
Genome-Wide Identification and Analysis of the AHL Gene Family in Pepper (Capsicum annuum L.)

 ,
,
Abstract
1. Introduction
2. Results
2.1. Identification and Basic Information on the CaAHL Gene Family
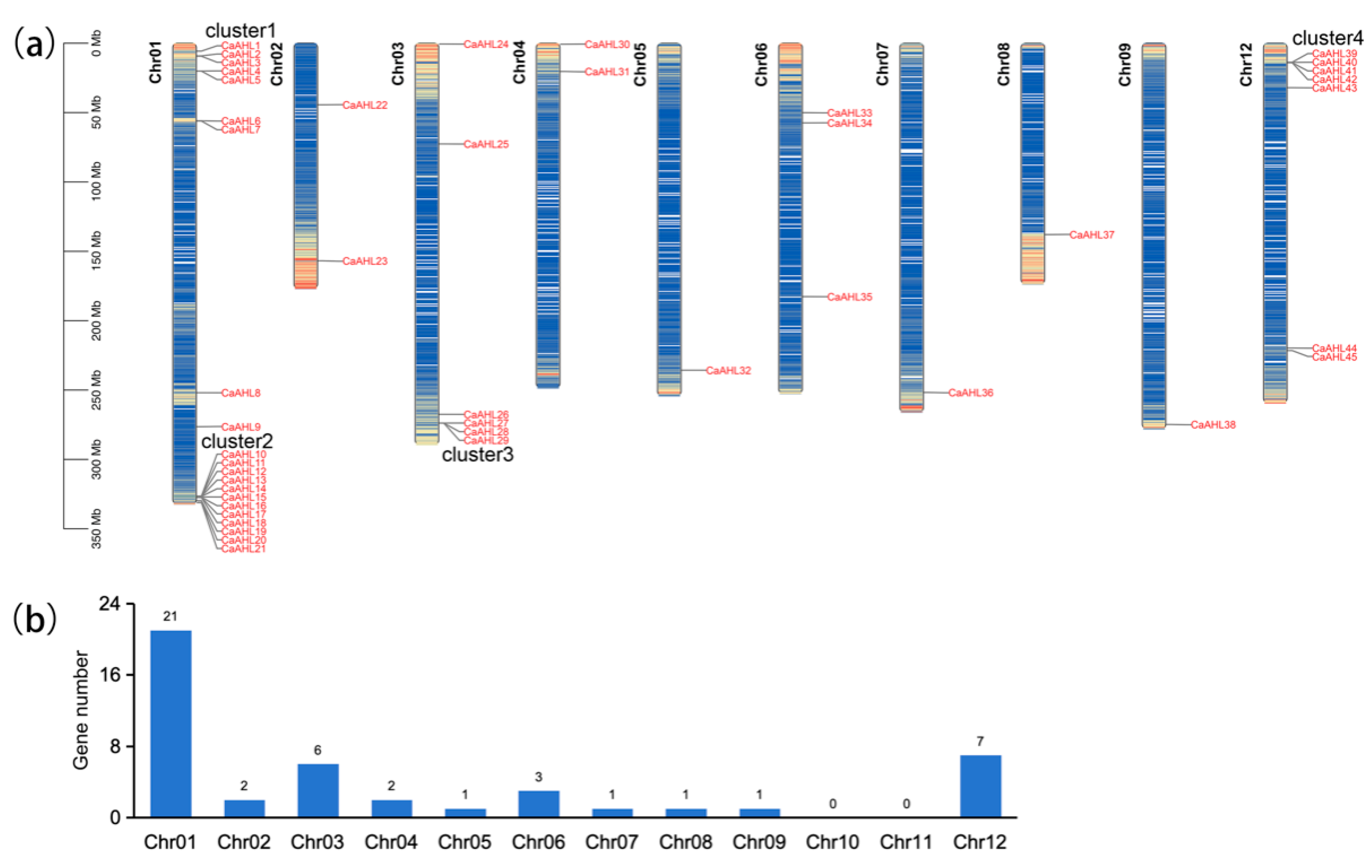
2.2. Chromosomal Distribution of the CaAHL Gene Family
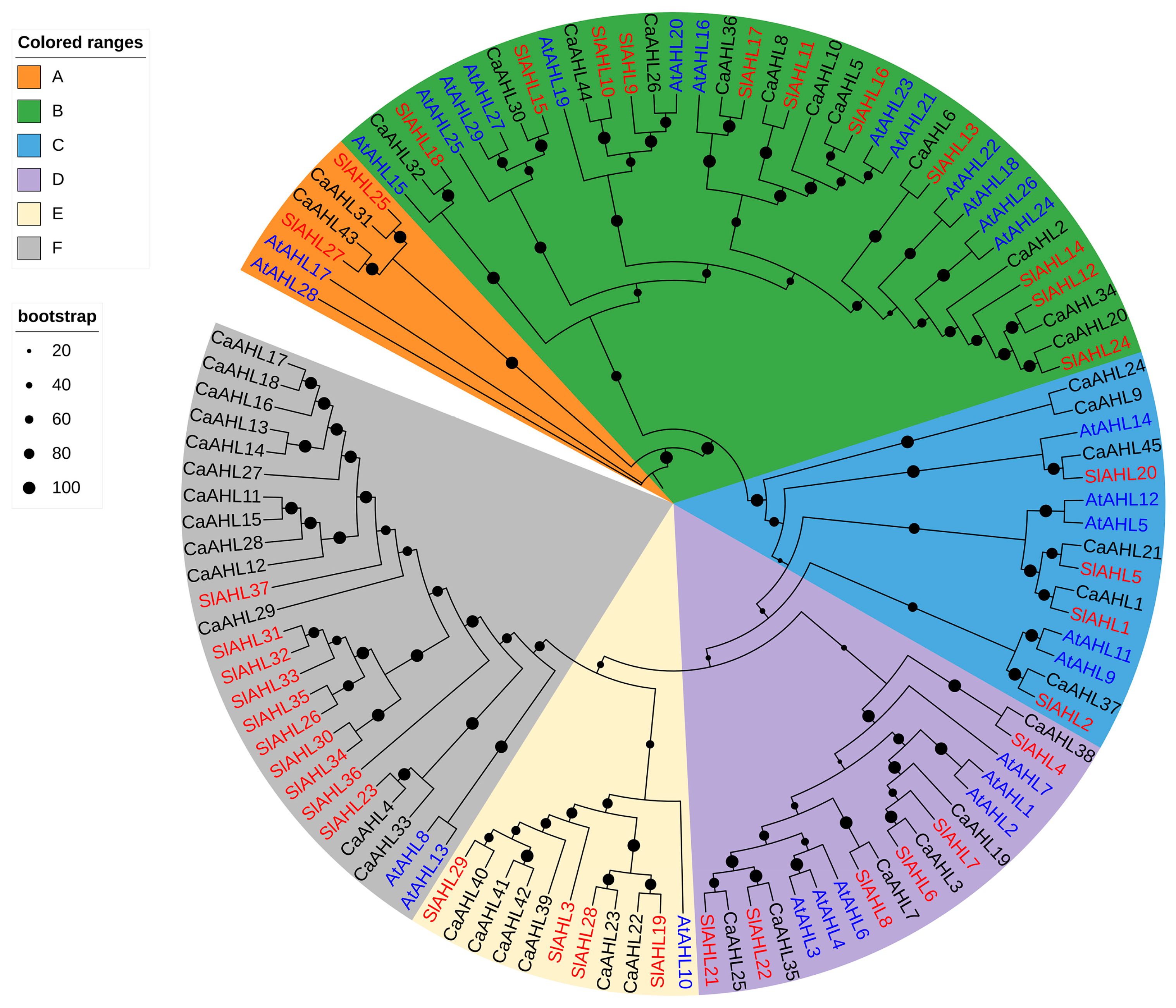
2.3. Phylogenetic Analysis of the CaAHL Gene Family
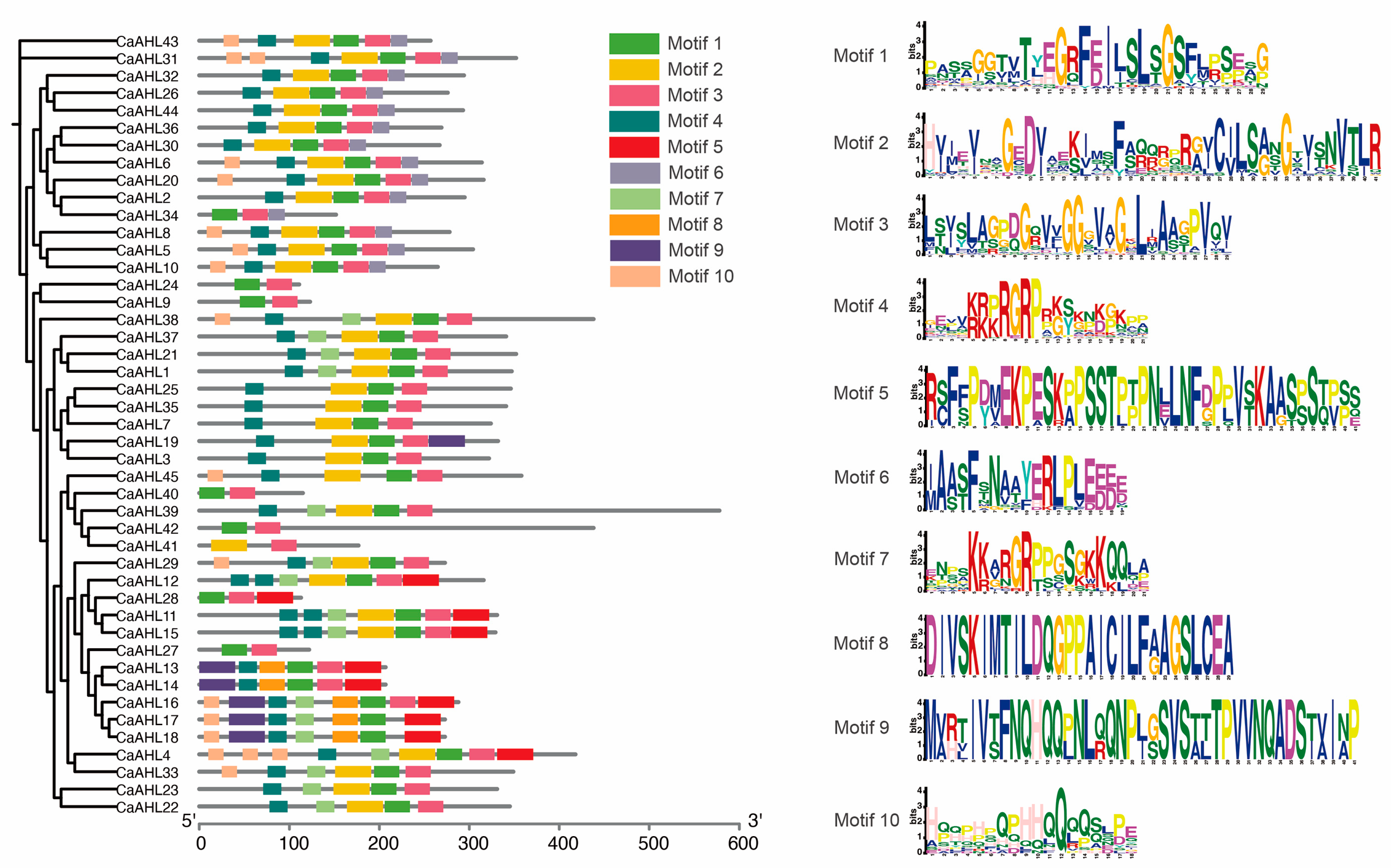
2.4. Analysis of Conserved CaAHL Motifs
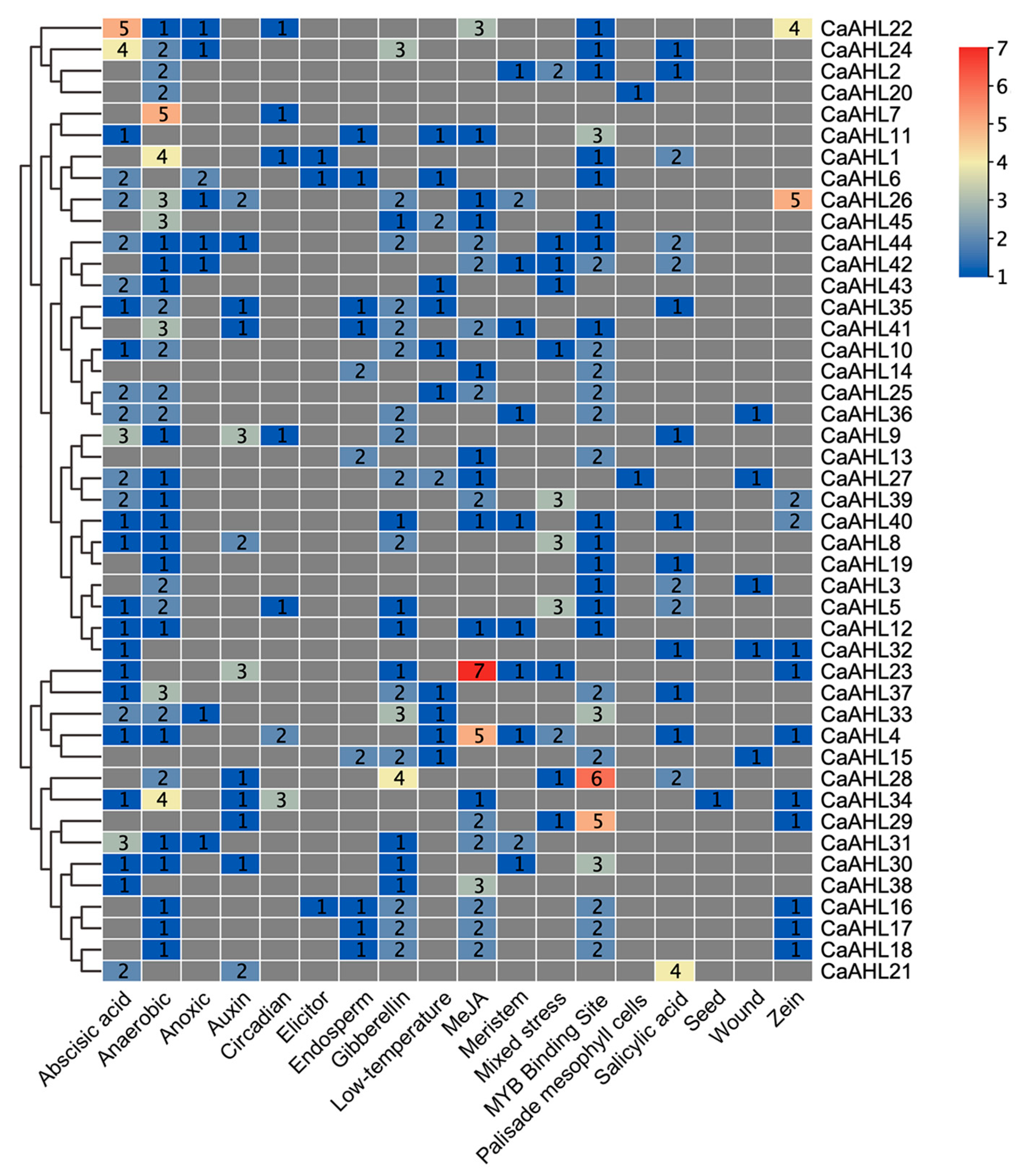
2.5. Cis-Regulatory Element Analysis of the CaAHL Promoters
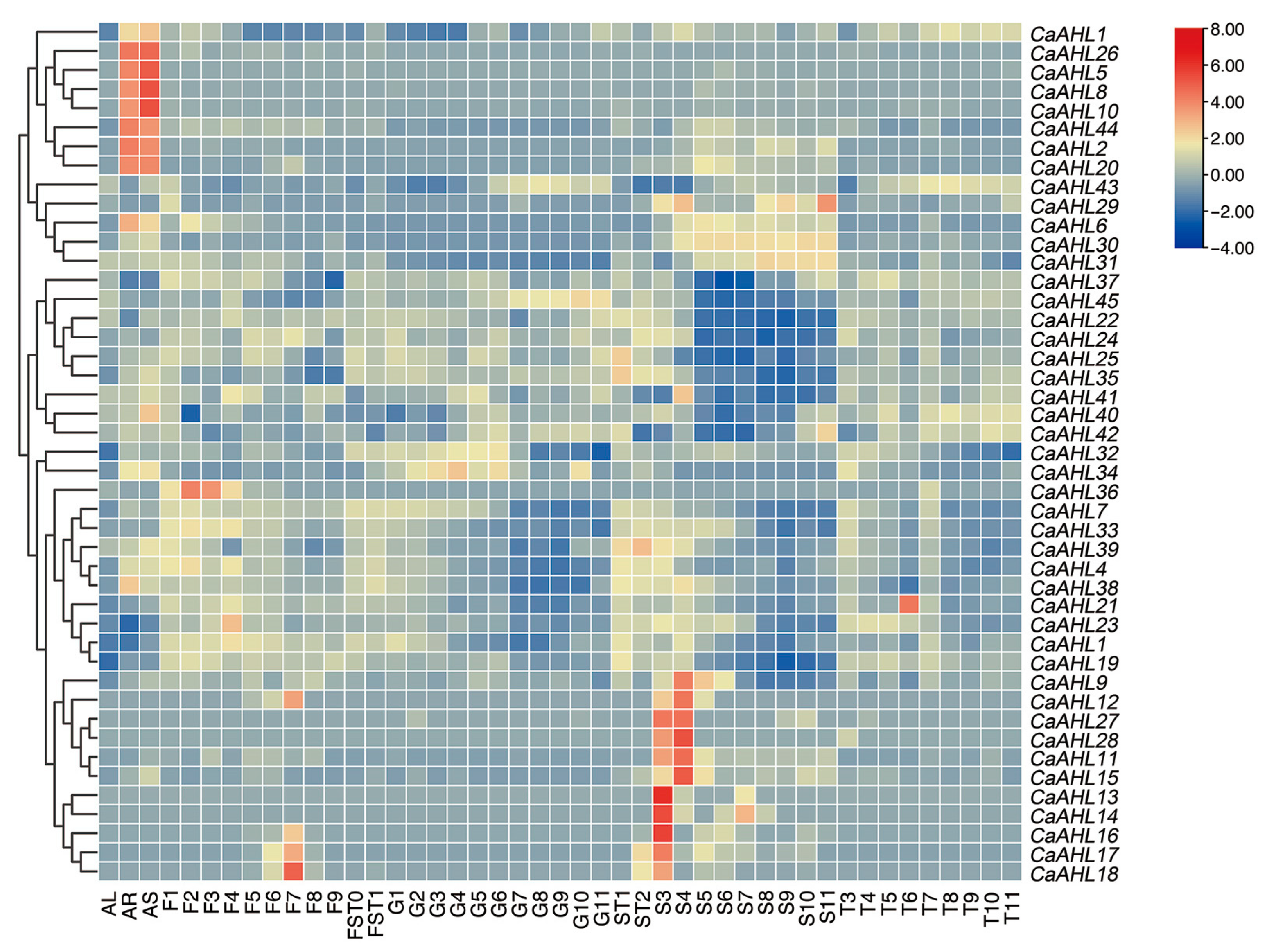
2.6. Tissue-Specific Expression Profiles of CaAHLs in Pepper
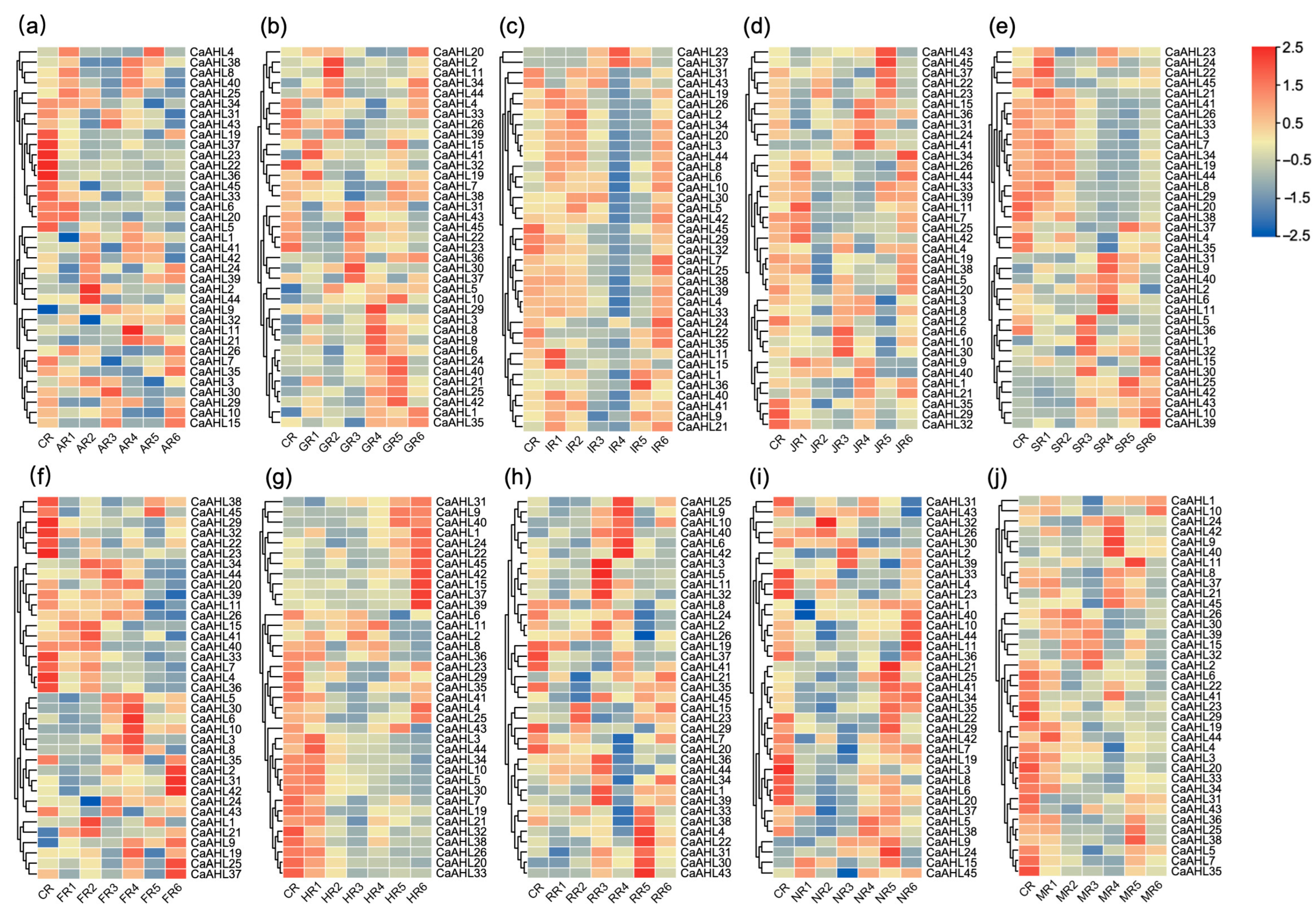
2.7. Expression Profiles of CaAHL Genes in Response to Exogenous Hormones and Abiotic Stresses
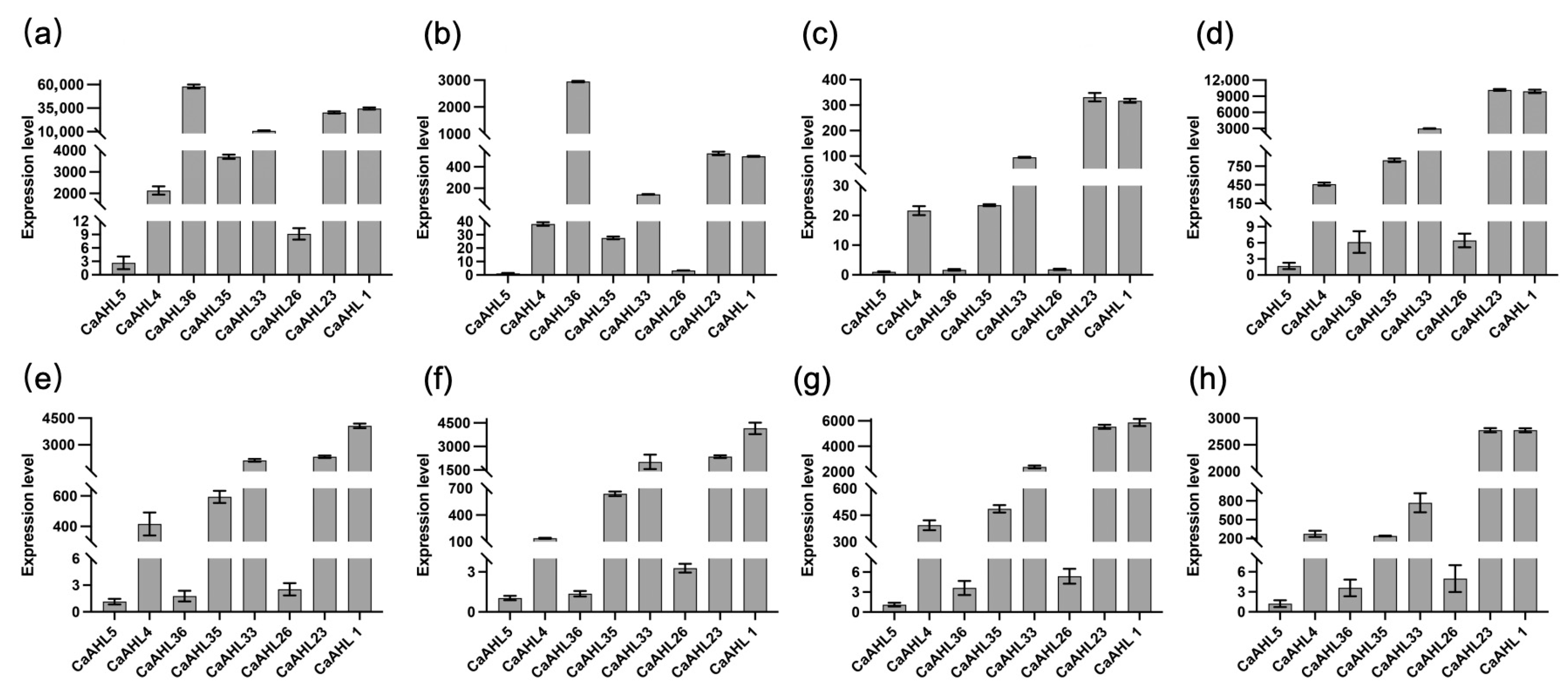
2.8. Relative Expression Levels of Eight CaAHL Genes in Bud Development
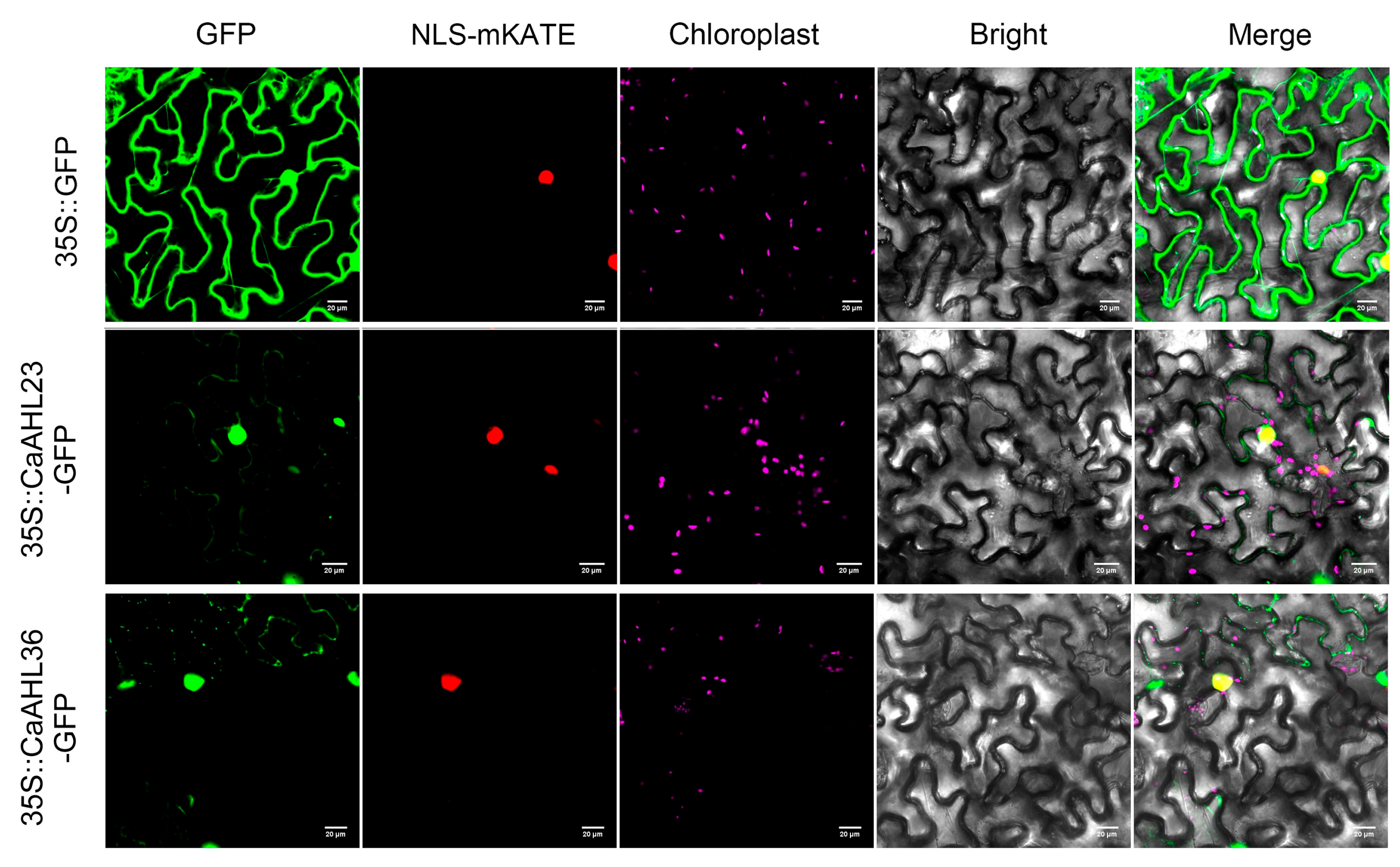
2.9. Subcellular Localization of CaAHL23 and CaAHL36
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Retrieval and Identification of AHL Genes in Pepper
4.3. Sequence Analysis and Structural Characteristics
4.4. Chromosome Localization, Tandem Duplication, and Synteny Analysis
4.5. Phylogenetic Analysis
4.6. RNA-Seq Analysis of CaAHL Genes
4.7. RNA Extraction and RT-qPCR Analysis
4.8. Subcellular Localization
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kim, S.; Park, M.; Yeom, S.-I.; Kim, Y.-M.; Lee, J.M.; Lee, H.-A.; Seo, E.; Choi, J.; Cheong, K.; Kim, K.-T.; et al. Genome sequence of the hot pepper provides insights into the evolution of pungency in Capsicum species. Nat. Genet. 2014, 46, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, U.; Li, X.; Fan, Y.; Chang, W.; Niu, Y.; Li, J.; Qu, C.; Lu, K. Multi-omics revolution to promote plant breeding efficiency. Front. Plant Sci. 2022, 13, 1062952. [Google Scholar] [CrossRef]
