
GeneHarmony: A Knowledge-Based Tool for Biomarker Discovery in Disease: Sjögren’s Disease vs. Rheumatoid Arthritis and Systemic Lupus Erythematosus


Abstract
1. Introduction
2. Results
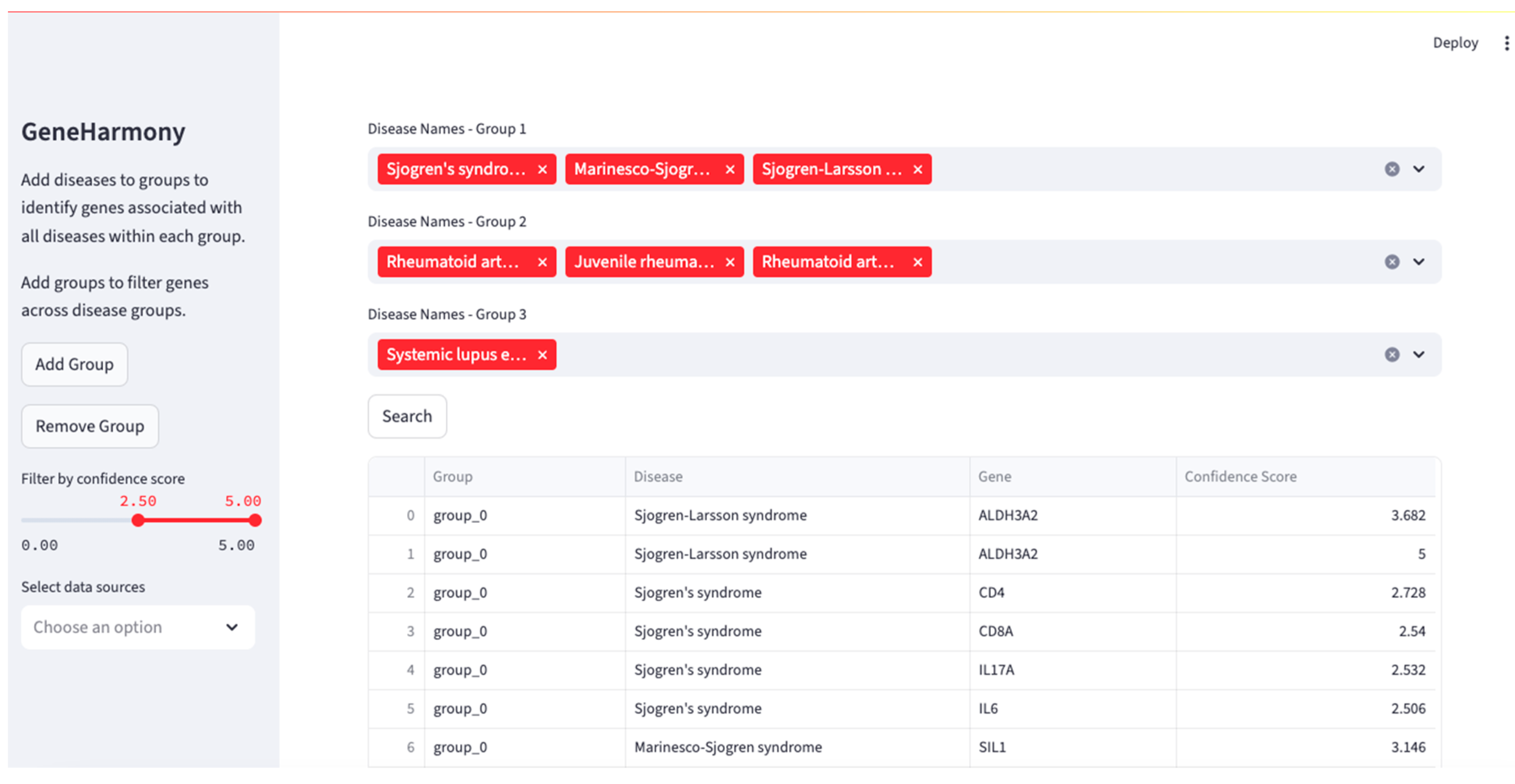
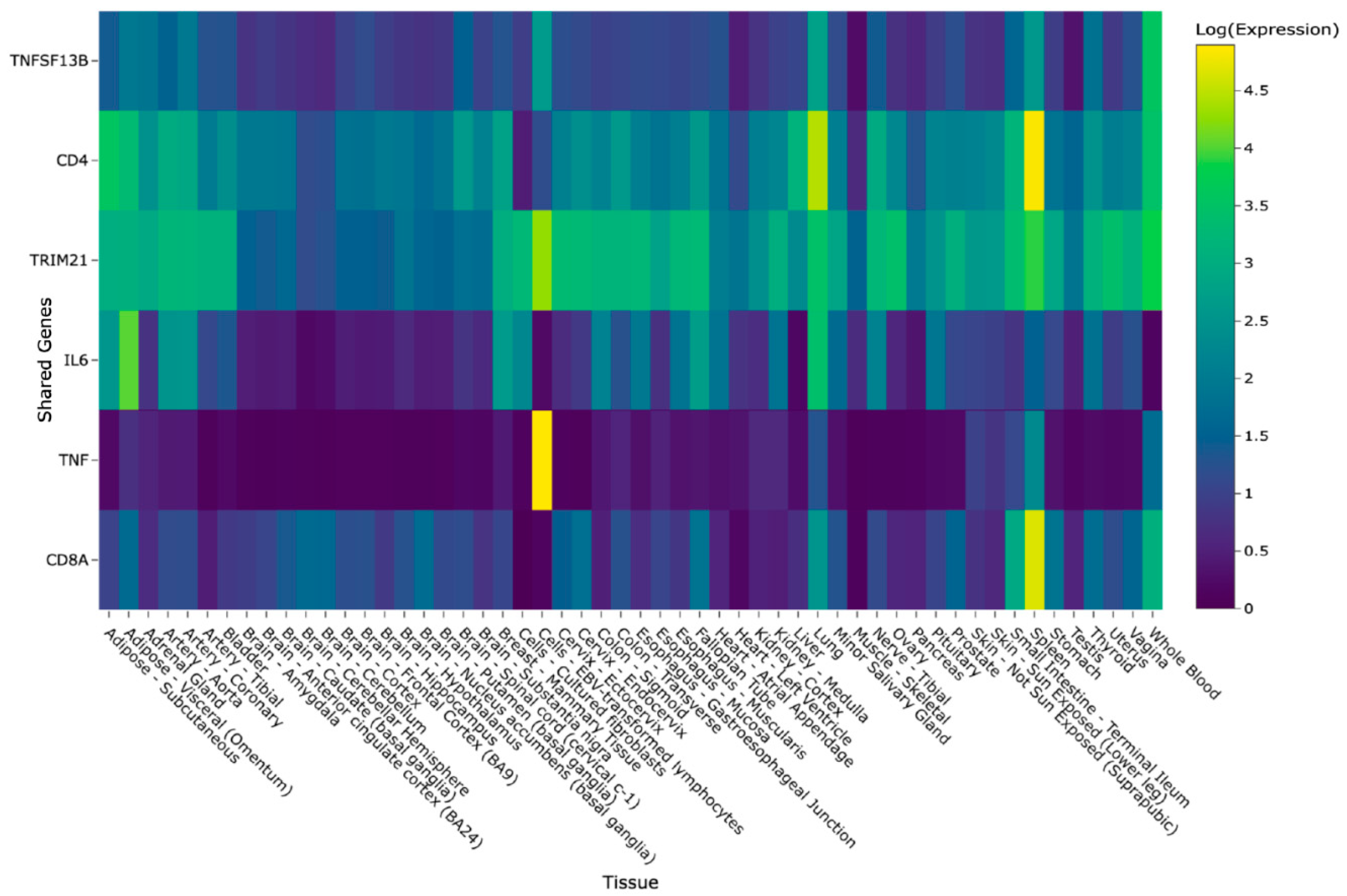
Data Usage and Application Results
3. Discussion
4. Methods
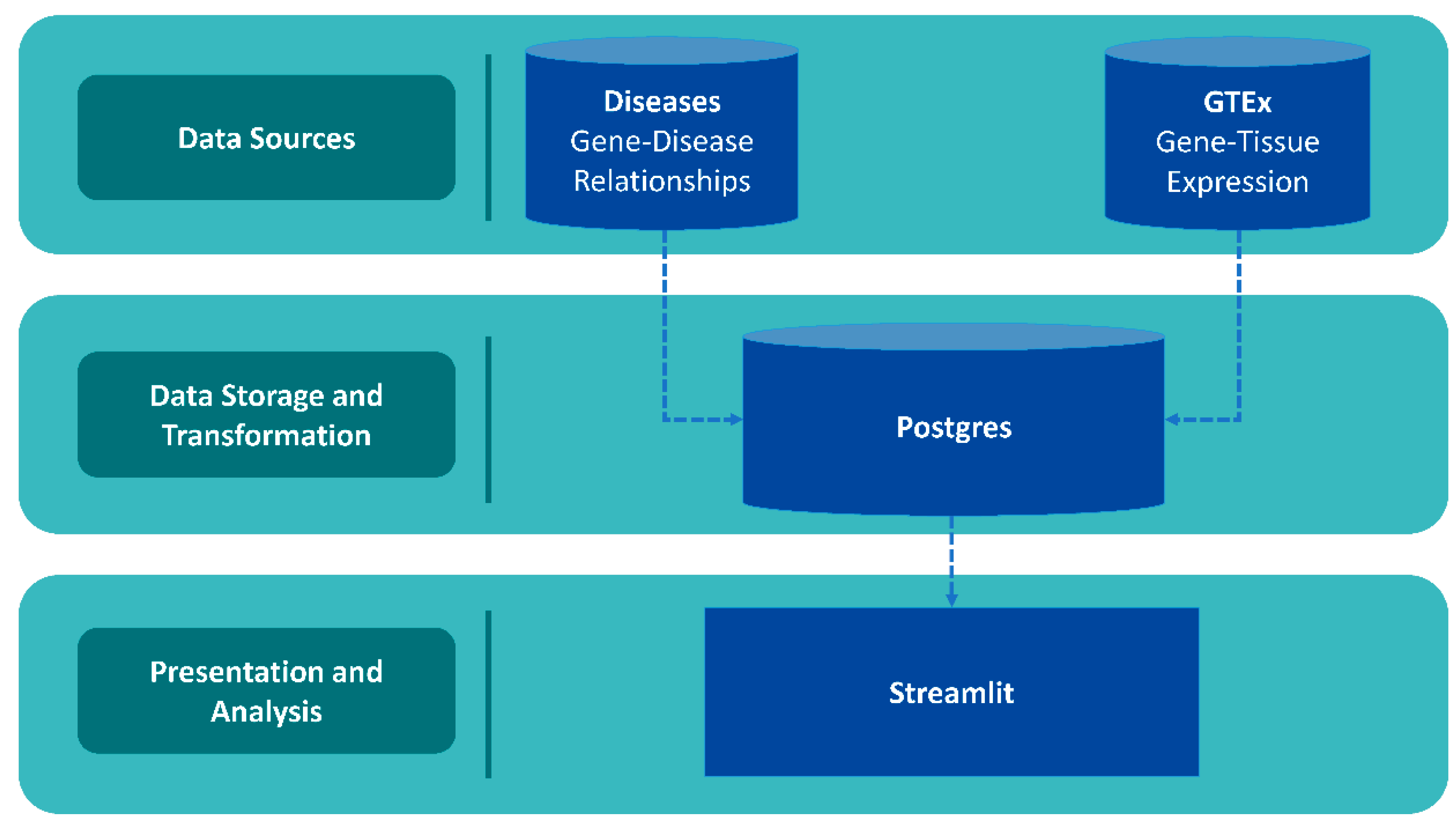
4.1. Database Architecture and Data Collection
4.2. Download and Usage
4.3. Application: Utilizing GeneHarmony for Cross-Disease Gene Association Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wang, L.; Wang, F.S.; Gershwin, M.E. Human autoimmune diseases: A comprehensive update. J. Intern. Med. 2015, 278, 369–395. [Google Scholar] [CrossRef] [PubMed]
- Ortíz-Fernández, L.; Martín, J.; Alarcón-Riquelme, M.E. A Summary on the Genetics of Systemic Lupus Erythematosus, Rheumatoid Arthritis, Systemic Sclerosis, and Sjögren’s Syndrome. Clin. Rev. Allergy Immunol. 2023, 64, 392–411. [Google Scholar] [CrossRef] [PubMed]
- Grissa, D.; Junge, A.; Oprea, T.I.; Jensen, L.J. Diseases 2.0: A weekly updated database of disease-gene associations from text mining and data integration. Database J. Biol. Databases Curation 2022, 2022, baac019. [Google Scholar] [CrossRef] [PubMed]
- Lonsdale, J.; Thomas, J.; Salvatore, M.; Phillips, R.; Lo, E.; Shad, S.; Hasz, R.; Walters, G.; Garcia, F.; Young, N.; et al. GTEx Consortium the Genotype-Tissue Expression (GTEx) project. Nat. Genet. 2013, 45, 580–585. [Google Scholar] [CrossRef]
- Pletscher-Frankild, S.; Pallejà, A.; Tsafou, K.; Binder, J.X.; Jensen, L.J. DISEASES: Text mining and data integration of disease-gene associations. Methods 2015, 74, 83–89. [Google Scholar] [CrossRef]
- Mills, K.H.G. IL-17 and IL-17-producing cells in protection versus pathology. Nat. Rev. Immunol. 2023, 23, 38–54. [Google Scholar] [CrossRef]
