
Cloning, Expression and Functional Characterization of V. vinifera CAT2 Arginine Transporter




 and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Cloning, Expression and Purification
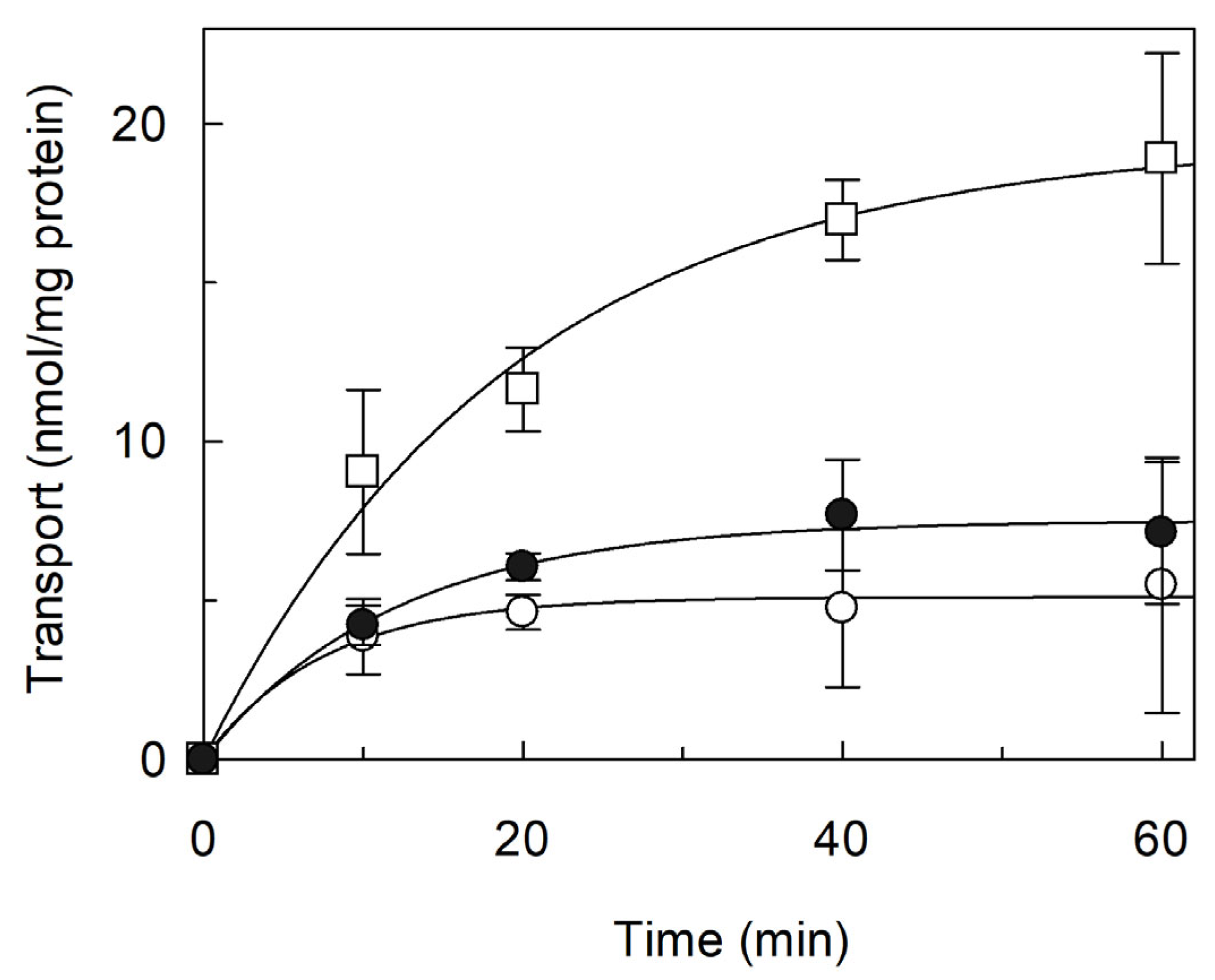
2.2. Functional Characterization and Regulation
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. RNA Extraction, cDNA Production and Cloning of VvCAT2
4.3. Protein Production
4.4. Protein Purification
4.5. Reconstitution of the SlCAT2 Transporter into Liposomes
4.6. Transport Measurements
4.7. Other Methods
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| APC | Amino acid-Polyamine/Choline |
| ATF | Amino acid Transporter superFamily |
| BTA | Benzyltriethylammonium |
| CAT | Cation Amino acid Transporter |
| CHS | Cholesteryl HemiSuccinate |
| Cu-Phe | Copper Phenanthroline |
| DDM | n-Dodecyl-β-D-Maltoside |
| DTE | Dithioerythritol |
| EDTA | Ethylenediaminetetraacetic Acid |
| IPTG | Isopropil-β-D-tiogalattopiranoside |
| TEA | Tetraethylammonium |
| TMA | Tetramethylammonium |
References
- Kuang, L.; Chen, S.; Guo, Y.; Ma, H. Quantitative Proteome Analysis Reveals Changes in the Protein Landscape During Grape Berry Development With a Focus on Vacuolar Transport Proteins. Front. Plant Sci. 2019, 10, 641. [Google Scholar] [CrossRef]
- Averilla, J.N.; Oh, J.; Wu, Z.; Liu, K.H.; Jang, C.H.; Kim, H.J.; Kim, J.S.; Kim, J.S. Improved extraction of resveratrol and antioxidants from grape peel using heat and enzymatic treatments. J. Sci. Food Agric. 2019, 99, 4043–4053. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.; Cai, L.; Udeani, G.O.; Slowing, K.V.; Thomas, C.F.; Beecher, C.W.; Fong, H.H.; Farnsworth, N.R.; Kinghorn, A.D.; Mehta, R.G.; et al. Cancer chemopreventive activity of resveratrol, a natural product derived from grapes. Science 1997, 275, 218–220. [Google Scholar] [CrossRef] [PubMed]
- Brown, K.; Theofanous, D.; Britton, R.G.; Aburido, G.; Pepper, C.; Sri Undru, S.; Howells, L. Resveratrol for the Management of Human Health: How Far Have We Come? A Systematic Review of Resveratrol Clinical Trials to Highlight Gaps and Opportunities. Int. J. Mol. Sci. 2024, 25, 747. [Google Scholar] [CrossRef]
- Yao, X.; Nie, J.; Bai, R.; Sui, X. Amino Acid Transporters in Plants: Identification and Function. Plants 2020, 9, 972. [Google Scholar] [CrossRef] [PubMed]
- Etxeberria, E.; Pozueta-Romero, J.; Gonzalez, P. In and out of the plant storage vacuole. Plant Sci. 2012, 190, 52–61. [Google Scholar] [CrossRef]
- Carter, C.; Pan, S.; Zouhar, J.; Avila, E.L.; Girke, T.; Raikhel, N.V. The vegetative vacuole proteome of Arabidopsis thaliana reveals predicted and unexpected proteins. Plant Cell 2004, 16, 3285–3303. [Google Scholar] [CrossRef]
- Schumacher, K. pH in the plant endomembrane system-an import and export business. Curr. Opin. Plant Biol. 2014, 22, 71–76. [Google Scholar] [CrossRef]
