
Metabolomic Analysis Identifies Betaine as a Key Mediator of TAp73α-Induced Ferroptosis in Ovarian Granulosa Cells

Abstract
1. Introduction
2. Results
2.1. TAp73α Overexpression Promotes Ferroptosis in Granulosa Cells
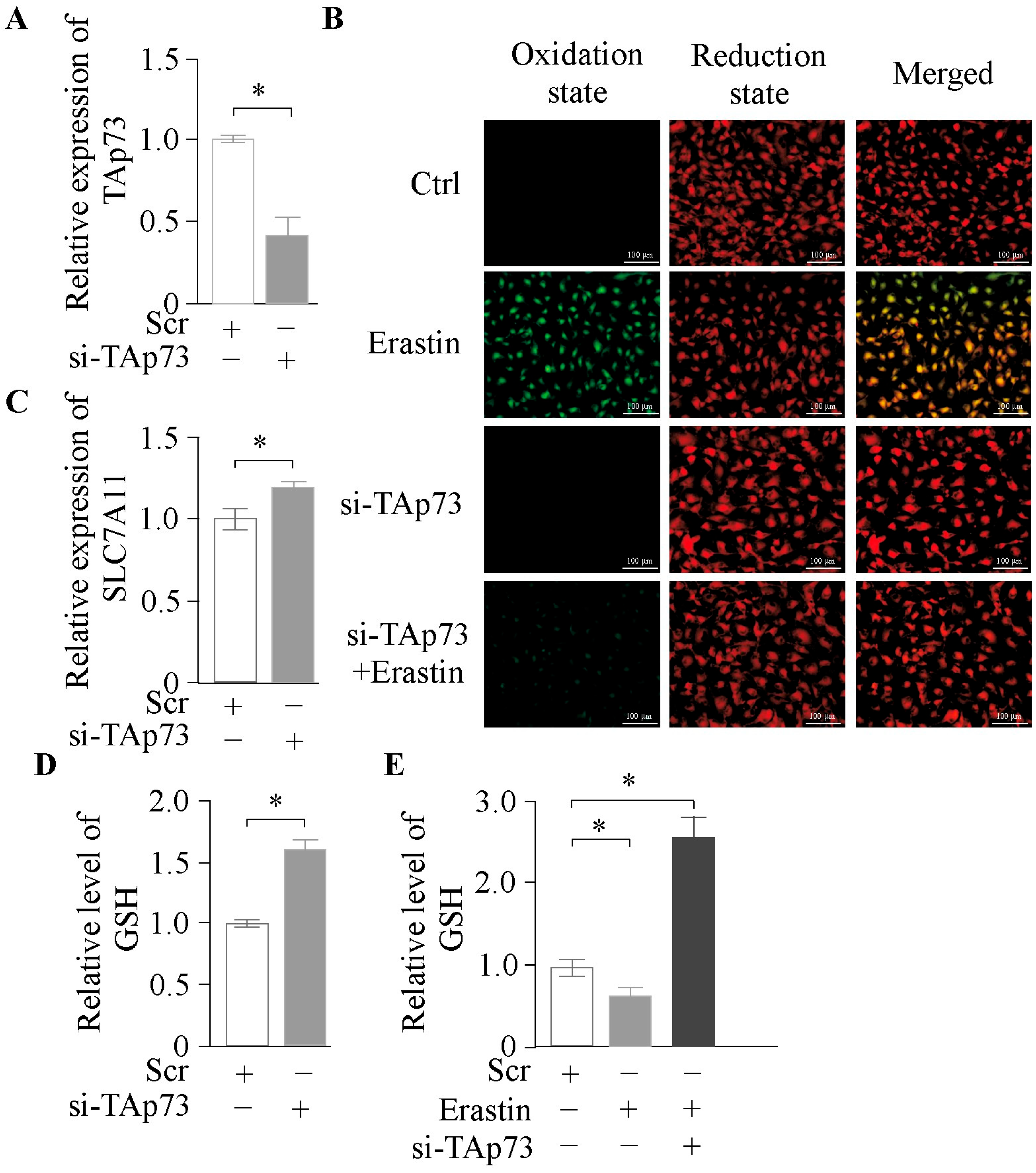
2.2. TAp73α Knockdown Attenuates Ferroptosis in Granulosa Cells
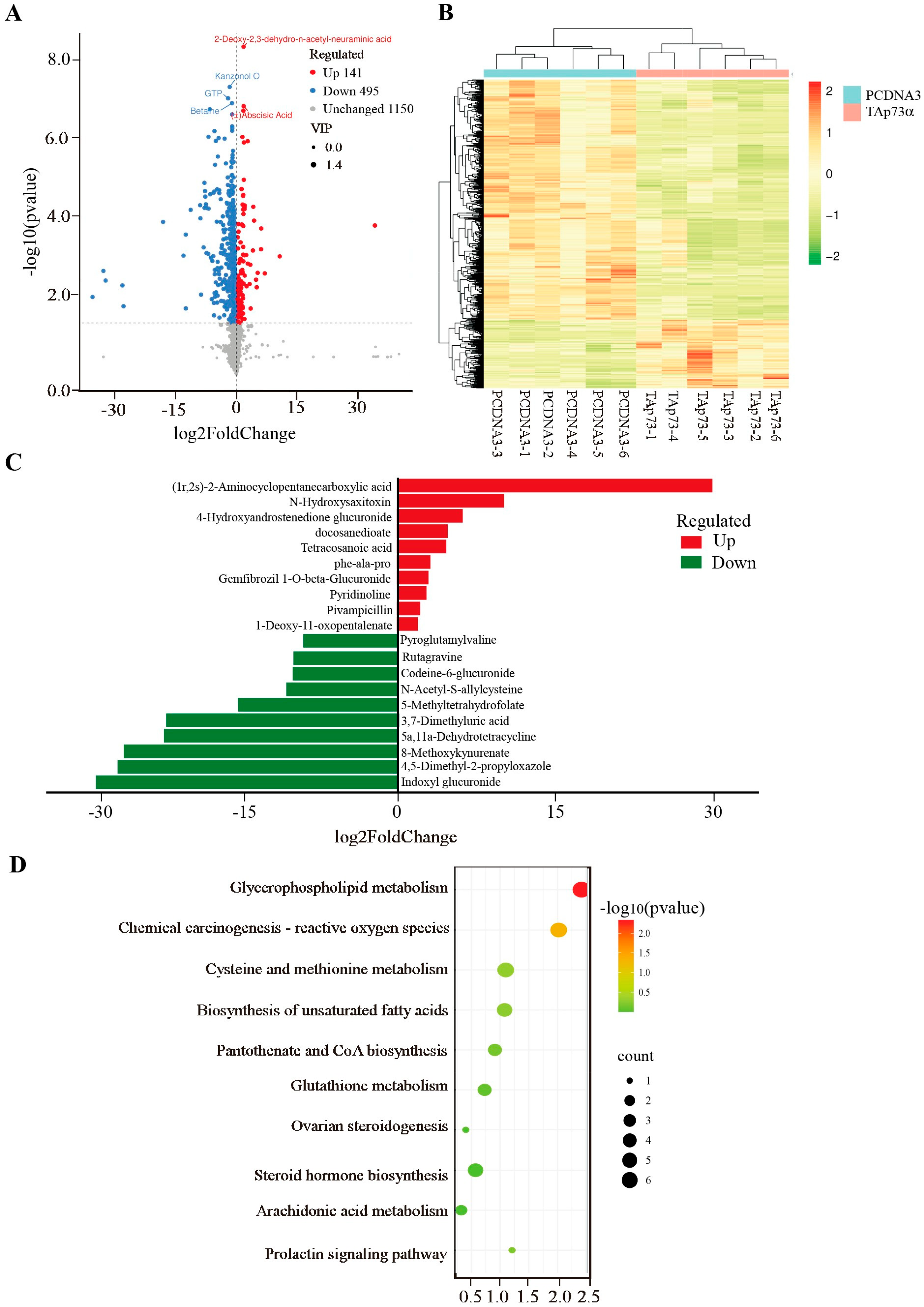
2.3. Effect of TAp73α Overexpression on the Metabolomics of GCs in Positive Mode
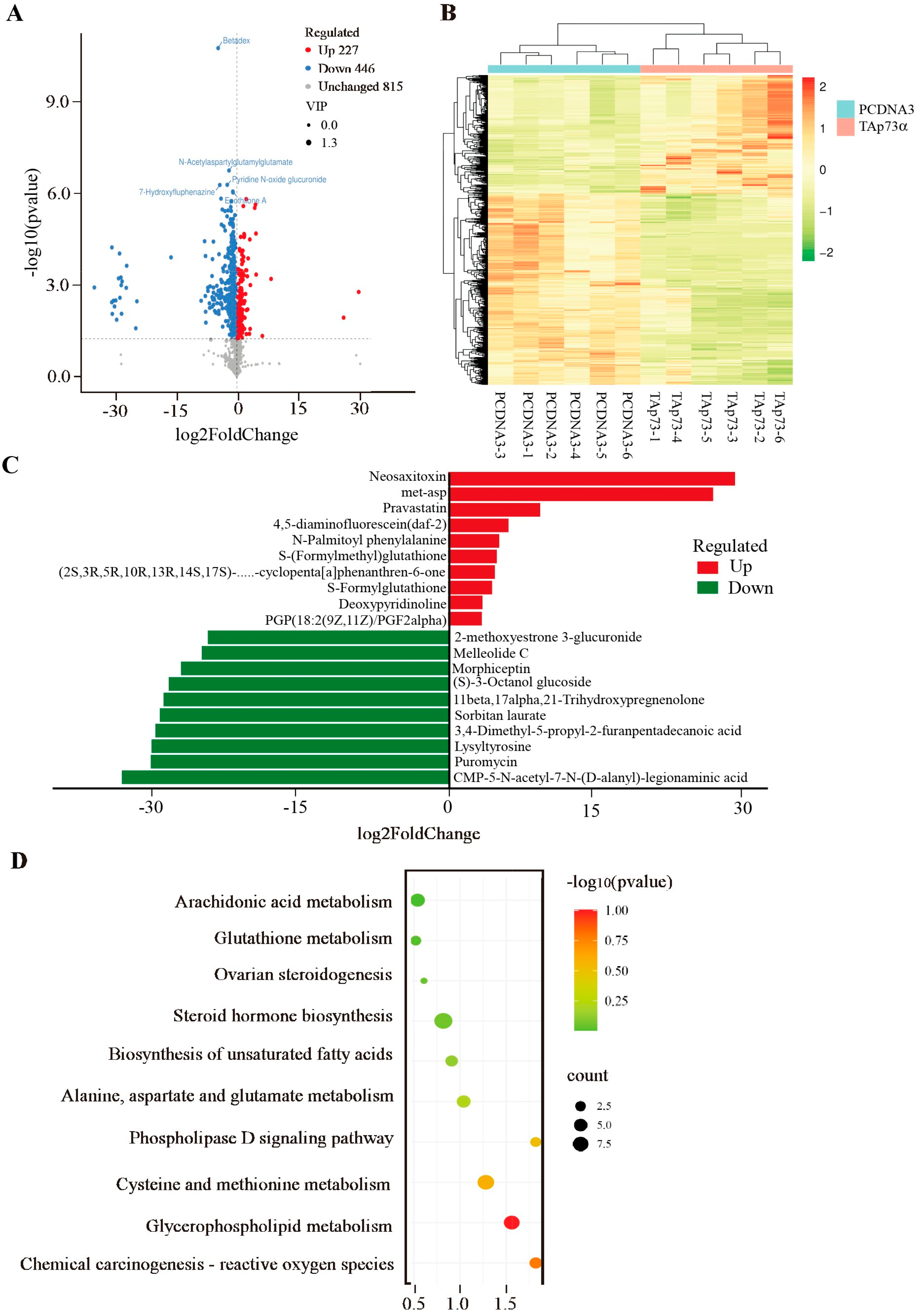
2.4. Effect of TAp73α Overexpression on the Metabolomics of GCs in Negative Mode
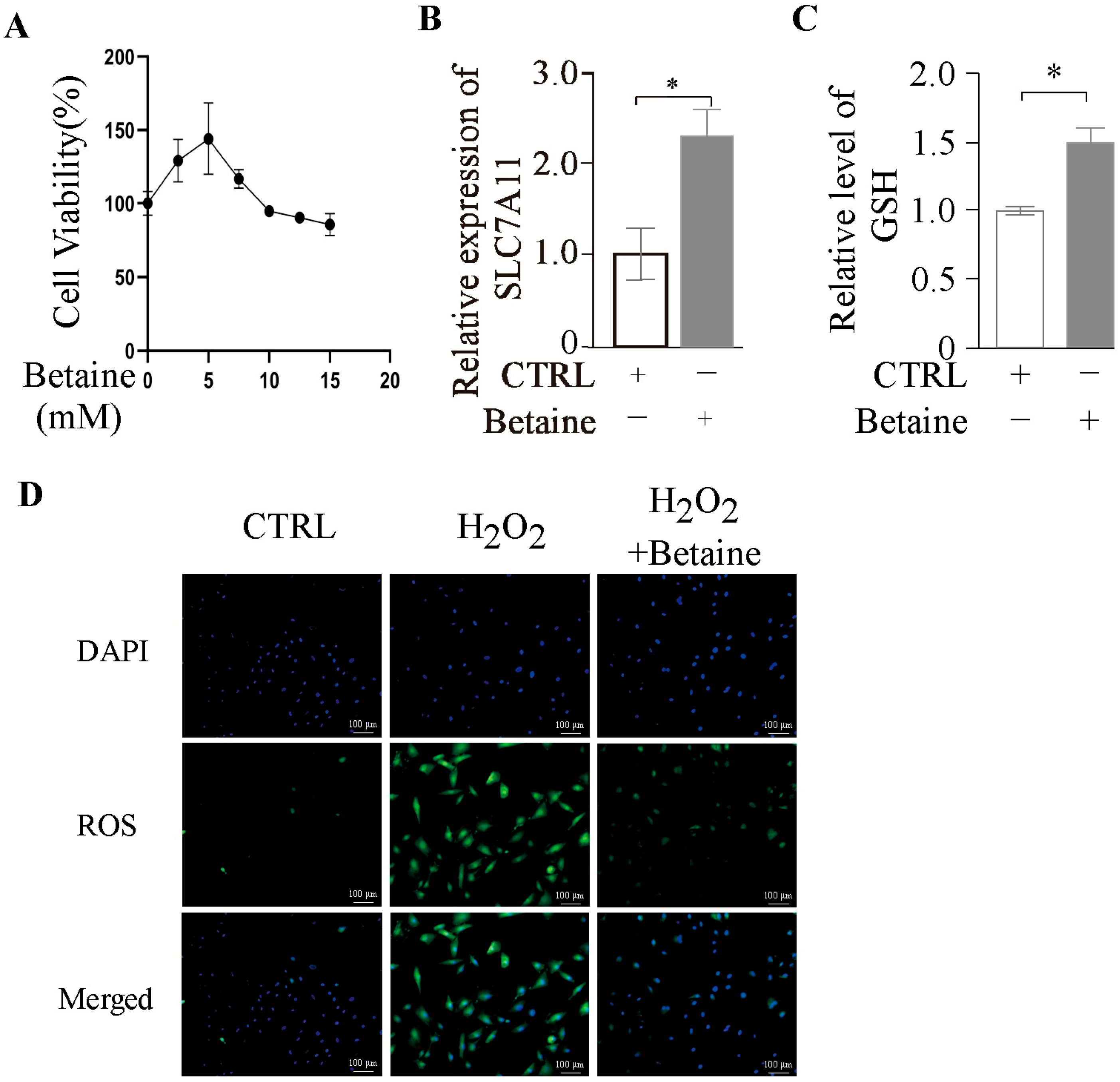
2.5. Betaine Enhances Antioxidant Capacity of GCs
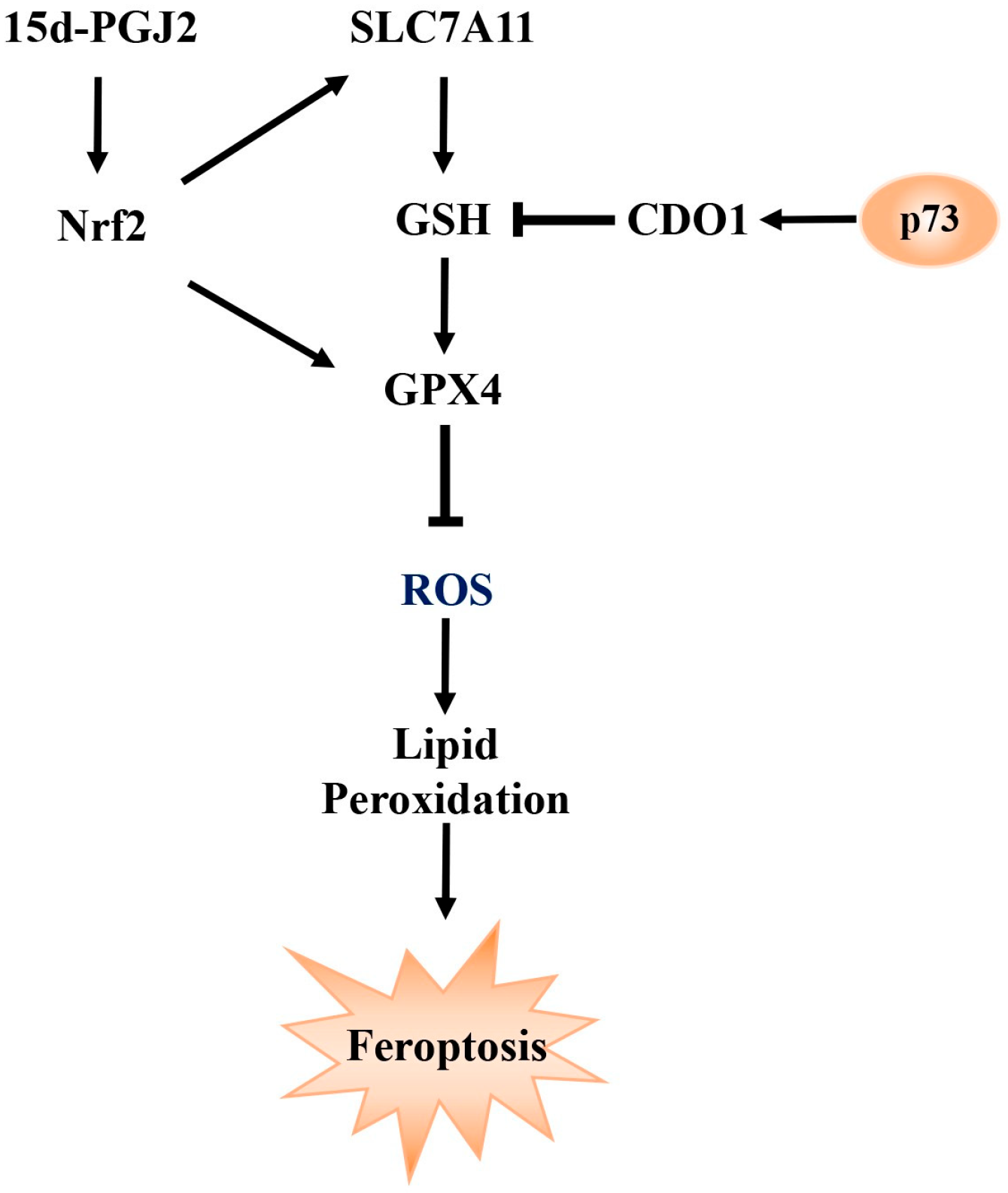
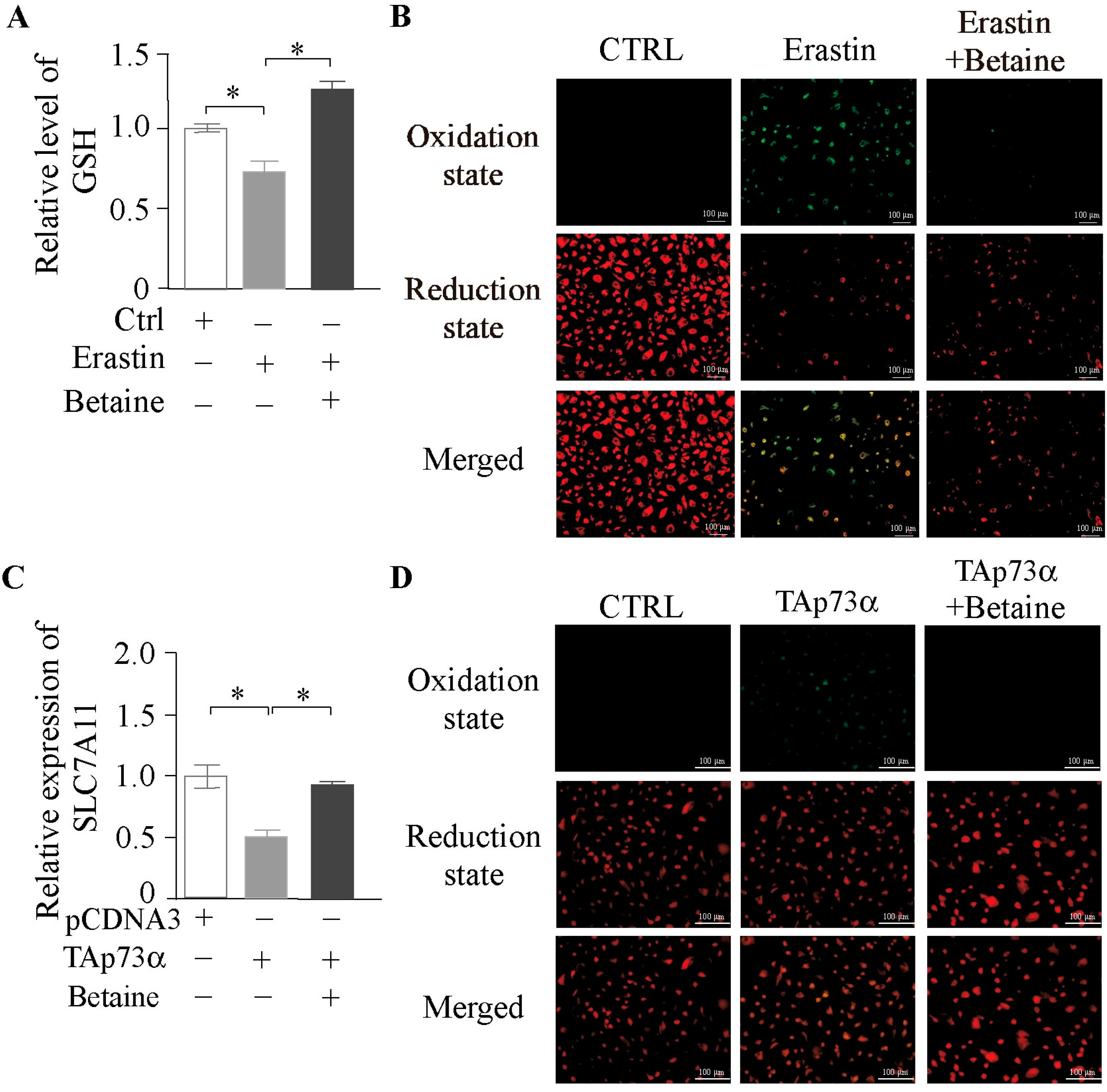
2.6. Betaine as a Key Metabolite in the Modulation of Ferroptosis by TAp73α
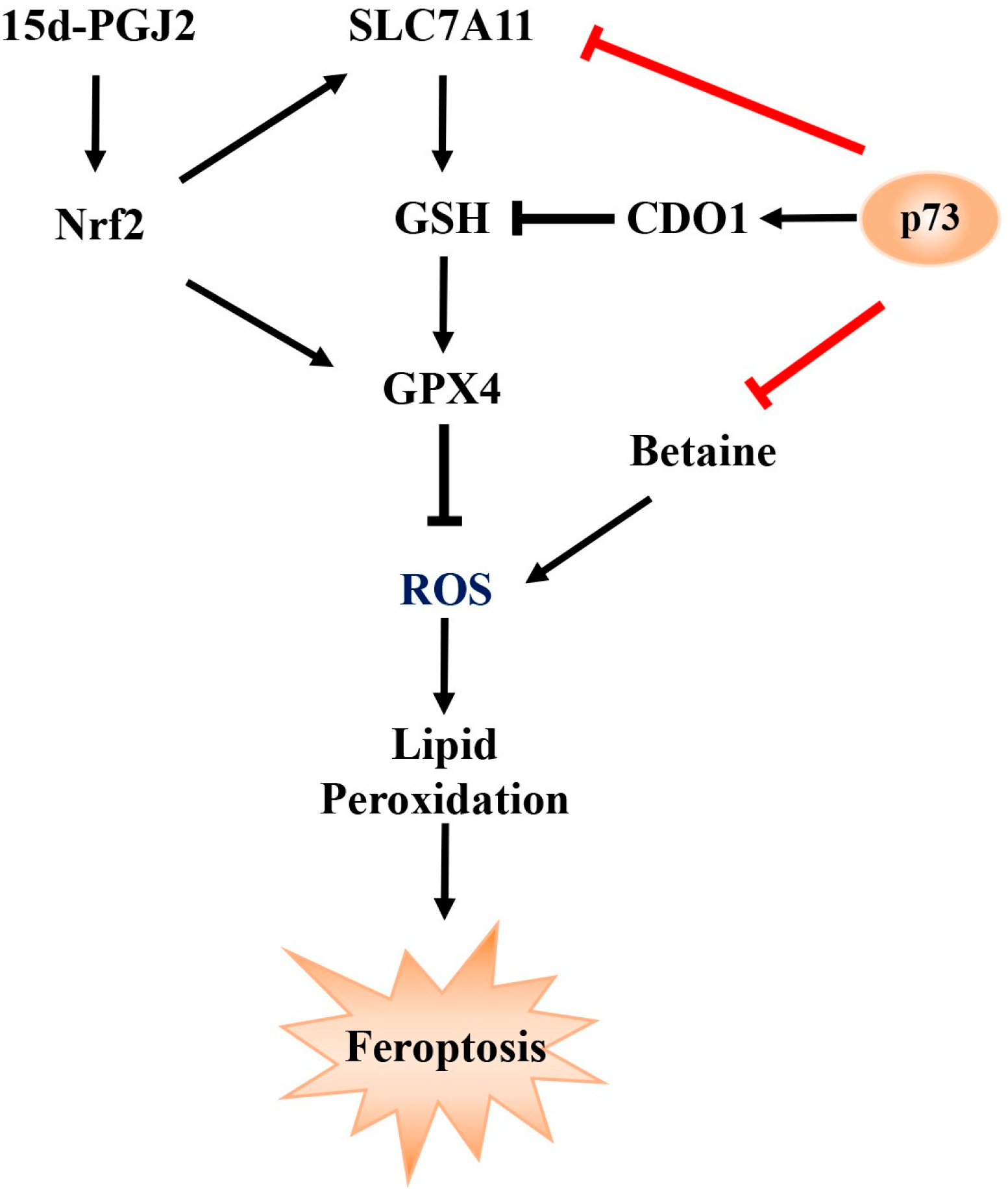
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Cell Transfection
4.3. Metabolite Extraction
4.4. LC-MS/MS Analysis
4.5. Data Preprocessing and Annotation
4.6. GSH Determination
4.7. Detection of Lipid Peroxidation
4.8. ROS Determination
4.9. CCK-8 Assay
4.10. Real-Time Quantitative PCR Analysis
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Betaine | Trimethylglycine |
| CCK-8 | Cell counting kit-8 |
| FSH | Follicle-stimulating hormone |
| GC | Granulosa cell |
| GPX | Glutathione peroxidase |
| G6PD | Glucose-6-phosphate dehydrogenase |
| GSH | Glutathione |
| LH | Luteinizing hormone |
| MDA | Malondialdehyde |
| PGJ2 | Prostaglandin J2 |
| 15d-PGJ2 | 15deoxy-Δ12,14-prostaglandin J2 |
