
Sex and Circadian Rhythm Dependent Behavioral Effects of Chronic Stress in Mice and Modulation of Clock Genes in the Prefrontal Cortex

 , , ,
, , ,  ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
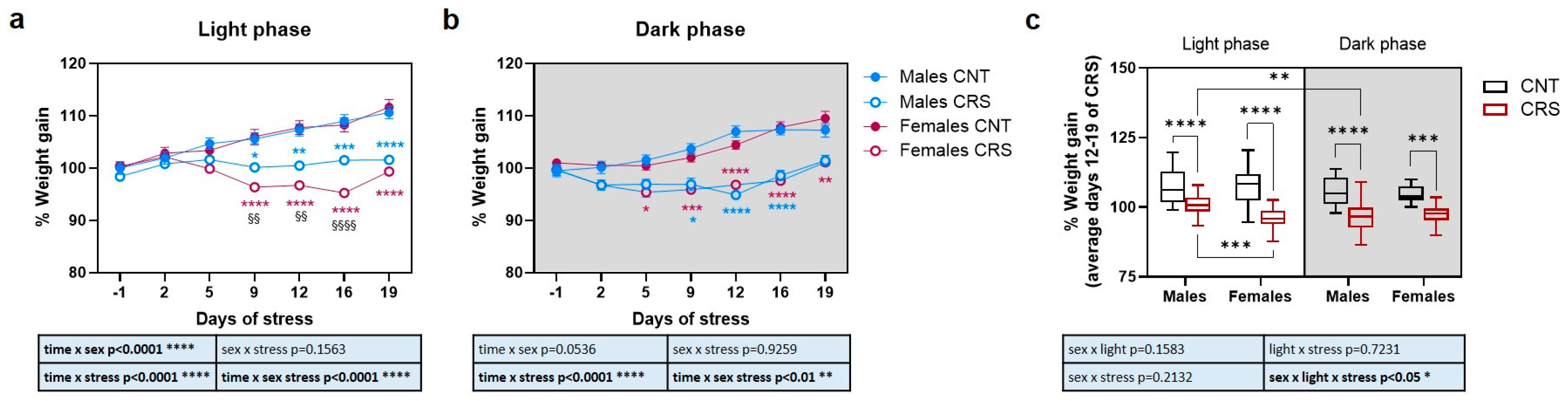
2.1. Sex and Circadian Rhythm Dependent Effects of Chronic Restraint Stress on Body Weight in Mice
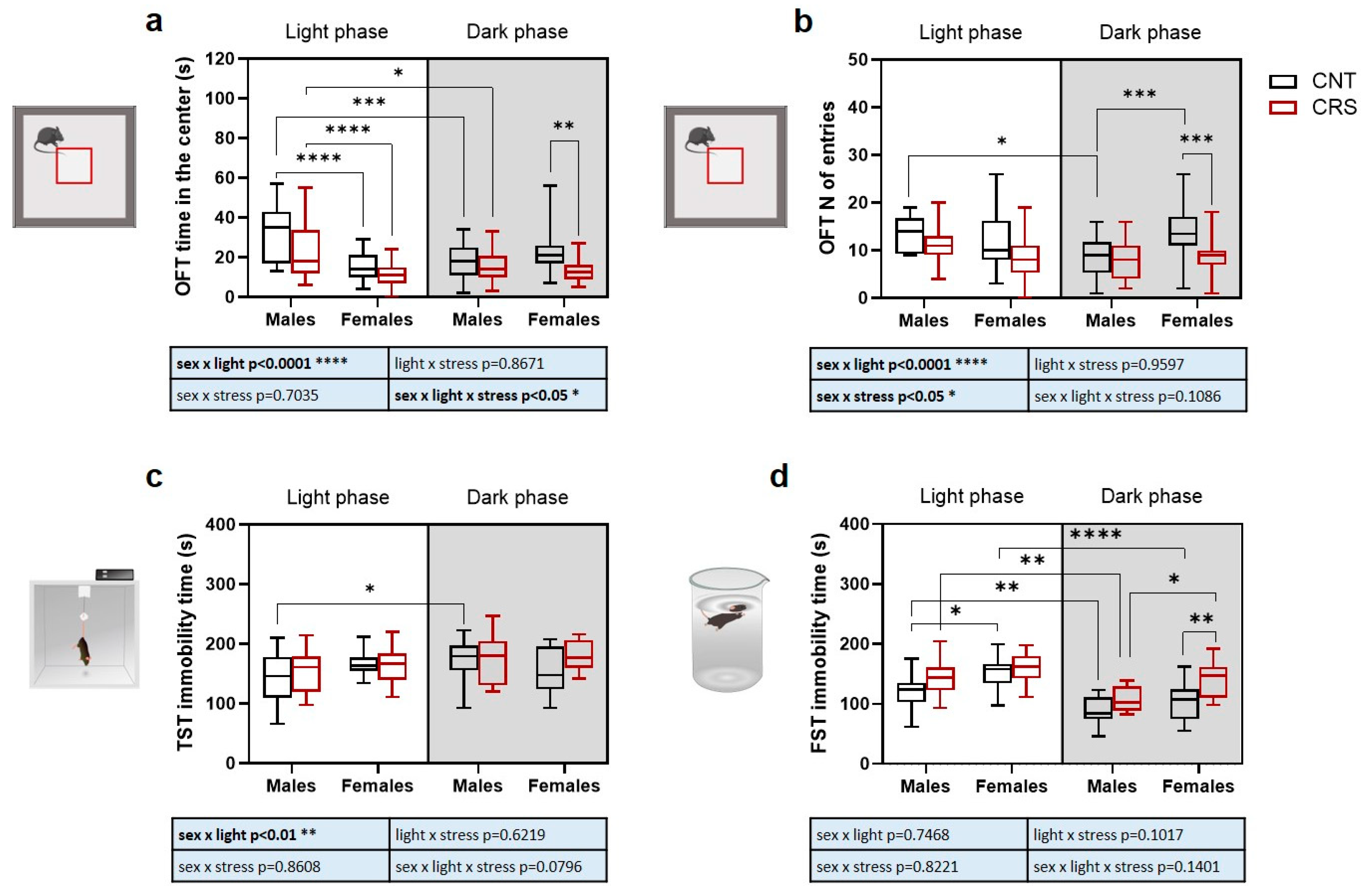
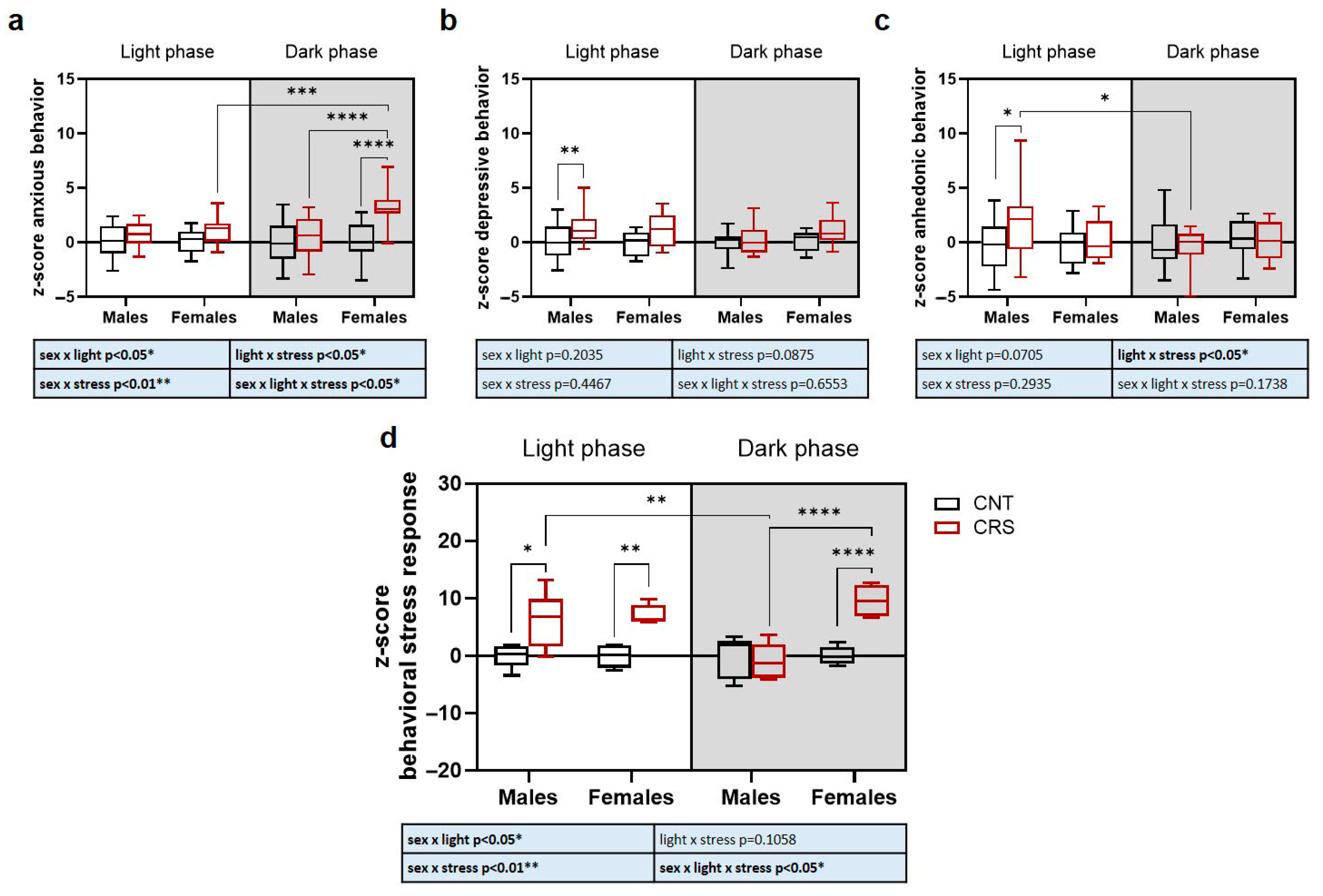
2.2. Sex and Circadian Rhythm Dependent Behavioral Effects of Chronic Restraint Stress in Mice
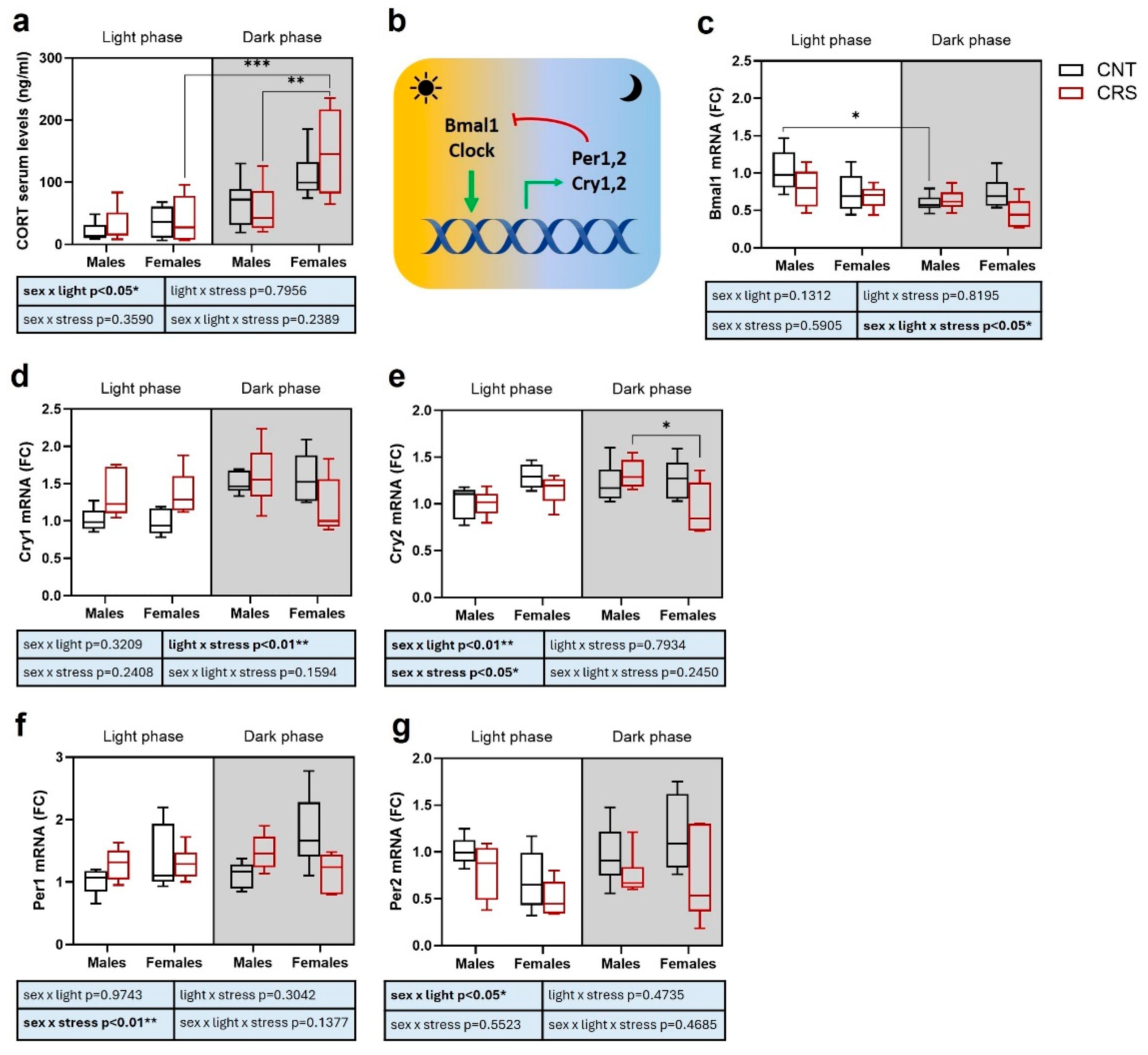
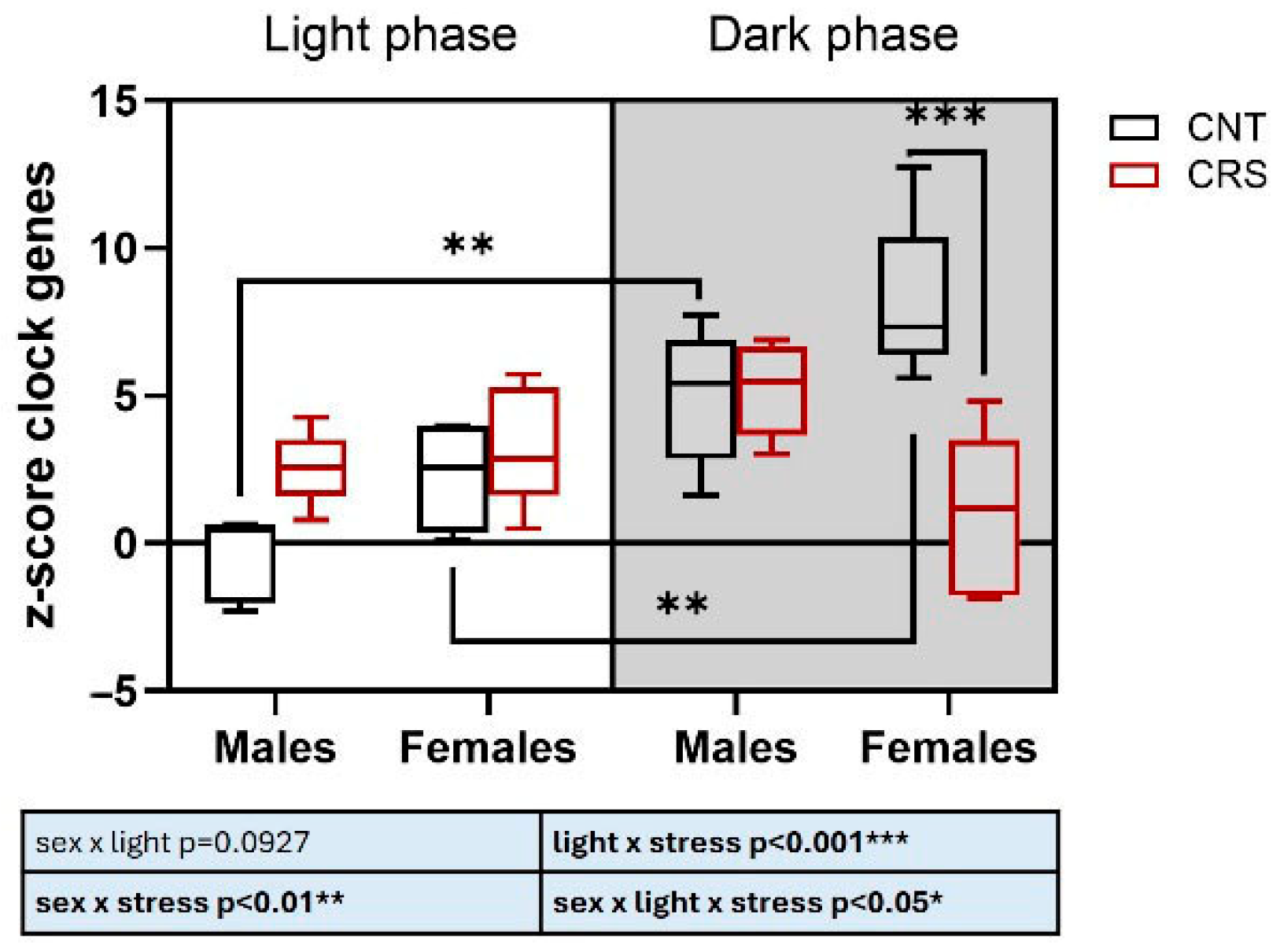
2.3. Sex and Circadian Rhythm Dependent Effects of Chronic Restraint Stress on Corticosterone Serum Levels and Expression of Clock Genes in the Prefrontal Cortex of Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Body Weight Measurements
4.3. Chronic Restraint Stress
4.4. Tail Suspension Test
4.5. Open Field Test
4.6. Sucrose Splash Test
