
Advances in Regenerative Therapies for Inflammatory Arthritis: Exploring the Potential of Mesenchymal Stem Cells and Extracellular Vesicles

 ,
,  ,
,
Abstract
1. Introduction
2. Cell Therapy
2.1. The Immune-Modulating Effect
2.2. ECM Synthesis
2.3. Chondrogenesis
2.4. Tissue Engineering and MSC
3. Therapy of OA and RA with Unmodified EV
4. Modified Extracellular Vesicles
4.1. Hypoxic Method
4.2. Preconditioning with Pro-Inflammatory Factors and 3D Cultivation Conditions
4.3. Genetic and Drug Modifications
4.4. Surface Modification of EV
4.5. Application of Tissue Bioengineering Approaches in EV Modification for Inflammatory Arthritis Therapy
5. Challenges and Prospects of Cell Therapy and MSC-Derived EV Therapy on the Path to Clinical Practice
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Lin, Y.-J.; Anzaghe, M.; Schülke, S. Update on the Pathomechanism, Diagnosis, and Treatment Options for Rheumatoid Arthritis. Cells 2020, 9, 880. [Google Scholar] [CrossRef] [PubMed]
- Hunter, D.J.; March, L.; Chew, M. Osteoarthritis in 2020 and beyond: A Lancet Commission. Lancet 2020, 396, 1711–1712. [Google Scholar] [CrossRef] [PubMed]
- Aletaha, D.; Smolen, J.S. Diagnosis and Management of Rheumatoid Arthritis: A Review. JAMA 2018, 320, 1360–1372. [Google Scholar] [CrossRef] [PubMed]
- Myasoedova, E.; Davis, J.; Matteson, E.L.; Crowson, C.S. Is the Epidemiology of Rheumatoid Arthritis Changing? Results from a Population-Based Incidence Study, 1985–2014. Ann. Rheum. Dis. 2020, 79, 440–444. [Google Scholar] [CrossRef]
- Zhao, X.; Shah, D.; Gandhi, K.; Wei, W.; Dwibedi, N.; Webster, L.; Sambamoorthi, U. Clinical, Humanistic, and Economic Burden of Osteoarthritis among Noninstitutionalized Adults in the United States. Osteoarthr. Cartil. 2019, 27, 1618–1626. [Google Scholar] [CrossRef]
- Katz, J.N.; Arant, K.R.; Loeser, R.F. Diagnosis and Treatment of Hip and Knee Osteoarthritis: A Review. JAMA 2021, 325, 568–578. [Google Scholar] [CrossRef]
- Conforti, A.; Di Cola, I.; Pavlych, V.; Ruscitti, P.; Berardicurti, O.; Ursini, F.; Giacomelli, R.; Cipriani, P. Beyond the Joints, the Extra-Articular Manifestations in Rheumatoid Arthritis. Autoimmun. Rev. 2021, 20, 102735. [Google Scholar] [CrossRef]
- Raychaudhuri, S.; Sandor, C.; Stahl, E.A.; Freudenberg, J.; Lee, H.-S.; Jia, X.; Alfredsson, L.; Padyukov, L.; Klareskog, L.; Worthington, J.; et al. Five Amino Acids in Three HLA Proteins Explain Most of the Association between MHC and Seropositive Rheumatoid Arthritis. Nat. Genet. 2012, 44, 291–296. [Google Scholar] [CrossRef]
- Darrah, E.; Andrade, F. Rheumatoid Arthritis and Citrullination. Curr. Opin. Rheumatol. 2018, 30, 72. [Google Scholar] [CrossRef]
- Sen, R.; Hurley, J.A. Osteoarthritis. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2024. [Google Scholar]
- Gelber, A.C.; Hochberg, M.C.; Mead, L.A.; Wang, N.-Y.; Wigley, F.M.; Klag, M.J. Joint Injury in Young Adults and Risk for Subsequent Knee and Hip Osteoarthritis. Ann. Intern. Med. 2000, 133, 321–328. [Google Scholar] [CrossRef]
- Sahin, V.; Karakaş, E.S.; Aksu, S.; Atlihan, D.; Turk, C.Y.; Halici, M. Traumatic Dislocation and Fracture-Dislocation of the Hip: A Long-Term Follow-up Study. J. Trauma 2003, 54, 520–529. [Google Scholar] [CrossRef] [PubMed]
- Abramoff, B.; Caldera, F.E. Osteoarthritis: Pathology, Diagnosis, and Treatment Options. Med. Clin. N. Am. 2020, 104, 293–311. [Google Scholar] [CrossRef] [PubMed]
- Mort, J.S.; Billington, C.J. Articular Cartilage and Changes in Arthritis: Matrix Degradation. Arthritis Res. Ther. 2001, 3, 337. [Google Scholar] [CrossRef] [PubMed]
- Xia, B.; Chen, D.; Zhang, J.; Hu, S.; Jin, H.; Tong, P. Osteoarthritis Pathogenesis: A Review of Molecular Mechanisms. Calcif. Tissue Int. 2014, 95, 495–505. [Google Scholar] [CrossRef]
- Lambert, C.; Zappia, J.; Sanchez, C.; Florin, A.; Dubuc, J.-E.; Henrotin, Y. The Damage-Associated Molecular Patterns (DAMPs) as Potential Targets to Treat Osteoarthritis: Perspectives from a Review of the Literature. Front. Med. 2021, 7, 607186. [Google Scholar] [CrossRef]
- Neogi, T. Clinical Significance of Bone Changes in Osteoarthritis. Ther. Adv. Musculoskelet. Dis. 2012, 4, 259–267. [Google Scholar] [CrossRef]
- Buckley, C.D.; McGettrick, H.M. Leukocyte Trafficking between Stromal Compartments: Lessons from Rheumatoid Arthritis. Nat. Rev. Rheumatol. 2018, 14, 476–487. [Google Scholar] [CrossRef]
- Alivernini, S.; MacDonald, L.; Elmesmari, A.; Finlay, S.; Tolusso, B.; Gigante, M.R.; Petricca, L.; Di Mario, C.; Bui, L.; Perniola, S.; et al. Distinct Synovial Tissue Macrophage Subsets Regulate Inflammation and Remission in Rheumatoid Arthritis. Nat. Med. 2020, 26, 1295–1306. [Google Scholar] [CrossRef]
- Zhao, S.; Grieshaber-Bouyer, R.; Rao, D.A.; Kolb, P.; Chen, H.; Andreeva, I.; Tretter, T.; Lorenz, H.-M.; Watzl, C.; Wabnitz, G.; et al. Effect of JAK Inhibition on the Induction of Proinflammatory HLA–DR+CD90+ Rheumatoid Arthritis Synovial Fibroblasts by Interferon-γ. Arthritis Rheumatol. 2022, 74, 441–452. [Google Scholar] [CrossRef]
- Floudas, A.; Neto, N.; Orr, C.; Canavan, M.; Gallagher, P.; Hurson, C.; Monaghan, M.G.; Nagpar, S.; Mullan, R.H.; Veale, D.J.; et al. Loss of Balance between Protective and Pro-Inflammatory Synovial Tissue T-Cell Polyfunctionality Predates Clinical Onset of Rheumatoid Arthritis. Ann. Rheum. Dis. 2022, 81, 193–205. [Google Scholar] [CrossRef]
- Smolen, J.S.; Aletaha, D.; Barton, A.; Burmester, G.R.; Emery, P.; Firestein, G.S.; Kavanaugh, A.; McInnes, I.B.; Solomon, D.H.; Strand, V.; et al. Rheumatoid Arthritis. Nat. Rev. Dis. Primer 2018, 4, 18001. [Google Scholar] [CrossRef]
- Luo, G.; Li, F.; Li, X.; Wang, Z.-G.; Zhang, B. TNF-α and RANKL Promote Osteoclastogenesis by Upregulating RANK via the NF-κB Pathway. Mol. Med. Rep. 2018, 17, 6605–6611. [Google Scholar] [CrossRef]
- Meednu, N.; Zhang, H.; Owen, T.; Sun, W.; Wang, V.; Cistrone, C.; Rangel-Moreno, J.; Xing, L.; Anolik, J.H. Production of RANKL by Memory B Cells. Arthritis Rheumatol. 2016, 68, 805–816. [Google Scholar] [CrossRef]
- Ouboussad, L.; Burska, A.N.; Melville, A.; Buch, M.H. Synovial Tissue Heterogeneity in Rheumatoid Arthritis and Changes with Biologic and Targeted Synthetic Therapies to Inform Stratified Therapy. Front. Med. 2019, 6, 45. [Google Scholar] [CrossRef]
- Peng, X.; Chen, X.; Zhang, Y.; Tian, Z.; Wang, M.; Chen, Z. Advances in the Pathology and Treatment of Osteoarthritis. J. Adv. Res. 2025, in press. [Google Scholar] [CrossRef]
- Han, P.; Liu, X.; He, J.; Han, L.; Li, J. Overview of Mechanisms and Novel Therapies on Rheumatoid Arthritis from a Cellular Perspective. Front. Immunol. 2024, 15, 1461756. [Google Scholar] [CrossRef]
- Coaccioli, S.; Sarzi-Puttini, P.; Zis, P.; Rinonapoli, G.; Varrassi, G. Osteoarthritis: New Insight on Its Pathophysiology. J. Clin. Med. 2022, 11, 6013. [Google Scholar] [CrossRef]
- Bannuru, R.R.; Osani, M.C.; Vaysbrot, E.E.; Arden, N.K.; Bennell, K.; Bierma-Zeinstra, S.M.A.; Kraus, V.B.; Lohmander, L.S.; Abbott, J.H.; Bhandari, M.; et al. OARSI Guidelines for the Non-Surgical Management of Knee, Hip, and Polyarticular Osteoarthritis. Osteoarthr. Cartil. 2019, 27, 1578–1589. [Google Scholar] [CrossRef]
- Archer, R.; Hock, E.; Hamilton, J.; Stevens, J.; Essat, M.; Poku, E.; Clowes, M.; Pandor, A.; Stevenson, M. Assessing Prognosis and Prediction of Treatment Response in Early Rheumatoid Arthritis: Systematic Reviews. Health Technol. Assess. Winch. Engl. 2018, 22, 1–294. [Google Scholar] [CrossRef]
- Kerola, A.M.; Rollefstad, S.; Semb, A.G. Atherosclerotic Cardiovascular Disease in Rheumatoid Arthritis: Impact of Inflammation and Antirheumatic Treatment. Eur. Cardiol. Rev. 2021, 16, e18. [Google Scholar] [CrossRef]
- Gomez, E.A.; Colas, R.A.; Souza, P.R.; Hands, R.; Lewis, M.J.; Bessant, C.; Pitzalis, C.; Dalli, J. Blood Pro-Resolving Mediators Are Linked with Synovial Pathology and Are Predictive of DMARD Responsiveness in Rheumatoid Arthritis. Nat. Commun. 2020, 11, 5420. [Google Scholar] [CrossRef]
- Bindu, S.; Mazumder, S.; Bandyopadhyay, U. Non-Steroidal Anti-Inflammatory Drugs (NSAIDs) and Organ Damage: A Current Perspective. Biochem. Pharmacol. 2020, 180, 114147. [Google Scholar] [CrossRef]
- Kloppenburg, M.; Berenbaum, F. Osteoarthritis Year in Review 2019: Epidemiology and Therapy. Osteoarthr. Cartil. 2020, 28, 242–248. [Google Scholar] [CrossRef]
- Tschopp, M.; Pfirrmann, C.W.A.; Fucentese, S.F.; Brunner, F.; Catanzaro, S.; Kühne, N.; Zwyssig, I.; Sutter, R.; Götschi, T.; Tanadini, M.; et al. A Randomized Trial of Intra-Articular Injection Therapy for Knee Osteoarthritis. Investig. Radiol. 2023, 58, 355–362. [Google Scholar] [CrossRef]
- Robinson, P.D.; McEwan, J.; Adukia, V.; Prabhakar, M. Osteoarthritis and Arthroplasty of the Hip and Knee. Br. J. Hosp. Med. 2018, 79, C54–C59. [Google Scholar] [CrossRef]
- Regmi, S.; Pathak, S.; Kim, J.O.; Yong, C.S.; Jeong, J.-H. Mesenchymal Stem Cell Therapy for the Treatment of Inflammatory Diseases: Challenges, Opportunities, and Future Perspectives. Eur. J. Cell Biol. 2019, 98, 151041. [Google Scholar] [CrossRef]
- Chung, M.-J.; Son, J.-Y.; Park, S.; Park, S.-S.; Hur, K.; Lee, S.-H.; Lee, E.-J.; Park, J.-K.; Hong, I.-H.; Kim, T.-H.; et al. Mesenchymal Stem Cell and MicroRNA Therapy of Musculoskeletal Diseases. Int. J. Stem Cells 2021, 14, 150–167. [Google Scholar] [CrossRef]
- Ragni, E.; Perucca Orfei, C.; De Luca, P.; Mondadori, C.; Viganò, M.; Colombini, A.; de Girolamo, L. Inflammatory Priming Enhances Mesenchymal Stromal Cell Secretome Potential as a Clinical Product for Regenerative Medicine Approaches through Secreted Factors and EV-miRNAs: The Example of Joint Disease. Stem Cell Res. Ther. 2020, 11, 165. [Google Scholar] [CrossRef]
- Hernigou, P.; Bouthors, C.; Bastard, C.; Flouzat Lachaniette, C.H.; Rouard, H.; Dubory, A. Subchondral Bone or Intra-Articular Injection of Bone Marrow Concentrate Mesenchymal Stem Cells in Bilateral Knee Osteoarthritis: What Better Postpone Knee Arthroplasty at Fifteen Years? A Randomized Study. Int. Orthop. 2021, 45, 391–399. [Google Scholar] [CrossRef]
- Liu, H.; Li, R.; Liu, T.; Yang, L.; Yin, G.; Xie, Q. Immunomodulatory Effects of Mesenchymal Stem Cells and Mesenchymal Stem Cell-Derived Extracellular Vesicles in Rheumatoid Arthritis. Front. Immunol. 2020, 11, 1912. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal Criteria for Defining Multipotent Mesenchymal Stromal Cells. The International Society for Cellular Therapy Position Statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Kobolak, J.; Dinnyes, A.; Memic, A.; Khademhosseini, A.; Mobasheri, A. Mesenchymal Stem Cells: Identification, Phenotypic Characterization, Biological Properties and Potential for Regenerative Medicine through Biomaterial Micro-Engineering of Their Niche. Methods 2016, 99, 62–68. [Google Scholar] [CrossRef]
- Ilas, D.C.; Churchman, S.M.; Baboolal, T.; Giannoudis, P.V.; Aderinto, J.; McGonagle, D.; Jones, E. The Simultaneous Analysis of Mesenchymal Stem Cells and Early Osteocytes Accumulation in Osteoarthritic Femoral Head Sclerotic Bone. Rheumatology 2019, 58, 1777–1783. [Google Scholar] [CrossRef]
- Turajane, T.; Chaveewanakorn, U.; Fongsarun, W.; Aojanepong, J.; Papadopoulos, K.I. Avoidance of Total Knee Arthroplasty in Early Osteoarthritis of the Knee with Intra-Articular Implantation of Autologous Activated Peripheral Blood Stem Cells versus Hyaluronic Acid: A Randomized Controlled Trial with Differential Effects of Growth Factor Addition. Stem Cells Int. 2017, 2017, 8925132. [Google Scholar] [CrossRef]
- Wu, X.; Wang, W.; Meng, C.; Yang, S.; Duan, D.; Xu, W.; Liu, X.; Tang, M.; Wang, H. Regulation of Differentiation in Trabecular Bone-derived Mesenchymal Stem Cells by T Cell Activation and Inflammation. Oncol. Rep. 2013, 30, 2211–2219. [Google Scholar] [CrossRef]