- Jang, D.I.; Lee, A.H.; Shin, H.Y.; Song, H.R.; Park, J.H.; Kang, T.B.; Lee, S.R.; Yang, S.H. The Role of Tumor Necrosis Factor Alpha (TNF-α) in Autoimmune Disease and Current TNF-α Inhibitors in Therapeutics. Int. J. Mol. Sci. 2021, 22, 2719. [Google Scholar] [CrossRef]
- Zhou, J.; Kawai, T.; Yu, Q. Pathogenic role of endogenous TNF-α in the development of Sjögren’s-like sialadenitis and secretory dysfunction in non-obese diabetic mice. Lab. Investig. A J. Tech. Methods Pathol. 2017, 97, 458–467. [Google Scholar] [CrossRef]
- Moutsopoulos, N.M.; Katsifis, G.E.; Angelov, N.; Leakan, R.A.; Sankar, V.; Pillemer, S.; Wahl, S.M. Lack of efficacy of etanercept in Sjögren syndrome correlates with failed suppression of tumour necrosis factor alpha and systemic immune activation. Ann. Rheum. Dis. 2008, 67, 1437–1443. [Google Scholar] [CrossRef]
- Santos-Moreno, P.; Rodríguez-Vargas, G.S.; Rodríguez-Linares, P.; Ibatá, L.; Martínez, S.; Rodríguez-Florido, F.; Rojas-Villarraga, A. Effectiveness and Persistence of Anti-TNFα Treatment in Patients with Rheumatoid Arthritis—A 7 Years Real-World Cohort Study. Biol. Targets Ther. 2024, 18, 339–347. [Google Scholar] [CrossRef]
- Aringer, M.; Smolen, J.S. The role of tumor necrosis factor-alpha in systemic lupus erythematosus. Arthritis Res. Ther. 2008, 10, 202. [Google Scholar] [CrossRef] [PubMed]
- Richter, P.; Macovei, L.A.; Mihai, I.R.; Cardoneanu, A.; Burlui, M.A.; Rezus, E. Cytokines in Systemic Lupus Erythematosus-Focus on TNF-α and IL-17. Int. J. Mol. Sci. 2023, 24, 14413. [Google Scholar] [CrossRef]
- Fischer, R.; Kontermann, R.E.; Pfizenmaier, K. Selective Targeting of TNF Receptors as a Novel Therapeutic Approach. Front. Cell Dev. Biol. 2020, 8, 401. [Google Scholar] [CrossRef]