| PPP | Pentose phosphate pathway |
| ROS | Reactive oxygen species |
| SLC7A11 | Solute carrier family 7 member 11 |
| TAp73α | Transactivation domain-containing isoform of p73α |
References
- Wang, C.; Sun, H.; Davis, J.S.; Wang, X.; Huo, L.; Sun, N.; Huang, Q.; Lv, X.; Wang, C.; He, C.; et al. FHL2 deficiency impairs follicular development and fertility by attenuating EGF/EGFR/YAP signaling in ovarian granulosa cells. Cell Death Dis. 2023, 14, 239. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Meng, X.; Liu, S.; Li, W.; Zhang, X.; Zhou, J.; Yao, W.; Dong, C.; Liu, Z.; Zhou, J.; et al. Oocytes and hypoxanthine orchestrate the G2-M switch mechanism in ovarian granulosa cells. Development 2020, 13, 147. [Google Scholar] [CrossRef] [PubMed]
- Osterburg, C.; Dotsch, V. Structural diversity of p63 and p73 isoforms. Cell Death Differ. 2022, 29, 921–937. [Google Scholar] [CrossRef] [PubMed]
- Tomasini, R.; Tsuchihara, K.; Wilhelm, M.; Fujitani, M.; Rufini, A.; Cheung, C.C.; Khan, F.; Itie-Youten, A.; Wakeham, A.; Tsao, M.S.; et al. TAp73 knockout shows genomic instability with infertility and tumor suppressor functions. Genes Dev. 2008, 22, 2677–2691. [Google Scholar] [CrossRef]
- Santos Guasch, G.L.; Beeler, J.S.; Marshall, C.B.; Shaver, T.M.; Sheng, Q.; Johnson, K.N.; Boyd, K.L.; Venters, B.J.; Cook, R.S.; Pietenpol, J.A. p73 Is Required for Ovarian Follicle Development and Regulates a Gene Network Involved in Cell-to-Cell Adhesion. iScience 2018, 8, 236–249. [Google Scholar] [CrossRef]