4.7. Forced Swim Test
4.8. Sacrifice
4.9. Corticosterone Measurement
4.10. RNA Isolation, Reverse Transcription, and Real-Time PCR
4.11. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CRS | Chronic Restraint Stress |
| FST | Forced swim test |
| HPA | Hypothalamic–pituitary–adrenal axis |
| OFT | Open Field test |
| PFC | Prefrontal cortex |
| SCN | Suprachiasmatic nucleus |
| SST | Sucrose splash test |
| TST | Tail suspension test |
References
- McEwen, B. Allostasis and Allostatic Load Implications for Neuropsychopharmacology. Neuropsychopharmacology 2000, 22, 108–124. [Google Scholar] [CrossRef]
- Sanacora, G.; Yan, Z.; Popoli, M. The Stressed Synapse 2.0: Pathophysiological Mechanisms in Stress-Related Neuropsychiatric Disorders. Nat. Rev. Neurosci. 2022, 23, 86–103. [Google Scholar] [CrossRef] [PubMed]
- Penner-Goeke, S.; Binder, E.B. Epigenetics and Depression. Dialogues Clin. Neurosci. 2019, 21, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Larrieu, T.; Sandi, C. Stress-Induced Depression: Is Social Rank a Predictive Risk Factor? BioEssays 2018, 40, 1800012. [Google Scholar] [CrossRef]
- Slavich, G.M.; Sacher, J. Stress, Sex Hormones, Inflammation, and Major Depressive Disorder: Extending Social Signal Transduction Theory of Depression to Account for Sex Differences in Mood Disorders. Psychopharmacology 2019, 236, 3063–3079. [Google Scholar] [CrossRef] [PubMed]
- Mengelkoch, S.; Slavich, G.M. Sex Differences in Stress Susceptibility as a Key Mechanism Underlying Depression Risk. Curr. Psychiatry Rep. 2024, 26, 157–165. [Google Scholar] [CrossRef]
- Hodes, G.E.; Bangasser, D.; Sotiropoulos, I.; Kokras, N.; Dalla, C. Sex Differences in Stress Response: Classical Mechanisms and Beyond. Curr. Neuropharmacol. 2024, 22, 475–494. [Google Scholar] [CrossRef]
- Kronfeld-Schor, N.; Einat, H. Circadian Rhythms and Depression: Human Psychopathology and Animal Models. Neuropharmacology 2012, 62, 101–114. [Google Scholar] [CrossRef]