- Gao, C.; Zhao, Q.; Jiang, L. Vacuoles protect plants from high magnesium stress. Proc. Natl. Acad. Sci. USA 2015, 112, 2931–2932. [Google Scholar] [CrossRef]
- Martinoia, E.; Meyer, S.; De Angeli, A.; Nagy, R. Vacuolar transporters in their physiological context. Annu. Rev. Plant Biol. 2012, 63, 183–213. [Google Scholar] [CrossRef]
- Hildebrandt, T.M.; Nunes Nesi, A.; Araujo, W.L.; Braun, H.P. Amino Acid Catabolism in Plants. Mol. Plant 2015, 8, 1563–1579. [Google Scholar] [CrossRef] [PubMed]
- Patil, S.S.; Prashant, R.; Kadoo, N.Y.; Upadhyay, A.; Gupta, V.S. Global study of MFS superfamily transporters in arabidopsis and grapes reveals their functional diversity in plants. Plant Gene 2019, 18, 100179. [Google Scholar] [CrossRef]
- Yang, G.; Wei, Q.; Huang, H.; Xia, J. Amino Acid Transporters in Plant Cells: A Brief Review. Plants 2020, 9, 967. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Zhu, X.; Shen, J.; Xing, H.; Zou, Z.; Ma, Y.; Wang, Y.; Fang, W. Genome-wide identification, characterization and expression analysis of the amino acid permease gene family in tea plants (Camellia sinensis). Genomics 2020, 112, 2866–2874. [Google Scholar] [CrossRef] [PubMed]
- Yuan, D.; Wu, X.; Jiang, X.; Gong, B.; Gao, H. Types of Membrane Transporters and the Mechanisms of Interaction between Them and Reactive Oxygen Species in Plants. Antioxidants 2024, 13, 221. [Google Scholar] [CrossRef]
- Chen, X.; Wu, Z.; Yin, Z.; Zhang, Y.; Rui, C.; Wang, J.; Malik, W.A.; Lu, X.; Wang, D.; Wang, J.; et al. Comprehensive genomic characterization of cotton cationic amino acid transporter genes reveals that GhCAT10D regulates salt tolerance. BMC Plant Biol. 2022, 22, 441. [Google Scholar] [CrossRef]
- Wan, Y.; King, R.; Mitchell, R.A.C.; Hassani-Pak, K.; Hawkesford, M.J. Spatiotemporal expression patterns of wheat amino acid transporters reveal their putative roles in nitrogen transport and responses to abiotic stress. Sci. Rep. 2017, 7, 5461. [Google Scholar] [CrossRef]
- Feng, L.; Yu, Y.; Lin, S.; Yang, T.; Chen, Q.; Liu, L.; Sun, J.; Zheng, P.; Zhang, Z.; Wan, X. Tonoplast-Localized Theanine Transporter CsCAT2 May Mediate Theanine Storage in the Root of Tea Plants (Camellia sinensis L.). Front. Plant Sci. 2021, 12, 797854. [Google Scholar] [CrossRef]
- Yang, H.; Krebs, M.; Stierhof, Y.D.; Ludewig, U. Characterization of the putative amino acid transporter genes AtCAT2, 3 &4: The tonoplast localized AtCAT2 regulates soluble leaf amino acids. J. Plant Physiol. 2014, 171, 594–601. [Google Scholar]
- Islam, M.N.; Rabby, M.G.; Hossen, M.M.; Bonny, M.; Hasan, M.M. Genome-wide identification following functional analysis of amino acid permease and cationic amino acid transporter gene families in maize and their role in drought stress. S. Afr. J. Bot. 2024, 168, 360–371. [Google Scholar] [CrossRef]