- Islam, K.; Momo, J.; Rawoof, A.; Vijay, A.; Anusree, V.K.; Kumar, A.; Ramchiary, N. Integrated Use of Molecular and Omics Approaches for Breeding High Yield and Stress Resistance Chili Peppers. In Smart Plant Breeding for Vegetable Crops in Post-Genomics Era; Springer Nature: Singapore, 2023; pp. 279–335. [Google Scholar]
- Street, I.H.; Shah, P.K.; Smith, A.M.; Avery, N.; Neff, M.M. The AT-hook-containing proteins SOB3/AHL29 and ESC/AHL27 are negative modulators of hypocotyl growth in Arabidopsis. Plant J. 2008, 54, 1–14. [Google Scholar] [CrossRef]
- Xiao, C.; Chen, F.; Yu, X.; Lin, C.; Fu, Y.-F. Over-expression of an AT-hook gene, AHL22, delays flowering and inhibits the elongation of the hypocotyl in Arabidopsis thaliana. Plant Mol. Biol. 2009, 71, 39–50. [Google Scholar] [CrossRef]
- Xu, L.; Zheng, S.; Witzel, K.; Van De Slijke, E.; Baekelandt, A.; Mylle, E.; Van Damme, D.; Cheng, J.; De Jaeger, G.; Inzé, D.; et al. Chromatin attachment to the nuclear matrix represses hypocotyl elongation in Arabidopsis thaliana. Nat. Commun. 2024, 15, 1286. [Google Scholar] [CrossRef] [PubMed]
- Tayengwa, R.; Koirala, P.S.; Pierce, C.F.; Werner, B.E.; Neff, M.M. Overexpression of AtAHL20 causes delayed flowering in Arabidopsis via repression of FT expression. BMC Plant Biol. 2020, 20, 559. [Google Scholar] [CrossRef]
- Uzair, M.; Xu, D.; Schreiber, L.; Shi, J.; Liang, W.; Jung, K.-H.; Chen, M.; Luo, Z.; Zhang, Y.; Yu, J.; et al. PERSISTENT TAPETAL CELL2 Is Required for Normal Tapetal Programmed Cell Death and Pollen Wall Patterning. Plant Physiol. 2020, 182, 962–976. [Google Scholar] [CrossRef]
- Machaj, G.; Grzebelus, D. Characteristics of the AT-Hook Motif Containing Nuclear Localized (AHL) Genes in Carrot Provides Insight into Their Role in Plant Growth and Storage Root Development. Genes 2021, 12, 764. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Yang, H.; Sun, H.; Lu, P.; Yan, P.; Zhao, W.; Zeng, L.; Li, Z.; Zhang, L.; Hou, W. Characterization of AHL transcription factors and functional analysis of IbAHL10 in storage root development in sweetpotato. Sci. Hortic. 2024, 338, 113718. [Google Scholar] [CrossRef]