- Talos, F.; Abraham, A.; Vaseva, A.V.; Holembowski, L.; Tsirka, S.E.; Scheel, A.; Bode, D.; Dobbelstein, M.; Brück, W.; Moll, U.M. p73 is an essential regulator of neural stem cell maintenance in embryonal and adult CNS neurogenesis. Cell Death Differ. 2010, 17, 1816–1829. [Google Scholar] [CrossRef]
- Abreu, F.M.; Coutinho da Silva, M.A.; Cruppe, L.H.; Mussard, M.L.; Bridges, G.A.; Harstine, B.R.; Smith, G.W.; Geary, T.W.; Day, M.L. Role of progesterone concentrations during early follicular development in beef cattle: I. Characteristics of LH secretion and oocyte quality. Anim. Reprod. Sci. 2018, 196, 59–68. [Google Scholar] [CrossRef]
- Zhou, J.; Peng, X.; Mei, S. Autophagy in Ovarian Follicular Development and Atresia. Int. J. Biol. Sci. 2019, 15, 726–737. [Google Scholar] [CrossRef]
- Tilly, J.L.; Kowalski, K.I.; Johnson, A.L.; Hsueh, A.J. Involvement of apoptosis in ovarian follicular atresia and postovulatory regression. Endocrinology 1991, 129, 2799–2801. [Google Scholar] [CrossRef]
- Kaur, S.; Kurokawa, M. Regulation of Oocyte Apoptosis: A View from Gene Knockout Mice. Int. J. Mol. Sci. 2023, 24, 1345. [Google Scholar] [CrossRef]
- Pan, Y.; Gan, M.; Wu, S.; He, Y.; Feng, J.; Jing, Y.; Li, J.; Chen, Q.; Tong, J.; Kang, L.; et al. tRF-Gly-GCC in Atretic Follicles Promotes Ferroptosis in Granulosa Cells by Down-Regulating MAPK1. Int. J. Mol. Sci. 2024, 25, 9061. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ni, F.; Huang, J.; Hu, Y.; Wang, J.; Wang, X.; Du, X.; Jiang, H. PPAR-α inhibits DHEA-induced ferroptosis in granulosa cells through upregulation of FADS2. Biochem. Biophys. Res. Commun. 2024, 715, 150005. [Google Scholar] [CrossRef] [PubMed]