- Neybecker, P.; Henrionnet, C.; Pape, E.; Mainard, D.; Galois, L.; Loeuille, D.; Gillet, P.; Pinzano, A. In Vitro and in Vivo Potentialities for Cartilage Repair from Human Advanced Knee Osteoarthritis Synovial Fluid-Derived Mesenchymal Stem Cells. Stem Cell Res. Ther. 2018, 9, 329. [Google Scholar] [CrossRef]
- Greif, D.N.; Kouroupis, D.; Murdock, C.J.; Griswold, A.J.; Kaplan, L.D.; Best, T.M.; Correa, D. Infrapatellar Fat Pad/Synovium Complex in Early-Stage Knee Osteoarthritis: Potential New Target and Source of Therapeutic Mesenchymal Stem/Stromal Cells. Front. Bioeng. Biotechnol. 2020, 8, 860. [Google Scholar] [CrossRef]
- Chen, X.; Zheng, J.; Yin, L.; Li, Y.; Liu, H. Transplantation of Three Mesenchymal Stem Cells for Knee Osteoarthritis, Which Cell and Type Are More Beneficial? A Systematic Review and Network Meta-Analysis. J. Orthop. Surg. 2024, 19, 366. [Google Scholar] [CrossRef]
- Lee, W.-S.; Kim, H.J.; Kim, K.-I.; Kim, G.B.; Jin, W. Intra-Articular Injection of Autologous Adipose Tissue-Derived Mesenchymal Stem Cells for the Treatment of Knee Osteoarthritis: A Phase IIb, Randomized, Placebo-Controlled Clinical Trial. Stem Cells Transl. Med. 2019, 8, 504–511. [Google Scholar] [CrossRef]
- Freitag, J.; Bates, D.; Wickham, J.; Shah, K.; Huguenin, L.; Tenen, A.; Paterson, K.; Boyd, R. Adipose-Derived Mesenchymal Stem Cell Therapy in the Treatment of Knee Osteoarthritis: A Randomized Controlled Trial. Regen. Med. 2019, 14, 213–230. [Google Scholar] [CrossRef]
- Wang, Z.-G.; He, Z.-Y.; Liang, S.; Yang, Q.; Cheng, P.; Chen, A.-M. Comprehensive Proteomic Analysis of Exosomes Derived from Human Bone Marrow, Adipose Tissue, and Umbilical Cord Mesenchymal Stem Cells. Stem Cell Res. Ther. 2020, 11, 511. [Google Scholar] [CrossRef]
- Wagenbrenner, M.; Heinz, T.; Horas, K.; Jakuscheit, A.; Arnholdt, J.; Herrmann, M.; Rudert, M.; Holzapfel, B.M.; Steinert, A.F.; Weißenberger, M. The Human Arthritic Hip Joint Is a Source of Mesenchymal Stromal Cells (MSCs) with Extensive Multipotent Differentiation Potential. BMC Musculoskelet. Disord. 2020, 21, 297. [Google Scholar] [CrossRef]
- Yu, Y.; Yoon, K.-A.; Kang, T.-W.; Jeon, H.-J.; Sim, Y.-B.; Choe, S.H.; Baek, S.Y.; Lee, S.; Seo, K.-W.; Kang, K.-S. Therapeutic Effect of Long-Interval Repeated Intravenous Administration of Human Umbilical Cord Blood-Derived Mesenchymal Stem Cells in DBA/1 Mice with Collagen-Induced Arthritis. J. Tissue Eng. Regen. Med. 2019, 13, 1134–1142. [Google Scholar] [CrossRef] [PubMed]
- Gowhari Shabgah, A.; Shariati-Sarabi, Z.; Tavakkol-Afshari, J.; Ghasemi, A.; Ghoryani, M.; Mohammadi, M. A Significant Decrease of BAFF, APRIL, and BAFF Receptors Following Mesenchymal Stem Cell Transplantation in Patients with Refractory Rheumatoid Arthritis. Gene 2020, 732, 144336. [Google Scholar] [CrossRef] [PubMed]
- Sobacchi, C.; Palagano, E.; Villa, A.; Menale, C. Soluble Factors on Stage to Direct Mesenchymal Stem Cells Fate. Front. Bioeng. Biotechnol. 2017, 5, 32. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Chen, X.; Huang, Y.; Li, W.; Li, J.; Cao, K.; Cao, G.; Zhang, L.; Li, F.; Roberts, A.I.; et al. Phylogenetic Distinction of iNOS and IDO Function in Mesenchymal Stem Cell-Mediated Immunosuppression in Mammalian Species. Cell Death Differ. 2014, 21, 388–396. [Google Scholar] [CrossRef]
- Sun, A.R.; Panchal, S.K.; Friis, T.; Sekar, S.; Crawford, R.; Brown, L.; Xiao, Y.; Prasadam, I. Obesity-Associated Metabolic Syndrome Spontaneously Induces Infiltration of pro-Inflammatory Macrophage in Synovium and Promotes Osteoarthritis. PLoS ONE 2017, 12, e0183693. [Google Scholar] [CrossRef]
- Weyand, C.M.; Goronzy, J.J. The Immunology of Rheumatoid Arthritis. Nat. Immunol. 2021, 22, 10–18. [Google Scholar] [CrossRef]
- Liu, B.; Zhang, M.; Zhao, J.; Zheng, M.; Yang, H. Imbalance of M1/M2 Macrophages Is Linked to Severity Level of Knee Osteoarthritis. Exp. Ther. Med. 2018, 16, 5009–5014. [Google Scholar] [CrossRef]
- Liu, K.-J.; Wang, C.-J.; Chang, C.-J.; Hu, H.-I.; Hsu, P.-J.; Wu, Y.-C.; Bai, C.-H.; Sytwu, H.-K.; Yen, B.L. Surface Expression of HLA-G Is Involved in Mediating Immunomodulatory Effects of Placenta-Derived Multipotent Cells (PDMCs) towards Natural Killer Lymphocytes. Cell Transplant. 2011, 20, 1721–1730. [Google Scholar] [CrossRef]
- Selmani, Z.; Naji, A.; Zidi, I.; Favier, B.; Gaiffe, E.; Obert, L.; Borg, C.; Saas, P.; Tiberghien, P.; Rouas-Freiss, N.; et al. Human Leukocyte Antigen-G5 Secretion by Human Mesenchymal Stem Cells Is Required to Suppress T Lymphocyte and Natural Killer Function and to Induce CD4+CD25highFOXP3+ Regulatory T Cells. Stem Cells 2008, 26, 212–222. [Google Scholar] [CrossRef]
- Schurgers, E.; Kelchtermans, H.; Mitera, T.; Geboes, L.; Matthys, P. Discrepancy between the in Vitro and in Vivo Effects of Murine Mesenchymal Stem Cells on T-Cell Proliferation and Collagen-Induced Arthritis. Arthritis Res. Ther. 2010, 12, R31. [Google Scholar] [CrossRef]
- Fan, X.-L.; Zhang, Z.; Ma, C.Y.; Fu, Q.-L. Mesenchymal Stem Cells for Inflammatory Airway Disorders: Promises and Challenges. Biosci. Rep. 2019, 39, BSR20182160. [Google Scholar] [CrossRef]
- Song, N.; Scholtemeijer, M.; Shah, K. Mesenchymal Stem Cell Immunomodulation: Mechanisms and Therapeutic Potential. Trends Pharmacol. Sci. 2020, 41, 653–664. [Google Scholar] [CrossRef]
- Hong, J.; Hueckelhoven, A.; Wang, L.; Schmitt, A.; Wuchter, P.; Tabarkiewicz, J.; Kleist, C.; Bieback, K.; Ho, A.D.; Schmitt, M. Indoleamine 2,3-Dioxygenase Mediates Inhibition of Virus-Specific CD8+ T Cell Proliferation by Human Mesenchymal Stromal Cells. Cytotherapy 2016, 18, 621–629. [Google Scholar] [CrossRef]
- Hwu, P.; Du, M.X.; Lapointe, R.; Do, M.; Taylor, M.W.; Young, H.A. Indoleamine 2,3-Dioxygenase Production by Human Dendritic Cells Results in the Inhibition of T Cell Proliferation. J. Immunol. 2000, 164, 3596–3599. [Google Scholar] [CrossRef]
- Duffy, M.M.; Pindjakova, J.; Hanley, S.A.; McCarthy, C.; Weidhofer, G.A.; Sweeney, E.M.; English, K.; Shaw, G.; Murphy, J.M.; Barry, F.P.; et al. Mesenchymal Stem Cell Inhibition of T-helper 17 Cell- Differentiation Is Triggered by Cell–Cell Contact and Mediated by Prostaglandin E2 via the EP4 Receptor. Eur. J. Immunol. 2011, 41, 2840–2851. [Google Scholar] [CrossRef]
- Steffen, U.; Schett, G.; Bozec, A. How Autoantibodies Regulate Osteoclast Induced Bone Loss in Rheumatoid Arthritis. Front. Immunol. 2019, 10, 1483. [Google Scholar] [CrossRef]
- Arai, F.; Miyamoto, T.; Ohneda, O.; Inada, T.; Sudo, T.; Brasel, K.; Miyata, T.; Anderson, D.M.; Suda, T. Commitment and Differentiation of Osteoclast Precursor Cells by the Sequential Expression of C-Fms and Receptor Activator of Nuclear Factor kappaB (RANK) Receptors. J. Exp. Med. 1999, 190, 1741–1754. [Google Scholar] [CrossRef]
- Xing, L.; Xiu, Y.; Boyce, B.F. Osteoclast Fusion and Regulation by RANKL-Dependent and Independent Factors. World J. Orthop. 2012, 3, 212–222. [Google Scholar] [CrossRef]
- Hamdalla, H.M.; Ahmed, R.R.; Galaly, S.R.; Ahmed, O.M.; Naguib, I.A.; Alghamdi, B.S.; Abdul-Hamid, M. Assessment of the Efficacy of Bone Marrow-Derived Mesenchymal Stem Cells against a Monoiodoacetate-Induced Osteoarthritis Model in Wistar Rats. Stem Cells Int. 2022, 2022, 1900403. [Google Scholar] [CrossRef] [PubMed]
- Shin, T.-H.; Kim, H.-S.; Kang, T.-W.; Lee, B.-C.; Lee, H.-Y.; Kim, Y.-J.; Shin, J.-H.; Seo, Y.; Won Choi, S.; Lee, S.; et al. Human Umbilical Cord Blood-Stem Cells Direct Macrophage Polarization and Block Inflammasome Activation to Alleviate Rheumatoid Arthritis. Cell Death Dis. 2016, 7, e2524. [Google Scholar] [CrossRef] [PubMed]
- Schelbergen, R.F.; van Dalen, S.; ter Huurne, M.; Roth, J.; Vogl, T.; Noël, D.; Jorgensen, C.; van den Berg, W.B.; van de Loo, F.A.; Blom, A.B.; et al. Treatment Efficacy of Adipose-Derived Stem Cells in Experimental Osteoarthritis Is Driven by High Synovial Activation and Reflected by S100A8/A9 Serum Levels. Osteoarthr. Cartil. 2014, 22, 1158–1166. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Lee, R.H.; Bazhanov, N.; Oh, J.Y.; Prockop, D.J. Anti-Inflammatory Protein TSG-6 Secreted by Activated MSCs Attenuates Zymosan-Induced Mouse Peritonitis by Decreasing TLR2/NF-κB Signaling in Resident Macrophages. Blood 2011, 118, 330–338. [Google Scholar] [CrossRef]
- Melief, S.M.; Geutskens, S.B.; Fibbe, W.E.; Roelofs, H. Multipotent Stromal Cells Skew Monocytes towards an Anti-Inflammatory Interleukin-10-Producing Phenotype by Production of Interleukin-6. Haematologica 2013, 98, 888–895. [Google Scholar] [CrossRef]
- Song, W.-J.; Li, Q.; Ryu, M.-O.; Ahn, J.-O.; Ha Bhang, D.; Chan Jung, Y.; Youn, H.-Y. TSG-6 Secreted by Human Adipose Tissue-Derived Mesenchymal Stem Cells Ameliorates DSS-Induced Colitis by Inducing M2 Macrophage Polarization in Mice. Sci. Rep. 2017, 7, 5187. [Google Scholar] [CrossRef]
- François, M.; Romieu-Mourez, R.; Li, M.; Galipeau, J. Human MSC Suppression Correlates with Cytokine Induction of Indoleamine 2,3-Dioxygenase and Bystander M2 Macrophage Differentiation. Mol. Ther. 2012, 20, 187–195. [Google Scholar] [CrossRef]
- Oshita, K.; Yamaoka, K.; Udagawa, N.; Fukuyo, S.; Sonomoto, K.; Maeshima, K.; Kurihara, R.; Nakano, K.; Saito, K.; Okada, Y.; et al. Human Mesenchymal Stem Cells Inhibit Osteoclastogenesis through Osteoprotegerin Production. Arthritis Rheum. 2011, 63, 1658–1667. [Google Scholar] [CrossRef]
- Galgaro, B.C.; Beckenkamp, L.R.; Nunnenkamp, M.v.d.M.; Korb, V.G.; Naasani, L.I.S.; Roszek, K.; Wink, M.R. The Adenosinergic Pathway in Mesenchymal Stem Cell Fate and Functions. Med. Res. Rev. 2021, 41, 2316–2349. [Google Scholar] [CrossRef]
- Saldanha-Araujo, F.; Ferreira, F.I.S.; Palma, P.V.; Araujo, A.G.; Queiroz, R.H.C.; Covas, D.T.; Zago, M.A.; Panepucci, R.A. Mesenchymal Stromal Cells Up-Regulate CD39 and Increase Adenosine Production to Suppress Activated T-Lymphocytes. Stem Cell Res. 2011, 7, 66–74. [Google Scholar] [CrossRef]
- Pinto-Cardoso, R.; Pereira-Costa, F.; Pedro Faria, J.; Bandarrinha, P.; Bessa-Andrês, C.; Correia-de-Sá, P.; Bernardo Noronha-Matos, J. Adenosinergic Signalling in Chondrogenesis and Cartilage Homeostasis: Friend or Foe? Biochem. Pharmacol. 2020, 174, 113784. [Google Scholar] [CrossRef] [PubMed]
- Strazzulla, L.C.; Cronstein, B.N. Regulation of Bone and Cartilage by Adenosine Signaling. Purinergic Signal. 2016, 12, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Hazrati, A.; Malekpour, K.; Khorramdelazad, H.; Rajaei, S.; Hashemi, S.M. Therapeutic and Immunomodulatory Potentials of Mesenchymal Stromal/Stem Cells and Immune Checkpoints Related Molecules. Biomark. Res. 2024, 12, 35. [Google Scholar] [CrossRef] [PubMed]
- Corciulo, C.; Lendhey, M.; Wilder, T.; Schoen, H.; Cornelissen, A.S.; Chang, G.; Kennedy, O.D.; Cronstein, B.N. Endogenous Adenosine Maintains Cartilage Homeostasis and Exogenous Adenosine Inhibits Osteoarthritis Progression. Nat. Commun. 2017, 8, 15019. [Google Scholar] [CrossRef]
- Luo, Y.; Wu, W.; Gu, J.; Zhang, X.; Dang, J.; Wang, J.; Zheng, Y.; Huang, F.; Yuan, J.; Xue, Y.; et al. Human Gingival Tissue-Derived MSC Suppress Osteoclastogenesis and Bone Erosion via CD39-Adenosine Signal Pathway in Autoimmune Arthritis. eBioMedicine 2019, 43, 620–631. [Google Scholar] [CrossRef]
- Ilchovska, D.; Barrow, D.M. An Overview of the NF-kB Mechanism of Pathophysiology in Rheumatoid Arthritis, Investigation of the NF-kB Ligand RANKL and Related Nutritional Interventions. Autoimmun. Rev. 2021, 20, 102741. [Google Scholar] [CrossRef]
