
Prediction of Rice Plant Height Using Linear Regression Model by Pyramiding Plant Height-Related Alleles
 and
and
Abstract
1. Introduction
2. Results
2.1. Significant Loci Associated with Rice Plant Height Detected by Association Mapping
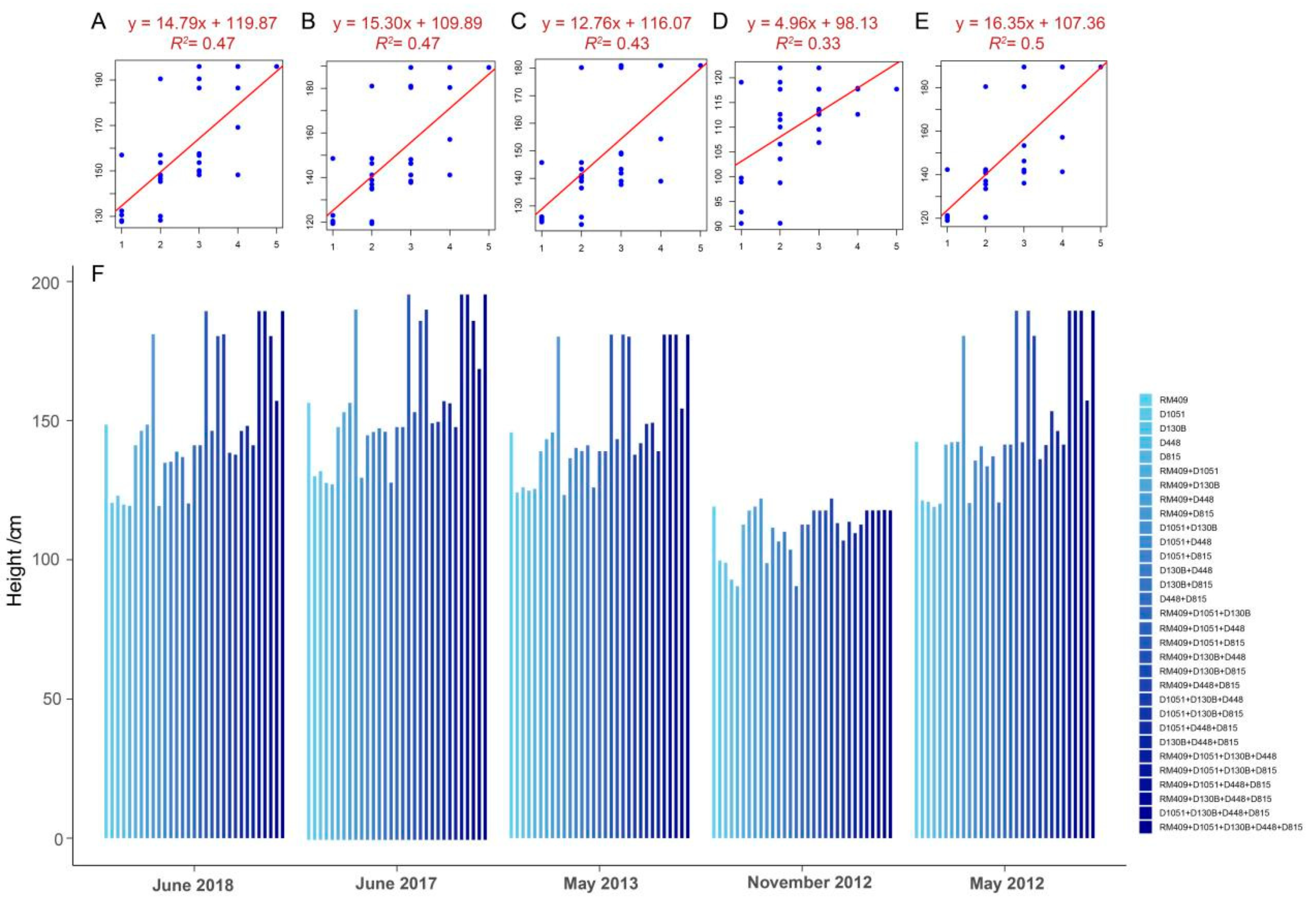
2.2. Pyramiding Plant Height-Related Alleles Could Promote the Plant Height of Rice
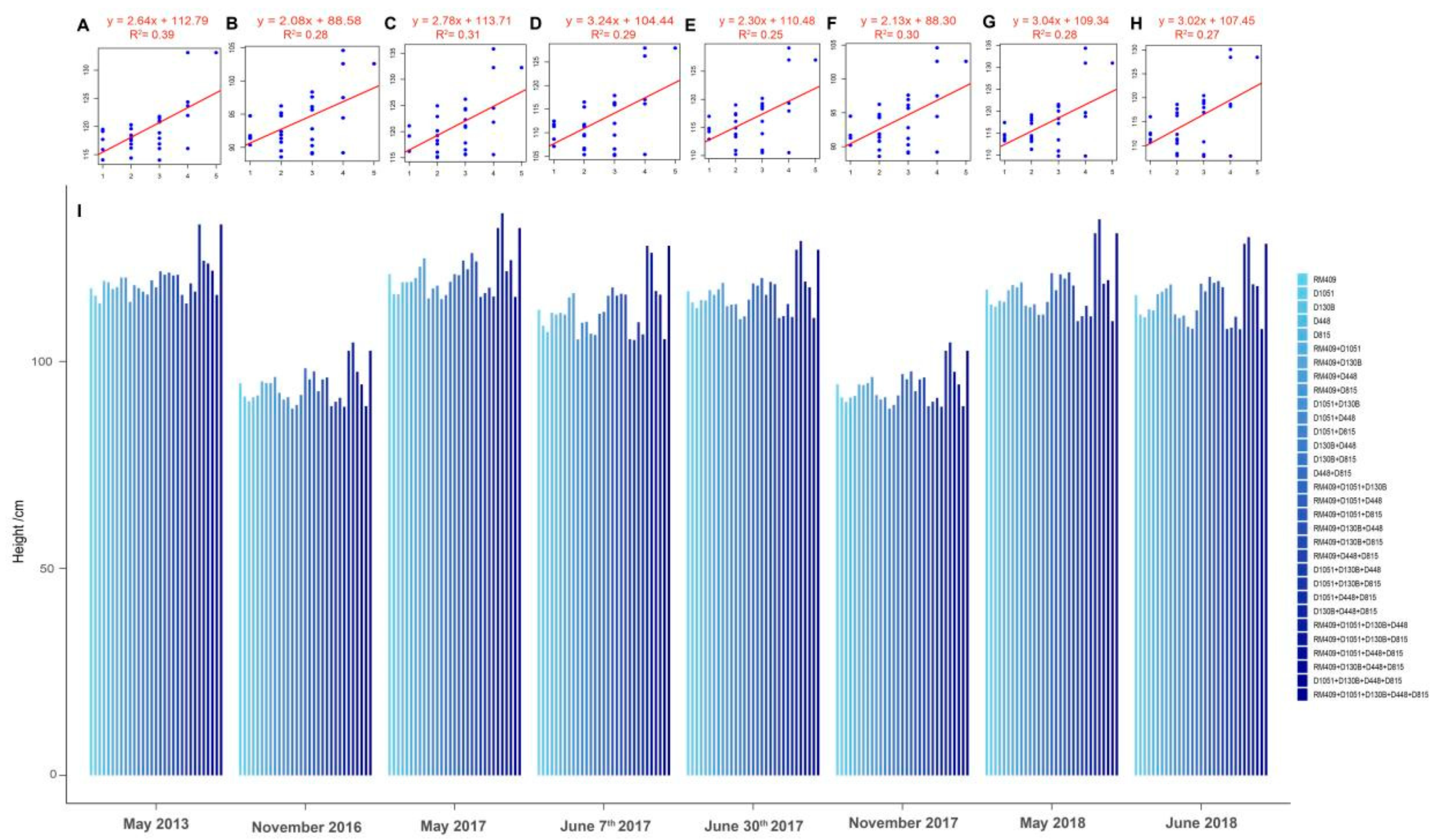
2.3. Confirmation the Effect of the Five Plant Height-Related Loci in Influencing Plant Height by Using Another Recombinant Inbred Line (RIL) Population
2.4. Building Linear Regression Model for Prediction of Rice Plant Height Based on Plant Height-Related Alleles
2.5. Genotype-Environment Interaction Influence Rice Plant Height
3. Discussion
4. Materials and Methods
4.1. Association Mapping Population for Detecting Plant Height-Related Marker Loci and Building Plant Height Prediction Model
4.2. Recombinant Inbred Line Population for Evaluating Plant Height Prediction Model
4.3. Association Mapping Using TASSEL
4.4. Building Linear Regression Model for Prediction of Plant Height
4.5. Evaluation of the Prediction Accuracy of the Prediction Model
4.6. Influence of Interaction Between Genotype and Environment on Plant Height Phenotype
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, L.; Huang, J.; Wang, Y.; Xu, R.; Yang, Z.; Zhao, Z.; Liu, S.; Tian, Y.; Zheng, X.; Li, F.; et al. Identification and genetic analysis of qCL1.2, a novel allele of the “green revolution” gene SD1 from wild rice (Oryza rufipogon) that enhances plant height. BMC Genet. 2020, 21, 62. [Google Scholar] [CrossRef]
- Wei, X.; Xu, J.; Guo, H.; Jiang, L.; Chen, S.; Yu, C.; Zhou, Z.; Hu, P.; Zhai, H.; Wan, J. DTH8 suppresses flowering in rice, influencing plant height and yield potential simultaneously. Plant Physiol. 2010, 153, 1747–1758. [Google Scholar] [CrossRef]
- Liu, F.; Wang, P.; Zhang, X.; Li, X.; Yan, X.; Fu, D.; Wu, G. The genetic and molecular basis of crop height based on a rice model. Planta 2018, 247, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Lu, S.C.; Wang, Y.F.; Zhang, X.; Lv, B.; Luo, L.Q.; Xi, D.D.; Shen, J.B.; Ma, H.; Ming, F. OsNAC2 encoding a NAC transcription factor that affects plant height through mediating the gibberellic acid pathway in rice. Plant J. 2015, 82, 302–314. [Google Scholar] [CrossRef] [PubMed]
