
A Novel Hybrid Peptide VLP-Aβ Efficiently Regulates Immunity by Stimulating Myeloid Differentiation Protein and Activating the NF-κB Pathway
 ,
,
Abstract
1. Introduction
2. Results
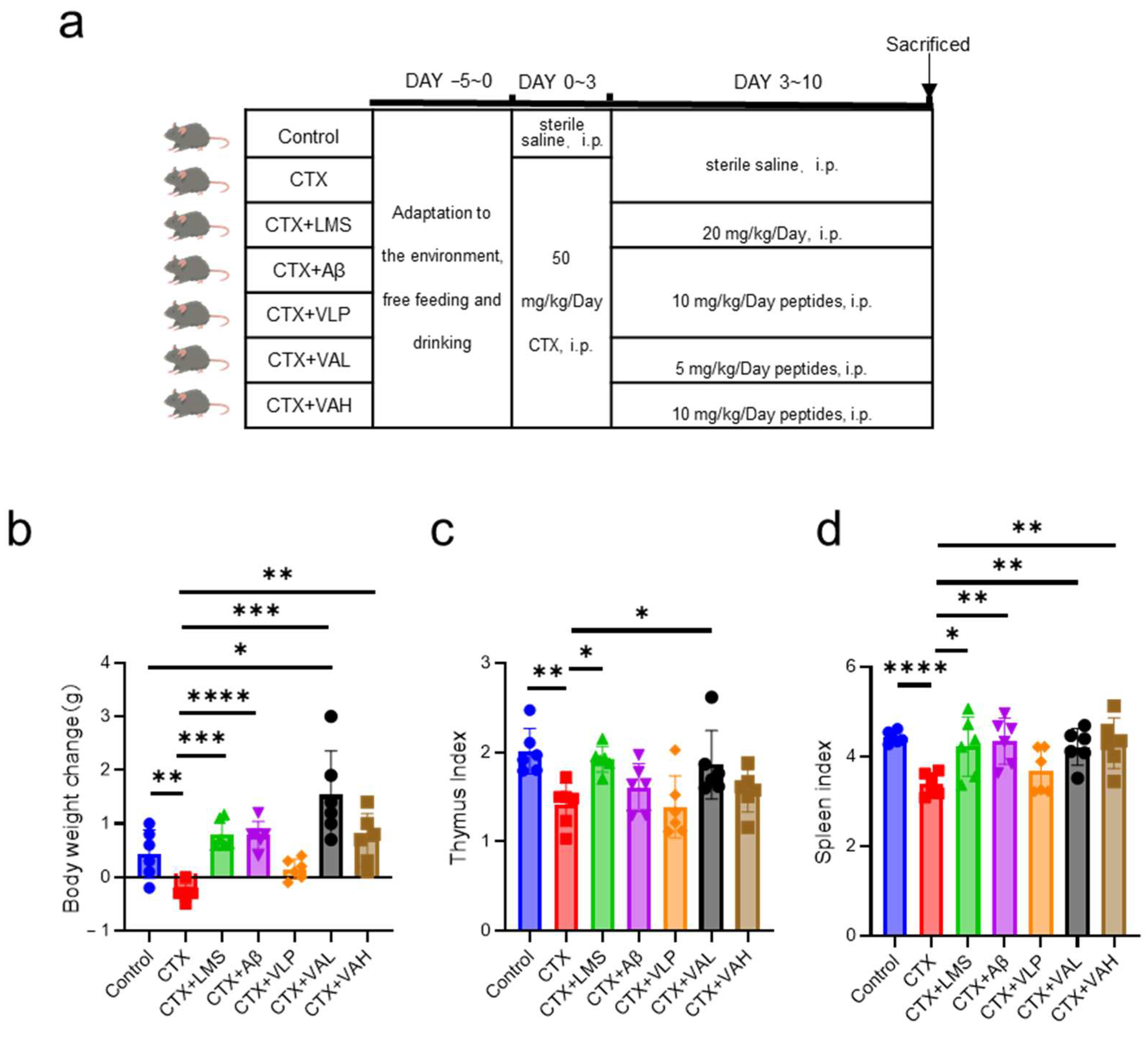
2.1. Hybrid Peptide VA Alleviates Cyclophosphamide-Induced Immunodeficiency in Mice
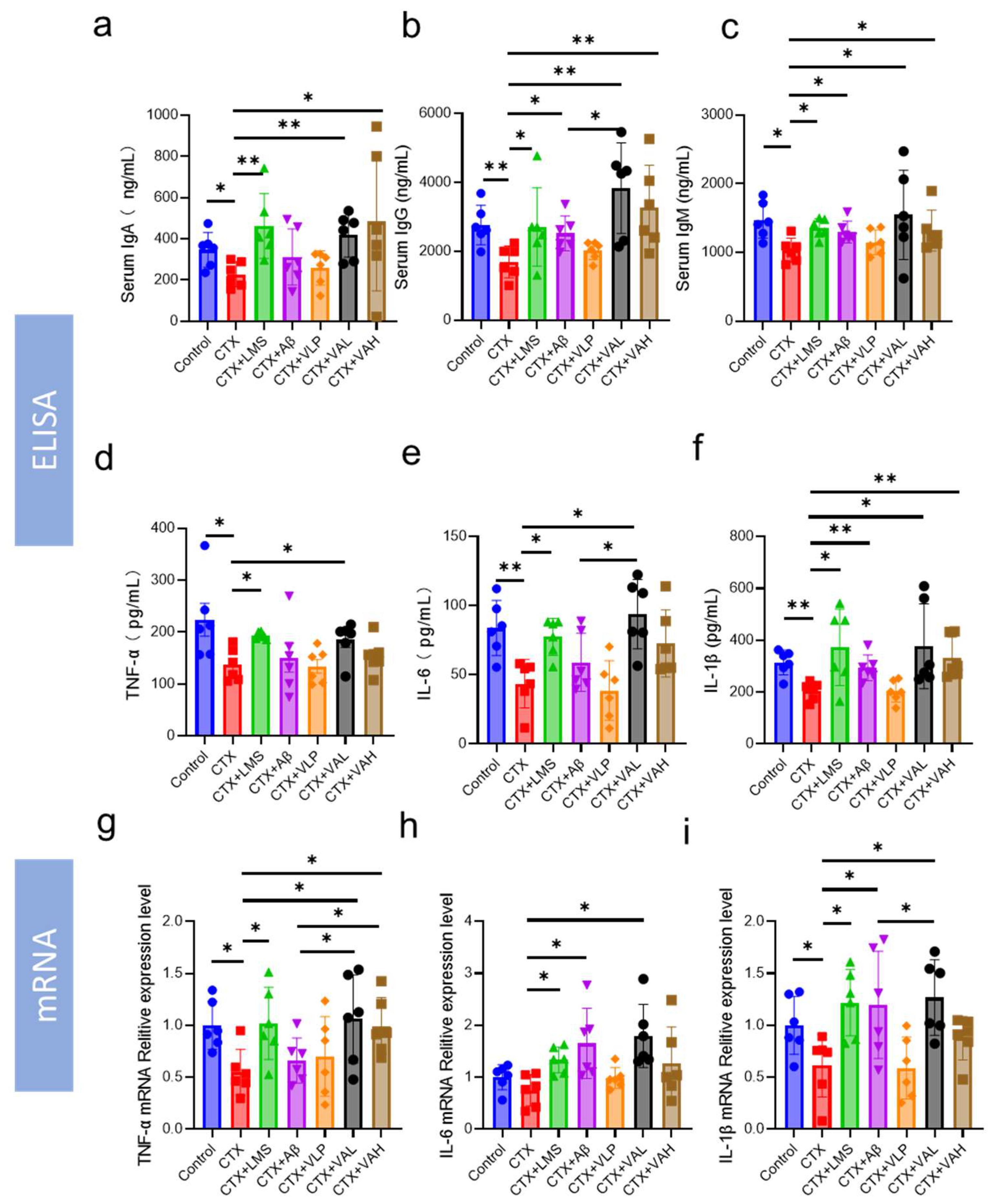
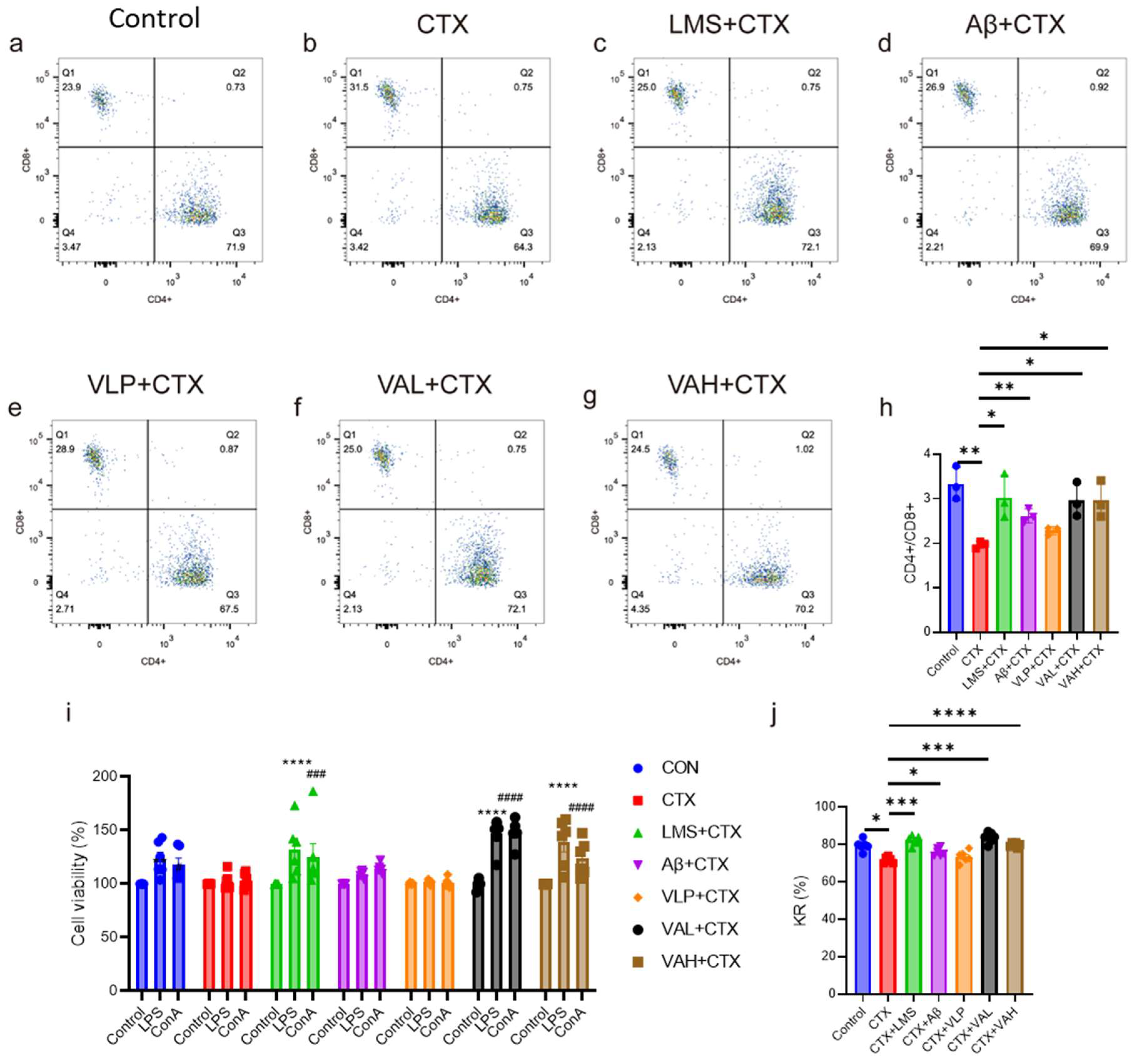
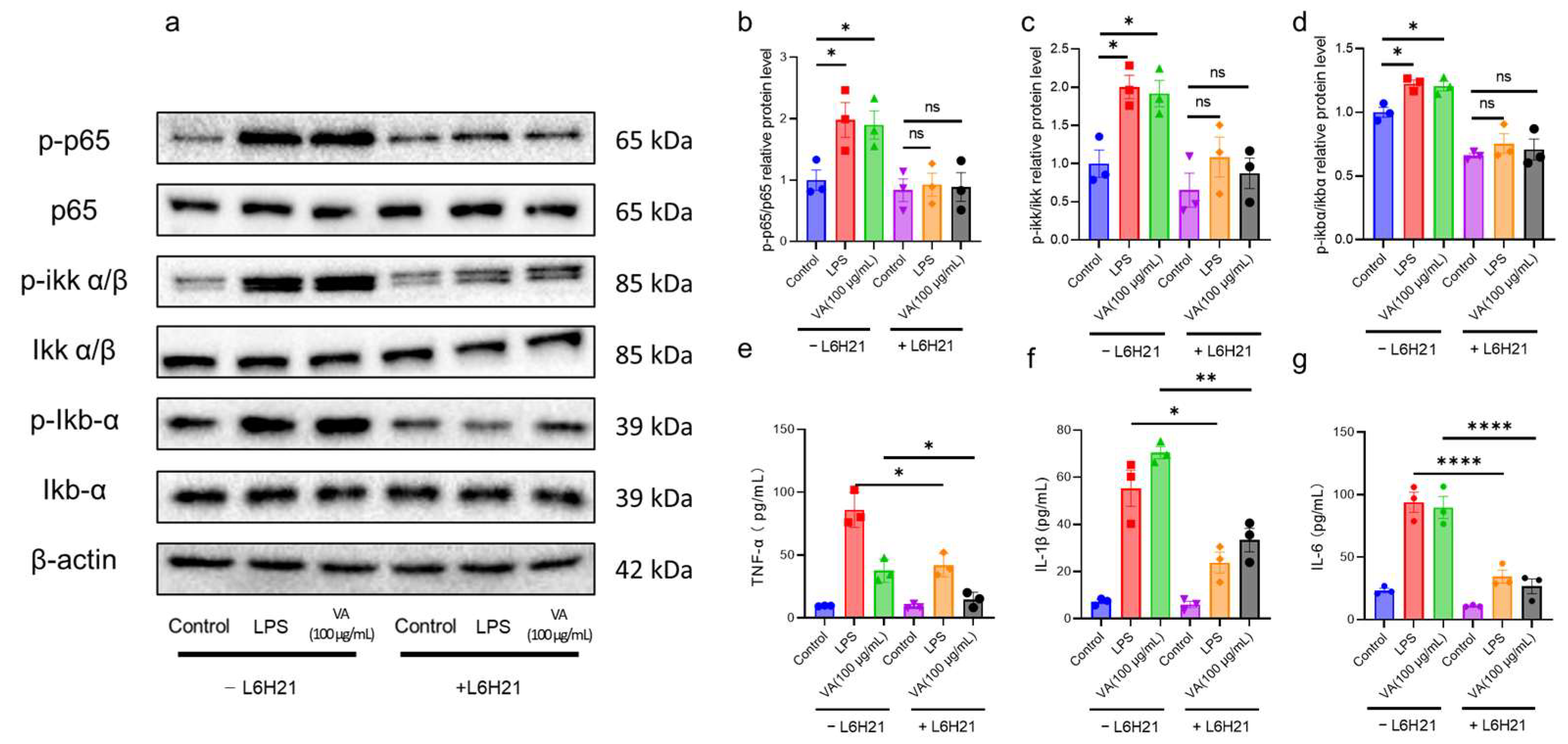
2.2. Hybrid Peptide VA Enhances Immunity by Activating the NF-κB Signaling Pathway
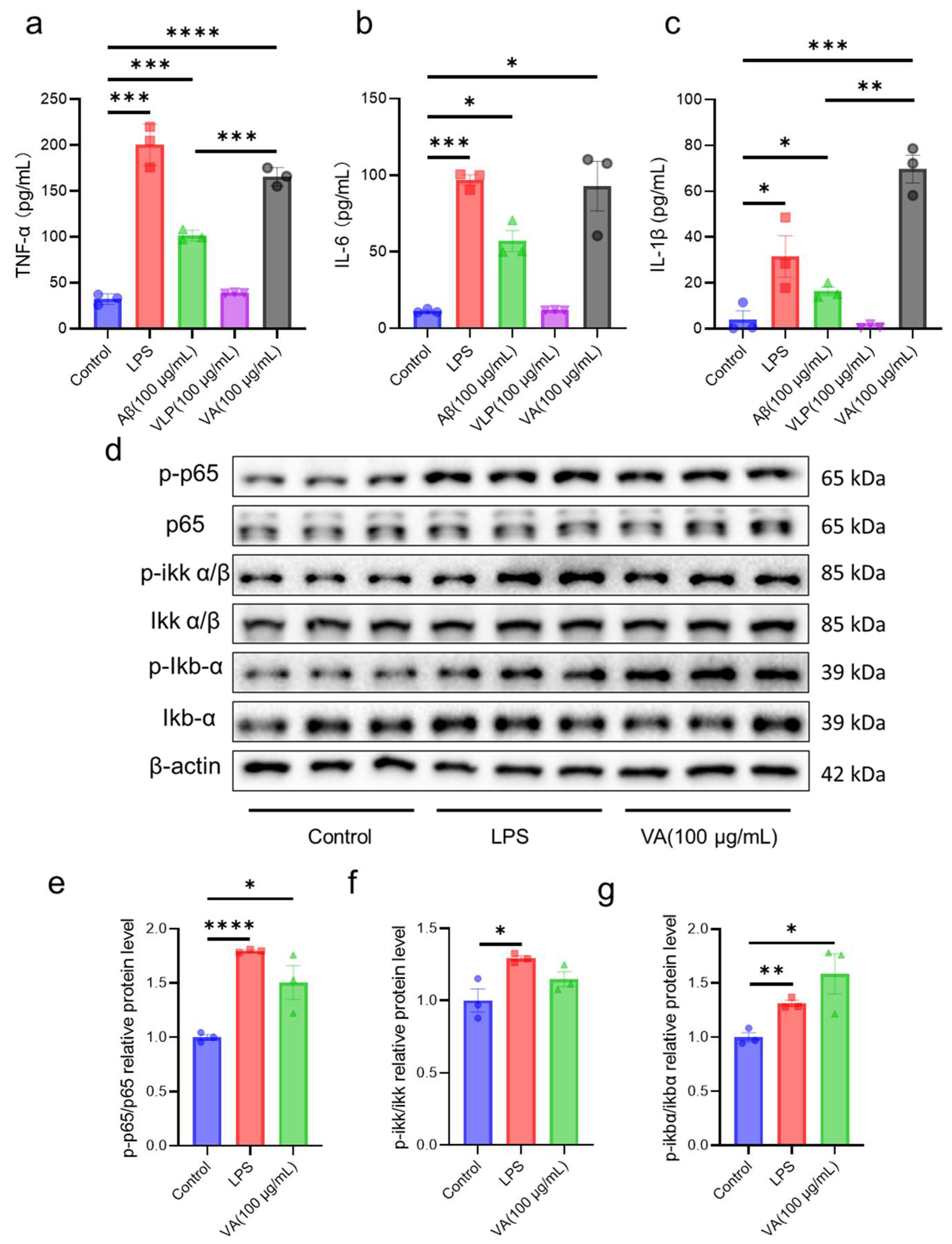
2.3. Hybrid Peptide VA Activates the NF-κB Pathway in an MD2-Dependent Manner
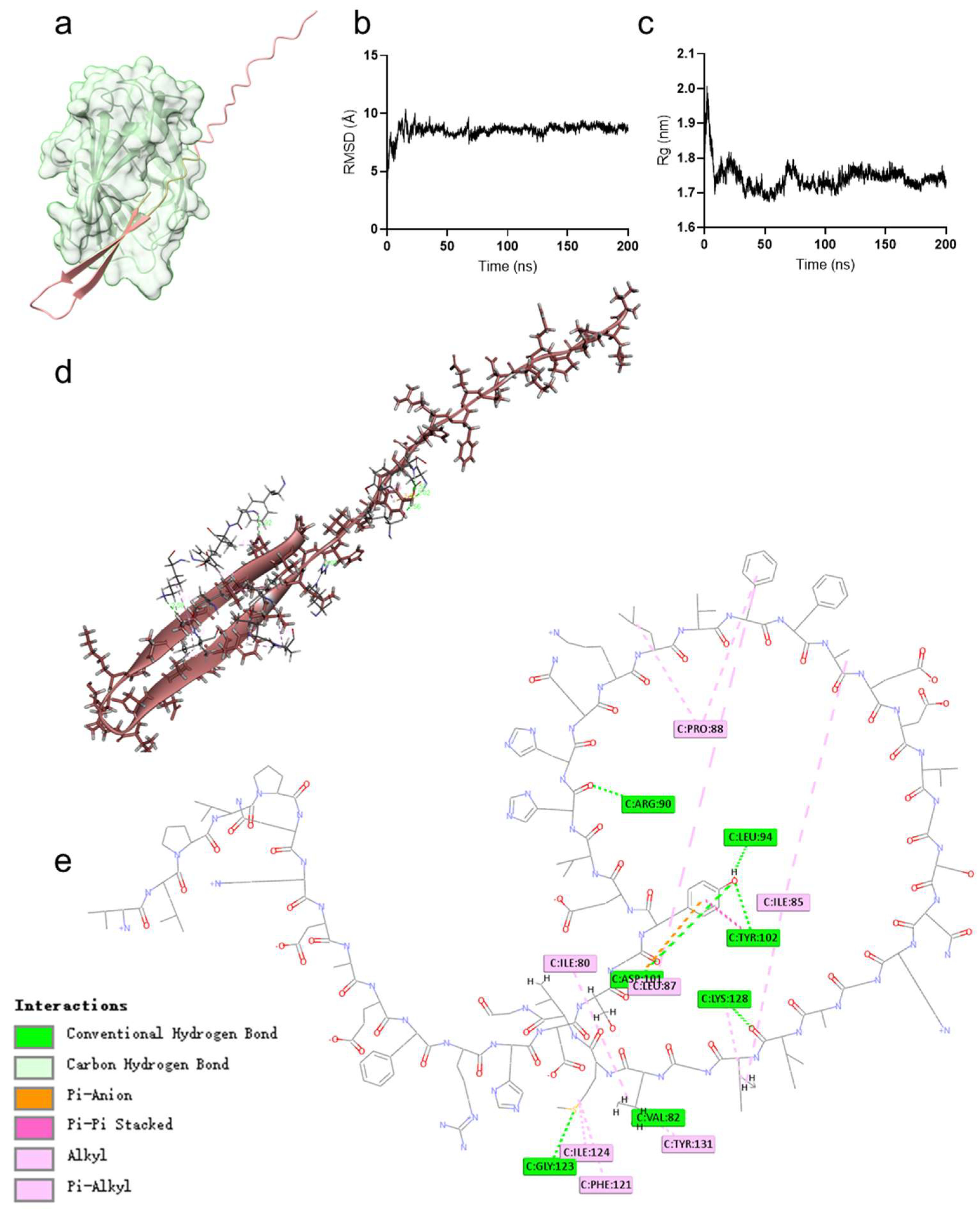
2.4. Hybrid Peptide VA Exerts Immunoregulatory Effects by Binding to the MD2 Hydrophobic Pocket
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Culture
4.3. Animal Models
4.3.1. Organ Index Calculation
4.3.2. ELISA Measurement
4.3.3. RT-PCR
4.3.4. Flow Cytometry Analysis of Spleen T Cell Subpopulations
4.3.5. Proliferation Assay for Spleen Lymphocytes
4.3.6. NK Cell Cytotoxicity Assay
4.4. Enzyme-Linked Immunosorbent Assay (ELISA)
4.5. Western Blot Analysis
4.6. Interaction Analysis of Hybrid Peptide VA with MD2
4.7. Molecular Docking and Dynamics Simulation
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, J.; Han, C.; Lu, Z.; Ge, P.; Cui, Y.; Zhao, D.; Yang, X.; Wu, B.; Qiang, L.; Zhang, Y.; et al. Simulated microgravity suppresses MAPK pathway-mediated innate immune response to bacterial infection and induces gut microbiota dysbiosis. FASEB J. 2020, 34, 14631–14644. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Park, S.; Hwang, S.H.; Lee, E.Y.; Kim, J.H.; Lee, G.S.; Lee, G.; Chang, D.H.; Lee, J.G.; Hwang, J.; et al. Secreted Akkermansia muciniphila threonyl-tRNA synthetase functions to monitor and modulate immune homeostasis. Cell Host Microbe 2023, 31, 1021–1037.e1010. [Google Scholar] [CrossRef]
- Wraith, D.C. The Future of Immunotherapy: A 20-Year Perspective. Front. Immunol. 2017, 8, 1668. [Google Scholar] [CrossRef]