- Takahashi, J.S. Transcriptional Architecture of the Mammalian Circadian Clock. Nat. Rev. Genet. 2017, 18, 164–179. [Google Scholar] [CrossRef]
- Begemann, K.; Rawashdeh, O.; Olejniczak, I.; Pilorz, V.; de Assis, L.V.M.; Osorio-Mendoza, J.; Oster, H. Endocrine Regulation of Circadian Rhythms. NPJ Biol. Timing Sleep 2025, 2, 10. [Google Scholar] [CrossRef]
- Boyd, H.M.; Frick, K.M.; Kwapis, J.L. Connecting the Dots: Potential Interactions Between Sex Hormones and the Circadian System During Memory Consolidation. J. Biol. Rhythms 2023, 38, 537–555. [Google Scholar] [CrossRef] [PubMed]
- Friedman, N.P.; Robbins, T.W. The Role of Prefrontal Cortex in Cognitive Control and Executive Function. Neuropsychopharmacology 2022, 47, 72–89. [Google Scholar] [CrossRef] [PubMed]
- Hare, B.D.; Duman, R.S. Prefrontal Cortex Circuits in Depression and Anxiety: Contribution of Discrete Neuronal Populations and Target Regions. Mol. Psychiatry 2020, 25, 2742–2758. [Google Scholar] [CrossRef]
- Hiser, J.; Koenigs, M. The Multifaceted Role of the Ventromedial Prefrontal Cortex in Emotion, Decision Making, Social Cognition, and Psychopathology. Biol. Psychiatry 2018, 83, 638–647. [Google Scholar] [CrossRef] [PubMed]
- Sarrazin, D.H.; Gardner, W.; Marchese, C.; Balzinger, M.; Ramanathan, C.; Schott, M.; Rozov, S.; Veleanu, M.; Vestring, S.; Normann, C.; et al. Prefrontal Cortex Molecular Clock Modulates Development of Depression-like Phenotype and Rapid Antidepressant Response in Mice. Nat. Commun. 2024, 15, 7257. [Google Scholar] [CrossRef]
- Roberts, B.L.; Karatsoreos, I.N. Circadian Desynchronization Disrupts Physiological Rhythms of Prefrontal Cortex Pyramidal Neurons in Mice. Sci. Rep. 2023, 13, 9181. [Google Scholar] [CrossRef]
- Smart.Servier.Com. Available online: https://smart.servier.com/ (accessed on 30 May 2025).