- Monna, L.; Kitazawa, N.; Yoshino, R.; Suzuki, J.; Masuda, H.; Maehara, Y.; Tanji, M.; Sato, M.; Nasu, S.; Minobe, Y. Positional cloning of rice semidwarfing gene, sd-1: Rice “green revolution gene” encodes a mutant enzyme involved in gibberellin synthesis. DNA Res. 2002, 9, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Lan, J.; Lin, Q.B.; Zhou, C.L.; Ren, Y.K.; Liu, X.; Miao, R.; Jing, R.N.; Mou, C.L.; Nguyen, T.; Zhu, X.J.; et al. Small grain and semi-dwarf 3, a WRKY transcription factor, negatively regulates plant height and grain size by stabilizing SLR1 expression in rice. Plant Mol. Biol. 2020, 104, 429–450. [Google Scholar] [CrossRef]
- Sasaki, A.; Ashikari, M.; Ueguchi-Tanaka, M.; Itoh, H.; Nishimura, A.; Swapan, D.; Ishiyama, K.; Saito, T.; Kobayashi, M.; Khush, G.S.; et al. Green revolution: A mutant gibberellin-synthesis gene in rice. Nature 2002, 416, 701–702. [Google Scholar] [CrossRef]
- Park, J.R.; Jang, Y.H.; Kim, E.G.; Hur, S.S.; Kim, K.M. Quantitative Trait Loci Mapping Identified Candidate Genes Involved in Plant Height Regulation in Rice. Int. J. Mol. Sci. 2023, 24, 16895. [Google Scholar] [CrossRef]
- Tang, L.Q.; Xu, H.Y.; Wang, Y.F.; Wang, H.M.; Li, Z.Y.; Liu, X.X.; Shu, Y.Z.; Li, G.; Liu, W.N.; Ying, J.Z.; et al. OsABF1 Represses Gibberellin Biosynthesis to Regulate Plant Height and Seed Germination in Rice (Oryza sativa L.). Int. J. Mol. Sci. 2021, 22, 2220. [Google Scholar] [CrossRef]
- Zhang, T.H.; Wang, J.F.; Luo, R.; Man, J.M.; Long, Q.; Xu, N. OsHLS1 regulates plant height and development by controlling active gibberellin accumulation in rice (Oryza sativa L.). Plant Sci. 2023, 326, 111508. [Google Scholar] [CrossRef]
- Umehara, M.; Hanada, A.; Yoshida, S.; Akiyama, K.; Arite, T.; Takeda-Kamiya, N.; Magome, H.; Kamiya, Y.; Shirasu, K.; Yoneyama, K.; et al. Inhibition of shoot branching by new terpenoid plant hormones. Nature 2008, 455, 195–200. [Google Scholar] [CrossRef]