- Abudureyimu, G.; Wu, Y.; Chen, Y.; Wang, L.; Hao, G.; Yu, J.; Wang, J.; Lin, J.; Huang, J. MiR-134-3p targets HMOX1 to inhibit ferroptosis in granulosa cells of sheep follicles. J. Ovarian Res. 2024, 17, 3. [Google Scholar] [CrossRef] [PubMed]
- Agostini, M.; Niklison-Chirou, M.V.; Annicchiarico-Petruzzelli, M.M.; Grelli, S.; Di Daniele, N.; Pestlikis, I.; Knight, R.A.; Melino, G.; Rufini, A. p73 Regulates Primary Cortical Neuron Metabolism: A Global Metabolic Profile. Mol. Neurobiol. 2018, 55, 3237–3250. [Google Scholar] [CrossRef]
- D’Alessandro, A.; Marrocco, C.; Rinalducci, S.; Peschiaroli, A.; Timperio, A.M.; Bongiorno-Borbone, L.; Finazzi Agrò, A.; Melino, G.; Zolla, L. Analysis of TAp73-dependent signaling via omics technologies. J. Proteome Res. 2013, 12, 4207–4220. [Google Scholar] [CrossRef]
- Zhang, J.; Sun, W.; Yan, W.; Kong, X.; Shen, T.; Laubach, K.; Chen, M.; Chen, X. TP73 Isoform-specific disruption reveals a critical role of TAp73beta in growth suppression and inflammatory response. Cell Death Dis. 2023, 14, 14. [Google Scholar] [CrossRef]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An iron-dependent form of nonapoptotic cell death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef]
- Chen, L.-L.; Shan, W.; Cai, D.-L.; Chen, J.-Y.; Lu, W.-J.; Su, X.-G.; Kuang, J.-F. Postharvest application of glycine betaine ameliorates chilling injury in cold-stored banana fruit by enhancing antioxidant system. Sci. Hortic. 2021, 287, 110264. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, Q.; Bai, F.; Hu, Y.; Xu, B.; Tang, Y.; Wu, J. Granulosa cell-specific FOXJ2 overexpression induces premature ovarian insufficiency by triggering apoptosis via mitochondrial calcium overload. J. Ovarian Res. 2025, 18, 75. [Google Scholar] [CrossRef]