- Ghorbaninezhad, F.; Leone, P.; Alemohammad, H.; Najafzadeh, B.; Nourbakhsh, N.S.; Prete, M.; Malerba, E.; Saeedi, H.; Tabrizi, N.J.; Racanelli, V.; et al. Tumor necrosis factorα in systemic lupus erythematosus: Structure, function and therapeutic implications (Review). Int. J. Mol. Med. 2022, 49, 43. [Google Scholar] [CrossRef]
- Zhu, L.J.; Yang, X.; Yu, X.Q. Anti-TNF-alpha therapies in systemic lupus erythematosus. J. Biomed. Biotechnol. 2010, 2010, 465898. [Google Scholar] [CrossRef] [PubMed]
- Grisius, M.M.; Bermudez, D.K.; Fox, P.C. Salivary and serum interleukin 6 in primary Sjögren’s syndrome. J. Rheumatol. 1997, 24, 1089–1091. [Google Scholar]
- Youinou, P.; Pers, J.O. Disturbance of cytokine networks in Sjögren’s syndrome. Arthritis Res. Ther. 2011, 13, 227. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, Y.; Takeuchi, T. An update on the pathogenic role of IL-6 in rheumatic diseases. Cytokine 2021, 146, 155645. [Google Scholar] [CrossRef]
- Verstappen, G.M.; Kroese, F.G.M.; Bootsma, H. T cells in primary Sjögren’s syndrome: Targets for early intervention. Rheumatology 2021, 60, 3088–3098. [Google Scholar] [CrossRef]
- Li, H.; Boulougoura, A.; Endo, Y.; Tsokos, G.C. Abnormalities of T cells in systemic lupus erythematosus: New insights in pathogenesis and therapeutic strategies. J. Autoimmun. 2022, 132, 102870. [Google Scholar] [CrossRef]
- Yang, P.; Qian, F.Y.; Zhang, M.F.; Xu, A.L.; Wang, X.; Jiang, B.P.; Zhou, L.L. Th17 cell pathogenicity and plasticity in rheumatoid arthritis. J. Leukoc. Biol. 2019, 106, 1233–1240. [Google Scholar] [CrossRef]
- St Clair, E.W.; Levesque, M.C.; Prak, E.T.; Vivino, F.B.; Alappatt, C.J.; Spychala, M.E.; Wedgwood, J.; McNamara, J.; Moser Sivils, K.L.; Fisher, L.; et al. Autoimmunity Centers of Excellence Rituximab therapy for primary Sjögren’s syndrome: An open-label clinical trial and mechanistic analysis. Arthritis Rheum. 2013, 65, 1097–1106. [Google Scholar] [CrossRef] [PubMed]