- Castro-Gomez, S.; Heneka, M.T. Innate immune activation in neurodegenerative diseases. Immunity 2024, 57, 790–814. [Google Scholar] [CrossRef]
- Wei, X.; Zhang, C. A universe of second messengers for cGLR-STING signaling. Immunity 2023, 56, 1975–1977. [Google Scholar] [CrossRef]
- Wang, J.; Tang, Y.; Zhao, X.; Ding, Z.; Ahmat, M.; Si, D.; Zhang, R.; Wei, X. Molecular hybridization modification improves the stability and immunomodulatory activity of TP5 peptide. Front. Immunol. 2024, 15, 1472839. [Google Scholar] [CrossRef] [PubMed]
- Goldszmid, R.S.; Dzutsev, A.; Trinchieri, G. Host immune response to infection and cancer: Unexpected commonalities. Cell Host Microbe 2014, 15, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Miao, X.; Liu, X.; Chen, H.; Wang, C.; Diao, J. Restoration and preservation effects of mung bean antioxidant peptides on H(2)O(2)-induced WRL-68 cells via Keap1-Nrf2 pathway. Food Sci. Nutr. 2023, 11, 7130–7144. [Google Scholar] [CrossRef]
- Pei, X.D.; He, Y.N.; Wu, Q.L.; Zhang, Y.M.; Li, F.; Jiao, D.Q.; Liu, X.L.; Wang, C.H. Novel Antioxidant Peptides Derived from Feather Keratin Alleviate H(2)O(2)-Induced Oxidative Damage in HepG2 Cells via Keap1/Nrf2 Pathway. J. Agric. Food Chem. 2023, 71, 20062–20072. [Google Scholar] [CrossRef]
- Xiao, Y.; Yang, J.; Deng, Y.; Zhang, L.; Xu, Q.; Li, H. Tartary buckwheat protein-derived peptide AFYRW alleviates H(2)O(2)-induced vascular injury via the PI3K/AKT/NF-κB pathway. Prostaglandins Other Lipid Mediat. 2023, 169, 106768. [Google Scholar] [CrossRef]
- Wang, J.; Zhou, Y.; Zhang, J.; Tong, Y.; Abbas, Z.; Zhao, X.; Li, Z.; Zhang, H.; Chen, S.; Si, D.; et al. Peptide TaY Attenuates Inflammatory Responses by Interacting with Myeloid Differentiation 2 and Inhibiting NF-κB Signaling Pathway. Molecules 2024, 29, 4843. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhang, R.; Wang, J.; Tong, Y.; Zhang, J.; Li, Z.; Zhang, H.; Abbas, Z.; Si, D.; Wei, X. Isolation, Characterization, and Functional Properties of Antioxidant Peptides from Mulberry Leaf Enzymatic Hydrolysates. Antioxidants 2024, 13, 854. [Google Scholar] [CrossRef]
