---|
| High-performance liquid chromatography | Q-Trap 5500 LC-MS/MS system with a turbo ion spray source | AFB1, AFB2, AFG1 | 0.04–0.05 | [68] |
| Liquid chromatography | TSQ Quantum Discovery system: a high-performance triple-stage quadrupole mass spectrometer with electrospray ionization | AFB1, AFG1, AFG2 | 0.04 | [69] |
| Liquid chromatography–tandem mass spectrometry | Triple-quadrupole mass spectrometer with an electrospray ionization source | AFB1 | 0.03 | [71] |
| High-performance liquid chromatography | Fluorescence detector | AFM1 | 0.002 | [72] |
| Online solid phase extraction coupled with ultra-high performance liquid chromatography | Triple-quadrupole mass spectrometer | AFM1 | 0.0007 | [73] |
| Ultrahigh-performance liquid chromatography | Triple-quadrupole electrospray ionization mass spectrometer | AFB1, AFB2, AFG1, AFG2 | 0.25–0.32 | [74] |
| Ultrahigh-performance liquid chromatography | Quadrupole orbitrap mass spectrometer | AFB1, AFB2, AFG1, AFG2, AFM1 | 0.0003–0.0080 | [75] |
| Liquid chromatography | Triple-quadrupole mass spectrometer with an electrospray ionization source | AFB1, AFB2, AFG1, AFG2, AFM1 | 0.02–10.14 | [76] |
| Ultrahigh-performance liquid chromatography | Tandem-quadrupole mass spectrometer | AFB1, AFB2, AFG1, AFG2, AFM1, AFM2 | 0.14 | [77] |
| Ultrahigh-performance liquid chromatography | Fluorescence detector | AFB1, AFB2, AFG1, AFG2 | 0.07 0.08 0.06 0.09 | [78] |
| High-performance liquid chromatography | Fluorescence detector | AFB1, AFB2, AFG1, AFG2, | 0.1 | [79] |
| High-performance liquid chromatography | Fluorescence detector | AFB1 | 16.5 | [80] |
| Quantitative thin-layer chromatography | Thin layer chromatography scanner (fluorescence) | AFB1, AFB2, AFG1, AFG2 | <2.0 | [81] |
| Ultrahigh-performance liquid chromatography | Fluorescence detector | AFB1, AFB2, AFG1, AFG2 | 2 | [82] |
| Method | Detected Aflatoxin | LOD for AFs [μg/kg] | Ref. |
|---|---|---|---|
| Fluorescence spectroscopy and multispectral imaging | AFB1 | Not calculated | [86] |
| Laser-induced fluorescence spectroscopy | Aflatoxins | Not calculated | [87] |
| Fourier transform near-infrared reflectance spectroscopy | AFB1, AFB2, AFG1, AFG2 | <4.0 | [88] |
| Colorimetric competitive enzyme immunoassay | AFB1 | 0.1 | [89] |
| Attenuated total reflectance–Fourier transform infrared (ATR-FTIR) spectrometry | AFM1 | 0.02 | [90] |
| Custom-built ultraviolet-visible–near infrared spectroscopy system (UV-vis–NIR) | Aflatoxins | Not calculated | [91] |
| Near-infrared and mid-infrared spectroscopy with chemometrics | AFB1, AFB2, AFG1, AFG2 | Not calculated | [92] |
| Multiplexing fiber optic laser-induced fluorescence spectroscopy system with one-, two-, and three-probe | AFB1 | Not calculated | [93] |
| Fluorescence spectroscopy | AFB1, AFB2 | 0.2 | [94] |
| Shortwave near-infrared spectroscopy | AFB1 | Not calculated | [95] |
| Visible-near-infrared spectroscopy | AFB1, aflatoxins | Not calculated | [96] |
| Terahertz spectroscopy with chemometric methods | AFB1 | 1.0 | [97] |
| On-line fluorescence spectroscopy system | AFB1 | <6.20 | [98] |
| Hyperspectral imaging technology (UV) | Aflatoxins | Not calculated | [99] |
| Near-infrared reflectance spectroscopy-based fast versicolorin A detection | AFB1 | 8.26 | [100] |
| Laser-induced fluorescence spectroscopy | AFB1 | 6.86 | [101] |
| Fourier transform infrared spectroscopy | AFM1 | 10.0 | [102] |