- Jakez-Ocampo, J.; Atisha-Fregoso, Y.; Llorente, L. Refractory primary Sjögren syndrome successfully treated with bortezomib. J. Clin. Rheumatol. Pract. Rep. Rheum. Musculoskelet. Dis. 2015, 21, 31–32. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.W.; Lee, S.T.; Kim, T.J.; Jun, J.S.; Chu, K. Bortezomib treatment for severe refractory anti-NMDA receptor encephalitis. Ann. Clin. Transl. Neurol. 2018, 5, 598–605. [Google Scholar] [CrossRef] [PubMed]
- Felten, R.; Devauchelle-Pensec, V.; Seror, R.; Duffau, P.; Saadoun, D.; Hachulla, E.; Pierre Yves, H.; Salliot, C.; Perdriger, A.; Morel, J.; et al. Interleukin 6 receptor inhibition in primary Sjögren syndrome: A multicentre double-blind randomised placebo-controlled trial. Ann. Rheum. Dis. 2021, 80, 329–338. [Google Scholar] [CrossRef]
- Eyre, H.; Siddarth, P.; Cyr, N.; Yang, H.; Cole, S.; Forbes, M.; Lavretsky, H. Comparing the Immune-Genomic Effects of Vilazodone and Paroxetine in Late-Life Depression: A Pilot Study. Pharmacopsychiatry 2017, 50, 256–263. [Google Scholar] [CrossRef]
- Hansen, A.; Reiter, K.; Ziprian, T.; Jacobi, A.; Hoffmann, A.; Gosemann, M.; Scholze, J.; Lipsky, P.E.; Dörner, T. Dysregulation of chemokine receptor expression and function by B cells of patients with primary Sjögren’s syndrome. Arthritis Rheum. 2005, 52, 2109–2119. [Google Scholar] [CrossRef]
- Peng, L.; Zhu, N.; Mao, J.; Huang, L.; Yang, Y.; Zhou, Z.; Wang, L.; Wu, B. Expression levels of CXCR4 and CXCL12 in patients with rheumatoid arthritis and its correlation with disease activity. Exp. Ther. Med. 2020, 20, 1925–1934. [Google Scholar] [CrossRef]
- Hanaoka, H.; Okazaki, Y.; Hashiguchi, A.; Yasuoka, H.; Takeuchi, T.; Kuwana, M. Overexpression of CXCR4 on circulating B cells in patients with active systemic lupus erythematosus. Clin. Exp. Rheumatol. 2015, 33, 863–870. [Google Scholar]