- Wang, L.F.; Chen, J.; Shan, H.J.; Li, W. Synthesis of TP3 fragment via one pot strategy and its immune regulatory activity. Chem. Res. Chin. Univ. 2005, 21, 566–568. [Google Scholar]
- Chimen, M.; McGettrick, H.M.; Yates, C.M.; Kennedy, A.; Martin, A.; Barone, F.; Walker, L.S.K.; Buckley, C.D.; Nash, G.B.; Rainger, G.E.; et al. A novel immune regulatory peptide, PEPITEM (peptide inhibitor of trans-endothelial migration), controls T-cell trafficking during inflammation, a tonic inhibitory pathway that is lost in Type 1 diabetes. Diabet. Med. 2014, 31, 46. [Google Scholar]
- Zou, X.G.; Chi, Y.; Cao, Y.Q.; Zheng, M.; Deng, Z.Y.; Cai, M.; Yang, K.; Sun, P.L. Preparation Process Optimization of Peptides from Agaricus blazei Murrill, and Comparison of Their Antioxidant and Immune-Enhancing Activities Separated by Ultrafiltration Membrane Technology. Foods 2023, 12, 251. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Mai, Y.; Wei, Y.; Yu, N.; Gao, T.; Yang, J. Therapeutic potential of tolerance-based peptide vaccines in autoimmune diseases. Int. Immunopharmacol. 2023, 116, 109740. [Google Scholar] [CrossRef]
- Wei, X.; Zhang, L.; Yang, Y.; Hou, Y.; Xu, Y.; Wang, Z.; Su, H.; Han, F.; Han, J.; Liu, P.; et al. LL-37 transports immunoreactive cGAMP to activate STING signaling and enhance interferon-mediated host antiviral immunity. Cell Rep. 2022, 39, 110880. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, R.; Zhao, X.; Zhang, J.; Tong, Y.; Abbas, Z.; Li, Z.; Zhang, H.; Si, D.; Wei, X. Hybridization Design and High-Throughput Screening of Peptides with Immunomodulatory and Antioxidant Activities. Int. J. Mol. Sci. 2025, 26, 505. [Google Scholar] [CrossRef]
- Emadi, A.; Jones, R.J.; Brodsky, R.A. Cyclophosphamide and cancer: Golden anniversary. Nat. Rev. Clin. Oncol. 2009, 6, 638–647. [Google Scholar] [CrossRef]
- Zhong, S.; Huang, M.; Yang, X.; Liang, L.; Wang, Y.; Romkes, M.; Duan, W.; Chan, E.; Zhou, S.F. Relationship of glutathione S-transferase genotypes with side-effects of pulsed cyclophosphamide therapy in patients with systemic lupus erythematosus. Br. J. Clin. Pharmacol. 2006, 62, 457–472. [Google Scholar] [CrossRef]