- Zimmermann, J.A.; Hettiaratchi, M.H.; McDevitt, T.C. Enhanced Immunosuppression of T Cells by Sustained Presentation of Bioactive Interferon-γ Within Three-Dimensional Mesenchymal Stem Cell Constructs. Stem Cells Transl. Med. 2017, 6, 223–237. [Google Scholar] [CrossRef]
- Fan, H.; Zhao, G.; Liu, L.; Liu, F.; Gong, W.; Liu, X.; Yang, L.; Wang, J.; Hou, Y. Pre-Treatment with IL-1β Enhances the Efficacy of MSC Transplantation in DSS-Induced Colitis. Cell. Mol. Immunol. 2012, 9, 473–481. [Google Scholar] [CrossRef]
- Bétous, R.; Renoud, M.-L.; Hoede, C.; Gonzalez, I.; Jones, N.; Longy, M.; Sensebé, L.; Cazaux, C.; Hoffmann, J.-S. Human Adipose-Derived Stem Cells Expanded Under Ambient Oxygen Concentration Accumulate Oxidative DNA Lesions and Experience Procarcinogenic DNA Replication Stress. Stem Cells Transl. Med. 2017, 6, 68–76. [Google Scholar] [CrossRef]
- Estrada, J.C.; Albo, C.; Benguría, A.; Dopazo, A.; López-Romero, P.; Carrera-Quintanar, L.; Roche, E.; Clemente, E.P.; Enríquez, J.A.; Bernad, A.; et al. Culture of Human Mesenchymal Stem Cells at Low Oxygen Tension Improves Growth and Genetic Stability by Activating Glycolysis. Cell Death Differ. 2012, 19, 743–755. [Google Scholar] [CrossRef]
- Lee, S.-G.; Joe, Y.A. Autophagy Mediates Enhancement of Proangiogenic Activity by Hypoxia in Mesenchymal Stromal/Stem Cells. Biochem. Biophys. Res. Commun. 2018, 501, 941–947. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Yang, Q.; Wang, Z.; Tong, H.; Ma, L.; Zhang, Y.; Shan, F.; Meng, Y.; Yuan, Z. Comparative Analysis of Human Mesenchymal Stem Cells from Fetal-Bone Marrow, Adipose Tissue, and Warton’s Jelly as Sources of Cell Immunomodulatory Therapy. Hum. Vaccines Immunother. 2016, 12, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, M.; Toupet, K.; Maumus, M.; Rozier, P.; Jorgensen, C.; Noël, D. TGFBI Secreted by Mesenchymal Stromal Cells Ameliorates Osteoarthritis and Is Detected in Extracellular Vesicles. Biomaterials 2020, 226, 119544. [Google Scholar] [CrossRef]
- Uzieliene, I.; Bernotas, P.; Mobasheri, A.; Bernotiene, E. The Role of Physical Stimuli on Calcium Channels in Chondrogenic Differentiation of Mesenchymal Stem Cells. Int. J. Mol. Sci. 2018, 19, 2998. [Google Scholar] [CrossRef]
- Parate, D.; Franco-Obregón, A.; Fröhlich, J.; Beyer, C.; Abbas, A.A.; Kamarul, T.; Hui, J.H.P.; Yang, Z. Enhancement of Mesenchymal Stem Cell Chondrogenesis with Short-Term Low Intensity Pulsed Electromagnetic Fields. Sci. Rep. 2017, 7, 9421. [Google Scholar] [CrossRef]
- Steward, A.J.; Kelly, D.J. Mechanical Regulation of Mesenchymal Stem Cell Differentiation. J. Anat. 2015, 227, 717–731. [Google Scholar] [CrossRef]
- Mueller, A.-L.; Payandeh, Z.; Mohammadkhani, N.; Mubarak, S.M.H.; Zakeri, A.; Alagheband Bahrami, A.; Brockmueller, A.; Shakibaei, M. Recent Advances in Understanding the Pathogenesis of Rheumatoid Arthritis: New Treatment Strategies. Cells 2021, 10, 3017. [Google Scholar] [CrossRef]
- Lee, D.H.; Kim, S.J.; Kim, S.A.; Ju, G.-I. Past, Present, and Future of Cartilage Restoration: From Localized Defect to Arthritis. Knee Surg. Relat. Res. 2022, 34, 1. [Google Scholar] [CrossRef]
- Deng, Y.; Lu, J.; Li, W.; Wu, A.; Zhang, X.; Tong, W.; Ho, K.K.; Qin, L.; Song, H.; Mak, K.K. Reciprocal Inhibition of YAP/TAZ and NF-κB Regulates Osteoarthritic Cartilage Degradation. Nat. Commun. 2018, 9, 4564. [Google Scholar] [CrossRef]
- Mehana, E.-S.E.; Khafaga, A.F.; El-Blehi, S.S. The Role of Matrix Metalloproteinases in Osteoarthritis Pathogenesis: An Updated Review. Life Sci. 2019, 234, 116786. [Google Scholar] [CrossRef]
- Chen, Y.; Cheng, R.-J.; Wu, Y.; Huang, D.; Li, Y.; Liu, Y. Advances in Stem Cell-Based Therapies in the Treatment of Osteoarthritis. Int. J. Mol. Sci. 2023, 25, 394. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Xiang, E.; Rao, W.; Zhang, Y.Q.; Xiao, C.H.; Li, C.Y.; Han, B.; Wu, D. Intra-Articular Injection of Human Umbilical Cord Mesenchymal Stem Cells Ameliorates Monosodium Iodoacetate-Induced Osteoarthritis in Rats by Inhibiting Cartilage Degradation and Inflammation. Bone Jt. Res. 2021, 10, 226–236. [Google Scholar] [CrossRef]
- Wang, A.-T.; Zhang, Q.-F.; Wang, N.-X.; Yu, C.-Y.; Liu, R.-M.; Luo, Y.; Zhao, Y.-J.; Xiao, J.-H. Cocktail of Hyaluronic Acid and Human Amniotic Mesenchymal Cells Effectively Repairs Cartilage Injuries in Sodium Iodoacetate-Induced Osteoarthritis Rats. Front. Bioeng. Biotechnol. 2020, 8, 87. [Google Scholar] [CrossRef]
- Guan, M.; Yu, Q.; Zhou, G.; Wang, Y.; Yu, J.; Yang, W.; Li, Z. Mechanisms of Chondrocyte Cell Death in Osteoarthritis: Implications for Disease Progression and Treatment. J. Orthop. Surg. 2024, 19, 550. [Google Scholar] [CrossRef]
- Dickinson, S.C.; Sutton, C.A.; Brady, K.; Salerno, A.; Katopodi, T.; Williams, R.L.; West, C.C.; Evseenko, D.; Wu, L.; Pang, S.; et al. The Wnt5a Receptor, Receptor Tyrosine Kinase-Like Orphan Receptor 2, Is a Predictive Cell Surface Marker of Human Mesenchymal Stem Cells with an Enhanced Capacity for Chondrogenic Differentiation. Stem Cells 2017, 35, 2280–2291. [Google Scholar] [CrossRef]
- Wang, H.; Yan, X.; Jiang, Y.; Wang, Z.; Li, Y.; Shao, Q. The Human Umbilical Cord Stem Cells Improve the Viability of OA Degenerated Chondrocytes. Mol. Med. Rep. 2018, 17, 4474–4482. [Google Scholar] [CrossRef]
- Jeon, H.-J.; Yoon, K.-A.; An, E.S.; Kang, T.-W.; Sim, Y.-B.; Ahn, J.; Choi, E.-K.; Lee, S.; Seo, K.-W.; Kim, Y.-B.; et al. Therapeutic Effects of Human Umbilical Cord Blood-Derived Mesenchymal Stem Cells Combined with Cartilage Acellular Matrix Mediated via Bone Morphogenic Protein 6 in a Rabbit Model of Articular Cruciate Ligament Transection. Stem Cell Rev. Rep. 2020, 16, 596–611. [Google Scholar] [CrossRef]
- Lin, S.; Lee, W.Y.W.; Xu, L.; Wang, Y.; Chen, Y.; Ho, K.K.W.; Qin, L.; Jiang, X.; Cui, L.; Li, G. Stepwise Preconditioning Enhances Mesenchymal Stem Cell-Based Cartilage Regeneration through Epigenetic Modification. Osteoarthr. Cartil. 2017, 25, 1541–1550. [Google Scholar] [CrossRef]
- Bolduc, J.A.; Collins, J.A.; Loeser, R.F. Reactive Oxygen Species, Aging and Articular Cartilage Homeostasis. Free Radic. Biol. Med. 2019, 132, 73–82. [Google Scholar] [CrossRef]
- Rousset, F.; Hazane-Puch, F.; Pinosa, C.; Nguyen, M.V.C.; Grange, L.; Soldini, A.; Rubens-Duval, B.; Dupuy, C.; Morel, F.; Lardy, B. IL-1beta Mediates MMP Secretion and IL-1beta Neosynthesis via Upregulation of P22phox and NOX4 Activity in Human Articular Chondrocytes. Osteoarthr. Cartil. 2015, 23, 1972–1980. [Google Scholar] [CrossRef]
- Platas, J.; Guillén, M.I.; del Caz, M.D.P.; Gomar, F.; Castejón, M.A.; Mirabet, V.; Alcaraz, M.J. Paracrine Effects of Human Adipose-Derived Mesenchymal Stem Cells in Inflammatory Stress-Induced Senescence Features of Osteoarthritic Chondrocytes. Aging 2016, 8, 1703–1717. [Google Scholar] [CrossRef] [PubMed]
- Coryell, P.R.; Diekman, B.O.; Loeser, R.F. Mechanisms and Therapeutic Implications of Cellular Senescence in Osteoarthritis. Nat. Rev. Rheumatol. 2021, 17, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Maimaitijuma, T.; Ma, Y.-Y.; Jiao, Y.; Cao, Y.-P. Mitochondrial Transfer from Bone-Marrow-Derived Mesenchymal Stromal Cells to Chondrocytes Protects against Cartilage Degenerative Mitochondrial Dysfunction in Rats Chondrocytes. Chin. Med. J. 2021, 134, 212. [Google Scholar] [CrossRef]
- Souza, A.G.; Silva, I.B.B.; Campos-Fernandez, E.; Barcelos, L.S.; Souza, J.B.; Marangoni, K.; Goulart, L.R.; Alonso-Goulart, V. Comparative Assay of 2D and 3D Cell Culture Models: Proliferation, Gene Expression and Anticancer Drug Response. Curr. Pharm. Des. 2018, 24, 1689–1694. [Google Scholar] [CrossRef]
- Bartosh, T.J.; Ylöstalo, J.H.; Mohammadipoor, A.; Bazhanov, N.; Coble, K.; Claypool, K.; Lee, R.H.; Choi, H.; Prockop, D.J. Aggregation of Human Mesenchymal Stromal Cells (MSCs) into 3D Spheroids Enhances Their Antiinflammatory Properties. Proc. Natl. Acad. Sci. USA 2010, 107, 13724–13729. [Google Scholar] [CrossRef]
- Wong, C.-C.; Sheu, S.-D.; Chung, P.-C.; Yeh, Y.-Y.; Chen, C.-H.; Chang, Y.-W.; Kuo, T.-F. Hyaluronic Acid Supplement as a Chondrogenic Adjuvant in Promoting the Therapeutic Efficacy of Stem Cell Therapy in Cartilage Healing. Pharmaceutics 2021, 13, 432. [Google Scholar] [CrossRef]
- Joyce, K.; Fabra, G.T.; Bozkurt, Y.; Pandit, A. Bioactive Potential of Natural Biomaterials: Identification, Retention and Assessment of Biological Properties. Signal Transduct. Target. Ther. 2021, 6, 122. [Google Scholar] [CrossRef]
- Zhan, X. Effect of Matrix Stiffness and Adhesion Ligand Density on Chondrogenic Differentiation of Mesenchymal Stem Cells. J. Biomed. Mater. Res. Part A 2019, 108, 675–683. [Google Scholar] [CrossRef]
- Shao, J.; Ding, Z.; Li, L.; Chen, Y.; Zhu, J.; Qian, Q. Improved Accumulation of TGF-β by Photopolymerized Chitosan/Silk Protein Bio-Hydrogel Matrix to Improve Differentiations of Mesenchymal Stem Cells in Articular Cartilage Tissue Regeneration. J. Photochem. Photobiol. B 2020, 203, 111744. [Google Scholar] [CrossRef]
- Baldari, S.; Di Rocco, G.; Piccoli, M.; Pozzobon, M.; Muraca, M.; Toietta, G. Challenges and Strategies for Improving the Regenerative Effects of Mesenchymal Stromal Cell-Based Therapies. Int. J. Mol. Sci. 2017, 18, 2087. [Google Scholar] [CrossRef]
- Park, Y.-B.; Ha, C.-W.; Lee, C.-H.; Yoon, Y.C.; Park, Y.-G. Cartilage Regeneration in Osteoarthritic Patients by a Composite of Allogeneic Umbilical Cord Blood-Derived Mesenchymal Stem Cells and Hyaluronate Hydrogel: Results from a Clinical Trial for Safety and Proof-of-Concept with 7 Years of Extended Follow-Up. Stem Cells Transl. Med. 2017, 6, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Fernandez, P.; Simula, L.; Jenni, S.; Jordan, O.; Allémann, E. Hyaluronan-Based Hydrogel Delivering Glucose to Mesenchymal Stem Cells Intended to Treat Osteoarthritis. Int. J. Pharm. 2024, 657, 124139. [Google Scholar] [CrossRef]
- Volarevic, V.; Markovic, B.S.; Gazdic, M.; Volarevic, A.; Jovicic, N.; Arsenijevic, N.; Armstrong, L.; Djonov, V.; Lako, M.; Stojkovic, M. Ethical and Safety Issues of Stem Cell-Based Therapy. Int. J. Med. Sci. 2018, 15, 36–45. [Google Scholar] [CrossRef]
- Ljujic, B.; Milovanovic, M.; Volarevic, V.; Murray, B.; Bugarski, D.; Przyborski, S.; Arsenijevic, N.; Lukic, M.L.; Stojkovic, M. Human Mesenchymal Stem Cells Creating an Immunosuppressive Environment and Promote Breast Cancer in Mice. Sci. Rep. 2013, 3, 2298. [Google Scholar] [CrossRef]
- Gazdic, M.; Volarevic, V.; Arsenijevic, N.; Stojkovic, M. Mesenchymal Stem Cells: A Friend or Foe in Immune-Mediated Diseases. Stem Cell Rev. Rep. 2015, 11, 280–287. [Google Scholar] [CrossRef]
- Zhu, X.; Ma, D.; Yang, B.; An, Q.; Zhao, J.; Gao, X.; Zhang, L. Research Progress of Engineered Mesenchymal Stem Cells and Their Derived Exosomes and Their Application in Autoimmune/Inflammatory Diseases. Stem Cell Res. Ther. 2023, 14, 71. [Google Scholar] [CrossRef]
- Musiał-Wysocka, A.; Kot, M.; Majka, M. The Pros and Cons of Mesenchymal Stem Cell-Based Therapies. Cell Transplant. 2019, 28, 801–812. [Google Scholar] [CrossRef]
- Deng, Y.; Zhang, Y.; Ye, L.; Zhang, T.; Cheng, J.; Chen, G.; Zhang, Q.; Yang, Y. Umbilical Cord-Derived Mesenchymal Stem Cells Instruct Monocytes Towards an IL10-Producing Phenotype by Secreting IL6 and HGF. Sci. Rep. 2016, 6, 37566. [Google Scholar] [CrossRef]
- Davies, L.C.; Heldring, N.; Kadri, N.; Le Blanc, K. Mesenchymal Stromal Cell Secretion of Programmed Death-1 Ligands Regulates T Cell Mediated Immunosuppression. Stem Cells 2017, 35, 766–776. [Google Scholar] [CrossRef]
- Lai, J.J.; Chau, Z.L.; Chen, S.-Y.; Hill, J.J.; Korpany, K.V.; Liang, N.-W.; Lin, L.-H.; Lin, Y.-H.; Liu, J.K.; Liu, Y.-C.; et al. Exosome Processing and Characterization Approaches for Research and Technology Development. Adv. Sci. 2022, 9, e2103222. [Google Scholar] [CrossRef]