- Favero, D.S.; Kawamura, A.; Shibata, M.; Takebayashi, A.; Jung, J.-H.; Suzuki, T.; Jaeger, K.E.; Ishida, T.; Iwase, A.; Wigge, P.A.; et al. AT-Hook Transcription Factors Restrict Petiole Growth by Antagonizing PIFs. Curr. Biol. 2020, 30, 1454–1466.e6. [Google Scholar] [CrossRef]
- Zhou, L.; Liu, Z.; Liu, Y.; Kong, D.; Li, T.; Yu, S.; Mei, H.; Xu, X.; Liu, H.; Chen, L.; et al. A novel gene OsAHL1 improves both drought avoidance and drought tolerance in rice. Sci. Rep. 2016, 6, 30264. [Google Scholar] [CrossRef] [PubMed]
- Howden, A.J.M.; Stam, R.; Heredia, V.M.; Motion, G.B.; Have, S.T.; Hodge, K.; Amaro, T.M.M.M.; Huitema, E. Quantitative analysis of the tomato nuclear proteome during Phytophthora capsici infection unveils regulators of immunity. New Phytol. 2017, 215, 309–322. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.-N.; Sun, H.-J.; Zuo, Z.-F.; Lee, D.H.; Song, P.-S.; Kang, H.-G.; Lee, H.-Y. Overexpression of ATHG1/AHL23 and ATPG3/AHL20, Arabidopsis AT-hook motif nuclear-localized genes, confers salt tolerance in transgenic Zoysia japonica. Plant Biotechnol. Rep. 2020, 14, 351–361. [Google Scholar] [CrossRef]
- Rayapuram, N.; Jarad, M.; Alhoraibi, H.M.; Bigeard, J.; Abulfaraj, A.A.; Völz, R.; Mariappan, K.G.; Almeida-Trapp, M.; Schlöffel, M.; Lastrucci, E.; et al. Chromatin phosphoproteomics unravels a function for AT-hook motif nuclear localized protein AHL13 in PAMP-triggered immunity. Proc. Natl. Acad. Sci. USA 2021, 118, e2004670118. [Google Scholar] [CrossRef]
- Bishop, E.H.; Kumar, R.; Luo, F.; Saski, C.; Sekhon, R.S. Genome-wide identification, expression profiling, and network analysis of AT-hook gene family in maize. Genomics 2020, 112, 1233–1244. [Google Scholar] [CrossRef]