| Colorimetric enzyme-linked immunosorbent assay | AFB1 | 0.06 | [103] |
| Ultraviolet-visible–near-infrared spectroscopy | Aflatoxins | Not calculated | [104] |
| Fluorescence spectroscopy and multispectral imaging | AFB1 | Not calculated | [105] |
| Fourier transform near-infrared spectroscopy | AFB1 | Not calculated | [106] |
| Handheld fluorescence spectrometry | AFB1 | 0.6 | [107] |
| Detection Method | Advantages | Limitations |
|---|---|---|
| Macroscopic Culture-Based | Low cost; simple setup; suitable for preliminary fungal screening | Time-consuming (32–120 h); requires microbiological expertise; not specific to toxins |
| Molecular Techniques | High sensitivity and specificity; enables detection of toxigenic strains | Does not detect actual toxin levels; costly equipment; technical expertise required |
| Immunochemical | Rapid; low cost; suitable for screening; can detect multiple toxins simultaneously | Cross-reactivity; matrix effects; lower quantitative accuracy; may need confirmation |
| Electrochemical Biosensors | High sensitivity; miniaturizable; potential for on-site use | Limited standardization; device-specific calibration; still in early-stage commercialization |
| Chromatographic | Gold standard; precise quantification; multi-analyte detection | Expensive instrumentation; time-consuming sample preparation; requires trained personnel |
| Spectroscopic Techniques | Rapid; non-destructive; no need for chemical reagents | Lower sensitivity; affected by matrix complexity; limited detection of low toxin concentrations |
| Aflatoxin and/or Fungus | Inhibitor | Model | Effects | References |
|---|---|---|---|---|
| A. parasiticus; aflatoxins | Lactobacillus plantarum L. delbrueckii subsp. Lactis | in vitro |
| [116] |
| AFB1 AFB2 AFM1 | Bacillus shackletonii strain L7 | in vitro |
| [112] |
| AFB1 | Pseudomonas fluorescens strain 3JW1 | in vitro |
| [9] |
| A. parasiticus AFB1 AFG1 | Bacillus amyloliquefaciens UTB2 B. subtilis UTB3 | in vitro |
| [117] |
| AFB1 | Bacillus velezensis | in vitro |
| [111] |
| A. flavus A. niger AFB1 | Lactobacillus plantarum strain ITEM 17215 | in vitro |
| [118] |
| A. parasiticus AFB1 | Bacillus mojavensis RC1A B. subtilis RC6A | in vitro |
| [119] |
| A. flavus AFB1 | Volatile organic compounds from Streptomyces alboflavus TD—1 | in vitro |
| [120] |
| AFB1 | Lactobacillus rhamnosus GG | Dairy cattle in vivo |
| [113] |
| AFB1 AFB2 AFG1 AFG2 | Recombinant laccase C30 from Saccharomyces cerevisiae | in vitro |
| [121] |
| A. flavus A. parasiticus AFB1 AFB2 AFG1 AFG2 | Lactobacillus rhamnosus L. gasseri L. plantarum | in vitro | On YES media:
| [122] |
| AFB1 AFM1 | Kluyveromyces marxianus CPY1 K. marxianus RSY5 Pichia kudriavzevii YSY2 | Dairy cattle in vivo |
| [114] |
| AFM1 | Lactobacillus paracasei 108 L. plantarum 49 L. fermentum 111 | in vitro |
| [123] |
| AFB1 | Lactobacillus plantarum | in vitro |
| [124] |
| AFB1 AFB2 AFG1 AFG2 | Aspergillus oryzae A. versicolor A. ochraceus Cladosporium subcinereum Bacillus albus B. velezensis | Human skin fibroblast cells in vitro |
| [125] |
| A. flavus AFB1 | Bacillus amyloliquefaciens WF2020 | in vitro |
| [126] |
| Aspergillus flavus AFB1 | Pseudomonas 50 isolates | in vitro |
| [127] |
| Aflatoxin and/or Fungus | Type of Botanical Agent | Model | Effects | References |
|---|---|---|---|---|
| AFB1 | Coumarin and chlorophyllin | Rats in vivo |