- Shirani, K.; Hassani, F.V.; Razavi-Azarkhiavi, K.; Heidari, S.; Zanjani, B.R.; Karimi, G. Phytotrapy of cyclophosphamide-induced immunosuppression. Environ. Toxicol. Pharmacol. 2015, 39, 1262–1275. [Google Scholar] [CrossRef] [PubMed]
- Mei, Y.X.; Chen, H.X.; Zhang, J.; Zhang, X.D.; Liang, Y.X. Protective effect of chitooligosaccharides against cyclophosphamide-induced immunosuppression in mice. Int. J. Biol. Macromol. 2013, 62, 330–335. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Guo, Z.; Song, L.; Ma, J.X.; Ma, Y.Q.; Shang, L.N.; Meng, Y.P.; Fan, Z.Q.; Hao, M.H.; Zhao, J. Role of DNA damage response in cyclophosphamide-induced premature ovarian failure in mice. J. Obstet. Gynaecol. Res. 2024, 50, 1655–1666. [Google Scholar] [CrossRef]
- Du, X.R.; Yan, Y.H.; Dai, Y.F.; Xu, R.J. Yogurt Alleviates Cyclophosphamide-Induced Immunosuppression in Mice through D-Lactate. Nutrients 2024, 16, 1395. [Google Scholar] [CrossRef] [PubMed]
- Seo, B.-c.; Kim, S.-J. The Effect of Angelica gigas Nakai on Immune Enhancement in Cyclophosphamide-induced Immune-suppressed Mice. Biomed. Sci. Lett. 2022, 28, 120–126. [Google Scholar] [CrossRef]
- Jo, K.A.; Kim, K.J.; Park, S.Y.; Jeon, J.Y.; Hwang, J.E.; Kim, J.Y. Evaluation of the Effects of Euglena gracilis on Enhancing Immune Responses in RAW264.7 Cells and a Cyclophosphamide-Induced Mouse Model. J. Microbiol. Biotechnol. 2023, 33, 493–499. [Google Scholar] [CrossRef]
- Liu, C.; Chu, D.W.; Kalantar-Zadeh, K.; George, J.; Young, H.A.; Liu, G.Z. Cytokines: From Clinical Significance to Quantification. Adv. Sci. 2021, 8, 29. [Google Scholar] [CrossRef]
- Pasricha, C.; Bansal, N.; Kaur, R.; Kumari, P.; Jangra, S.; Singh, R. Immunoglobulins: Mechanistic Approaches in Moderation of Various Inflammatory and Anti-Inflammatory Pathways. Curr. Pharm. Biotechnol. 2024, 26, 1950–1970. [Google Scholar] [CrossRef]
- Veremeyko, T.; Barteneva, N.S.; Vorobyev, I.; Ponomarev, E.D. The Emerging Role of Immunoglobulins and Complement in the Stimulation of Neuronal Activity and Repair: Not as Simple as We Thought. Biomolecules 2024, 14, 1323. [Google Scholar] [CrossRef]