- Liu, L.; Yu, X.C.; Yan, Y.; He, C.X.; Wang, J.; Sun, M.T.; Li, Y.S. Amino Acid Transporters on Amino Acid Absorption, Transport and Distribution in Crops. Horticulturae 2024, 10, 999. [Google Scholar] [CrossRef]
- Regina, T.M.R.; Galluccio, M.; Scalise, M.; Pochini, L.; Indiveri, C. Bacterial production and reconstitution in proteoliposomes of Solanum lycopersicum CAT2: A transporter of basic amino acids and organic cations. Plant Mol. Biol. 2017, 94, 657–667. [Google Scholar] [CrossRef] [PubMed]
- Du, X.Q.; Sun, S.S.; Zhou, T.; Zhang, L.; Feng, Y.N.; Zhang, K.L.; Hua, Y.P. Genome-Wide Identification of the CAT Genes and Molecular Characterization of Their Transcriptional Responses to Various Nutrient Stresses in Allotetraploid Rapeseed. Int. J. Mol. Sci. 2024, 25, 12658. [Google Scholar] [CrossRef]
- Feng, L.; Yang, T.Y.; Zhang, Z.L.; Li, F.D.; Chen, Q.; Sun, J.; Shi, C.Y.; Deng, W.W.; Tao, M.M.; Tai, Y.L.; et al. Identification and characterization of cationic amino acid transporters (CATs) in tea plant (Camellia sinensis). Plant Growth Regul. 2018, 84, 57–69. [Google Scholar] [CrossRef]
- Yang, H.; Stierhof, Y.D.; Ludewig, U. The putative Cationic Amino Acid Transporter 9 is targeted to vesicles and may be involved in plant amino acid homeostasis. Front. Plant Sci. 2015, 6, 212. [Google Scholar] [CrossRef] [PubMed]
- Martinoia, E. Vacuolar Transporters—Companions on a Longtime Journey. Plant Physiol. 2018, 176, 1384–1407. [Google Scholar] [CrossRef]
- Rogiers, S.Y.; Coetzee, Z.A.; Walker, R.R.; Deloire, A.; Tyerman, S.D. Potassium in the Grape (Vitis vinifera L.) Berry: Transport and Function. Front. Plant Sci. 2017, 8, 1629. [Google Scholar] [CrossRef]
- Shitan, N.; Yazaki, K. Dynamism of vacuoles toward survival strategy in plants. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183127. [Google Scholar] [CrossRef]
- Rosano, G.L.; Ceccarelli, E.A. Recombinant protein expression in Escherichia coli: Advances and challenges. Front. Microbiol. 2014, 5, 172. [Google Scholar] [CrossRef]
- Galluccio, M.; Mazza, T.; Scalise, M.; Tripicchio, M.; Scarpelli, M.; Tolomeo, M.; Pochini, L.; Indiveri, C. Over-Production of the Human SLC7A10 in E. coli and Functional Assay in Proteoliposomes. Int. J. Mol. Sci. 2023, 25, 536. [Google Scholar] [CrossRef]
- Galluccio, M.; Tripicchio, M.; Console, L.; Indiveri, C. Bacterial over-production of the functionally active human SLC38A2 (SNAT2) exploiting the mistic tag: A cheap and fast tool for testing ligands. Mol. Biol. Rep. 2024, 51, 336. [Google Scholar] [CrossRef] [PubMed]
- Dilworth, M.V.; Findlay, H.E.; Booth, P.J. Detergent-free purification and reconstitution of functional human serotonin transporter (SERT) using diisobutylene maleic acid (DIBMA) copolymer. Biochim. Biophys. Acta Biomembr. 2021, 1863, 183602. [Google Scholar] [CrossRef] [PubMed]