- Zhao, L.; Lü, Y.; Chen, W.; Yao, J.; Li, Y.; Li, Q.; Pan, J.; Fang, S.; Sun, J.; Zhang, Y. Genome-wide identification and analyses of the AHL gene family in cotton (Gossypium). BMC Genom. 2020, 21, 69. [Google Scholar] [CrossRef]
- Wang, M.; Chen, B.; Zhou, W.; Xie, L.; Wang, L.; Zhang, Y.; Zhang, Q. Genome-wide identification and expression analysis of the AT-hook Motif Nuclear Localized gene family in soybean. BMC Genom. 2021, 22, 361. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, S.; Mishra, A. Genome-wide identification and analyses of the AHL gene family in rice (Oryza sativa). 3 Biotech 2023, 13, 248. [Google Scholar] [CrossRef]
- Yun, J.; Kim, Y.-S.; Jung, J.-H.; Seo, P.J.; Park, C.-M. The AT-hook Motif-containing Protein AHL22 Regulates Flowering Initiation by Modifying FLOWERING LOCUS T Chromatin in Arabidopsis. J. Biol. Chem. 2012, 287, 15307–15316. [Google Scholar] [CrossRef]
- Xu, Y.; Gan, E.-S.; Ito, T. The AT-hook/PPC domain protein TEK negatively regulates floral repressors including MAF4 and MAF5. Plant Signal. Behav. 2013, 8, e25006. [Google Scholar] [CrossRef]
- Lou, Y.; Xu, X.-F.; Zhu, J.; Gu, J.-N.; Blackmore, S.; Yang, Z.-N. The tapetal AHL family protein TEK determines nexine formation in the pollen wall. Nat. Commun. 2014, 5, 3855. [Google Scholar] [CrossRef] [PubMed]
- Karami, O.; Rahimi, A.; Mak, P.; Horstman, A.; Boutilier, K.; Compier, M.; van der Zaal, B.; Offringa, R. An Arabidopsis AT-hook motif nuclear protein mediates somatic embryogenesis and coinciding genome duplication. Nat. Commun. 2021, 12, 2508. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Zhao, J.; Sun, H.; Xiong, C.; Sun, X.; Wang, X.; Wang, Z.; Jarret, R.; Wang, J.; Tang, B.; et al. Genomes of cultivated and wild Capsicum species provide insights into pepper domestication and population differentiation. Nat. Commun. 2023, 14, 5487. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Yu, H.; Deng, Y.; Zheng, J.; Liu, M.; Ou, L.; Yang, B.; Dai, X.; Ma, Y.; Feng, S.; et al. PepperHub, an Informatics Hub for the Chili Pepper Research Community. Mol. Plant 2017, 10, 1129–1132. [Google Scholar] [CrossRef]