- Wang, G.C.; Zhou, X.L.; Yang, M.; Ma, X.Y.; Liu, X. Norcantharidin alleviates cyclophosphamide-induced immunosuppression and leukopenia in mice through NF ΚB pathway. Trop. J. Pharm. Res. 2023, 22, 297–303. [Google Scholar] [CrossRef]
- Lewis, S.M.; Williams, A.; Eisenbarth, S.C. Structure and function of the immune system in the spleen. Sci. Immunol. 2019, 4, 12. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Qu, H.; Gao, Z.; Zeng, D.; Wang, J.; Baranenko, D.; Li, Y.; Lu, W. Protective effects of Ulva pertusa polysaccharide and polysaccharide-iron (III) complex on cyclophosphamide induced immunosuppression in mice. Int. J. Biol. Macromol. 2019, 133, 911–919. [Google Scholar] [CrossRef]
- Dai, J.; Chen, J.; Qi, J.; Ding, M.; Liu, W.; Shao, T.; Han, J.; Wang, G. Konjac Glucomannan from Amorphophallus konjac enhances immunocompetence of the cyclophosphamide-induced immunosuppressed mice. Food Sci. Nutr. 2021, 9, 728–735. [Google Scholar] [CrossRef] [PubMed]
- Saigusa, R.; Winkels, H.; Ley, K. T cell subsets and functions in atherosclerosis. Nat. Rev. Cardiol. 2020, 17, 387–401. [Google Scholar] [CrossRef] [PubMed]
- Laidlaw, B.J.; Craft, J.E.; Kaech, S.M. The multifaceted role of CD4+ T cells in CD8+ T cell memory. Nat. Rev. Immunol. 2016, 16, 102–111. [Google Scholar] [CrossRef]
- Wang, Y.; Qi, Q.; Li, A.; Yang, M.; Huang, W.; Xu, H.; Zhao, Z.; Li, S. Immuno-enhancement effects of Yifei Tongluo Granules on cyclophosphamide-induced immunosuppression in Balb/c mice. J. Ethnopharmacol. 2016, 194, 72–82. [Google Scholar] [CrossRef]
- Yu, Q.; Nie, S.P.; Wang, J.Q.; Liu, X.Z.; Yin, P.F.; Huang, D.F.; Li, W.J.; Gong, D.M.; Xie, M.Y. Chemoprotective effects of Ganoderma atrum polysaccharide in cyclophosphamide-induced mice. Int. J. Biol. Macromol. 2014, 64, 395–401. [Google Scholar] [CrossRef]
- Myers, J.A.; Miller, J.S. Exploring the NK cell platform for cancer immunotherapy. Nat. Rev. Clin. Oncol. 2021, 18, 85–100. [Google Scholar] [CrossRef]