- Gomez-Roldan, V.; Fermas, S.; Brewer, P.B.; Puech-Pagès, V.; Dun, E.A.; Pillot, J.P.; Letisse, F.; Matusova, R.; Danoun, S.; Portais, J.C.; et al. Strigolactone inhibition of shoot branching. Nature 2008, 455, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Mizutori, Y.; Nishiyama, K.; Seto, Y. Inhibition of shoot branching in Arabidopsis by the artificially biosynthesized canonical strigolactone. Biosci. Biotechnol. Biochem. 2025, 89, 406–412. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Wang, R.; Qian, Q.; Yan, M.; Meng, X.; Fu, Z.; Yan, C.; Jiang, B.; Su, Z.; Li, J.; et al. DWARF27, an iron-containing protein required for the biosynthesis of strigolactones, regulates rice tiller bud outgrowth. Plant Cell 2009, 21, 1512–1525. [Google Scholar] [CrossRef] [PubMed]
- Sazuka, T.; Kamiya, N.; Nishimura, T.; Ohmae, K.; Sato, Y.; Imamura, K.; Nagato, Y.; Koshiba, T.; Nagamura, Y.; Ashikari, M.; et al. A rice tryptophan deficient dwarf mutant, tdd1, contains a reduced level of indole acetic acid and develops abnormal flowers and organless embryos. Plant J. 2009, 60, 227–241. [Google Scholar] [CrossRef]
- Dai, Z.; Wang, J.; Yang, X.; Lu, H.; Miao, X.; Shi, Z. Modulation of plant architecture by the miR156f-OsSPL7-OsGH3.8 pathway in rice. J. Exp. Bot. 2018, 69, 5117–5130. [Google Scholar] [CrossRef]
- Lu, G.W.; Coneva, V.; Casaretto, J.A.; Ying, S.; Mahmood, K.; Liu, F.; Nambara, E.; Bi, Y.M.; Rothstein, S.J. OsPIN5b modulates rice (Oryza sativa) plant architecture and yield by changing auxin homeostasis. Plant J. 2015, 83, 913–925. [Google Scholar] [CrossRef]
- Shi, F.C.; Wang, M.; An, Y.X. Overexpression of a B-type cytokinin response regulator (OsORR2) reduces plant height in rice. Plant Signal. Behav. 2020, 15, 1780405. [Google Scholar] [CrossRef]
- Zhang, X.B.; Wang, Y.; Wang, X.W.; Zhu, Z.; Zhang, X.F.; Jia, L.Q.; Li, Y.Y.; Tian, W.J.; Chen, H.Y.; Zhu, X.Y.; et al. A very-long-chain fatty acid synthesis gene, SD38, inflfluences plant height by activating ethylene biosynthesis in rice. Plant J. 2022, 112, 1084–1097. [Google Scholar] [CrossRef]
- Yan, W.H.; Wang, P.; Chen, H.X.; Zhou, H.J.; Li, Q.P.; Wang, C.R.; Ding, Z.H.; Zhang, Y.S.; Yu, S.B.; Xing, Y.Z.; et al. A major QTL, Ghd8, plays pleiotropic roles in regulating grain productivity, plant height, and heading date in rice. Mol. Plant 2011, 4, 319–330. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, C.; Lin, J.; Liu, J.; Liu, B.; Wang, J.; Huang, A.; Li, H.; Zhao, T. OsMPH1 regulates plant height and improves grain yield in rice. PLoS ONE 2017, 12, e0180825. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Zhou, H.; Xie, D.; Li, J.; Yang, M.; Chang, T.; Wang, D.; Hu, L.; Xie, G.; Wang, J.; et al. Genome-Wide Association Study in Rice Revealed a Novel Gene in Determining Plant Height and Stem Development, by Encoding a WRKY Transcription Factor. Int. J. Mol. Sci. 2021, 22, 8192. [Google Scholar] [CrossRef]