- Yan, R.; Zhao, X.; Lei, J.; Zhou, Q. Structure of the human LAT1-4F2hc heteromeric amino acid transporter complex. Nature 2019, 568, 127–130. [Google Scholar] [CrossRef]
- Sonawane, P.D.; Pollier, J.; Panda, S.; Szymanski, J.; Massalha, H.; Yona, M.; Unger, T.; Malitsky, S.; Arendt, P.; Pauwels, L.; et al. Plant cholesterol biosynthetic pathway overlaps with phytosterol metabolism. Nat. Plants 2016, 3, 16205. [Google Scholar] [CrossRef]
- Kumar, K.; Gibbs, H.C.; Yeh, A.T.; Griffing, L.R. The Sterol Trafficking Pathway in Arabidopsis thaliana. Front. Plant Sci. 2021, 12, 616631. [Google Scholar] [CrossRef]
- Cosco, J.; Regina, T.M.R.; Scalise, M.; Galluccio, M.; Indiveri, C. Regulatory Aspects of the Vacuolar CAT2 Arginine Transporter of S. lycopersicum: Role of Osmotic Pressure and Cations. Int. J. Mol. Sci. 2019, 20, 906. [Google Scholar] [CrossRef]
- Pochini, L.; Pappacoda, G.; Galluccio, M.; Pastore, F.; Scalise, M.; Indiveri, C. Effect of Cholesterol on the Organic Cation Transporter OCTN1 (SLC22A4). Int. J. Mol. Sci. 2020, 21, 1091. [Google Scholar] [CrossRef] [PubMed]
- Scalise, M.; Pochini, L.; Cosco, J.; Aloe, E.; Mazza, T.; Console, L.; Esposito, A.; Indiveri, C. Interaction of Cholesterol With the Human SLC1A5 (ASCT2): Insights Into Structure/Function Relationships. Front. Mol. Biosci. 2019, 6, 110. [Google Scholar] [CrossRef]
- Dickens, D.; Chiduza, G.N.; Wright, G.S.; Pirmohamed, M.; Antonyuk, S.V.; Hasnain, S.S. Modulation of LAT1 (SLC7A5) transporter activity and stability by membrane cholesterol. Sci. Rep. 2017, 7, 43580. [Google Scholar] [CrossRef]
- Endler, A.; Meyer, S.; Schelbert, S.; Schneider, T.; Weschke, W.; Peters, S.W.; Keller, F.; Baginsky, S.; Martinoia, E.; Schmidt, U.G. Identification of a vacuolar sucrose transporter in barley and Arabidopsis mesophyll cells by a tonoplast proteomic approach. Plant Physiol. 2006, 141, 196–207. [Google Scholar] [CrossRef]
- Dietz, K.J.; Martinoia, E.; Heber, U. Mobilization of Vacuolar Amino-Acids in Leaf-Cells as Affected by Atp and the Level of Cytosolic Amino-Acids—Atp Regulates but Appears Not to Energize Vacuolar Amino-Acid Release. Biochim. Biophys. Acta 1989, 984, 57–62. [Google Scholar] [CrossRef]
- Savchenko, T.; Tikhonov, K. Oxidative Stress-Induced Alteration of Plant Central Metabolism. Life 2021, 11, 304. [Google Scholar] [CrossRef]
- Tonazzi, A.; Console, L.; Indiveri, C. Inhibition of mitochondrial carnitine/acylcarnitine transporter by H2O2: Molecular mechanism and possible implication in pathophysiology. Chem. Biol. Interact. 2013, 203, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Moses, T.; Goossens, A. Plants for human health: Greening biotechnology and synthetic biology. J. Exp. Bot. 2017, 68, 4009–4011. [Google Scholar] [CrossRef]