- Ambadas, D.A.; Singh, A.; Jha, R.K.; Chauhan, D.; Santhosh, B.; Sharma, V.K. Genome-wide dissection of AT-hook motif nuclear-localized gene family and their expression profiling for drought and salt stress in rice (Oryza sativa). Front. Plant Sci. 2023, 14, 1283555. [Google Scholar] [CrossRef]
- Qiao, X.; Li, Q.; Yin, H.; Qi, K.; Li, L.; Wang, R.; Zhang, S.; Paterson, A.H. Gene duplication and evolution in recurring polyploidization–diploidization cycles in plants. Genome Biol. 2019, 20, 38. [Google Scholar] [CrossRef]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef]
- Chen, C.; Wu, Y.; Li, J.; Wang, X.; Zeng, Z.; Xu, J.; Liu, Y.; Feng, J.; Chen, H.; He, Y. TBtools-II: A “one for all, all for one” bioinformatics platform for biological big-data mining. Mol. Plant 2023, 16, 1733–1742. [Google Scholar] [CrossRef]
- Horton, P.; Park, K.-J.; Obayashi, T.; Fujita, N.; Harada, H.; Adams-Collier, C.J.; Nakai, K. WoLF PSORT: Protein localization predictor. Nucleic Acids Res. 2007, 35, W585–W587. [Google Scholar] [CrossRef]
- Bailey, T.L.; Elkan, C. Fitting a mixture model by expectation maximization to discover motifs in biopolymers. In Proceedings of the Intelligent Systems for Molecular Biology, Stanford University, Palo Alto, CA, USA, 14–17 August 1994; Volume 2, pp. 28–36. [Google Scholar]
- Chao, J.-T.; Kong, Y.-Z.; Wang, Q.; Sun, Y.; Gong, D.; Lv, J.; Liu, G. MapGene2Chrom, a tool to draw gene physical map based on Perl and SVG languages. Yi Chuan 2015, 37, 91–97. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v6: Recent updates to the phylogenetic tree display and annotation tool. Nucleic Acids Res. 2024, 52, W78–W82. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Gene ID | Chromosome Location | Protein Length | Molecular Weight (kDa) | Theoretical pI | Instability Index | Grand Average of Hydropathicity | Subcellular Localization 1 |