- Smirnova, O.; Efremov, Y.; Klyucherev, T.; Peshkova, M.; Senkovenko, A.; Svistunov, A.; Timashev, P. Direct and Cell-Mediated EV-ECM Interplay. Acta Biomater. 2024, 186, 63–84. [Google Scholar] [CrossRef]
- Jeppesen, D.K.; Zhang, Q.; Franklin, J.L.; Coffey, R.J. Extracellular Vesicles and Nanoparticles: Emerging Complexities. Trends Cell Biol. 2023, 33, 667–681. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V.S. The Biology, Function, and Biomedical Applications of Exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef]
- Welsh, J.A.; Goberdhan, D.C.I.; O’Driscoll, L.; Buzas, E.I.; Blenkiron, C.; Bussolati, B.; Cai, H.; Di Vizio, D.; Driedonks, T.A.P.; Erdbrügger, U.; et al. Minimal Information for Studies of Extracellular Vesicles (MISEV2023): From Basic to Advanced Approaches. J. Extracell. Vesicles 2024, 13, e12404. [Google Scholar] [CrossRef]
- Willms, E.; Cabañas, C.; Mäger, I.; Wood, M.J.A.; Vader, P. Extracellular Vesicle Heterogeneity: Subpopulations, Isolation Techniques, and Diverse Functions in Cancer Progression. Front. Immunol. 2018, 9, 738. [Google Scholar] [CrossRef]
- Hessvik, N.P.; Llorente, A. Current Knowledge on Exosome Biogenesis and Release. Cell. Mol. Life Sci. CMLS 2018, 75, 193–208. [Google Scholar] [CrossRef]
- Zhao, R.; Zhao, T.; He, Z.; Cai, R.; Pang, W. Composition, Isolation, Identification and Function of Adipose Tissue-Derived Exosomes. Adipocyte 2021, 10, 587–604. [Google Scholar] [CrossRef]
- Yáñez-Mó, M.; Siljander, P.R.-M.; Andreu, Z.; Zavec, A.B.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological Properties of Extracellular Vesicles and Their Physiological Functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef]
- Caruso, S.; Poon, I.K.H. Apoptotic Cell-Derived Extracellular Vesicles: More Than Just Debris. Front. Immunol. 2018, 9, 1486. [Google Scholar] [CrossRef]
- Dostert, G.; Mesure, B.; Menu, P.; Velot, É. How Do Mesenchymal Stem Cells Influence or Are Influenced by Microenvironment through Extracellular Vesicles Communication? Front. Cell Dev. Biol. 2017, 5, 6. [Google Scholar] [CrossRef]
- Wang, L.; Wang, C.; Jia, X.; Yu, J. Circulating Exosomal miR-17 Inhibits the Induction of Regulatory T Cells via Suppressing TGFBR II Expression in Rheumatoid Arthritis. Cell. Physiol. Biochem. 2018, 50, 1754–1763. [Google Scholar] [CrossRef]
- Yan, J.; Shen, M.; Sui, B.; Lu, W.; Han, X.; Wan, Q.; Liu, Y.; Kang, J.; Qin, W.; Zhang, Z.; et al. Autophagic LC3+ Calcified Extracellular Vesicles Initiate Cartilage Calcification in Osteoarthritis. Sci. Adv. 2022, 8, eabn1556. [Google Scholar] [CrossRef]
- Zeng, G.; Deng, G.; Xiao, S.; Li, F. Fibroblast-like Synoviocytes-Derived Exosomal PCGEM1 Accelerates IL-1β-Induced Apoptosis and Cartilage Matrix Degradation by miR-142-5p/RUNX2 in Chondrocytes. Immunol. Investig. 2022, 51, 1284–1301. [Google Scholar] [CrossRef]
- Kolhe, R.; Hunter, M.; Liu, S.; Jadeja, R.N.; Pundkar, C.; Mondal, A.K.; Mendhe, B.; Drewry, M.; Rojiani, M.V.; Liu, Y.; et al. Gender-Specific Differential Expression of Exosomal miRNA in Synovial Fluid of Patients with Osteoarthritis. Sci. Rep. 2017, 7, 2029. [Google Scholar] [CrossRef]
- Michael, B.N.R.; Kommoju, V.; Kavadichanda Ganapathy, C.; Negi, V.S. Characterization of Cell-Derived Microparticles in Synovial Fluid and Plasma of Patients with Rheumatoid Arthritis. Rheumatol. Int. 2019, 39, 1377–1387. [Google Scholar] [CrossRef]
- Jouybari, M.T.; Mojtahedi, F.; Babaahmadi, M.; Faeed, M.; Eslaminejad, M.B.; Taghiyar, L. Advancements in Extracellular Vesicle Targeted Therapies for Rheumatoid Arthritis: Insights into Cellular Origins, Current Perspectives, and Emerging Challenges. Stem Cell Res. Ther. 2024, 15, 276. [Google Scholar] [CrossRef]
- Jeon, O.H.; Wilson, D.R.; Clement, C.C.; Rathod, S.; Cherry, C.; Powell, B.; Lee, Z.; Khalil, A.M.; Green, J.J.; Campisi, J.; et al. Senescence Cell–Associated Extracellular Vesicles Serve as Osteoarthritis Disease and Therapeutic Markers. JCI Insight 2019, 4, e125019. [Google Scholar] [CrossRef]
- Wu, X.; Crawford, R.; Xiao, Y.; Mao, X.; Prasadam, I. Osteoarthritic Subchondral Bone Release Exosomes That Promote Cartilage Degeneration. Cells 2021, 10, 251. [Google Scholar] [CrossRef]
- Arntz, O.J.; Pieters, B.C.H.; Thurlings, R.M.; Wenink, M.H.; van Lent, P.L.E.M.; Koenders, M.I.; van den Hoogen, F.H.J.; van der Kraan, P.M.; van de Loo, F.A.J. Rheumatoid Arthritis Patients with Circulating Extracellular Vesicles Positive for IgM Rheumatoid Factor Have Higher Disease Activity. Front. Immunol. 2018, 9, 2388. [Google Scholar] [CrossRef]
- Burbano, C.; Villar-Vesga, J.; Vásquez, G.; Muñoz-Vahos, C.; Rojas, M.; Castaño, D. Proinflammatory Differentiation of Macrophages Through Microparticles That Form Immune Complexes Leads to T- and B-Cell Activation in Systemic Autoimmune Diseases. Front. Immunol. 2019, 10, 2058. [Google Scholar] [CrossRef]
- Song, J.E.; Kim, J.S.; Shin, J.H.; Moon, K.W.; Park, J.K.; Park, K.S.; Lee, E.Y. Role of Synovial Exosomes in Osteoclast Differentiation in Inflammatory Arthritis. Cells 2021, 10, 120. [Google Scholar] [CrossRef]
- Neumann, E.; Lefèvre, S.; Zimmermann, B.; Gay, S.; Müller-Ladner, U. Rheumatoid Arthritis Progression Mediated by Activated Synovial Fibroblasts. Trends Mol. Med. 2010, 16, 458–468. [Google Scholar] [CrossRef]
- Frank-Bertoncelj, M.; Pisetsky, D.S.; Kolling, C.; Michel, B.A.; Gay, R.E.; Jüngel, A.; Gay, S. TLR3 Ligand Poly(I:C) Exerts Distinct Actions in Synovial Fibroblasts When Delivered by Extracellular Vesicles. Front. Immunol. 2018, 9, 28. [Google Scholar] [CrossRef]
- Cavallo, C.; Merli, G.; Borzì, R.M.; Zini, N.; D’Adamo, S.; Guescini, M.; Grigolo, B.; Di Martino, A.; Santi, S.; Filardo, G. Small Extracellular Vesicles from Adipose Derived Stromal Cells Significantly Attenuate in Vitro the NF-κB Dependent Inflammatory/Catabolic Environment of Osteoarthritis. Sci. Rep. 2021, 11, 1053. [Google Scholar] [CrossRef]
- Lu, L.; Wang, J.; Fan, A.; Wang, P.; Chen, R.; Lu, L.; Yin, F. Synovial Mesenchymal Stem Cell-Derived Extracellular Vesicles Containing microRN555A-26a-5p Ameliorate Cartilage Damage of Osteoarthritis. J. Gene Med. 2021, 23, e3379. [Google Scholar] [CrossRef]
- Tian, X.; Wei, W.; Cao, Y.; Ao, T.; Huang, F.; Javed, R.; Wang, X.; Fan, J.; Zhang, Y.; Liu, Y.; et al. Gingival Mesenchymal Stem Cell-Derived Exosomes Are Immunosuppressive in Preventing Collagen-Induced Arthritis. J. Cell. Mol. Med. 2022, 26, 693–708. [Google Scholar] [CrossRef]
- Wu, X.; Zhu, D.; Tian, J.; Tang, X.; Guo, H.; Ma, J.; Xu, H.; Wang, S. Granulocytic Myeloid-Derived Suppressor Cell Exosomal Prostaglandin E2 Ameliorates Collagen-Induced Arthritis by Enhancing IL-10+ B Cells. Front. Immunol. 2020, 11, 588500. [Google Scholar] [CrossRef]
- Cosenza, S.; Ruiz, M.; Toupet, K.; Jorgensen, C.; Noël, D. Mesenchymal Stem Cells Derived Exosomes and Microparticles Protect Cartilage and Bone from Degradation in Osteoarthritis. Sci. Rep. 2017, 7, 16214. [Google Scholar] [CrossRef]
- Varderidou-Minasian, S.; Lorenowicz, M.J. Mesenchymal Stromal/Stem Cell-Derived Extracellular Vesicles in Tissue Repair: Challenges and Opportunities. Theranostics 2020, 10, 5979–5997. [Google Scholar] [CrossRef]
- Kim, G.B.; Shon, O.-J.; Seo, M.-S.; Choi, Y.; Park, W.T.; Lee, G.W. Mesenchymal Stem Cell-Derived Exosomes and Their Therapeutic Potential for Osteoarthritis. Biology 2021, 10, 285. [Google Scholar] [CrossRef]
- Casado-Díaz, A.; Quesada-Gómez, J.M.; Dorado, G. Extracellular Vesicles Derived From Mesenchymal Stem Cells (MSC) in Regenerative Medicine: Applications in Skin Wound Healing. Front. Bioeng. Biotechnol. 2020, 8, 146. [Google Scholar] [CrossRef]
- Elahi, F.M.; Farwell, D.G.; Nolta, J.A.; Anderson, J.D. Preclinical Translation of Exosomes Derived from Mesenchymal Stem/Stromal Cells. Stem Cells 2020, 38, 15–21. [Google Scholar] [CrossRef]
- Sun, L.; Xu, R.; Sun, X.; Duan, Y.; Han, Y.; Zhao, Y.; Qian, H.; Zhu, W.; Xu, W. Safety Evaluation of Exosomes Derived from Human Umbilical Cord Mesenchymal Stromal Cell. Cytotherapy 2016, 18, 413–422. [Google Scholar] [CrossRef]
- Chang, T.-H.; Wu, C.-S.; Chiou, S.-H.; Chang, C.-H.; Liao, H.-J. Adipose-Derived Stem Cell Exosomes as a Novel Anti-Inflammatory Agent and the Current Therapeutic Targets for Rheumatoid Arthritis. Biomedicines 2022, 10, 1725. [Google Scholar] [CrossRef]
- Alvarez-Erviti, L.; Seow, Y.; Yin, H.; Betts, C.; Lakhal, S.; Wood, M.J.A. Delivery of siRNA to the Mouse Brain by Systemic Injection of Targeted Exosomes. Nat. Biotechnol. 2011, 29, 341–345. [Google Scholar] [CrossRef]
- Zhang, J.; Li, S.; Li, L.; Li, M.; Guo, C.; Yao, J.; Mi, S. Exosome and Exosomal microRNA: Trafficking, Sorting, and Function. Genom. Proteom. Bioinform. 2015, 13, 17–24. [Google Scholar] [CrossRef]
- Vader, P.; Mol, E.A.; Pasterkamp, G.; Schiffelers, R.M. Extracellular Vesicles for Drug Delivery. Adv. Drug Deliv. Rev. 2016, 106, 148–156. [Google Scholar] [CrossRef]
- Lee, E.S.; Ko, H.; Kim, C.H.; Kim, H.-C.; Choi, S.-K.; Jeong, S.W.; Lee, S.-G.; Lee, S.-J.; Na, H.-K.; Park, J.H.; et al. Disease-Microenvironment Modulation by Bare- or Engineered-Exosome for Rheumatoid Arthritis Treatment. Biomater. Res. 2023, 27, 81. [Google Scholar] [CrossRef]
- Mellado, M.; Martínez-Muñoz, L.; Cascio, G.; Lucas, P.; Pablos, J.L.; Rodríguez-Frade, J.M. T Cell Migration in Rheumatoid Arthritis. Front. Immunol. 2015, 6, 384. [Google Scholar] [CrossRef]
- Lotfi, N.; Thome, R.; Rezaei, N.; Zhang, G.-X.; Rezaei, A.; Rostami, A.; Esmaeil, N. Roles of GM-CSF in the Pathogenesis of Autoimmune Diseases: An Update. Front. Immunol. 2019, 10, 1265. [Google Scholar] [CrossRef]
- Wang, B.; Zhao, P.; Zhou, Y.; Meng, L.; Zhu, W.; Jiang, C.; Wang, L.; Cai, Y.; Lu, S.; Hou, W. Increased Expression of Th17 Cytokines and Interleukin-22 Correlates with Disease Activity in Pristane-Induced Arthritis in Rats. PLoS ONE 2017, 12, e0188199. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Feng, X.; Gao, W. Proliferation of Rheumatoid Arthritis Fibroblast-like Synoviocytes Is Enhanced by IL-17-Mediated Autophagy through STAT3 Activation. Connect. Tissue Res. 2019, 60, 358–366. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Qian, W.; Qian, Y.; Giltiay, N.V.; Lu, Y.; Swaidani, S.; Misra, S.; Deng, L.; Chen, Z.J.; Li, X. Act1, a U-Box E3 Ubiquitin Ligase for IL-17 Signaling. Sci. Signal. 2009, 2, ra63. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Xu, K.; Zhang, G.; Liu, Y.; Gao, J.; Tian, M.; Wei, C.; Li, J.; Zhang, L. Immunomodulatory Effect of Human Umbilical Cord Mesenchymal Stem Cells on T Lymphocytes in Rheumatoid Arthritis. Int. Immunopharmacol. 2019, 74, 105687. [Google Scholar] [CrossRef]
- Cosenza, S.; Toupet, K.; Maumus, M.; Luz-Crawford, P.; Blanc-Brude, O.; Jorgensen, C.; Noël, D. Mesenchymal Stem Cells-Derived Exosomes Are More Immunosuppressive than Microparticles in Inflammatory Arthritis. Theranostics 2018, 8, 1399–1410. [Google Scholar] [CrossRef]
- Komatsu, N.; Takayanagi, H. Mechanisms of Joint Destruction in Rheumatoid Arthritis—Immune Cell-Fibroblast-Bone Interactions. Nat. Rev. Rheumatol. 2022, 18, 415–429. [Google Scholar] [CrossRef]
- Koedderitzsch, K.; Zezina, E.; Li, L.; Herrmann, M.; Biesemann, N. TNF Induces Glycolytic Shift in Fibroblast like Synoviocytes via GLUT1 and HIF1A. Sci. Rep. 2021, 11, 19385. [Google Scholar] [CrossRef]
- Zhang, B.; Gu, J.; Wang, Y.; Guo, L.; Xie, J.; Yang, M. TNF-α Stimulated Exosome Derived from Fibroblast-like Synoviocytes Isolated from Rheumatoid Arthritis Patients Promotes HUVEC Migration, Invasion and Angiogenesis by Targeting the miR-200a-3p/KLF6/VEGFA Axis. Autoimmunity 2023, 56, 2282939. [Google Scholar] [CrossRef]