- Bruno, K.A.; Morales-Lara, A.C.; Bittencourt, E.B.; Siddiqui, H.; Bommarito, G.; Patel, J.; Sousou, J.M.; Salomon, G.R.; Paloka, R.; Watford, S.T.; et al. Sex differences in comorbidities associated with Sjögren’s disease. Front. Med. 2022, 9, 958670. [Google Scholar] [CrossRef]
- Maranini, B.; Bortoluzzi, A.; Silvagni, E.; Govoni, M. Focus on Sex and Gender: What We Need to Know in the Management of Rheumatoid Arthritis. J. Pers. Med. 2022, 12, 499. [Google Scholar] [CrossRef]
- Vieira, A.A.; Almada-Correia, I.; Inácio, J.; Costa-Reis, P.; da Rocha, S.T. Female-bias in systemic lupus erythematosus: How much is the X chromosome to blame? Biol. Sex Differ. 2024, 15, 76. [Google Scholar] [CrossRef]
- Panimolle, F.; Tiberti, C.; Spaziani, M.; Riitano, G.; Lucania, G.; Anzuini, A.; Lenzi, A.; Gianfrilli, D.; Sorice, M.; Radicioni, A.F. Non-organ-specific autoimmunity in adult 47,XXY Klinefelter patients and higher-grade X-chromosome aneuploidies. Clin. Exp. Immunol. 2021, 205, 316–325. [Google Scholar] [CrossRef]
- Liu, K.; Kurien, B.T.; Zimmerman, S.L.; Kaufman, K.M.; Taft, D.H.; Kottyan, L.C.; Lazaro, S.; Weaver, C.A.; Ice, J.A.; Adler, A.J.; et al. X Chromosome Dose and Sex Bias in Autoimmune Diseases: Increased Prevalence of 47,XXX in Systemic Lupus Erythematosus and Sjögren’s Syndrome. Arthritis Rheumatol. 2016, 68, 1290–1300. [Google Scholar] [CrossRef]
- Cooney, C.M.; Bruner, G.R.; Aberle, T.; Namjou-Khales, B.; Myers, L.K.; Feo, L.; Li, S.; D’Souza, A.; Ramirez, A.; Harley, J.B.; et al. 46,X,del(X)(q13) Turner’s syndrome women with systemic lupus erythematosus in a pedigree multiplex for SLE. Genes Immun. 2009, 10, 478–481. [Google Scholar] [CrossRef]