| [149] |
| AFB1 AFB2 AFG1 AFG2 | Jatropha pomace extract; jojoba oil; jojoba pomace extract; jatropha oil | in vitro |
| [148] |
| AFB1 | Ginger extract | HepG2 cells in vitro; rats in vivo |
| [150] |
| AFs | Banana peel powder | Rats in vivo and in vitro |
| [147] |
| AFB1 AFG1 | Cape golden berry | Rats in vivo |
| [151] |
| Aflatoxins | Date palm (Phoenix dactylifera) seeds | Chickens in vivo |
| [152] |
| AFB1 | Black tea and/or curcumin | Rats in vivo |
| [153] |
| AFB1 | Purple waxy corn extract | HepG2 cells in vitro |
| [154] |
| AFB1 | Curcumin | Ducks in vivo |
| [155] |
| Aflatoxins | Lycopene and silymarin | Ducks in vivo |
| [156] |
| AFB1 | Artichoke leaf extract | Rats in vivo |
| [157] |
| AFB1 | Curcumin | Ducks in vivo |
| [158] |
| Aflatoxins | Barley microgreen | Rats in vivo |
| [159] |
| A. parasiticus aflatoxins | Essential oils: lavandin grosso and abrial, Origanum virens, Rosmarinus officinalis; Phenolic acids: caffeic, chlorogenic, ferulic, p-coumaric | in vitro |
| [133] |
| AFB1 | Curcumin | Mice in vivo |
| [160] |
| AFB1 | Curcumin | Mice in vivo |
| [161] |
| AFB1 | Quercetin | Bovine fetal hepatocyte-derived cells (BFH12) in vitro |
| [162] |
| AFB1 AFB2 | Vegetable biocholine | Pigs in vivo |
| [163] |
| A. parasiticus—15, 16, 24 A. favus—18; aflatoxins | Halimeda opuntia extract; Turbinaria decurrens extract; Jania rubens extract | in vitro |
| [18] |
| AFB1 | Marjoram essential oil | Rabbits in vivo |
| [164] |
| AFB1 | Morin | Rats in vivo |
| [165] |
| AFB1 | Aqueous extracts of H. bacciferum (HB), O. dhofarense (OD), and Z. multiflora (ZM) | Mice in vitro, in vivo |
| [166] |
| A. flavus AFs | Pumpkin seed oil (PSO); pumpkin seed oil nanoparticles (PSO-NPs) | in vitro |
| [167] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kępka-Borkowska, K.; Chałaśkiewicz, K.; Ogłuszka, M.; Borkowski, M.; Lepczyński, A.; Pareek, C.S.; Starzyński, R.R.; Lichwiarska, E.; Sultana, S.; Kalra, G.; et al. Current Approaches to Aflatoxin B1 Control in Food and Feed Safety: Detection, Inhibition, and Mitigation. Int. J. Mol. Sci. 2025, 26, 6534. https://doi.org/10.3390/ijms26136534
Kępka-Borkowska K, Chałaśkiewicz K, Ogłuszka M, Borkowski M, Lepczyński A, Pareek CS, Starzyński RR, Lichwiarska E, Sultana S, Kalra G, et al. Current Approaches to Aflatoxin B1 Control in Food and Feed Safety: Detection, Inhibition, and Mitigation. International Journal of Molecular Sciences. 2025; 26(13):6534. https://doi.org/10.3390/ijms26136534
Chicago/Turabian StyleKępka-Borkowska, Katarzyna, Katarzyna Chałaśkiewicz, Magdalena Ogłuszka, Mateusz Borkowski, Adam Lepczyński, Chandra Shekhar Pareek, Rafał Radosław Starzyński, Elżbieta Lichwiarska, Sharmin Sultana, Garima Kalra, and et al. 2025. "Current Approaches to Aflatoxin B1 Control in Food and Feed Safety: Detection, Inhibition, and Mitigation" International Journal of Molecular Sciences 26, no. 13: 6534. https://doi.org/10.3390/ijms26136534
APA StyleKępka-Borkowska, K., Chałaśkiewicz, K., Ogłuszka, M., Borkowski, M., Lepczyński, A., Pareek, C. S., Starzyński, R. R., Lichwiarska, E., Sultana, S., Kalra, G., Purohit, N., Gralak, B., Poławska, E., & Pierzchała, M. (2025). Current Approaches to Aflatoxin B1 Control in Food and Feed Safety: Detection, Inhibition, and Mitigation. International Journal of Molecular Sciences, 26(13), 6534. https://doi.org/10.3390/ijms26136534