- Kuti, D.; Winkler, Z.; Horváth, K.; Juhász, B.; Szilvásy-Szabó, A.; Fekete, C.; Ferenczi, S.; Kovács, K.J. The Metabolic Stress Response: Adaptation to Acute-, Repeated- and Chronic Challenges in Mice. iScience 2022, 25, 104693. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.Y.; Lee, D.H.; Kang, S.S. Effects of Chronic Restraint Stress on Body Weight, Food Intake, and Hypothalamic Gene Expressions in Mice. Endocrinol. Metab. 2013, 28, 288. [Google Scholar] [CrossRef]
- Harris, R.B.S.; Palmondon, J.; Leshin, S.; Flatt, W.P.; Richard, D. Chronic Disruption of Body Weight but Not of Stress Peptides or Receptors in Rats Exposed to Repeated Restraint Stress. Horm. Behav. 2006, 49, 615–625. [Google Scholar] [CrossRef]
- Kamens, H.M.; Anziano, E.K.; Horton, W.J.; Cavigelli, S.A. Chronic Adolescent Restraint Stress Downregulates MiRNA-200a Expression in Male and Female C57BL/6J and BALB/CJ Mice. Genes 2024, 15, 873. [Google Scholar] [CrossRef]
- Wang, J.; Sun, L.; You, J.; Peng, H.; Yan, H.; Wang, J.; Sun, F.; Cui, M.; Wang, S.; Zhang, Z.; et al. Role and Mechanism of PVN–Sympathetic–Adipose Circuit in Depression and Insulin Resistance Induced by Chronic Stress. EMBO Rep. 2023, 24, e57176. [Google Scholar] [CrossRef] [PubMed]
- Olave, F.A.; Aguayo, F.I.; Román-Albasini, L.; Corrales, W.A.; Silva, J.P.; González, P.I.; Lagos, S.; García, M.A.; Alarcón-Mardones, M.; Rojas, P.S.; et al. Chronic Restraint Stress Produces Sex-Specific Behavioral and Molecular Outcomes in the Dorsal and Ventral Rat Hippocampus. Neurobiol. Stress 2022, 17, 100440. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Liu, Q.; Mao, Y.; Wen, J.; Chen, G. Different Action of Glucocorticoid Receptor in Adipose Tissue Remodelling to Modulate Energy Homeostasis by Chronic Restraint Stress. Lipids Health Dis. 2025, 24, 121. [Google Scholar] [CrossRef]
- Mannan, M.; Mamun, A.; Doi, S.; Clavarino, A. Is There a Bi-Directional Relationship between Depression and Obesity among Adult Men and Women? Systematic Review and Bias-Adjusted Meta Analysis. Asian J. Psychiatr. 2016, 21, 51–66. [Google Scholar] [CrossRef]
- Maxwell, M.A.; Cole, D.A. Weight Change and Appetite Disturbance as Symptoms of Adolescent Depression: Toward an Integrative Biopsychosocial Model. Clin. Psychol. Rev. 2009, 29, 260–273. [Google Scholar] [CrossRef] [PubMed]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders; American Psychiatric Association: Washington, DC, USA, 2013; ISBN 0-89042-555-8. [Google Scholar]
- Aslani, S.; Harb, M.R.; Costa, P.S.; Almeida, O.F.X.; Sousa, N.; Palha, J.A. Day and Night: Diurnal Phase Influences the Response to Chronic Mild Stress. Front. Behav. Neurosci. 2014, 8, 82. [Google Scholar] [CrossRef]
- Spencer, J.L.; Waters, E.M.; Milner, T.A.; Lee, F.S.; McEwen, B.S. BDNF Variant Val66Met Interacts with Estrous Cycle in the Control of Hippocampal Function. Proc. Natl. Acad. Sci. USA 2010, 107, 4395–4400. [Google Scholar] [CrossRef]
- Zhao, W.; Li, Q.; Ma, Y.; Wang, Z.; Fan, B.; Zhai, X.; Hu, M.; Wang, Q.; Zhang, M.; Zhang, C.; et al. Behaviors Related to Psychiatric Disorders and Pain Perception in C57BL/6J Mice During Different Phases of Estrous Cycle. Front. Neurosci. 2021, 15, 650793. [Google Scholar] [CrossRef]
- Cryan, J.F.; Slattery, D.A. Animal Models of Mood Disorders: Recent Developments. Curr. Opin. Psychiatry 2007, 20, 1–7. [Google Scholar] [CrossRef]
- Bowman, R.; Frankfurt, M.; Luine, V. Sex Differences in Anxiety and Depression: Insights from Adult Rodent Models of Chronic Stress and Neural Plasticity. Front. Behav. Neurosci. 2025, 19, 1591973. [Google Scholar] [CrossRef]