- Scofield, R.H.; Bruner, G.R.; Namjou, B.; Kimberly, R.P.; Ramsey-Goldman, R.; Petri, M.; Reveille, J.D.; Alarcón, G.S.; Vilá, L.M.; Reid, J.; et al. Klinefelter’s syndrome (47,XXY) in male systemic lupus erythematosus patients: Support for the notion of a gene-dose effect from the X chromosome. Arthritis Rheum. 2008, 58, 2511–2517. [Google Scholar] [CrossRef]
- Odhams, C.A.; Roberts, A.L.; Vester, S.K.; Duarte, C.S.T.; Beales, C.T.; Clarke, A.J.; Lindinger, S.; Daffern, S.J.; Zito, A.; Chen, L.; et al. Interferon inducible X-linked gene CXorf21 may contribute to sexual dimorphism in Systemic Lupus Erythematosus. Nat. Commun. 2019, 10, 2164. [Google Scholar] [CrossRef]
- Harris, V.M.; Harley, I.T.W.; Kurien, B.T.; Koelsch, K.A.; Scofield, R.H. Lysosomal pH Is Regulated in a Sex Dependent Manner in Immune Cells Expressing CXorf21. Front. Immunol. 2019, 10, 578. [Google Scholar] [CrossRef]
- Groom, J.R.; Luster, A.D. CXCR3 ligands: Redundant, collaborative and antagonistic functions. Immunol. Cell Biol. 2011, 89, 207–215. [Google Scholar] [CrossRef]
- Roberts, A.L.; Morea, A.; Amar, A.; Zito, A.; El-Sayed Moustafa, J.S.; Tomlinson, M.; Bowyer, R.C.E.; Zhang, X.; Christiansen, C.; Costeira, R.; et al. Age acquired skewed X chromosome inactivation is associated with adverse health outcomes in humans. eLife 2022, 11, e78263. [Google Scholar] [CrossRef]
- Mayer, M.; Fey, K.; Heinze, E.; Wick, C.R.; Abboud, M.I.; Yeh, T.L.; Tumber, A.; Orth, N.; Schley, G.; Buchholz, B.; et al. A Fluorescent Benzo[g]isoquinoline-Based HIF Prolyl Hydroxylase Inhibitor for Cellular Imaging. ChemMedChem 2019, 14, 94–99. [Google Scholar] [CrossRef]
- Kou, Y.; Xu, X.; Zhu, Z.; Dai, L.; Tan, Y. Microbe-set enrichment analysis facilitates functional interpretation of microbiome profiling data. Sci. Rep. 2020, 10, 21466. [Google Scholar] [CrossRef]
- Integrative HMP (iHMP) Research Network Consortium. The Integrative Human Microbiome Project. Nature 2019, 569, 641–648. [Google Scholar] [CrossRef]