- Cuiling, L.; Wei, Y.; Zhaoyuan, H.; Yixun, L. Granulosa cell proliferation differentiation and its role in follicular development. Chin. Sci. Bull. 2005, 50, 2665–2671. [Google Scholar] [CrossRef]
- Holembowski, L.; Kramer, D.; Riedel, D.; Sordella, R.; Nemajerova, A.; Dobbelstein, M.; Moll, U.M. TAp73 is essential for germ cell adhesion and maturation in testis. J. Cell Biol. 2014, 204, 1173–1190. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Chen, C.; Gao, Y.; Cui, X.; Zhang, Y.; Gu, L.; He, Y.; Li, J.; Gao, S.; Gao, R.; et al. Transcriptome Dynamics and Cell Dialogs Between Oocytes and Granulosa Cells in Mouse Follicle Development. Genom. Proteom. Bioinform. 2024, 22, qzad001. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, L.; Li, W.; Chen, T.; Bin, Z.; Zhao, L.; Wang, H.; Wang, X.; Xu, L.; Liu, X.; et al. TAp73-induced phosphofructokinase-1 transcription promotes the Warburg effect and enhances cell proliferation. Nat. Commun. 2018, 9, 4683. [Google Scholar] [CrossRef]
- Sang, M.; Ando, K.; Okoshi, R.; Koida, N.; Li, Y.; Zhu, Y.; Shimozato, O.; Geng, C.; Shan, B.; Nakagawara, A.; et al. Plk3 inhibits pro-apoptotic activity of p73 through physical interaction and phosphorylation. Genes Cells Devoted Mol. Cell. Mech. 2009, 14, 775–788. [Google Scholar] [CrossRef]