- Oyola, M.G.; Handa, R.J. Hypothalamic–Pituitary–Adrenal and Hypothalamic–Pituitary–Gonadal Axes: Sex Differences in Regulation of Stress Responsivity. Stress 2017, 20, 476–494. [Google Scholar] [CrossRef] [PubMed]
- Seedat, S.; Scott, K.M.; Angermeyer, M.C.; Berglund, P.; Bromet, E.J.; Brugha, T.S.; Demyttenaere, K.; de Girolamo, G.; Haro, J.M.; Jin, R.; et al. Cross-National Associations Between Gender and Mental Disorders in the World Health Organization World Mental Health Surveys. Arch. Gen. Psychiatry 2009, 66, 785. [Google Scholar] [CrossRef] [PubMed]
- McLean, C.P.; Asnaani, A.; Litz, B.T.; Hofmann, S.G. Gender Differences in Anxiety Disorders: Prevalence, Course of Illness, Comorbidity and Burden of Illness. J. Psychiatr. Res. 2011, 45, 1027–1035. [Google Scholar] [CrossRef] [PubMed]
- Koch, C.E.; Leinweber, B.; Drengberg, B.C.; Blaum, C.; Oster, H. Interaction between Circadian Rhythms and Stress. Neurobiol. Stress 2017, 6, 57–67. [Google Scholar] [CrossRef]
- Herman, J.P.; McKlveen, J.M.; Ghosal, S.; Kopp, B.; Wulsin, A.; Makinson, R.; Scheimann, J.; Myers, B. Regulation of the Hypothalamic-Pituitary-Adrenocortical Stress Response. In Comprehensive Physiology; Wiley: Hoboken, NJ, USA, 2016; pp. 603–621. [Google Scholar]
- Malisch, J.L.; Saltzman, W.; Gomes, F.R.; Rezende, E.L.; Jeske, D.R.; Garland, T., Jr. Baseline and Stress-Induced Plasma Corticosterone Concentrations of Mice Selectively Bred for High Voluntary Wheel Running. Physiol. Biochem. Zool. 2007, 80, 146–156. [Google Scholar] [CrossRef]
- Musazzi, L.; Sala, N.; Tornese, P.; Gallivanone, F.; Belloli, S.; Conte, A.; Di Grigoli, G.; Chen, F.; Ikinci, A.; Treccani, G.; et al. Acute Inescapable Stress Rapidly Increases Synaptic Energy Metabolism in Prefrontal Cortex and Alters Working Memory Performance. Cereb. Cortex 2019, 29, 4948–4957. [Google Scholar] [CrossRef]
- Bonini, D.; Mora, C.; Tornese, P.; Sala, N.; Filippini, A.; La Via, L.; Milanese, M.; Calza, S.; Bonanno, G.; Racagni, G.; et al. Acute Footshock Stress Induces Time-Dependent Modifications of AMPA/NMDA Protein Expression and AMPA Phosphorylation. Neural Plast. 2016, 2016, 7267865. [Google Scholar] [CrossRef] [PubMed]
- Karin, O.; Raz, M.; Tendler, A.; Bar, A.; Korem Kohanim, Y.; Milo, T.; Alon, U. A New Model for the HPA Axis Explains Dysregulation of Stress Hormones on the Timescale of Weeks. Mol. Syst. Biol. 2020, 16, e9510. [Google Scholar] [CrossRef]
- Borrow, A.P.; Heck, A.L.; Miller, A.M.; Sheng, J.A.; Stover, S.A.; Daniels, R.M.; Bales, N.J.; Fleury, T.K.; Handa, R.J. Chronic Variable Stress Alters Hypothalamic-Pituitary-Adrenal Axis Function in the Female Mouse. Physiol. Behav. 2019, 209, 112613. [Google Scholar] [CrossRef]
- Tornese, P.; Sala, N.; Bonini, D.; Bonifacino, T.; La Via, L.; Milanese, M.; Treccani, G.; Seguini, M.; Ieraci, A.; Mingardi, J.; et al. Chronic Mild Stress Induces Anhedonic Behavior and Changes in Glutamate Release, BDNF Trafficking and Dendrite Morphology Only in Stress Vulnerable Rats. The Rapid Restorative Action of Ketamine. Neurobiol. Stress 2019, 10, 100160. [Google Scholar] [CrossRef]
- Calabrese, F.; Savino, E.; Papp, M.; Molteni, R.; Riva, M.A. Chronic Mild Stress-Induced Alterations of Clock Gene Expression in Rat Prefrontal Cortex: Modulatory Effects of Prolonged Lurasidone Treatment. Pharmacol. Res. 2016, 104, 140–150. [Google Scholar] [CrossRef]