- Yan, P.; Zhu, Y.; Wang, Y.; Ma, F.; Lan, D.; Niu, F.; Dong, S.; Zhang, X.; Hu, J.; Liu, S.; et al. A New RING Finger Protein, PLANT ARCHITECTURE and GRAIN NUMBER 1, Affects Plant Architecture and Grain Yield in Rice. Int. J. Mol. Sci. 2022, 23, 824. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Zhang, S.; Luo, Y.; Li, F.; Tan, J.; Wang, B.; Zhao, Z.; Lin, H.; Zhang, T.; Liu, J.; et al. Rice OsUBR7 modulates plant height by regulating histone H2B monoubiquitination and cell proliferation. Plant Commun. 2022, 3, 100412. [Google Scholar] [CrossRef]
- Kurotani, K.I.; Hattori, T.; Takeda, S. Overexpression of a CYP94 family gene CYP94C2b increases internode length and plant height in rice. Plant Signal. Behav. 2015, 10, e1046667. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.T.; Chen, P.W.; Chen, L.C.; Yang, C.C.; Chen, S.Y.; Huang, G.; Lin, T.C.; Ku, H.M.; Chen, J.J.W. Suppressive effect of microRNA319 expression on rice plant height. Theor. Appl. Genet. 2017, 130, 1507–1518. [Google Scholar] [CrossRef]
- Xin, W.; Liu, H.; Yang, L.; Ma, T.; Wang, J.; Zheng, H.; Liu, W.; Zou, D. BSA-Seq and Fine Linkage Mapping for the Identification of a Novel Locus (qPH9) for Mature Plant Height in Rice (Oryza sativa). Rice 2022, 15, 26. [Google Scholar] [CrossRef]
- Wang, H.; Wang, G.; Qin, R.; Gong, C.; Zhou, D.; Li, D.; Luo, B.; Jin, J.; Deng, Q.; Wang, S.; et al. Improvement of Quality and Disease Resistance for a Heavy-Panicle Hybrid Restorer Line, R600, in Rice (Oryza sativa L.) by Gene Pyramiding Breeding. Curr. Issues Mol. Biol. 2024, 46, 10762–10778. [Google Scholar] [CrossRef]
- Abe, T.; Ito, M.; Takahashi, R.; Honma, T.; Kuramata, M.; Ishikawa, S. QTL Pyramiding and Its Use in Breeding for Increasing the Phytoextraction Efficiency of Soil Cd via High-Cd-Accumulating Rice. Plants 2022, 11, 2178. [Google Scholar] [CrossRef]
- Lin, C.; Zhu, T.; Ogorek, L.L.P.; Wang, Y.; Sauter, M.; Pedersen, O. The Pyramiding of Three Key Root Traits Aid Breeding of Flood-Tolerant Rice. Plants 2022, 11, 2033. [Google Scholar] [CrossRef]