- Amelio, I.; Markert, E.K.; Rufini, A.; Antonov, A.V.; Sayan, B.S.; Tucci, P.; Agostini, M.; Mineo, T.C.; Levine, A.J.; Melino, G. p73 regulates serine biosynthesis in cancer. Oncogene 2014, 33, 5039–5046. [Google Scholar] [CrossRef]
- Li, X.Y.; Luo, B.L.; Wang, L.J.; Zhang, W.D.; Liu, Z.G. 15-Deoxy-prostaglandin J2 anti-inflammation in a rat model of chronic obstructive pulmonary disease and human bronchial epithelial cells via Nrf2 activation. Genet. Mol. Res. GMR 2015, 14, 14037–14042. [Google Scholar] [CrossRef]
- Dodson, M.; Portuguez, R.C.; Zhang, D.D. NRF2 plays a critical role in mitigating lipid peroxidation and ferroptosis. Redox Biol. 2019, 23, 101107. [Google Scholar] [CrossRef]
- Xue, X.; Wang, M.; Cui, J.; Yang, M.; Ma, L.; Kang, R.; Tang, D.; Wang, J. Glutathione metabolism in ferroptosis and cancer therapy. Cancer Lett. 2025, 621, 217697. [Google Scholar] [CrossRef]
- Stockwell, B.R.; Friedmann Angeli, J.P.; Bayir, H.; Bush, A.I.; Conrad, M.; Dixon, S.J.; Fulda, S.; Gascón, S.; Hatzios, S.K.; Kagan, V.E.; et al. Ferroptosis: A Regulated Cell Death Nexus Linking Metabolism, Redox Biology, and Disease. Cell 2017, 171, 273–285. [Google Scholar] [CrossRef]
- Mohammadi, N.; Hemmati, M.; Motlagh, B.; Biyabani, A. Betaine postpones hyperglycemia-related senescence in ovarian and testicular cells: Involvement of RAGE and β-galactosidase. Cell Biochem. Funct. 2024, 42, e3973. [Google Scholar] [CrossRef]