- Musaelyan, K.; Yildizoglu, S.; Bozeman, J.; Du Preez, A.; Egeland, M.; Zunszain, P.A.; Pariante, C.M.; Fernandes, C.; Thuret, S. Chronic Stress Induces Significant Gene Expression Changes in the Prefrontal Cortex alongside Alterations in Adult Hippocampal Neurogenesis. Brain Commun. 2020, 2, fcaa153. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, S.; Bouzinova, E.; Fahrenkrug, J.; Wiborg, O. Altered Expression Pattern of Clock Genes in a Rat Model of Depression. Int. J. Neuropsychopharmacol. 2016, 19, pyw061. [Google Scholar] [CrossRef] [PubMed]
- Peirson, S.N.; Brown, L.A.; Pothecary, C.A.; Benson, L.A.; Fisk, A.S. Light and the Laboratory Mouse. J. Neurosci. Methods 2018, 300, 26–36. [Google Scholar] [CrossRef]
- Musazzi, L.; Tornese, P.; Sala, N.; Lee, F.S.; Popoli, M.; Ieraci, A. Acute Stress Induces an Aberrant Increase of Presynaptic Release of Glutamate and Cellular Activation in the Hippocampus of BDNF Val/Met Mice. J. Cell Physiol. 2022, 237, 3834–3844. [Google Scholar] [CrossRef] [PubMed]
- Ieraci, A.; Mallei, A.; Popoli, M. Social Isolation Stress Induces Anxious-Depressive-Like Behavior and Alterations of Neuroplasticity-Related Genes in Adult Male Mice. Neural Plast. 2016, 2016, 6212983. [Google Scholar] [CrossRef]
- Bouguiyoud, N.; Roullet, F.; Bronchti, G.; Frasnelli, J.; Al Aïn, S. Anxiety and Depression Assessments in a Mouse Model of Congenital Blindness. Front. Neurosci. 2022, 15, 807434. [Google Scholar] [CrossRef]
- Barbieri, S.S.; Sandrini, L.; Musazzi, L.; Popoli, M.; Ieraci, A. Apocynin Prevents Anxiety-Like Behavior and Histone Deacetylases Overexpression Induced by Sub-Chronic Stress in Mice. Biomolecules 2021, 11, 885. [Google Scholar] [CrossRef]
- Bonifacino, T.; Mingardi, J.; Facchinetti, R.; Sala, N.; Frumento, G.; Ndoj, E.; Valenza, M.; Paoli, C.; Ieraci, A.; Torazza, C.; et al. Changes at Glutamate Tripartite Synapses in the Prefrontal Cortex of a New Animal Model of Resilience/Vulnerability to Acute Stress. Transl. Psychiatry 2023, 13, 62. [Google Scholar] [CrossRef]
- Giacopuzzi, E.; Gennarelli, M.; Sacco, C.; Filippini, A.; Mingardi, J.; Magri, C.; Barbon, A. Genome-Wide Analysis of Consistently RNA Edited Sites in Human Blood Reveals Interactions with MRNA Processing Genes and Suggests Correlations with Cell Types and Biological Variables. BMC Genom. 2018, 19, 963. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mingardi, J.; Giovenzana, M.; Nicosia, N.; Misztak, P.; Ieraci, A.; Musazzi, L. Sex and Circadian Rhythm Dependent Behavioral Effects of Chronic Stress in Mice and Modulation of Clock Genes in the Prefrontal Cortex. Int. J. Mol. Sci. 2025, 26, 6410. https://doi.org/10.3390/ijms26136410
Mingardi J, Giovenzana M, Nicosia N, Misztak P, Ieraci A, Musazzi L. Sex and Circadian Rhythm Dependent Behavioral Effects of Chronic Stress in Mice and Modulation of Clock Genes in the Prefrontal Cortex. International Journal of Molecular Sciences. 2025; 26(13):6410. https://doi.org/10.3390/ijms26136410
Chicago/Turabian StyleMingardi, Jessica, Mattia Giovenzana, Noemi Nicosia, Paulina Misztak, Alessandro Ieraci, and Laura Musazzi. 2025. "Sex and Circadian Rhythm Dependent Behavioral Effects of Chronic Stress in Mice and Modulation of Clock Genes in the Prefrontal Cortex" International Journal of Molecular Sciences 26, no. 13: 6410. https://doi.org/10.3390/ijms26136410
APA StyleMingardi, J., Giovenzana, M., Nicosia, N., Misztak, P., Ieraci, A., & Musazzi, L. (2025). Sex and Circadian Rhythm Dependent Behavioral Effects of Chronic Stress in Mice and Modulation of Clock Genes in the Prefrontal Cortex. International Journal of Molecular Sciences, 26(13), 6410. https://doi.org/10.3390/ijms26136410