|---|---|---|---|---|---|---|---|---|
| CaAHL1 | Caz01g03310.1 | Chr01:5691094–5697294 | 347 | 35.27 | 9.62 | 38.59 | −0.33 | nucl |
| CaAHL2 | Caz01g04670.1 | Chr01:9114966–9116603 | 295 | 31.17 | 5.79 | 52.67 | −0.54 | nucl |
| CaAHL3 | Caz01g04810.1 | Chr01:9383031–9386809 | 322 | 33.18 | 8.74 | 50.79 | −0.17 | chlo |
| CaAHL4 | Caz01g08370.1 | Chr01:20063956–20073612 | 418 | 43.66 | 8.82 | 60.17 | −0.76 | nucl |
| CaAHL5 | Caz01g08400.1 | Chr01:20111560–20112474 | 304 | 31.69 | 5.75 | 51.83 | −0.27 | nucl |
| CaAHL6 | Caz01g13980.1 | Chr01:55893363–55894391 | 314 | 33.10 | 6.83 | 53.54 | −0.60 | nucl |
| CaAHL7 | Caz01g14080.1 | Chr01:56075436–56079692 | 324 | 33.72 | 9.57 | 47.83 | −0.25 | chlo |
| CaAHL8 | Caz01g31040.1 | Chr01:251924071–251924907 | 278 | 29.43 | 6.54 | 50.25 | −0.47 | chlo |
| CaAHL9 | Caz01g35190.1 | Chr01:276409484–276418256 | 123 | 13.45 | 5.87 | 47.09 | −0.20 | cyto |
| CaAHL10 | Caz01g40510.1 | Chr01:326409380–326414186 | 265 | 28.25 | 5.96 | 44.58 | −0.44 | nucl |
| CaAHL11 | Caz01g40520.1 | Chr01:326434617–326442536 | 331 | 35.20 | 9.51 | 49.49 | −0.46 | chlo |
| CaAHL12 | Caz01g40530.1 | Chr01:326492721–326618469 | 316 | 33.80 | 10.04 | 51.86 | −0.37 | nucl |
| CaAHL13 | Caz01g40540.1 | Chr01:326621531–326626277 | 207 | 22.30 | 6.96 | 44.75 | −0.13 | nucl |
| CaAHL14 | Caz01g40550.1 | Chr01:326687022–326691770 | 207 | 22.34 | 7.84 | 44.38 | −0.14 | chlo |
| CaAHL15 | Caz01g40640.1 | Chr01:327024609–327031508 | 329 | 34.72 | 9.65 | 48.09 | −0.47 | chlo |
| CaAHL16 | Caz01g40680.1 | Chr01:327253871–327258557 | 288 | 30.70 | 8.76 | 47.03 | −0.45 | chlo |
| CaAHL17 | Caz01g40710.1 | Chr01:327378313–327383011 | 273 | 29.34 | 7.79 | 47.92 | −0.53 | chlo |
| CaAHL18 | Caz01g40720.1 | Chr01:327501110–327505809 | 273 | 29.30 | 7.79 | 51.24 | −0.53 | chlo |