- Kawai, T.; Ikegawa, M.; Ori, D.; Akira, S. Decoding Toll-like receptors: Recent insights and perspectives in innate immunity. Immunity 2024, 57, 649–673. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, H.; Lee, J.H.; Hwangbo, C. Toll-like receptor 4 (TLR4): New insight immune and aging. Immun. Ageing 2023, 20, 67. [Google Scholar] [CrossRef]
- Fitzgerald, K.A.; Kagan, J.C. Toll-like Receptors and the Control of Immunity. Cell 2020, 180, 1044–1066. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.Y.; Xu, X.Q.; Fu, D.; Gu, Y.B.; Fan, R.; Yi, H.M.; He, X.Y.; Wang, C.F.; Ouyang, B.S.; Zhao, P.; et al. Butyrate-producing Eubacterium rectale suppresses lymphomagenesis by alleviating the TNF-induced TLR4/MyD88/NF-κB axis. Cell Host Microbe 2022, 30, 1139–1150. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.K.; Kim, S.J.; Rah, S.H.; Kang, J.I.; Jung, H.E.; Lee, D.; Lee, H.K.; Lee, J.O.; Park, B.S.; Yoon, T.Y.; et al. Reconstruction of LPS Transfer Cascade Reveals Structural Determinants within LBP, CD14, and TLR4-MD2 for Efficient LPS Recognition and Transfer. Immunity 2017, 46, 38–50. [Google Scholar] [CrossRef]
- Wang, Y.; Shan, X.; Chen, G.; Jiang, L.; Wang, Z.; Fang, Q.; Liu, X.; Wang, J.; Zhang, Y.; Wu, W.; et al. MD-2 as the target of a novel small molecule, L6H21, in the attenuation of LPS-induced inflammatory response and sepsis. Br. J. Pharmacol. 2015, 172, 4391–4405. [Google Scholar] [CrossRef]
- Park, S.H.; Kim, N.D.; Jung, J.K.; Lee, C.K.; Han, S.B.; Kim, Y. Myeloid differentiation 2 as a therapeutic target of inflammatory disorders. Pharmacol. Ther. 2012, 133, 291–298. [Google Scholar] [CrossRef]
- Kim, H.M.; Park, B.S.; Kim, J.I.; Kim, S.E.; Lee, J.; Oh, S.C.; Enkhbayar, P.; Matsushima, N.; Lee, H.; Yoo, O.J.; et al. Crystal structure of the TLR4-MD-2 complex with bound endotoxin antagonist Eritoran. Cell 2007, 130, 906–917. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Chen, G.; Chen, L.; Liu, X.; Fu, W.; Zhang, Y.; Li, C.; Liang, G.; Cai, Y. Insights into the binding mode of curcumin to MD-2: Studies from molecular docking, molecular dynamics simulations and experimental assessments. Mol. Biosyst. 2015, 11, 1933–1938. [Google Scholar] [CrossRef]