- Yoshitomi, H. Regulation of Immune Responses and Chronic Inflammation by Fibroblast-Like Synoviocytes. Front. Immunol. 2019, 10, 1395. [Google Scholar] [CrossRef]
- del Rey, M.J.; Izquierdo, E.; Caja, S.; Usategui, A.; Santiago, B.; Galindo, M.; Pablos, J.L. Human Inflammatory Synovial Fibroblasts Induce Enhanced Myeloid Cell Recruitment and Angiogenesis through a Hypoxia-Inducible Transcription Factor 1alpha/Vascular Endothelial Growth Factor-Mediated Pathway in Immunodeficient Mice. Arthritis Rheum. 2009, 60, 2926–2934. [Google Scholar] [CrossRef]
- Machado, C.R.L.; Dias, F.F.; Resende, G.G.; de Oliveira, P.G.; Xavier, R.M.; de Andrade, M.V.M.; Kakehasi, A.M. Morphofunctional Analysis of Fibroblast-like Synoviocytes in Human Rheumatoid Arthritis and Mouse Collagen-Induced Arthritis. Adv. Rheumatol. 2023, 63, 1. [Google Scholar] [CrossRef] [PubMed]
- Machado, C.R.L.; Resende, G.G.; Macedo, R.B.V.; do Nascimento, V.C.; Branco, A.S.; Kakehasi, A.M.; Andrade, M.V. Fibroblast-like Synoviocytes from Fluid and Synovial Membrane from Primary Osteoarthritis Demonstrate Similar Production of Interleukin 6, and Metalloproteinases 1 and 3. Clin. Exp. Rheumatol. 2019, 37, 306–309. [Google Scholar] [PubMed]
- Ma, W.; Tang, F.; Xiao, L.; Han, S.; Yao, X.; Zhang, Q.; Zhou, J.; Wang, Y.; Zhou, J. miR-205-5p in Exosomes Divided from Chondrogenic Mesenchymal Stem Cells Alleviated Rheumatoid Arthritis via Regulating MDM2 in Fibroblast-like Synoviocytes. J. Musculoskelet. Neuronal Interact. 2022, 22, 132–141. [Google Scholar] [PubMed]
- Xu, W.; Liu, X.; Qu, W.; Wang, X.; Su, H.; Li, W.; Cheng, Y. Exosomes Derived from Fibrinogen-like Protein 1-Overexpressing Bone Marrow-Derived Mesenchymal Stem Cells Ameliorates Rheumatoid Arthritis. Bioengineered 2022, 13, 14545–14561. [Google Scholar] [CrossRef]
- Ostojic, M.; Zevrnja, A.; Vukojevic, K.; Soljic, V. Immunofluorescence Analysis of NF-kB and iNOS Expression in Different Cell Populations during Early and Advanced Knee Osteoarthritis. Int. J. Mol. Sci. 2021, 22, 6461. [Google Scholar] [CrossRef]
- Scanzello, C.R. Role of Low-Grade Inflammation in Osteoarthritis. Curr. Opin. Rheumatol. 2017, 29, 79–85. [Google Scholar] [CrossRef]
- Teo, K.Y.W.; Zhang, S.; Loh, J.T.; Lai, R.C.; Hey, H.W.D.; Lam, K.-P.; Lim, S.K.; Toh, W.S. Mesenchymal Stromal Cell Exosomes Mediate M2-like Macrophage Polarization through CD73/Ecto-5′-Nucleotidase Activity. Pharmaceutics 2023, 15, 1489. [Google Scholar] [CrossRef]
- Lepetsos, P.; Papavassiliou, A.G. ROS/Oxidative Stress Signaling in Osteoarthritis. Biochim. Biophys. Acta 2016, 1862, 576–591. [Google Scholar] [CrossRef]
- Li, X.; Wang, Y.; Cai, Z.; Zhou, Q.; Li, L.; Fu, P. Exosomes from Human Umbilical Cord Mesenchymal Stem Cells Inhibit ROS Production and Cell Apoptosis in Human Articular Chondrocytes via the miR-100-5p/NOX4 Axis. Cell Biol. Int. 2021, 45, 2096–2106. [Google Scholar] [CrossRef]
- Zhou, H.; Shen, X.; Yan, C.; Xiong, W.; Ma, Z.; Tan, Z.; Wang, J.; Li, Y.; Liu, J.; Duan, A.; et al. Extracellular Vesicles Derived from Human Umbilical Cord Mesenchymal Stem Cells Alleviate Osteoarthritis of the Knee in Mice Model by Interacting with METTL3 to Reduce m6A of NLRP3 in Macrophage. Stem Cell Res. Ther. 2022, 13, 322. [Google Scholar] [CrossRef]
- Huleihel, L.; Hussey, G.S.; Naranjo, J.D.; Zhang, L.; Dziki, J.L.; Turner, N.J.; Stolz, D.B.; Badylak, S.F. Matrix-Bound Nanovesicles within ECM Bioscaffolds. Sci. Adv. 2016, 2, e1600502. [Google Scholar] [CrossRef] [PubMed]
- Peshkova, M.; Korneev, A.; Revokatova, D.; Smirnova, O.; Klyucherev, T.; Shender, V.; Arapidi, G.; Kosheleva, N.; Timashev, P. Four Sides to the Story: A Proteomic Comparison of Liquid-Phase and Matrix-Bound Extracellular Vesicles in 2D and 3D Cell Cultures. Proteomics 2024, 24, e2300375. [Google Scholar] [CrossRef] [PubMed]
- Huleihel, L.; Bartolacci, J.G.; Dziki, J.L.; Vorobyov, T.; Arnold, B.; Scarritt, M.E.; Pineda Molina, C.; LoPresti, S.T.; Brown, B.N.; Naranjo, J.D.; et al. Matrix-Bound Nanovesicles Recapitulate Extracellular Matrix Effects on Macrophage Phenotype. Tissue Eng. Part A 2017, 23, 1283–1294. [Google Scholar] [CrossRef] [PubMed]
- Crum, R.J.; Hall, K.; Molina, C.P.; Hussey, G.S.; Graham, E.; Li, H.; Badylak, S.F. Immunomodulatory Matrix-Bound Nanovesicles Mitigate Acute and Chronic Pristane-Induced Rheumatoid Arthritis. NPJ Regen. Med. 2022, 7, 13. [Google Scholar] [CrossRef]
- Kim, H.; Back, J.H.; Han, G.; Lee, S.J.; Park, Y.E.; Gu, M.B.; Yang, Y.; Lee, J.E.; Kim, S.H. Extracellular Vesicle-Guided in Situ Reprogramming of Synovial Macrophages for the Treatment of Rheumatoid Arthritis. Biomaterials 2022, 286, 121578. [Google Scholar] [CrossRef]
- Zhu, D.; Tian, J.; Wu, X.; Li, M.; Tang, X.; Rui, K.; Guo, H.; Ma, J.; Xu, H.; Wang, S. G-MDSC-Derived Exosomes Attenuate Collagen-Induced Arthritis by Impairing Th1 and Th17 Cell Responses. Biochim. Biophys. Acta Mol. Basis Dis. 2019, 1865, 165540. [Google Scholar] [CrossRef]
- Headland, S.E.; Jones, H.R.; Norling, L.V.; Kim, A.; Souza, P.R.; Corsiero, E.; Gil, C.D.; Nerviani, A.; Dell’Accio, F.; Pitzalis, C.; et al. Neutrophil-Derived Microvesicles Enter Cartilage and Protect the Joint in Inflammatory Arthritis. Sci. Transl. Med. 2015, 7, 315ra190. [Google Scholar] [CrossRef]
- Da-Wa, Z.X.; Jun, M.; Chao-Zheng, L.; Sen-Lin, Y.; Chuan, L.; De-Chun, L.; Zu-Nan, D.; Hong-Tao, Z.; Shu-Qing, W.; Xian-Wei, P.; et al. Exosomes Derived from M2 Macrophages Exert a Therapeutic Effect via Inhibition of the PI3K/AKT/mTOR Pathway in Rats with Knee Osteoarthritic. BioMed Res. Int. 2021, 2021, 7218067. [Google Scholar] [CrossRef]
- Bai, J.; Zhang, Y.; Zheng, X.; Huang, M.; Cheng, W.; Shan, H.; Gao, X.; Zhang, M.; Sheng, L.; Dai, J.; et al. LncRNA MM2P-Induced, Exosome-Mediated Transfer of Sox9 from Monocyte-Derived Cells Modulates Primary Chondrocytes. Cell Death Dis. 2020, 11, 763. [Google Scholar] [CrossRef]
- Qin, L.; Yang, J.; Su, X.; Li, X.; Lei, Y.; Dong, L.; Chen, H.; Chen, C.; Zhao, C.; Zhang, H.; et al. The miR-21-5p Enriched in the Apoptotic Bodies of M2 Macrophage-Derived Extracellular Vesicles Alleviates Osteoarthritis by Changing Macrophage Phenotype. Genes Dis. 2023, 10, 1114–1129. [Google Scholar] [CrossRef]
- Rhys, H.I.; Dell’Accio, F.; Pitzalis, C.; Moore, A.; Norling, L.V.; Perretti, M. Neutrophil Microvesicles from Healthy Control and Rheumatoid Arthritis Patients Prevent the Inflammatory Activation of Macrophages. eBioMedicine 2018, 29, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Feng, D.; Zhao, W.-L.; Ye, Y.-Y.; Bai, X.-C.; Liu, R.-Q.; Chang, L.-F.; Zhou, Q.; Sui, S.-F. Cellular Internalization of Exosomes Occurs through Phagocytosis. Traffic 2010, 11, 675–687. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Liang, Y.; Li, X.; Ouyang, K.; Wang, M.; Cao, T.; Li, W.; Liu, J.; Xiong, J.; Li, B.; et al. Exosome-Mediated Delivery of Kartogenin for Chondrogenesis of Synovial Fluid-Derived Mesenchymal Stem Cells and Cartilage Regeneration. Biomaterials 2021, 269, 120539. [Google Scholar] [CrossRef] [PubMed]
- You, D.G.; Lim, G.T.; Kwon, S.; Um, W.; Oh, B.H.; Song, S.H.; Lee, J.; Jo, D.-G.; Cho, Y.W.; Park, J.H. Metabolically Engineered Stem Cell-Derived Exosomes to Regulate Macrophage Heterogeneity in Rheumatoid Arthritis. Sci. Adv. 2021, 7, eabe0083. [Google Scholar] [CrossRef]
- Pan, Z.; Sun, W.; Chen, Y.; Tang, H.; Lin, W.; Chen, J.; Chen, C. Extracellular Vesicles in Tissue Engineering: Biology and Engineered Strategy. Adv. Healthc. Mater. 2022, 11, e2201384. [Google Scholar] [CrossRef]
- Lee, E.S.; Sul, J.H.; Shin, J.M.; Shin, S.; Lee, J.A.; Kim, H.K.; Cho, Y.; Ko, H.; Son, S.; Lee, J.; et al. Reactive Oxygen Species-Responsive Dendritic Cell-Derived Exosomes for Rheumatoid Arthritis. Acta Biomater. 2021, 128, 462–473. [Google Scholar] [CrossRef]
- Duan, A.; Shen, K.; Li, B.; Li, C.; Zhou, H.; Kong, R.; Shao, Y.; Qin, J.; Yuan, T.; Ji, J.; et al. Extracellular Vesicles Derived from LPS-Preconditioned Human Synovial Mesenchymal Stem Cells Inhibit Extracellular Matrix Degradation and Prevent Osteoarthritis of the Knee in a Mouse Model. Stem Cell Res. Ther. 2021, 12, 427. [Google Scholar] [CrossRef]
- Rong, Y.; Zhang, J.; Jiang, D.; Ji, C.; Liu, W.; Wang, J.; Ge, X.; Tang, P.; Yu, S.; Cui, W.; et al. Hypoxic Pretreatment of Small Extracellular Vesicles Mediates Cartilage Repair in Osteoarthritis by Delivering miR-216a-5p. Acta Biomater. 2021, 122, 325–342. [Google Scholar] [CrossRef]
- Ragni, E.; Perucca Orfei, C.; De Luca, P.; Colombini, A.; Viganò, M.; de Girolamo, L. Secreted Factors and EV-miRNAs Orchestrate the Healing Capacity of Adipose Mesenchymal Stem Cells for the Treatment of Knee Osteoarthritis. Int. J. Mol. Sci. 2020, 21, 1582. [Google Scholar] [CrossRef]
- Yan, L.; Wu, X. Exosomes Produced from 3D Cultures of Umbilical Cord Mesenchymal Stem Cells in a Hollow-Fiber Bioreactor Show Improved Osteochondral Regeneration Activity. Cell Biol. Toxicol. 2020, 36, 165. [Google Scholar] [CrossRef]
- Zhang, B.; Tian, X.; Qu, Z.; Hao, J.; Zhang, W. Hypoxia-Preconditioned Extracellular Vesicles from Mesenchymal Stem Cells Improve Cartilage Repair in Osteoarthritis. Membranes 2022, 12, 225. [Google Scholar] [CrossRef] [PubMed]
- Saller, M.M.; Prall, W.C.; Docheva, D.; Schönitzer, V.; Popov, T.; Anz, D.; Clausen-Schaumann, H.; Mutschler, W.; Volkmer, E.; Schieker, M.; et al. Increased Stemness and Migration of Human Mesenchymal Stem Cells in Hypoxia Is Associated with Altered Integrin Expression. Biochem. Biophys. Res. Commun. 2012, 423, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-King, H.; García, N.A.; Ontoria-Oviedo, I.; Ciria, M.; Montero, J.A.; Sepúlveda, P. Hypoxia Inducible Factor-1α Potentiates Jagged 1-Mediated Angiogenesis by Mesenchymal Stem Cell-Derived Exosomes. Stem Cells 2017, 35, 1747–1759. [Google Scholar] [CrossRef] [PubMed]
- Gorin, C.; Rochefort, G.Y.; Bascetin, R.; Ying, H.; Lesieur, J.; Sadoine, J.; Beckouche, N.; Berndt, S.; Novais, A.; Lesage, M.; et al. Priming Dental Pulp Stem Cells With Fibroblast Growth Factor-2 Increases Angiogenesis of Implanted Tissue-Engineered Constructs Through Hepatocyte Growth Factor and Vascular Endothelial Growth Factor Secretion. Stem Cells Transl. Med. 2016, 5, 392–404. [Google Scholar] [CrossRef]
- Rocha, S.; Carvalho, J.; Oliveira, P.; Voglstaetter, M.; Schvartz, D.; Thomsen, A.R.; Walter, N.; Khanduri, R.; Sanchez, J.-C.; Keller, A.; et al. 3D Cellular Architecture Affects MicroRNA and Protein Cargo of Extracellular Vesicles. Adv. Sci. 2019, 6, 1800948. [Google Scholar] [CrossRef]
- Haraszti, R.A.; Miller, R.; Stoppato, M.; Sere, Y.Y.; Coles, A.; Didiot, M.-C.; Wollacott, R.; Sapp, E.; Dubuke, M.L.; Li, X.; et al. Exosomes Produced from 3D Cultures of MSCs by Tangential Flow Filtration Show Higher Yield and Improved Activity. Mol. Ther. J. Am. Soc. Gene Ther. 2018, 26, 2838–2847. [Google Scholar] [CrossRef]
- Kim, H.; Lee, M.J.; Bae, E.-H.; Ryu, J.S.; Kaur, G.; Kim, H.J.; Kim, J.Y.; Barreda, H.; Jung, S.Y.; Choi, J.M.; et al. Comprehensive Molecular Profiles of Functionally Effective MSC-Derived Extracellular Vesicles in Immunomodulation. Mol. Ther. J. Am. Soc. Gene Ther. 2020, 28, 1628–1644. [Google Scholar] [CrossRef]
- Rashedi, I.; Gómez-Aristizábal, A.; Wang, X.-H.; Viswanathan, S.; Keating, A. TLR3 or TLR4 Activation Enhances Mesenchymal Stromal Cell-Mediated Treg Induction via Notch Signaling. Stem Cells 2017, 35, 265–275. [Google Scholar] [CrossRef]
- Cheng, A.; Choi, D.; Lora, M.; Shum-Tim, D.; Rak, J.; Colmegna, I. Human Multipotent Mesenchymal Stromal Cells Cytokine Priming Promotes RAB27B-Regulated Secretion of Small Extracellular Vesicles with Immunomodulatory Cargo. Stem Cell Res. Ther. 2020, 11, 539. [Google Scholar] [CrossRef]