- Luo, J.; Li, S.; Xu, J.; Yan, L.; Ma, Y.; Xia, L. Pyramiding favorable alleles in an elite wheat variety in one generation by CRISPR-Cas9-mediated multiplex gene editing. Mol. Plant 2021, 14, 847–850. [Google Scholar] [CrossRef] [PubMed]
- Gholipourshahraki, T.; Bai, Z.; Shrestha, M.; Hjelholt, A.; Hu, S.; Kjolby, M.; Rohde, P.D.; Sorensen, P. Evaluation of Bayesian Linear Regression models for gene set prioritization in complex diseases. PLoS Genet. 2024, 20, e1011463. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Dong, J.; Ji, Z.; Yang, C.; Liang, Y. A Linear Regression Model for the Prediction of Rice Sheath Blight Field Resistance. Plant Dis. 2021, 105, 2964–2969. [Google Scholar] [CrossRef]
- Zhao, H.; Liu, X.; Wang, J.; Qian, Q.; Zhang, G. The coordinated regulation mechanism of rice plant architecture and its tolerance to stress. Front. Plant Sci. 2022, 13, 1087378. [Google Scholar] [CrossRef]
- Jiao, Y.; Wang, Y.; Xue, D.; Wang, J.; Yan, M.; Liu, G.; Dong, G.; Zeng, D.; Lu, Z.; Zhu, X.; et al. Regulation of OsSPL14 by OsmiR156 defines ideal plant architecture in rice. Nat. Genet. 2010, 42, 541–544. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chen, L.; Zhao, Z.; Jiang, N.; Khan, N.U.; Hu, Q.; Liu, R.; Liu, Z.; Qian, X.; Zhu, X.; et al. Exploring genomic regions and genes modulating plant height and flag leaf morphology in rice. Plant J. 2025, 122, e70145. [Google Scholar] [CrossRef]
- Shailani, A.; Joshi, R.; Singla-Pareek, S.L.; Pareek, A. Stacking for future: Pyramiding genes to improve drought and salinity tolerance in rice. Physiol. Plant. 2021, 172, 1352–1362. [Google Scholar] [CrossRef]
- Bakare, M.A.; Kayondo, S.I.; Aghogho, C.I.; Wolfe, M.D.; Parkes, E.Y.; Kulakow, P.; Egesi, C.; Rabbi, I.Y.; Jannink, J.L. Exploring genotype by environment interaction on cassava yield and yield related traits using classical statistical methods. PLoS ONE 2022, 17, e0268189. [Google Scholar] [CrossRef]
- Zeng, Y.; Shi, J.; Ji, Z.; Wen, Z.; Liang, Y.; Yang, C. Genotype by Environment Interaction: The Greatest Obstacle in Precise Determination of Rice Sheath Blight Resistance in the Field. Plant Dis. 2017, 101, 1795–1801. [Google Scholar] [CrossRef]