- Mendoza, S.M.; Boyd, R.D.; Remus, J.; Wilcock, P.; Martinez, G.E.; van Heugten, E. Sow performance in response to natural betaine fed during lactation and post-weaning during summer and non-summer months. J. Anim. Sci. Biotechnol. 2020, 11, 69. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.; Jing, M.; Zhang, A.; Yi, J.; Zhang, Y. Effects of Dietary Betaine on the Laying Performance, Antioxidant Capacity, and Uterus and Ovary Function of Laying Hens at the Late Stage of Production. Anim. 2023, 13, 3283. [Google Scholar] [CrossRef] [PubMed]
- Omer, N.A.; Hu, Y.; Hu, Y.; Idriss, A.A.; Abobaker, H.; Hou, Z.; Dong, H.; Zhao, R. Dietary betaine activates hepatic VTGII expression in laying hens associated with hypomethylation of GR gene promoter and enhanced GR expression. J. Anim. Sci. Biotechnol. 2018, 9, 2. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wang, Y.; Li, L.; Han, Z.; Mao, S.; Wang, G. Betaine protects against heat exposure–induced oxidative stress and apoptosis in bovine mammary epithelial cells via regulation of ROS production. Cell Stress Chaperones 2019, 24, 453–460. [Google Scholar] [CrossRef]
- Chen, R.; Yang, M.; Song, Y.D.; Wang, R.X.; Wen, C.; Liu, Q.; Zhou, Y.M.; Zhuang, S. Effect of anhydrous betaine and hydrochloride betaine on growth performance, meat quality, postmortem glycolysis, and antioxidant capacity of broilers. Poult. Sci. 2022, 101, 101687. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Kawashima, M.; Ishiguro-Watanabe, M. KEGG for taxonomy-based analysis of pathways and genomes. Nucleic Acids Res. 2023, 51, D587–D592. [Google Scholar] [CrossRef]
- Wishart, D.S.; Guo, A.; Oler, E.; Wang, F.; Anjum, A.; Peters, H.; Dizon, R.; Sayeeda, Z.; Tian, S.; Lee, B.L.; et al. HMDB 5.0: The Human Metabolome Database for 2022. Nucleic Acids Res. 2022, 50, D622–D631. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Primer Sequence (5′-3′) |