- Domenis, R.; Cifù, A.; Quaglia, S.; Pistis, C.; Moretti, M.; Vicario, A.; Parodi, P.C.; Fabris, M.; Niazi, K.R.; Soon-Shiong, P.; et al. Pro Inflammatory Stimuli Enhance the Immunosuppressive Functions of Adipose Mesenchymal Stem Cells-Derived Exosomes. Sci. Rep. 2018, 8, 13325. [Google Scholar] [CrossRef]
- Li, S.; Stöckl, S.; Lukas, C.; Herrmann, M.; Brochhausen, C.; König, M.A.; Johnstone, B.; Grässel, S. Curcumin-Primed Human BMSC-Derived Extracellular Vesicles Reverse IL-1β-Induced Catabolic Responses of OA Chondrocytes by Upregulating miR-126-3p. Stem Cell Res. Ther. 2021, 12, 252. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Xu, B. TGF-Β1-Modified MSC-Derived Exosomal miR-135b Attenuates Cartilage Injury via Promoting M2 Synovial Macrophage Polarization by Targeting MAPK6. Cell Tissue Res. 2021, 384, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-H.; Lechman, E.R.; Bianco, N.; Menon, R.; Keravala, A.; Nash, J.; Mi, Z.; Watkins, S.C.; Gambotto, A.; Robbins, P.D. Exosomes Derived from IL-10-Treated Dendritic Cells Can Suppress Inflammation and Collagen-Induced Arthritis. J. Immunol. 2005, 174, 6440–6448. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Jin, Z. Bone Mesenchymal Stem Cell-Derived Extracellular Vesicles Containing Long Noncoding RNA NEAT1 Relieve Osteoarthritis. Oxid. Med. Cell. Longev. 2022, 2022, 5517648. [Google Scholar] [CrossRef]
- Sage, E.K.; Thakrar, R.M.; Janes, S.M. Genetically Modified Mesenchymal Stromal Cells in Cancer Therapy. Cytotherapy 2016, 18, 1435–1445. [Google Scholar] [CrossRef]
- Oggu, G.S.; Sasikumar, S.; Reddy, N.; Ella, K.K.R.; Rao, C.M.; Bokara, K.K. Gene Delivery Approaches for Mesenchymal Stem Cell Therapy: Strategies to Increase Efficiency and Specificity. Stem Cell Rev. Rep. 2017, 13, 725–740. [Google Scholar] [CrossRef]
- Doi, K.; Takeuchi, Y. Gene therapy using retrovirus vectors: Vector development and biosafety at clinical trials. Uirusu 2015, 65, 27–36. [Google Scholar] [CrossRef]
- Ricks, D.M.; Kutner, R.; Zhang, X.-Y.; Welsh, D.A.; Reiser, J. Optimized Lentiviral Transduction of Mouse Bone Marrow-Derived Mesenchymal Stem Cells. Stem Cells Dev. 2008, 17, 441–450. [Google Scholar] [CrossRef]
- Zaiss, A.-K.; Liu, Q.; Bowen, G.P.; Wong, N.C.W.; Bartlett, J.S.; Muruve, D.A. Differential Activation of Innate Immune Responses by Adenovirus and Adeno-Associated Virus Vectors. J. Virol. 2002, 76, 4580–4590. [Google Scholar] [CrossRef]
- Shen, T.; Alvarez-Garcia, O.; Li, Y.; Olmer, M.; Lotz, M.K. Suppression of Sestrins in Aging and Osteoarthritic Cartilage: Dysfunction of an Important Stress Defense Mechanism. Osteoarthr. Cartil. 2017, 25, 287–296. [Google Scholar] [CrossRef]
- Meng, H.-Y.; Chen, L.-Q.; Chen, L.-H. The Inhibition by Human MSCs-Derived miRNA-124a Overexpression Exosomes in the Proliferation and Migration of Rheumatoid Arthritis-Related Fibroblast-like Synoviocyte Cell. BMC Musculoskelet. Disord. 2020, 21, 150. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Dai, L.; Wu, H.; Zhang, Z.-R.; Wang, W.-Y.; Fu, J.; Deng, R.; Li, F.; Dai, X.-J.; Zhan, X. Novel Anti-Inflammatory Target of Geniposide: Inhibiting Itgβ1/Ras-Erk1/2 Signal Pathway via the miRNA-124a in Rheumatoid Arthritis Synovial Fibroblasts. Int. Immunopharmacol. 2018, 65, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Bianco, N.R.; Kim, S.H.; Ruffner, M.A.; Robbins, P.D. Therapeutic Effect of Exosomes from Indoleamine 2,3-Dioxygenase-Positive Dendritic Cells in Collagen-Induced Arthritis and Delayed-Type Hypersensitivity Disease Models. Arthritis Rheum. 2009, 60, 380–389. [Google Scholar] [CrossRef]
- Yin, H.; Kanasty, R.L.; Eltoukhy, A.A.; Vegas, A.J.; Dorkin, J.R.; Anderson, D.G. Non-Viral Vectors for Gene-Based Therapy. Nat. Rev. Genet. 2014, 15, 541–555. [Google Scholar] [CrossRef] [PubMed]
- Hamann, A.; Nguyen, A.; Pannier, A.K. Nucleic Acid Delivery to Mesenchymal Stem Cells: A Review of Nonviral Methods and Applications. J. Biol. Eng. 2019, 13, 7. [Google Scholar] [CrossRef]
- Park, J.S.; Na, K.; Woo, D.G.; Yang, H.N.; Kim, J.M.; Kim, J.H.; Chung, H.-M.; Park, K.-H. Non-Viral Gene Delivery of DNA Polyplexed with Nanoparticles Transfected into Human Mesenchymal Stem Cells. Biomaterials 2010, 31, 124–132. [Google Scholar] [CrossRef]
- Liu, W.; Liu, A.; Li, X.; Sun, Z.; Sun, Z.; Liu, Y.; Wang, G.; Huang, D.; Xiong, H.; Yu, S.; et al. Dual-Engineered Cartilage-Targeting Extracellular Vesicles Derived from Mesenchymal Stem Cells Enhance Osteoarthritis Treatment via miR-223/NLRP3/Pyroptosis Axis: Toward a Precision Therapy. Bioact. Mater. 2023, 30, 169–183. [Google Scholar] [CrossRef]
- Chang, L.; Kan, L. Mesenchymal Stem Cell-Originated Exosomal Circular RNA circFBXW7 Attenuates Cell Proliferation, Migration and Inflammation of Fibroblast-Like Synoviocytes by Targeting miR-216a-3p/HDAC4 in Rheumatoid Arthritis. J. Inflamm. Res. 2021, 14, 6157–6171. [Google Scholar] [CrossRef]
- Yan, L.; Liu, G.; Wu, X. The Umbilical Cord Mesenchymal Stem Cell-Derived Exosomal lncRNA H19 Improves Osteochondral Activity through miR-29b-3p/FoxO3 Axis. Clin. Transl. Med. 2021, 11, e255. [Google Scholar] [CrossRef]
- Saliminejad, K.; Khorram Khorshid, H.R.; Soleymani Fard, S.; Ghaffari, S.H. An Overview of microRNAs: Biology, Functions, Therapeutics, and Analysis Methods. J. Cell. Physiol. 2019, 234, 5451–5465. [Google Scholar] [CrossRef]
- Lu, T.X.; Rothenberg, M.E. MicroRNA. J. Allergy Clin. Immunol. 2018, 141, 1202–1207. [Google Scholar] [CrossRef] [PubMed]
- Nygaard, G.; Firestein, G.S. Restoring Synovial Homeostasis in Rheumatoid Arthritis by Targeting Fibroblast-like Synoviocytes. Nat. Rev. Rheumatol. 2020, 16, 316–333. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, H.; Deng, R. Angiogenesis as a Potential Treatment Strategy for Rheumatoid Arthritis. Eur. J. Pharmacol. 2021, 910, 174500. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, Y.; Liu, Y.; Dai, X.; Li, W.; Cai, X.; Yin, Y.; Wang, Q.; Xue, Y.; Wang, C.; et al. Microvesicle-Mediated Transfer of microRNA-150 from Monocytes to Endothelial Cells Promotes Angiogenesis. J. Biol. Chem. 2013, 288, 23586–23596. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, H.; Xia, Y.; Yan, F.; Lu, Y. Therapeutic Potential of Mesenchymal Cell-Derived miRNA-150-5p-Expressing Exosomes in Rheumatoid Arthritis Mediated by the Modulation of MMP14 and VEGF. J. Immunol. 2018, 201, 2472–2482. [Google Scholar] [CrossRef]
- Lin, Z.; Li, W.; Wang, Y.; Lang, X.; Sun, W.; Zhu, X.; Bian, R.; Ma, Y.; Wei, X.; Zhang, J.; et al. SMSCs-Derived sEV Overexpressing miR-433-3p Inhibits Angiogenesis Induced by sEV Released from Synoviocytes under Triggering of Ferroptosis. Int. Immunopharmacol. 2023, 116, 109875. [Google Scholar] [CrossRef]
- Wang, Z.; Yan, K.; Ge, G.; Zhang, D.; Bai, J.; Guo, X.; Zhou, J.; Xu, T.; Xu, M.; Long, X.; et al. Exosomes Derived from miR-155-5p-Overexpressing Synovial Mesenchymal Stem Cells Prevent Osteoarthritis via Enhancing Proliferation and Migration, Attenuating Apoptosis, and Modulating Extracellular Matrix Secretion in Chondrocytes. Cell Biol. Toxicol. 2021, 37, 85–96. [Google Scholar] [CrossRef]
- Chen, D.; Kim, D.J.; Shen, J.; Zou, Z.; O’Keefe, R.J. Runx2 Plays a Central Role in Osteoarthritis Development. J. Orthop. Transl. 2020, 23, 132–139. [Google Scholar] [CrossRef]
- Huang, Y.; Zhang, X.; Zhan, J.; Yan, Z.; Chen, D.; Xue, X.; Pan, X. Bone Marrow Mesenchymal Stem Cell-Derived Exosomal miR-206 Promotes Osteoblast Proliferation and Differentiation in Osteoarthritis by Reducing Elf3. J. Cell. Mol. Med. 2021, 25, 7734–7745. [Google Scholar] [CrossRef]
- Wondimu, E.B.; Culley, K.L.; Quinn, J.; Chang, J.; Dragomir, C.L.; Plumb, D.A.; Goldring, M.B.; Otero, M. Elf3 Contributes to Cartilage Degradation in Vivo in a Surgical Model of Post-Traumatic Osteoarthritis. Sci. Rep. 2018, 8, 6438. [Google Scholar] [CrossRef]
- Conde, J.; Otero, M.; Scotece, M.; Abella, V.; Gómez, R.; López, V.; Pino, J.; Mera, A.; Goldring, M.B.; Gualillo, O. E74-Like Factor (ELF3) and Leptin, a Novel Loop Between Obesity and Inflammation Perpetuating a Pro-Catabolic State in Cartilage. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2018, 45, 2401–2410. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Liu, J.; Liu, S.; Jiao, W.; Wang, X. Mesenchymal Stem Cell-Derived Extracellular Vesicles Prevent the Development of Osteoarthritis via the circHIPK3/miR-124-3p/MYH9 Axis. J. Nanobiotechnol. 2021, 19, 194. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Liang, Y.-H.; Yang, Y.-T.; He, M.; Cai, Z.-J.; Xiao, W.-F.; Li, Y.-S. Circular RNA in Osteoarthritis: An Updated Insight into the Pathophysiology and Therapeutics. Am. J. Transl. Res. 2021, 13, 11–23. [Google Scholar] [PubMed]
- Kristensen, L.S.; Andersen, M.S.; Stagsted, L.V.W.; Ebbesen, K.K.; Hansen, T.B.; Kjems, J. The Biogenesis, Biology and Characterization of Circular RNAs. Nat. Rev. Genet. 2019, 20, 675–691. [Google Scholar] [CrossRef]
- Mao, G.; Xu, Y.; Long, D.; Sun, H.; Li, H.; Xin, R.; Zhang, Z.; Li, Z.; Yang, Z.; Kang, Y. Exosome-Transported circRNA_0001236 Enhances Chondrogenesis and Suppress Cartilage Degradation via the miR-3677-3p/Sox9 Axis. Stem Cell Res. Ther. 2021, 12, 389. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, Y.; Ma, Y.; Luo, L.; Chu, M.; Zhang, Z. Therapeutic Potential of Exosomal circRNA Derived from Synovial Mesenchymal Cells via Targeting circEDIL3/miR-485-3p/PIAS3/STAT3/VEGF Functional Module in Rheumatoid Arthritis. Int. J. Nanomed. 2021, 16, 7977–7994. [Google Scholar] [CrossRef]
- Yang, Q.; Yao, Y.; Zhao, D.; Zou, H.; Lai, C.; Xiang, G.; Wang, G.; Luo, L.; Shi, Y.; Li, Y.; et al. LncRNA H19 Secreted by Umbilical Cord Blood Mesenchymal Stem Cells through microRNA-29a-3p/FOS Axis for Central Sensitization of Pain in Advanced Osteoarthritis. Am. J. Transl. Res. 2021, 13, 1245–1256. [Google Scholar]
- Su, Y.; Liu, Y.; Ma, C.; Guan, C.; Ma, X.; Meng, S. Mesenchymal Stem Cell-Originated Exosomal lncRNA HAND2-AS1 Impairs Rheumatoid Arthritis Fibroblast-like Synoviocyte Activation through miR-143-3p/TNFAIP3/NF-κB Pathway. J. Orthop. Surg. 2021, 16, 116. [Google Scholar] [CrossRef]
- Wang, Q.; Yu, J.; Kadungure, T.; Beyene, J.; Zhang, H.; Lu, Q. ARMMs as a Versatile Platform for Intracellular Delivery of Macromolecules. Nat. Commun. 2018, 9, 960. [Google Scholar] [CrossRef]
- Kim, S.H.; Bianco, N.R.; Shufesky, W.J.; Morelli, A.E.; Robbins, P.D. Effective Treatment of Inflammatory Disease Models with Exosomes Derived from Dendritic Cells Genetically Modified to Express IL-4. J. Immunol. 2007, 179, 2242–2249. [Google Scholar] [CrossRef]
- Xie, A.; Xue, J.; Wang, Y.; Yang, C.; Xu, M.; Jiang, Y. Kartogenin Induced Adipose-Derived Stem Cell Exosomes Enhance the Chondrogenic Differentiation Ability of Adipose-Derived Stem Cells. Dis. Mark. 2022, 2022, 6943630. [Google Scholar] [CrossRef] [PubMed]
- Junxu, L.; Yuanbo, W.; Yanuan, H.; Biaobing, Y. Recent Advances and Applications of Kartogenin in Cartilage Regeneration and Repair. Chin. J. Tissue Eng. Res. 2020, 24, 5221. [Google Scholar] [CrossRef]
- Li, H.; Feng, Y.; Zheng, X.; Jia, M.; Mei, Z.; Wang, Y.; Zhang, Z.; Zhou, M.; Li, C. M2-Type Exosomes Nanoparticles for Rheumatoid Arthritis Therapy via Macrophage Re-Polarization. J. Control. Release 2022, 341, 16–30. [Google Scholar] [CrossRef] [PubMed]
- Mao, G.; Zhang, Z.; Hu, S.; Zhang, Z.; Chang, Z.; Huang, Z.; Liao, W.; Kang, Y. Exosomes Derived from miR-92a-3p-Overexpressing Human Mesenchymal Stem Cells Enhance Chondrogenesis and Suppress Cartilage Degradation via Targeting WNT5A. Stem Cell Res. Ther. 2018, 9, 247. [Google Scholar] [CrossRef]