- Hao, N.; Han, D.G.; Huang, K.; Du, Y.L.; Yang, J.J.; Zhang, J.; Wen, C.L.; Wu, T. Genome-based breeding approaches in major vegetable crops. Theor. Appl. Genet. 2020, 133, 1739–1752. [Google Scholar] [CrossRef]
- Desta, Z.A.; Ortiz, R. Genomic selection: Genome-wide prediction in plant improvement. Trends Plant Sci. 2014, 19, 592–601. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.X.; Ji, Z.J.; Wen, Z.H.; Liang, Y.; Yang, C.D. Combination of Eight Alleles at Four Quantitative Trait Loci Determines Grain Length in Rice. PLoS ONE 2016, 11, e0150832. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G. Principle and Method of Silver Staining of Proteins Separated by Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis. Methods Mol. Biol. 2018, 1853, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef]
- Sullivan, S.G.; Greenland, S. Bayesian regression in SAS software. Int. J. Epidemiol. 2013, 42, 308–317. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Planting Time and Location | Marker | Chr. | F Value | p Value | Marker R2 | Allele Associated with Highest Plant Height (Marker Band Size in bp) |
|---|---|---|---|---|---|---|
| May 2012 in Hangzhou | D815 | 8 | 10.72 | 0.0013 | 0.059 | 130 |
| May 2012 in Hangzhou | D1051 | 10 | 10.58 | 0.0014 | 0.058 | 96 |
| May 2012 in Hangzhou | RM409 | 9 | 5.99 | 0.0030 | 0.064 | 95 |
| May 2012 in Hangzhou | D224C | 2 | 7.50 | 0.0068 | 0.040 | 88 |
| May 2012 in Hangzhou | D206C | 2 | 5.07 | 0.0072 | 0.054 | 175 |
| May 2012 in Hangzhou | D1126 | 11 | 7.28 | 0.0076 | 0.039 | 175 |
| May 2012 in Hangzhou | D818 | 8 | 4.75 | 0.0097 | 0.052 | 180 |
| November 2012 in Hainan | D142C | 1 | 6.36 | 0.0004 | 0.094 | 225 |
| November 2012 in Hainan | D448 | 4 | 11.80 | 0.0007 | 0.058 | 140 |
| November 2012 in Hainan | RM3589 | 7 | 4.91 | 0.0008 | 0.096 | 100 |
| November 2012 in Hainan | RM523 | 3 | 4.43 | 0.0019 | 0.091 | 152 |
| November 2012 in Hainan | D118A | 1 | 6.12 | 0.0026 | 0.060 | 150 |
| November 2012 in Hainan | D116C | 1 | 5.38 | 0.0053 | 0.053 | 165 |
| November 2012 in Hainan | D622 | 6 | 7.82 | 0.0057 | 0.038 | 135 |
| November 2012 in Hainan | D120A | 1 | 5.24 | 0.0060 | 0.051 | 178 |
| November 2012 in Hainan | D134B | 1 | 5.00 | 0.0076 | 0.049 | 115 |
| November 2012 in Hainan | D128A | 1 | 4.96 | 0.0079 | 0.049 | 100 |