- Park, B.S.; Song, D.H.; Kim, H.M.; Choi, B.S.; Lee, H.; Lee, J.O. The structural basis of lipopolysaccharide recognition by the TLR4-MD-2 complex. Nature 2009, 458, 1191–1195. [Google Scholar] [CrossRef]
- Li, Y.; Yu, Z.; Schenk, M.; Lagovsky, I.; Illig, D.; Walz, C.; Rohlfs, M.; Conca, R.; Muise, A.M.; Snapper, S.B.; et al. Human MD2 deficiency-an inborn error of immunity with pleiotropic features. J. Allergy Clin. Immunol. 2023, 151, 791–796.e797. [Google Scholar] [CrossRef]
- Roh, E.; Lee, H.S.; Kwak, J.A.; Hong, J.T.; Nam, S.Y.; Jung, S.H.; Lee, J.Y.; Kim, N.D.; Han, S.B.; Kim, Y. MD-2 as the target of nonlipid chalcone in the inhibition of endotoxin LPS-induced TLR4 activity. J. Infect. Dis. 2011, 203, 1012–1020. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Sequence |
|---|---|
| Aβ | DAEFRHDSGYEVHHQKLVFFAEDVGSNKGAIIGLMVG |
| VLP | VLPVPQK |
| VLP-Aβ | VLPVPQKDAEFRHDSGYEVHHQKLVFFAEDVGSNKGAIIGLMVG |
| Name | Distance | Category | From | To |
|---|---|---|---|---|
| C:ARG90:HH21–A:HIS20:O | 3.07 | Hydrogen Bond | C:ARG90:HH21 | A:HIS20:O |
| C:ASP101:HN–A:TYR17:OH | 2.02 | Hydrogen Bond | C:ASP101:HN | A:TYR17:OH |
| C:TYR102:HN–A:TYR17:OH | 2.57 | Hydrogen Bond | C:TYR102:HN | A:TYR17:OH |
| C:GLY123:HN–A:MET42:SD | 2.92 | Hydrogen Bond | C:GLY123:HN | A:MET42:SD |
| C:LYS128:HZ1–A:ILE38:O | 2.58 | Hydrogen Bond | C:LYS128:HZ1 | A:ILE38:O |
| A:TYR17:HH–C:LEU94:O | 1.56 | Hydrogen Bond | A:TYR17:HH | C:LEU94:O |
| C:LYS128:HE2–A:ILE38:O | 1.71 | Hydrogen Bond | C:LYS128:HE2 | A:ILE38:O |
| C:ASP101:OD2–A:TYR17 | 3.46 | Electrostatic | C:ASP101:OD2 | A:TYR17 |
| C:TYR102–A:TYR17 | 3.79 | Hydrophobic | C:TYR102 | A:TYR17 |
| C:ILE80–A:LEU41 | 4.71 | Hydrophobic | C:ILE80 | A:LEU41 |
| C:ILE85–A:ILE39 | 5.00 | Hydrophobic | C:ILE85 | A:ILE39 |
| C:PRO88–A:LEU24 | 5.43 | Hydrophobic | C:PRO88 | A:LEU24 |
| C:ILE124–A:MET42 | 4.27 | Hydrophobic | C:ILE124 | A:MET42 |
| C:LYS128–A:ILE39 | 5.12 | Hydrophobic | C:LYS128 | A:ILE39 |