- Guo, X.; He, N.; Huang, B.; Chen, C.; Zhang, Y.; Yang, X.; Li, J.; Dong, Z. Genome-Wide Identification and Expression Analysis of Amino Acid/Auxin Permease (AAAP) Genes in Grapes (Vitis vinifera L.) Under Abiotic Stress and During Development. Plants 2025, 14, 128. [Google Scholar] [CrossRef]
- Martinoia, E.; Massonneau, A.; Frangne, N. Transport processes of solutes across the vacuolar membrane of higher plants. Plant Cell Physiol. 2000, 41, 1175–1186. [Google Scholar] [CrossRef]
- Mohseni, F.; Pakkish, Z.; Panahi, B. Arginine Impact on Yield and Fruit Qualitative Characteristics of Strawberry. Agric. Conspec. Sci. 2017, 82, 19–26. [Google Scholar]
- Dzhambazova, T.; Kondakova, V.; Tsvetkov, I.; Batchvarova, R. Grape Secondary Metabolites—Benefits for Human Health. In Advanced Understanding of Neurodegenerative Diseases; Chang, R.C.-C., Ed.; InTech: London, UK, 2011; pp. 285–298. [Google Scholar]
- Francisco, R.M.; Regalado, A.; Ageorges, A.; Burla, B.J.; Bassin, B.; Eisenach, C.; Zarrouk, O.; Vialet, S.; Marlin, T.; Chaves, M.M.; et al. ABCC1, an ATP binding cassette protein from grape berry, transports anthocyanidin 3-O-Glucosides. Plant Cell 2013, 25, 1840–1854. [Google Scholar] [CrossRef]
- Perez-Diaz, R.; Ryngajllo, M.; Perez-Diaz, J.; Pena-Cortes, H.; Casaretto, J.A.; Gonzalez-Villanueva, E.; Ruiz-Lara, S. VvMATE1 and VvMATE2 encode putative proanthocyanidin transporters expressed during berry development in Vitis vinifera L. Plant Cell Rep. 2014, 33, 1147–1159. [Google Scholar] [CrossRef]
- Galluccio, M.; Mazza, T.; Scalise, M.; Sarubbi, M.C.; Indiveri, C. Bacterial over-expression of functionally active human CT2 (SLC22A16) carnitine transporter. Mol. Biol. Rep. 2022, 49, 8185–8193. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pochini, L.; Regina, T.M.R.; Cerbelli, M.I.; Gallo, N.; Costantino, F.; Galluccio, M.; Indiveri, C. Cloning, Expression and Functional Characterization of V. vinifera CAT2 Arginine Transporter. Int. J. Mol. Sci. 2025, 26, 6259. https://doi.org/10.3390/ijms26136259
Pochini L, Regina TMR, Cerbelli MI, Gallo N, Costantino F, Galluccio M, Indiveri C. Cloning, Expression and Functional Characterization of V. vinifera CAT2 Arginine Transporter. International Journal of Molecular Sciences. 2025; 26(13):6259. https://doi.org/10.3390/ijms26136259
Chicago/Turabian StylePochini, Lorena, Teresa Maria Rosaria Regina, Maria Iolanda Cerbelli, Nicoletta Gallo, Federica Costantino, Michele Galluccio, and Cesare Indiveri. 2025. "Cloning, Expression and Functional Characterization of V. vinifera CAT2 Arginine Transporter" International Journal of Molecular Sciences 26, no. 13: 6259. https://doi.org/10.3390/ijms26136259
APA StylePochini, L., Regina, T. M. R., Cerbelli, M. I., Gallo, N., Costantino, F., Galluccio, M., & Indiveri, C. (2025). Cloning, Expression and Functional Characterization of V. vinifera CAT2 Arginine Transporter. International Journal of Molecular Sciences, 26(13), 6259. https://doi.org/10.3390/ijms26136259