- Feng, K.; Xie, X.; Yuan, J.; Gong, L.; Zhu, Z.; Zhang, J.; Li, H.; Yang, Y.; Wang, Y. Reversing the Surface Charge of MSC-Derived Small Extracellular Vesicles by εPL-PEG-DSPE for Enhanced Osteoarthritis Treatment. J. Extracell. Vesicles 2021, 10, e12160. [Google Scholar] [CrossRef]
- Wu, J.; Kuang, L.; Chen, C.; Yang, J.; Zeng, W.-N.; Li, T.; Chen, H.; Huang, S.; Fu, Z.; Li, J.; et al. miR-100-5p-Abundant Exosomes Derived from Infrapatellar Fat Pad MSCs Protect Articular Cartilage and Ameliorate Gait Abnormalities via Inhibition of mTOR in Osteoarthritis. Biomaterials 2019, 206, 87–100. [Google Scholar] [CrossRef]
- Tao, Y.; Zhou, J.; Wang, Z.; Tao, H.; Bai, J.; Ge, G.; Li, W.; Zhang, W.; Hao, Y.; Yang, X.; et al. Human Bone Mesenchymal Stem Cells-Derived Exosomal miRNA-361-5p Alleviates Osteoarthritis by Downregulating DDX20 and Inactivating the NF-κB Signaling Pathway. Bioorg. Chem. 2021, 113, 104978. [Google Scholar] [CrossRef]
- Wu, H.; Zhou, X.; Wang, X.; Cheng, W.; Hu, X.; Wang, Y.; Luo, B.; Huang, W.; Gu, J. miR-34a in Extracellular Vesicles from Bone Marrow Mesenchymal Stem Cells Reduces Rheumatoid Arthritis Inflammation via the Cyclin I/ATM/ATR/P53 Axis. J. Cell. Mol. Med. 2021, 25, 1896–1910. [Google Scholar] [CrossRef]
- Meng, Q.; Qiu, B. Exosomal MicroRNA-320a Derived From Mesenchymal Stem Cells Regulates Rheumatoid Arthritis Fibroblast-Like Synoviocyte Activation by Suppressing CXCL9 Expression. Front. Physiol. 2020, 11, 441. [Google Scholar] [CrossRef]
- Jin, Z.; Ren, J.; Qi, S. Exosomal miR-9-5p Secreted by Bone Marrow-Derived Mesenchymal Stem Cells Alleviates Osteoarthritis by Inhibiting Syndecan-1. Cell Tissue Res. 2020, 381, 99–114. [Google Scholar] [CrossRef]
- Wiklander, O.P.B.; Nordin, J.Z.; O’Loughlin, A.; Gustafsson, Y.; Corso, G.; Mäger, I.; Vader, P.; Lee, Y.; Sork, H.; Seow, Y.; et al. Extracellular Vesicle in Vivo Biodistribution Is Determined by Cell Source, Route of Administration and Targeting. J. Extracell. Vesicles 2015, 4, 26316. [Google Scholar] [CrossRef] [PubMed]
- Smyth, T.; Kullberg, M.; Malik, N.; Smith-Jones, P.; Graner, M.W.; Anchordoquy, T.J. Biodistribution and Delivery Efficiency of Unmodified Tumor-Derived Exosomes. J. Control. Release 2015, 199, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.P.; Mardini, O.; Ericsson, M.; Prabhakar, S.; Maguire, C.; Chen, J.W.; Tannous, B.A.; Breakefield, X.O. Dynamic Biodistribution of Extracellular Vesicles in Vivo Using a Multimodal Imaging Reporter. ACS Nano 2014, 8, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Byrska-Bishop, M.; Zhang, C.T.; Kastrup, C.J.; Hwang, N.S.; Tai, A.K.; Lee, W.W.; Xu, X.; Nahrendorf, M.; Langer, R.; et al. Stem Cell Membrane Engineering for Cell Rolling Using Peptide Conjugation and Tuning of Cell-Selectin Interaction Kinetics. Biomaterials 2012, 33, 5004–5012. [Google Scholar] [CrossRef]
- Mura, S.; Nicolas, J.; Couvreur, P. Stimuli-Responsive Nanocarriers for Drug Delivery. Nat. Mater. 2013, 12, 991–1003. [Google Scholar] [CrossRef]
- Agatemor, C.; Buettner, M.J.; Ariss, R.; Muthiah, K.; Saeui, C.T.; Yarema, K.J. Exploiting Metabolic Glycoengineering to Advance Healthcare. Nat. Rev. Chem. 2019, 3, 605–620. [Google Scholar] [CrossRef]
- Nagasundaram, M.; Horstkorte, R.; Gnanapragassam, V.S. Sialic Acid Metabolic Engineering of Breast Cancer Cells Interferes with Adhesion and Migration. Molecules 2020, 25, 2632. [Google Scholar] [CrossRef]
- Li, S.; Yu, B.; Wang, J.; Zheng, Y.; Zhang, H.; Walker, M.J.; Yuan, Z.; Zhu, H.; Zhang, J.; Wang, P.G.; et al. Biomarker-Based Metabolic Labeling for Redirected and Enhanced Immune Response. ACS Chem. Biol. 2018, 13, 1686–1694. [Google Scholar] [CrossRef]
- Xia, Y.; Rao, L.; Yao, H.; Wang, Z.; Ning, P.; Chen, X. Engineering Macrophages for Cancer Immunotherapy and Drug Delivery. Adv. Mater. 2020, 32, e2002054. [Google Scholar] [CrossRef]
- Rao, L.; Zhao, S.-K.; Wen, C.; Tian, R.; Lin, L.; Cai, B.; Sun, Y.; Kang, F.; Yang, Z.; He, L.; et al. Activating Macrophage-Mediated Cancer Immunotherapy by Genetically Edited Nanoparticles. Adv. Mater. 2020, 32, e2004853. [Google Scholar] [CrossRef]
- Yoon, H.Y.; Koo, H.; Kim, K.; Kwon, I.C. Molecular Imaging Based on Metabolic Glycoengineering and Bioorthogonal Click Chemistry. Biomaterials 2017, 132, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Ahern, B.J.; Parvizi, J.; Boston, R.; Schaer, T.P. Preclinical Animal Models in Single Site Cartilage Defect Testing: A Systematic Review. Osteoarthr. Cartil. 2009, 17, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Xu, X.; Li, X.; Xiong, J.; Li, B.; Duan, L.; Wang, D.; Xia, J. Chondrocyte-Targeted MicroRNA Delivery by Engineered Exosomes toward a Cell-Free Osteoarthritis Therapy. ACS Appl. Mater. Interfaces 2020, 12, 36938–36947. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Nishikawa, M.; Shinotsuka, H.; Matsui, Y.; Ohara, S.; Imai, T.; Takakura, Y. Visualization and in Vivo Tracking of the Exosomes of Murine Melanoma B16-BL6 Cells in Mice after Intravenous Injection. J. Biotechnol. 2013, 165, 77–84. [Google Scholar] [CrossRef]
- Ranjan, P.; Colin, K.; Dutta, R.K.; Verma, S.K. Challenges and Future Scope of Exosomes in the Treatment of Cardiovascular Diseases. J. Physiol. 2023, 601, 4873–4893. [Google Scholar] [CrossRef]
- Chen, P.; Zheng, L.; Wang, Y.; Tao, M.; Xie, Z.; Xia, C.; Gu, C.; Chen, J.; Qiu, P.; Mei, S.; et al. Desktop-Stereolithography 3D Printing of a Radially Oriented Extracellular Matrix/Mesenchymal Stem Cell Exosome Bioink for Osteochondral Defect Regeneration. Theranostics 2019, 9, 2439–2459. [Google Scholar] [CrossRef]
- Zhao, C.; Song, W.; Ma, J.; Wang, N. Macrophage-Derived Hybrid Exosome-Mimic Nanovesicles Loaded with Black Phosphorus for Multimodal Rheumatoid Arthritis Therapy. Biomater. Sci. 2022, 10, 6731–6739. [Google Scholar] [CrossRef]
- Quiñonez-Flores, C.M.; González-Chávez, S.A.; Del Río Nájera, D.; Pacheco-Tena, C. Oxidative Stress Relevance in the Pathogenesis of the Rheumatoid Arthritis: A Systematic Review. BioMed Res. Int. 2016, 2016, 6097417. [Google Scholar] [CrossRef]
- Rambhia, K.J.; Ma, P.X. Controlled Drug Release for Tissue Engineering. J. Control. Release 2015, 219, 119–128. [Google Scholar] [CrossRef]
- Jiang, S.; Tian, G.; Yang, Z.; Gao, X.; Wang, F.; Li, J.; Tian, Z.; Huang, B.; Wei, F.; Sang, X.; et al. Enhancement of Acellular Cartilage Matrix Scaffold by Wharton’s Jelly Mesenchymal Stem Cell-Derived Exosomes to Promote Osteochondral Regeneration. Bioact. Mater. 2021, 6, 2711–2728. [Google Scholar] [CrossRef]
- Bao, W.; Li, M.; Yang, Y.; Wan, Y.; Wang, X.; Bi, N.; Li, C. Advancements and Frontiers in the High Performance of Natural Hydrogels for Cartilage Tissue Engineering. Front. Chem. 2020, 8, 53. [Google Scholar] [CrossRef] [PubMed]
- Lenzini, S.; Bargi, R.; Chung, G.; Shin, J.-W. Matrix Mechanics and Water Permeation Regulate Extracellular Vesicle Transport. Nat. Nanotechnol. 2020, 15, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Xiong, J.; Yang, L.; Zhang, J.; Sun, S.; Liang, Y. Cell-Free Exosome-Laden Scaffolds for Tissue Repair. Nanoscale 2021, 13, 8740–8750. [Google Scholar] [CrossRef] [PubMed]
- Pang, L.; Jin, H.; Lu, Z.; Xie, F.; Shen, H.; Li, X.; Zhang, X.; Jiang, X.; Wu, L.; Zhang, M.; et al. Treatment with Mesenchymal Stem Cell-Derived Nanovesicle-Containing Gelatin Methacryloyl Hydrogels Alleviates Osteoarthritis by Modulating Chondrogenesis and Macrophage Polarization. Adv. Healthc. Mater. 2023, 12, e2300315. [Google Scholar] [CrossRef]
- Zhang, H.; Huang, J.; Alahdal, M. Exosomes Loaded with Chondrogenic Stimuli Agents Combined with 3D Bioprinting Hydrogel in the Treatment of Osteoarthritis and Cartilage Degeneration. Biomed. Pharmacother. 2023, 168, 115715. [Google Scholar] [CrossRef]
- Wang, Y.; Qin, J.-Z.; Xie, C.-Y.; Peng, X.-Z.; Wang, J.-H.; Wang, S.-J. Kartogenin-Loaded Exosomes Derived from Bone Marrow Mesenchymal Stem Cells Enhance Chondrogenesis and Expedite Tendon Enthesis Healing in a Rat Model of Rotator Cuff Injury. Am. J. Sports Med. 2024, 52, 3520–3535. [Google Scholar] [CrossRef]
- Cai, J.; Xu, J.; Ye, Z.; Wang, L.; Zheng, T.; Zhang, T.; Li, Y.; Jiang, J.; Zhao, J. Exosomes Derived from Kartogenin-Preconditioned Mesenchymal Stem Cells Promote Cartilage Formation and Collagen Maturation for Enthesis Regeneration in a Rat Model of Chronic Rotator Cuff Tear. Am. J. Sports Med. 2023, 51, 1267–1276. [Google Scholar] [CrossRef]
- Liu, X.; Yang, Y.; Li, Y.; Niu, X.; Zhao, B.; Wang, Y.; Bao, C.; Xie, Z.; Lin, Q.; Zhu, L. Integration of Stem Cell-Derived Exosomes with in Situ Hydrogel Glue as a Promising Tissue Patch for Articular Cartilage Regeneration. Nanoscale 2017, 9, 4430–4438. [Google Scholar] [CrossRef]
- Woo, C.H.; Kim, H.K.; Jung, G.Y.; Jung, Y.J.; Lee, K.S.; Yun, Y.E.; Han, J.; Lee, J.; Kim, W.S.; Choi, J.S.; et al. Small Extracellular Vesicles from Human Adipose-Derived Stem Cells Attenuate Cartilage Degeneration. J. Extracell. Vesicles 2020, 9, 1735249. [Google Scholar] [CrossRef]
- Lopez-Santalla, M.; Fernandez-Perez, R.; Garin, M.I. Mesenchymal Stem/Stromal Cells for Rheumatoid Arthritis Treatment: An Update on Clinical Applications. Cells 2020, 9, 1852. [Google Scholar] [CrossRef]
- Pers, Y.-M.; Rackwitz, L.; Ferreira, R.; Pullig, O.; Delfour, C.; Barry, F.; Sensebe, L.; Casteilla, L.; Fleury, S.; Bourin, P.; et al. Adipose Mesenchymal Stromal Cell-Based Therapy for Severe Osteoarthritis of the Knee: A Phase I Dose-Escalation Trial. Stem Cells Transl. Med. 2016, 5, 847–856. [Google Scholar] [CrossRef] [PubMed]
- Orozco, L.; Munar, A.; Soler, R.; Alberca, M.; Soler, F.; Huguet, M.; Sentís, J.; Sánchez, A.; García-Sancho, J. Treatment of Knee Osteoarthritis with Autologous Mesenchymal Stem Cells: A Pilot Study. Transplantation 2013, 95, 1535–1541. [Google Scholar] [CrossRef] [PubMed]
- Álvaro-Gracia, J.M.; Jover, J.A.; García-Vicuña, R.; Carreño, L.; Alonso, A.; Marsal, S.; Blanco, F.; Martínez-Taboada, V.M.; Taylor, P.; Martín-Martín, C.; et al. Intravenous Administration of Expanded Allogeneic Adipose-Derived Mesenchymal Stem Cells in Refractory Rheumatoid Arthritis (Cx611): Results of a Multicentre, Dose Escalation, Randomised, Single-Blind, Placebo-Controlled Phase Ib/IIa Clinical Trial. Ann. Rheum. Dis. 2017, 76, 196–202. [Google Scholar] [CrossRef] [PubMed]
- Garay-Mendoza, D.; Villarreal-Martínez, L.; Garza-Bedolla, A.; Pérez-Garza, D.M.; Acosta-Olivo, C.; Vilchez-Cavazos, F.; Diaz-Hutchinson, C.; Gómez-Almaguer, D.; Jaime-Pérez, J.C.; Mancías-Guerra, C. The Effect of Intra-Articular Injection of Autologous Bone Marrow Stem Cells on Pain and Knee Function in Patients with Osteoarthritis. Int. J. Rheum. Dis. 2018, 21, 140–147. [Google Scholar] [CrossRef]
- Shadmanfar, S.; Labibzadeh, N.; Emadedin, M.; Jaroughi, N.; Azimian, V.; Mardpour, S.; Kakroodi, F.A.; Bolurieh, T.; Hosseini, S.E.; Chehrazi, M.; et al. Intra-Articular Knee Implantation of Autologous Bone Marrow-Derived Mesenchymal Stromal Cells in Rheumatoid Arthritis Patients with Knee Involvement: Results of a Randomized, Triple-Blind, Placebo-Controlled Phase 1/2 Clinical Trial. Cytotherapy 2018, 20, 499–506. [Google Scholar] [CrossRef]
- Bastos, R.; Mathias, M.; Andrade, R.; Amaral, R.J.F.C.; Schott, V.; Balduino, A.; Bastos, R.; Miguel Oliveira, J.; Reis, R.L.; Rodeo, S.; et al. Intra-Articular Injection of Culture-Expanded Mesenchymal Stem Cells with or without Addition of Platelet-Rich Plasma Is Effective in Decreasing Pain and Symptoms in Knee Osteoarthritis: A Controlled, Double-Blind Clinical Trial. Knee Surg. Sports Traumatol. Arthrosc. 2020, 28, 1989–1999. [Google Scholar] [CrossRef]
- Copp, G.; Robb, K.P.; Viswanathan, S. Culture-Expanded Mesenchymal Stromal Cell Therapy: Does It Work in Knee Osteoarthritis? A Pathway to Clinical Success. Cell. Mol. Immunol. 2023, 20, 626–650. [Google Scholar] [CrossRef]