|---|---|
| ACTB | GGTGCCCATCTATGAGGGGTACG |
| TTCTCCTTGATGTCACGGACGATTTC | |
| GAPDH | CACTTTGGCATCGTGGAGGGACTT |
| AACAGACACGTTGGGAGTGGGGAC | |
| p73 | TGCCTGCTAACGGTGAGATGAACG |
| GTCCCTGAGAGGTGAAGTACTCGATGC | |
| SLC7A11 | GTCCTGTCGCTGTTTGGAGCCTTGT |
| CTGACACTCATGCTATTTAGGACCATCACC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mei, L.; Chen, L.; Zhang, B.; Jia, X.; Gan, X.; Sun, W. Metabolomic Analysis Identifies Betaine as a Key Mediator of TAp73α-Induced Ferroptosis in Ovarian Granulosa Cells. Int. J. Mol. Sci. 2025, 26, 6045. https://doi.org/10.3390/ijms26136045
Mei L, Chen L, Zhang B, Jia X, Gan X, Sun W. Metabolomic Analysis Identifies Betaine as a Key Mediator of TAp73α-Induced Ferroptosis in Ovarian Granulosa Cells. International Journal of Molecular Sciences. 2025; 26(13):6045. https://doi.org/10.3390/ijms26136045
Chicago/Turabian StyleMei, Liping, Le Chen, Bingfei Zhang, Xianbo Jia, Xiang Gan, and Wenqiang Sun. 2025. "Metabolomic Analysis Identifies Betaine as a Key Mediator of TAp73α-Induced Ferroptosis in Ovarian Granulosa Cells" International Journal of Molecular Sciences 26, no. 13: 6045. https://doi.org/10.3390/ijms26136045
APA StyleMei, L., Chen, L., Zhang, B., Jia, X., Gan, X., & Sun, W. (2025). Metabolomic Analysis Identifies Betaine as a Key Mediator of TAp73α-Induced Ferroptosis in Ovarian Granulosa Cells. International Journal of Molecular Sciences, 26(13), 6045. https://doi.org/10.3390/ijms26136045