- PostgreSQLGlobal Development Group. PostgreSQL, version 16; PostgreSQLGlobal Development Group: Santa Barbara, CA, USA; Available online: http://www.postgresql.org/ (accessed on 3 October 2023).
- Snowflake Inc. Streamlit, version 1.27.2; Snowflake Inc.: Bozeman, MT, USA; Available online: https://streamlit.io/ (accessed on 12 November 2023).
- Hunter, J.D. Matplotlib: A 2D Graphics Environment. Comput. Sci. Eng. 2007, 9, 90–95. [Google Scholar] [CrossRef]
- Plotly Technologies Inc. Collaborative Data Science; Plotly Technologies Inc.: Montréal, QC, Canada, 2015; Available online: https://plot.ly (accessed on 3 October 2023).
- Miniconda, Version 2-2.4.0; Anaconda Software Distribution. 2016. Available online: https://anaconda.com (accessed on 3 October 2023).
- The Pip Developers. Pip, Version 25.0.1. Available online: https://pip.pypa.io (accessed on 3 October 2023).
- The Psycopg Team. Psycopg, Version 2.9.10. Available online: https://www.psycopg.org (accessed on 3 October 2023).
- McKinney, W. Pandas: A Foundational Python Library for Data Analysis and Statistics. 2011. Available online: https://www.pandas.pydata.org (accessed on 3 October 2023).
- Harris, C.R.; Millman, K.J.; van der Walt, S.J.; Gommers, R.; Virtanen, P.; Cournapeau, D.; Wieser, E.; Taylor, J.; Berg, S.; Smith, N.J.; et al. Array programming with NumPy. Nature 2020, 585, 357–362. [Google Scholar] [CrossRef]
- Nastou, K.C.; Nasi, G.I.; Tsiolaki, P.L.; Litou, Z.I.; Iconomidou, V.A. AmyCo: The amyloidoses collection. Amyloid Int. J. Exp. Clin. Investig. Off. J. Int. Soc. Amyloidosis 2019, 26, 112–117. [Google Scholar] [CrossRef]
- MedlinePlus [Internet]; National Library of Medicine (US): Bethesda, MD, USA, 2024. Available online: https://medlineplus.gov/ (accessed on 1 July 2020).