| CaAHL19 | Caz01g41290.1 | Chr01:329720440–329727085 | 332 | 34.13 | 9.54 | 44.92 | −0.10 | chlo |
| CaAHL20 | Caz01g41410.1 | Chr01:329900501–329901727 | 316 | 33.79 | 6.05 | 59.8 | −0.63 | chlo |
| CaAHL21 | Caz01g41740.1 | Chr01:331181444–331186914 | 352 | 35.75 | 9.44 | 49.49 | −0.27 | nucl |
| CaAHL22 | Caz02g02600.1 | Chr02:44359225–44368231 | 345 | 34.69 | 8.97 | 55.44 | −0.11 | E.R. |
| CaAHL23 | Caz02g20690.1 | Chr02:156842236–156848762 | 331 | 33.33 | 9.99 | 50.43 | −0.23 | nucl |
| CaAHL24 | Caz03g00210.1 | Chr03:551605–558146 | 111 | 12.02 | 6.39 | 30.43 | 0.01 | chlo |
| CaAHL25 | Caz03g21400.1 | Chr03:72579177–72587401 | 346 | 36.17 | 6.33 | 45.59 | −0.38 | nucl |
| CaAHL26 | Caz03g34730.1 | Chr03:267518820–267519650 | 276 | 28.85 | 5.45 | 57.84 | −0.47 | cyto |
| CaAHL27 | Caz03g36660.1 | Chr03:273808903–273810928 | 122 | 12.87 | 5.19 | 34.02 | 0.18 | cyto |
| CaAHL28 | Caz03g36670.1 | Chr03:273824211–273825851 | 113 | 12.01 | 6.01 | 47.05 | −0.25 | nucl |
| CaAHL29 | Caz03g36680.1 | Chr03:273829352–273850851 | 273 | 28.76 | 9.64 | 49.08 | −0.30 | vacu |
| CaAHL30 | Caz04g00390.1 | Chr04:713251–718475 | 267 | 26.36 | 6.42 | 48.45 | −0.13 | nucl |
| CaAHL31 | Caz04g08200.1 | Chr04:20375417–20376564 | 352 | 37.79 | 7.05 | 63.62 | −0.66 | nucl |
| CaAHL32 | Caz05g17580.1 | Chr05:235719249–235723745 | 294 | 31.56 | 5.35 | 52.68 | −0.61 | nucl |
| CaAHL33 | Caz06g17080.1 | Chr06:50064634–50076684 | 349 | 36.50 | 9.34 | 53.47 | −0.51 | nucl |
| CaAHL34 | Caz06g17990.1 | Chr06:57415662–57416120 | 152 | 15.59 | 4.44 | 51.44 | −0.03 | nucl |
| CaAHL35 | Caz06g24080.1 | Chr06:182665460–182672828 | 341 | 35.70 | 7.02 | 40.07 | −0.30 | plas |
| CaAHL36 | Caz07g19270.1 | Chr07:251735483–251736292 | 269 | 28.24 | 9.24 | 33.67 | −0.13 | cyto |
| CaAHL37 | Caz08g07460.1 | Chr08:137932240–137938844 | 341 | 34.99 | 10.26 | 58.12 | −0.22 | nucl |
| CaAHL38 | Caz09g21720.1 | Chr09:274809109–274813725 | 438 | 45.01 | 9.38 | 50.94 | −0.34 | nucl |