| November 2012 in Hainan | D122E | 1 | 4.83 | 0.0089 | 0.047 | 112 |
| November 2012 in Hainan | D142A | 1 | 4.80 | 0.0092 | 0.047 | 203 |
| November 2012 in Hainan | D1051 | 10 | 6.86 | 0.0095 | 0.037 | 96 |
| May 2013 in Hangzhou | RM3589 | 7 | 4.95 | 0.0008 | 0.096 | 100 |
| May 2013 in Hangzhou | RM523 | 3 | 3.83 | 0.0050 | 0.076 | 152 |
| May 2013 in Hangzhou | RM409 | 9 | 5.37 | 0.0054 | 0.052 | 95 |
| June 2017 in Hangzhou | D1051 | 10 | 10.78 | 0.0012 | 0.044 | 96 |
| June 2017 in Hangzhou | RM3589 | 7 | 4.65 | 0.0012 | 0.072 | 100 |
| June 2017 in Hangzhou | RM409 | 9 | 6.85 | 0.0013 | 0.053 | 95 |
| June 2017 in Hangzhou | D1126 | 11 | 7.40 | 0.0070 | 0.028 | 175 |
| June 2017 in Hangzhou | RM6103 | 3 | 7.14 | 0.0080 | 0.027 | 188 |
| June 2018 in Hangzhou | RM409 | 9 | 11.10 | 0.0000 | 0.083 | 95 |
| June 2018 in Hangzhou | RM3589 | 7 | 5.32 | 0.0004 | 0.080 | 100 |
| June 2018 in Hangzhou | RM6103 | 3 | 9.88 | 0.0019 | 0.037 | 188 |
| June 2018 in Hangzhou | D130B | 1 | 5.76 | 0.0036 | 0.043 | 105 |
| June 2018 in Hangzhou | D304B | 3 | 4.43 | 0.0047 | 0.050 | 180 |
| June 2018 in Hangzhou | D815 | 8 | 7.82 | 0.0055 | 0.030 | 130 |
| June 2018 in Hangzhou | D206C | 2 | 4.79 | 0.0091 | 0.037 | 175 |
| Number of Loci Used in Building the Prediction Model | F Value (p < 0.0001) | Plant Height Prediction Model Developed Using the 273 Rice Genotype * | Predicted Average Plant Height of the 219 RILs (cm) | Average Plant Height of the 219 RILs Grown in Six Years (cm) | Average Absolute Error Between Predicted Plant Height and the Real Plant Height of the 219 RILs (cm) |
|---|---|---|---|---|---|
| 2 (RM409, RM6103) | 42.23 | y = 5.47816 x + 137.12733 | 123.85 | 115.17 | 14.02 |
| 3 (RM409, RM6103, D130B) | 30.09 | y = 2.60955 x + 129.0469 | 121.85 | 115.17 | 12.82 |
| 4 (RM409, RM6103, D130B, D224C) | 24.06 | y = 1.59275 x + 126.14526 | 120.89 | 115.17 | 12.14 |
| 5 (RM409, RM6103, D130B, D224C, D142C) | 39.89 | y = 1.92841 x + 127.87717 | 117.66 | 115.17 | 11.91 |
| 6 (RM409, RM6103, D130B, D224C, D142C, RM3589) | 47.99 | y = 1.97065 x + 131.53479 | 117.15 | 115.17 | 11.91 |
| 7 (RM409, RM6103, D130B, D224C, D142C, RM3589, D448) | 77.65 | y = 2.45987 x + 131.22406 | 113.63 | 115.17 | 13.11 |
| 8 (RM409, RM6103, D130B, D224C, D142C, RM3589, D448, D1051) | 57.79 | y = 1.74358 x + 128.60196 | 115.07 | 115.17 | 11.91 |