- Yang, J.J.; Grissa, D.; Lambert, C.G.; Bologa, C.G.; Mathias, S.L.; Waller, A.; Wild, D.J.; Jensen, L.J.; Oprea, T.I. TIGA: Target illumination GWAS analytics. Bioinformatics 2021, 37, 3865–3873. [Google Scholar] [CrossRef]
- UniProt Consortium. UniProt: The Universal Protein Knowledgebase in 2023. Nucleic Acids Res. 2023, 51, D523–D531. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| KEGG Pathway a | Pathway ID b | Total # Genes c | # Genes Included in Network d | Genes Included from Analysis in Network e | FDR f |
|---|---|---|---|---|---|
| Antifolate resistance | hsa01523 | 31 | 2 | IL6; TNF | 0.0017 |
| Graft-versus-host disease | hsa05332 | 36 | 2 | IL6; TNF | 0.0021 |
| African trypanosomiasis | hsa05143 | 36 | 2 | IL6; TNF | 0.0021 |
| Primary immunodeficiency | hsa05340 | 37 | 2 | CD4; CD8A | 0.0021 |
| Inflammatory bowel disease | hsa05321 | 59 | 3 | IL6; IL17A; TNF | 6.91 × 10−5 |
| Rheumatoid arthritis | hsa05323 | 83 | 4 | IL6; IL17A; TNF; TNFSF13B | 4.13 × 10−6 |
| Antigen processing and presentation | hsa04612 | 64 | 3 | CD4; CD8A; TNF | 7.29 × 10−5 |
| Intestinal immune network for IgA production | hsa04672 | 43 | 2 | IL6; TNFSF13B | 0.0024 |
| Hematopoietic cell lineage | hsa04640 | 90 | 4 | CD4; CD8A; IL6; TNF | 4.13 × 10−6 |
| Malaria | hsa05144 | 46 | 2 | IL6; TNF | 0.0025 |
| Legionellosis | hsa05134 | 55 | 2 | IL6; TNF | 0.0034 |
| IL-17 signaling pathway | hsa04657 | 91 | 3 | IL6; IL17A; TNF | 0.00017 |
| Yersinia infection | hsa05135 | 124 | 4 | CD4; CD8A; IL6; TNF | 4.92 × 10−6 |
| T cell receptor signaling pathway | hsa04660 | 100 | 3 | CD4; CD8A; TNF | 0.00019 |
| Th17 cell differentiation | hsa04659 | 99 | 3 | CD4; IL6; IL17A | 0.00019 |
| Pertussis | hsa05133 | 73 | 2 | IL6; TNF | 0.0055 |
| Hypertrophic cardiomyopathy | hsa05410 | 88 | 2 | IL6; TNF | 0.0075 |
| Systemic lupus erythematosus | hsa05322 | 94 | 2 | TNF; TRIM21 | 0.0082 |
| AGE-RAGE signaling pathway in diabetic complications | hsa04933 | 96 | 2 | IL6; TNF | 0.0082 |
| Viral protein interaction with cytokine and cytokine receptor | hsa04061 | 96 | 2 | IL6; TNF | 0.0082 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beckman, M.F.; Alexander, A.; Mougeot, J.-L.C.; Bahrani Mougeot, F. GeneHarmony: A Knowledge-Based Tool for Biomarker Discovery in Disease: Sjögren’s Disease vs. Rheumatoid Arthritis and Systemic Lupus Erythematosus. Int. J. Mol. Sci. 2025, 26, 6379. https://doi.org/10.3390/ijms26136379
Beckman MF, Alexander A, Mougeot J-LC, Bahrani Mougeot F. GeneHarmony: A Knowledge-Based Tool for Biomarker Discovery in Disease: Sjögren’s Disease vs. Rheumatoid Arthritis and Systemic Lupus Erythematosus. International Journal of Molecular Sciences. 2025; 26(13):6379. https://doi.org/10.3390/ijms26136379
Chicago/Turabian StyleBeckman, Micaela F., Adam Alexander, Jean-Luc C. Mougeot, and Farah Bahrani Mougeot. 2025. "GeneHarmony: A Knowledge-Based Tool for Biomarker Discovery in Disease: Sjögren’s Disease vs. Rheumatoid Arthritis and Systemic Lupus Erythematosus" International Journal of Molecular Sciences 26, no. 13: 6379. https://doi.org/10.3390/ijms26136379
APA StyleBeckman, M. F., Alexander, A., Mougeot, J.-L. C., & Bahrani Mougeot, F. (2025). GeneHarmony: A Knowledge-Based Tool for Biomarker Discovery in Disease: Sjögren’s Disease vs. Rheumatoid Arthritis and Systemic Lupus Erythematosus. International Journal of Molecular Sciences, 26(13), 6379. https://doi.org/10.3390/ijms26136379