| CaAHL39 | Caz12g05920.1 | Chr12:13682382–13697371 | 578 | 60.51 | 7.72 | 52.36 | −0.30 | chlo |
| CaAHL40 | Caz12g05950.1 | Chr12:13811174–13813071 | 115 | 11.71 | 11.25 | 59.27 | −0.01 | chlo |
| CaAHL41 | Caz12g06070.1 | Chr12:14037114–14041618 | 177 | 18.10 | 4.89 | 59.53 | −0.20 | chlo |
| CaAHL42 | Caz12g06080.1 | Chr12:14056732–14062760 | 438 | 47.47 | 9.27 | 50.27 | −0.21 | chlo |
| CaAHL43 | Caz12g08880.1 | Chr12:32039622–32040395 | 257 | 28.05 | 7.83 | 49.44 | −0.42 | nucl |
| CaAHL44 | Caz12g18220.1 | Chr12:219722263–219725994 | 293 | 29.60 | 6.16 | 46.17 | −0.33 | nucl |
| CaAHL45 | Caz12g18510.1 | Chr12:221530027–221540499 | 358 | 37.13 | 9.57 | 50.03 | −0.37 | cyto |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sui, X.-Y.; Li, Y.-L.; Wang, X.; Zhong, Y.; Cui, Q.-Z.; Luo, Y.; Tang, B.-Q.; Liu, F.; Zou, X.-X. Genome-Wide Identification and Analysis of the AHL Gene Family in Pepper (Capsicum annuum L.). Int. J. Mol. Sci. 2025, 26, 6527. https://doi.org/10.3390/ijms26136527
Sui X-Y, Li Y-L, Wang X, Zhong Y, Cui Q-Z, Luo Y, Tang B-Q, Liu F, Zou X-X. Genome-Wide Identification and Analysis of the AHL Gene Family in Pepper (Capsicum annuum L.). International Journal of Molecular Sciences. 2025; 26(13):6527. https://doi.org/10.3390/ijms26136527
Chicago/Turabian StyleSui, Xiao-Yan, Yan-Long Li, Xi Wang, Yi Zhong, Qing-Zhi Cui, Yin Luo, Bing-Qian Tang, Feng Liu, and Xue-Xiao Zou. 2025. "Genome-Wide Identification and Analysis of the AHL Gene Family in Pepper (Capsicum annuum L.)" International Journal of Molecular Sciences 26, no. 13: 6527. https://doi.org/10.3390/ijms26136527
APA StyleSui, X.-Y., Li, Y.-L., Wang, X., Zhong, Y., Cui, Q.-Z., Luo, Y., Tang, B.-Q., Liu, F., & Zou, X.-X. (2025). Genome-Wide Identification and Analysis of the AHL Gene Family in Pepper (Capsicum annuum L.). International Journal of Molecular Sciences, 26(13), 6527. https://doi.org/10.3390/ijms26136527