| A:ALA28–C:ILE85 | 4.28 | Hydrophobic | A:ALA28 | C:ILE85 |
| C:PHE121–A:MET42 | 4.60 | Hydrophobic | C:PHE121 | A:MET42 |
| C:TYR131–A:LEU41 | 5.09 | Hydrophobic | C:TYR131 | A:LEU41 |
| A:PHE26–C:LEU87 | 4.08 | Hydrophobic | A:PHE26 | C:LEU87 |
| A:PHE26–C:PRO88 | 5.00 | Hydrophobic | A:PHE26 | C:PRO88 |
| Gene | Sequence (5′–3′) | Length | |
|---|---|---|---|
| TNF-α | F | GGCCAACGGCATGGATCTCAAA | 22 |
| R | TAGCAAATCGGCTGACGGTGTG | 22 | |
| IL-6 | F | TCTTGGGACTGATGCTGGTGA | 21 |
| R | TTGGGAGTGGTATCCTCTGTGAA | 23 | |
| IL-1β | F | AATCTCGCAGCAGCACATCAACA | 23 |
| R | ACACCAGCAGGTTATCATCATCATCC | 26 | |
| β-actin | F | TCACTATTGGCAACGAGCGGTTC | 23 |
| R | CAGCACTGTGTTGGCATAGAGGTC | 24 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Zhao, X.; Zhang, R.; Zhang, J.; Tong, Y.; Abbas, Z.; Si, D.; Wei, X. A Novel Hybrid Peptide VLP-Aβ Efficiently Regulates Immunity by Stimulating Myeloid Differentiation Protein and Activating the NF-κB Pathway. Int. J. Mol. Sci. 2025, 26, 5834. https://doi.org/10.3390/ijms26125834
Wang J, Zhao X, Zhang R, Zhang J, Tong Y, Abbas Z, Si D, Wei X. A Novel Hybrid Peptide VLP-Aβ Efficiently Regulates Immunity by Stimulating Myeloid Differentiation Protein and Activating the NF-κB Pathway. International Journal of Molecular Sciences. 2025; 26(12):5834. https://doi.org/10.3390/ijms26125834
Chicago/Turabian StyleWang, Junyong, Xuelian Zhao, Rijun Zhang, Jing Zhang, Yucui Tong, Zaheer Abbas, Dayong Si, and Xubiao Wei. 2025. "A Novel Hybrid Peptide VLP-Aβ Efficiently Regulates Immunity by Stimulating Myeloid Differentiation Protein and Activating the NF-κB Pathway" International Journal of Molecular Sciences 26, no. 12: 5834. https://doi.org/10.3390/ijms26125834
APA StyleWang, J., Zhao, X., Zhang, R., Zhang, J., Tong, Y., Abbas, Z., Si, D., & Wei, X. (2025). A Novel Hybrid Peptide VLP-Aβ Efficiently Regulates Immunity by Stimulating Myeloid Differentiation Protein and Activating the NF-κB Pathway. International Journal of Molecular Sciences, 26(12), 5834. https://doi.org/10.3390/ijms26125834