| 9 (RM409, RM6103, D130B, D224C, D142C, RM3589, D448, D1051, D1126) | 31.42 | y = 1.14012 x + 126.32044 | 116.50 | 115.17 | 11.05 |
| 10 (RM409, RM6103, D130B, D224C, D142C, RM3589, D448, D1051,D1126, D815) | 52.69 | y = 1.54178 x + 126.80416 | 116.60 | 115.17 | 11.96 |
| DF | Type 1 SS | Mean Square | F Value | p | % of Genotype + Environment + Genotype × Environment | |
|---|---|---|---|---|---|---|
| Genotype | 218 | 531,630.10 | 2438.67 | 227.09 | <0.0001 | 76.44 |
| Environment | 4 | 25,125.86 | 6281.47 | 584.94 | <0.0001 | 3.61 |
| Genotype × environment | 872 | 138,735.51 | 159.10 | 14.82 | <0.0001 | 19.95 |
| Planted in May 2013 | Planted on 29 May 2017 | Planted on 7 June 2017 | Planted on 30 June 2017 | Planted on 23 May 2018 | Planted on 2 June 2018 | |
|---|---|---|---|---|---|---|
| Planted in May 2013 | - | 12.56 | 12.08 | 11.53 | 11.82 | 12.79 |
| Planted on 29 May 2017 | - | 11.11 | 9.25 | 9.79 | 10.67 | |
| Planted on 7 June 2017 | - | 8.33 | 8.37 | 7.25 | ||
| Planted on 30 June 2017 | - | 6.96 | 7.95 | |||
| Planted on 23 May 2018 | - | 5.72 | ||||
| Planted on 2 June 2018 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Y.; Xie, Z.; Chen, D.; Chen, H.; Zeng, Y.; Dai, S. Prediction of Rice Plant Height Using Linear Regression Model by Pyramiding Plant Height-Related Alleles. Int. J. Mol. Sci. 2025, 26, 6249. https://doi.org/10.3390/ijms26136249
Huang Y, Xie Z, Chen D, Chen H, Zeng Y, Dai S. Prediction of Rice Plant Height Using Linear Regression Model by Pyramiding Plant Height-Related Alleles. International Journal of Molecular Sciences. 2025; 26(13):6249. https://doi.org/10.3390/ijms26136249
Chicago/Turabian StyleHuang, Yongxiang, Zhihao Xie, Daming Chen, Haomin Chen, Yuxiang Zeng, and Shuangfeng Dai. 2025. "Prediction of Rice Plant Height Using Linear Regression Model by Pyramiding Plant Height-Related Alleles" International Journal of Molecular Sciences 26, no. 13: 6249. https://doi.org/10.3390/ijms26136249
APA StyleHuang, Y., Xie, Z., Chen, D., Chen, H., Zeng, Y., & Dai, S. (2025). Prediction of Rice Plant Height Using Linear Regression Model by Pyramiding Plant Height-Related Alleles. International Journal of Molecular Sciences, 26(13), 6249. https://doi.org/10.3390/ijms26136249