- Schioppo, T.; Ubiali, T.; Ingegnoli, F.; Bollati, V.; Caporali, R. The Role of Extracellular Vesicles in Rheumatoid Arthritis: A Systematic Review. Clin. Rheumatol. 2021, 40, 3481–3497. [Google Scholar] [CrossRef]
- Schmitz, C.; Alt, C.; Pearce, D.A.; Furia, J.P.; Maffulli, N.; Alt, E.U. Methodological Flaws in Meta-Analyses of Clinical Studies on the Management of Knee Osteoarthritis with Stem Cells: A Systematic Review. Cells 2022, 11, 965. [Google Scholar] [CrossRef]
- Wiggers, T.G.; Winters, M.; Van den Boom, N.A.; Haisma, H.J.; Moen, M.H. Autologous Stem Cell Therapy in Knee Osteoarthritis: A Systematic Review of Randomised Controlled Trials. Br. J. Sports Med. 2021, 55, 1161–1169. [Google Scholar] [CrossRef]
- Carneiro, D.d.C.; Araújo, L.T.d.; Santos, G.C.; Damasceno, P.K.F.; Vieira, J.L.; Santos, R.R.d.; Barbosa, J.D.V.; Soares, M.B.P. Clinical Trials with Mesenchymal Stem Cell Therapies for Osteoarthritis: Challenges in the Regeneration of Articular Cartilage. Int. J. Mol. Sci. 2023, 24, 9939. [Google Scholar] [CrossRef] [PubMed]
- Sarsenova, M.; Issabekova, A.; Abisheva, S.; Rutskaya-Moroshan, K.; Ogay, V.; Saparov, A. Mesenchymal Stem Cell-Based Therapy for Rheumatoid Arthritis. Int. J. Mol. Sci. 2021, 22, 11592. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.R.; Szczodry, M.; Bruno, S. Animal Models for Cartilage Regeneration and Repair. Tissue Eng. Part B Rev. 2010, 16, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Haraszti, R.A.; Miller, R.; Dubuke, M.L.; Rockwell, H.E.; Coles, A.H.; Sapp, E.; Didiot, M.-C.; Echeverria, D.; Stoppato, M.; Sere, Y.Y.; et al. Serum Deprivation of Mesenchymal Stem Cells Improves Exosome Activity and Alters Lipid and Protein Composition. iScience 2019, 16, 230–241. [Google Scholar] [CrossRef]
- Otsuru, S.; Hofmann, T.J.; Raman, P.; Olson, T.S.; Guess, A.J.; Dominici, M.; Horwitz, E.M. Genomic and Functional Comparison of Mesenchymal Stromal Cells Prepared Using Two Isolation Methods. Cytotherapy 2015, 17, 262–270. [Google Scholar] [CrossRef]
- Toh, W.S.; Brittberg, M.; Farr, J.; Foldager, C.B.; Gomoll, A.H.; Hui, J.H.P.; Richardson, J.B.; Roberts, S.; Spector, M. Cellular Senescence in Aging and Osteoarthritis. Acta Orthop. 2016, 87, 6–14. [Google Scholar] [CrossRef]
- Siegel, G.; Kluba, T.; Hermanutz-Klein, U.; Bieback, K.; Northoff, H.; Schäfer, R. Phenotype, Donor Age and Gender Affect Function of Human Bone Marrow-Derived Mesenchymal Stromal Cells. BMC Med. 2013, 11, 146. [Google Scholar] [CrossRef]
- McKee, C.; Chaudhry, G.R. Advances and Challenges in Stem Cell Culture. Colloids Surf. B Biointerfaces 2017, 159, 62–77. [Google Scholar] [CrossRef]
- Liu, Z.; Zhuang, Y.; Fang, L.; Yuan, C.; Wang, X.; Lin, K. Breakthrough of Extracellular Vesicles in Pathogenesis, Diagnosis and Treatment of Osteoarthritis. Bioact. Mater. 2022, 22, 423–452. [Google Scholar] [CrossRef]
- Coppin, L.; Sokal, E.; Stéphenne, X. Thrombogenic Risk Induced by Intravascular Mesenchymal Stem Cell Therapy: Current Status and Future Perspectives. Cells 2019, 8, 1160. [Google Scholar] [CrossRef]
- Lee, R.H.; Pulin, A.A.; Seo, M.J.; Kota, D.J.; Ylostalo, J.; Larson, B.L.; Semprun-Prieto, L.; Delafontaine, P.; Prockop, D.J. Intravenous hMSCs Improve Myocardial Infarction in Mice Because Cells Embolized in Lung Are Activated to Secrete the Anti-Inflammatory Protein TSG-6. Cell Stem Cell 2009, 5, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Phinney, D.G.; Pittenger, M.F. Concise Review: MSC-Derived Exosomes for Cell-Free Therapy. Stem Cells 2017, 35, 851–858. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal Information for Studies of Extracellular Vesicles 2018 (MISEV2018): A Position Statement of the International Society for Extracellular Vesicles and Update of the MISEV2014 Guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed]
- Jeyaram, A.; Jay, S.M. Preservation and Storage Stability of Extracellular Vesicles for Therapeutic Applications. AAPS J. 2017, 20, 1. [Google Scholar] [CrossRef]
- Wiklander, O.P.B.; Brennan, M.Á.; Lötvall, J.; Breakefield, X.O.; El Andaloussi, S. Advances in Therapeutic Applications of Extracellular Vesicles. Sci. Transl. Med. 2019, 11, eaav8521. [Google Scholar] [CrossRef]
- Moll, G.; Alm, J.J.; Davies, L.C.; von Bahr, L.; Heldring, N.; Stenbeck-Funke, L.; Hamad, O.A.; Hinsch, R.; Ignatowicz, L.; Locke, M.; et al. Do Cryopreserved Mesenchymal Stromal Cells Display Impaired Immunomodulatory and Therapeutic Properties? Stem Cells 2014, 32, 2430–2442. [Google Scholar] [CrossRef]
- Allan, D.; Tieu, A.; Lalu, M.; Burger, D. Mesenchymal Stromal Cell-Derived Extracellular Vesicles for Regenerative Therapy and Immune Modulation: Progress and Challenges toward Clinical Application. Stem Cells Transl. Med. 2020, 9, 39–46. [Google Scholar] [CrossRef]
- Kam, M.; Bekkers, R. Comparative Analysis of Standardized Protocols for EV Roaming; Report D6.1 for the evRoaming4EU Project; European Commission: Brussels, Belgium, 2020. [CrossRef]
- Toh, W.S.; Yarani, R.; Andaloussi, S.E.; Cho, B.S.; Choi, C.; Corteling, R.; Fougerolles, A.D.; Gimona, M.; Herz, J.; Khoury, M.; et al. A Report on the International Society for Cell & Gene Therapy 2022 Scientific Signature Series, “Therapeutic Advances with Native and Engineered Human Extracellular Vesicles”. Cytotherapy 2023, 25, 810–814. [Google Scholar] [CrossRef]
- Claridge, B.; Lozano, J.; Poh, Q.H.; Greening, D.W. Development of Extracellular Vesicle Therapeutics: Challenges, Considerations, and Opportunities. Front. Cell Dev. Biol. 2021, 9, 734720. [Google Scholar] [CrossRef]
- Wang, C.-K.; Tsai, T.-H.; Lee, C.-H. Regulation of Exosomes as Biologic Medicines: Regulatory Challenges Faced in Exosome Development and Manufacturing Processes. Clin. Transl. Sci. 2024, 17, e13904. [Google Scholar] [CrossRef] [PubMed]
- U.S. Food and Drug Administration. Regulatory Considerations for Human Cells, Tissues, and Cellular and Tissue-Based Products: Minimal Manipulation and Homologous Use. In Guidance for Industry and Food and Drug Administration Staff; U.S. Food and Drug Administration: Silver Spring, MD, USA, 2020. [Google Scholar]
- Lerussi, G.; Villagrasa-Araya, V.; Moltó-Abad, M.; del Toro, M.; Pintos-Morell, G.; Seras-Franzoso, J.; Abasolo, I. Extracellular Vesicles as Tools for Crossing the Blood–Brain Barrier to Treat Lysosomal Storage Diseases. Life 2025, 15, 70. [Google Scholar] [CrossRef] [PubMed]
- Shami-shah, A.; Travis, B.G.; Walt, D.R. Advances in Extracellular Vesicle Isolation Methods: A Path towards Cell-Type Specific EV Isolation. Extracell. Vesicles Circ. Nucleic Acids 2023, 4, 447–460. [Google Scholar] [CrossRef] [PubMed]
- Nelson, B.C.; Maragh, S.; Ghiran, I.C.; Jones, J.C.; DeRose, P.C.; Elsheikh, E.; Vreeland, W.N.; Wang, L. Measurement and Standardization Challenges for Extracellular Vesicle Therapeutic Delivery Vectors. Nanomedicine 2020, 15, 2149–2170. [Google Scholar] [CrossRef]
- Momen-Heravi, F. Isolation of Extracellular Vesicles by Ultracentrifugation. Methods Mol Biol. 2017, 1660, 25–32. [Google Scholar] [CrossRef]
- Ramirez, M.I.; Amorim, M.G.; Gadelha, C.; Milic, I.; Welsh, J.A.; Freitas, V.M.; Nawaz, M.; Akbar, N.; Couch, Y.; Makin, L.; et al. Technical Challenges of Working with Extracellular Vesicles. Nanoscale 2018, 10, 881–906. [Google Scholar] [CrossRef]
- Cricrì, G.; Bellucci, L.; Montini, G.; Collino, F. Urinary Extracellular Vesicles: Uncovering the Basis of the Pathological Processes in Kidney-Related Diseases. Int. J. Mol. Sci. 2021, 22, 6507. [Google Scholar] [CrossRef]
- Jones, M.T.; Manioci, S.W.; Russell, A.E. Size Exclusion Chromatography for Separating Extracellular Vesicles from Conditioned Cell Culture Media. J. Vis. Exp. JoVE 2022, 183, e63614. [Google Scholar] [CrossRef]
- Sidhom, K.; Obi, P.O.; Saleem, A. A Review of Exosomal Isolation Methods: Is Size Exclusion Chromatography the Best Option? Int. J. Mol. Sci. 2020, 21, 6466. [Google Scholar] [CrossRef]
- Paganini, C.; Palmiero, U.C.; Pocsfalvi, G.; Touzet, N.; Bongiovanni, A.; Arosio, P. Scalable Production and Isolation of Extracellular Vesicles: Available Sources and Lessons from Current Industrial Bioprocesses. Biotechnol. J. 2019, 14, e1800528. [Google Scholar] [CrossRef]
- Chernyshev, V.S.; Yashchenok, A.; Ivanov, M.; Silachev, D.N. Filtration-Based Technologies for Isolation, Purification and Analysis of Extracellular Vesicles. Phys. Chem. Chem. Phys. 2023, 25, 23344–23357. [Google Scholar] [CrossRef] [PubMed]
- Szatanek, R.; Baran, J.; Siedlar, M.; Baj-Krzyworzeka, M. Isolation of Extracellular Vesicles: Determining the Correct Approach (Review). Int. J. Mol. Med. 2015, 36, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Mas-Bargues, C.; Borrás, C. Importance of Stem Cell Culture Conditions for Their Derived Extracellular Vesicles Therapeutic Effect. Free Radic. Biol. Med. 2021, 168, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Wang, B.; Tang, T.; Lv, L.; Ding, Z.; Li, Z.; Hu, R.; Wei, Q.; Shen, A.; Fu, Y.; et al. Three-Dimensional Culture of MSCs Produces Exosomes with Improved Yield and Enhanced Therapeutic Efficacy for Cisplatin-Induced Acute Kidney Injury. Stem Cell Res. Ther. 2020, 11, 206. [Google Scholar] [CrossRef]
- Patel, D.B.; Luthers, C.R.; Lerman, M.J.; Fisher, J.P.; Jay, S.M. Enhanced Extracellular Vesicle Production and Ethanol-Mediated Vascularization Bioactivity via a 3D-Printed Scaffold-Perfusion Bioreactor System. Acta Biomater. 2019, 95, 236–244. [Google Scholar] [CrossRef]
- Gobin, J.; Muradia, G.; Mehic, J.; Westwood, C.; Couvrette, L.; Stalker, A.; Bigelow, S.; Luebbert, C.C.; Bissonnette, F.S.-D.; Johnston, M.J.W.; et al. Hollow-Fiber Bioreactor Production of Extracellular Vesicles from Human Bone Marrow Mesenchymal Stromal Cells Yields Nanovesicles That Mirrors the Immuno-Modulatory Antigenic Signature of the Producer Cell. Stem Cell Res. Ther. 2021, 12, 127. [Google Scholar] [CrossRef]
- Garcia, S.G.; Sanroque-Muñoz, M.; Clos-Sansalvador, M.; Font-Morón, M.; Monguió-Tortajada, M.; Borràs, F.E.; Franquesa, M. Hollow Fiber Bioreactor Allows Sustained Production of Immortalized Mesenchymal Stromal Cell-Derived Extracellular Vesicles. Extracell. Vesicles Circ. Nucleic Acids 2024, 5, 201–220. [Google Scholar] [CrossRef]
- Ludlow, J.W.; Buehrer, B.M. Analyses and Utilization of Selectively Tuned Human Adipose-Derived Stromal/Stem Cell Exosomes. Methods Mol Biol. 2024, 2783, 309–322. [Google Scholar] [CrossRef]
- Gao, W.-X.; Sun, Y.-Q.; Shi, J.; Li, C.-L.; Fang, S.-B.; Wang, D.; Deng, X.-Q.; Wen, W.; Fu, Q.-L. Effects of Mesenchymal Stem Cells from Human Induced Pluripotent Stem Cells on Differentiation, Maturation, and Function of Dendritic Cells. Stem Cell Res. Ther. 2017, 8, 48. [Google Scholar] [CrossRef]
- Spitzhorn, L.-S.; Megges, M.; Wruck, W.; Rahman, M.S.; Otte, J.; Degistirici, Ö.; Meisel, R.; Sorg, R.V.; Oreffo, R.O.C.; Adjaye, J. Human iPSC-Derived MSCs (iMSCs) from Aged Individuals Acquire a Rejuvenation Signature. Stem Cell Res. Ther. 2019, 10, 100. [Google Scholar] [CrossRef]
- Lee, J.H.; Ha, D.H.; Go, H.-K.; Youn, J.; Kim, H.-K.; Jin, R.C.; Miller, R.B.; Kim, D.-H.; Cho, B.S.; Yi, Y.W. Reproducible Large-Scale Isolation of Exosomes from Adipose Tissue-Derived Mesenchymal Stem/Stromal Cells and Their Application in Acute Kidney Injury. Int. J. Mol. Sci. 2020, 21, 4774. [Google Scholar] [CrossRef]
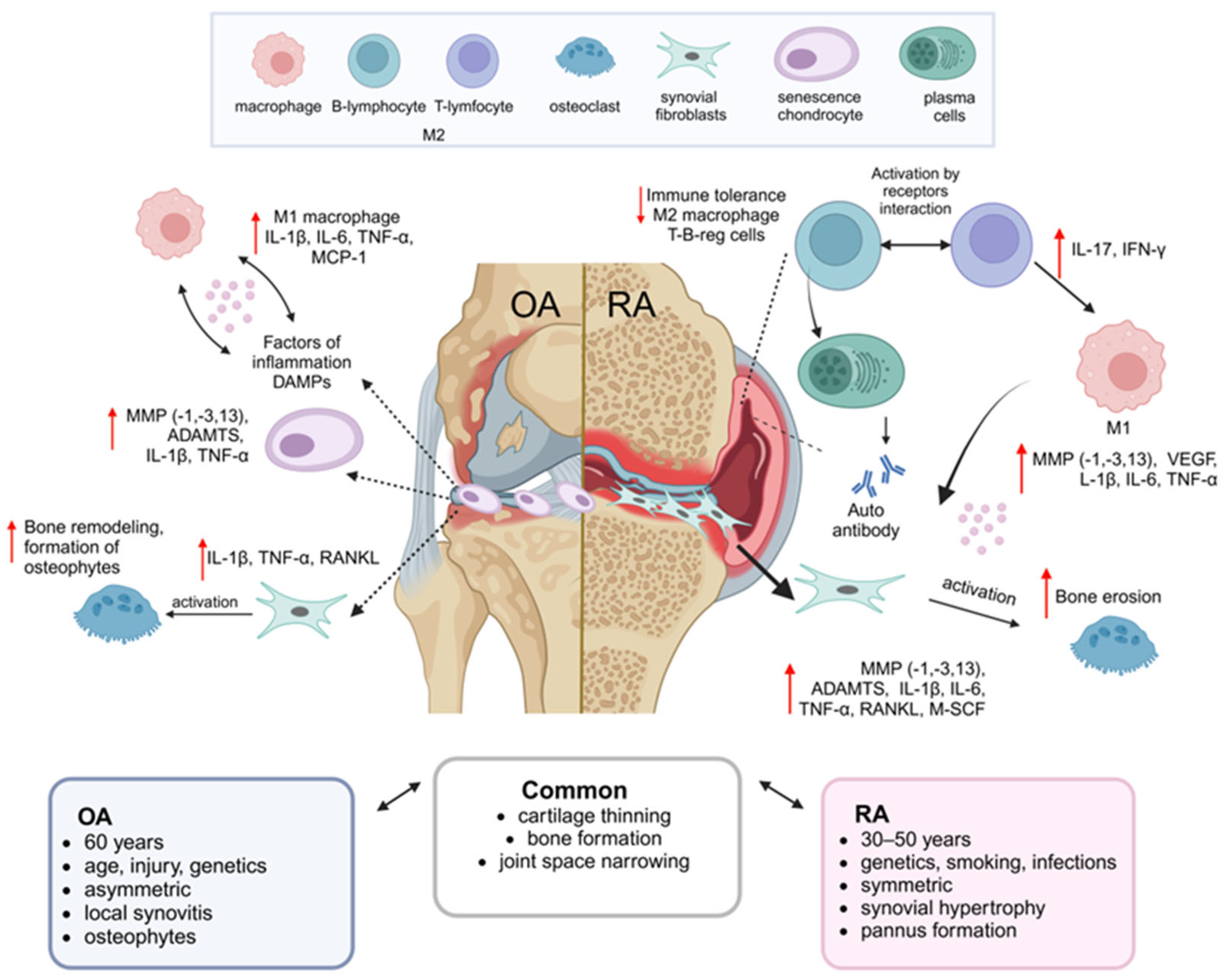
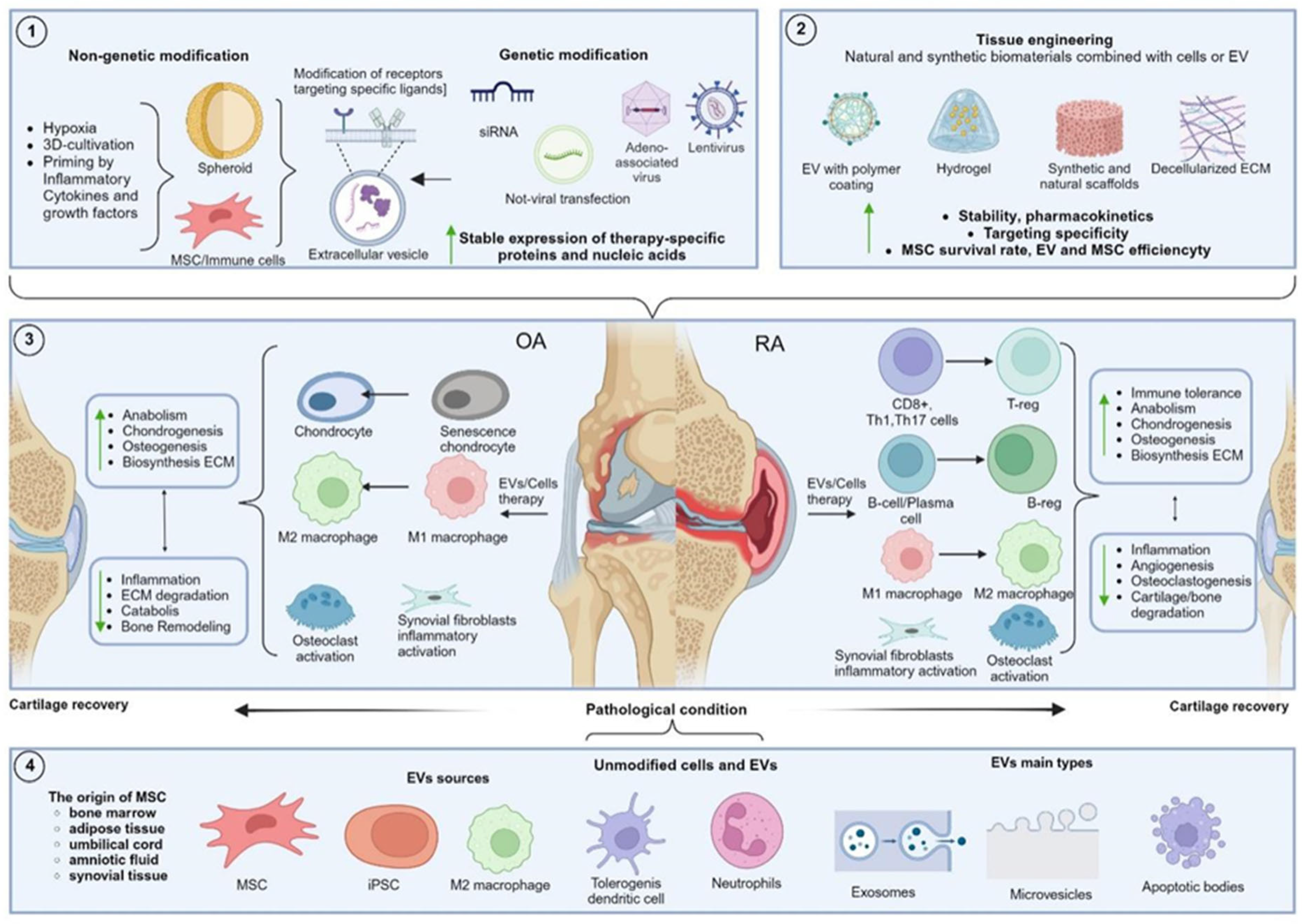

{kind=link}
{kind=link}
| Type Arthritis | Model of Arthritis/Type of Animal | Source EV | Cargo | The Cells Modification/EV Approach | Method of Administration, Concentration | Therapeutic Effects | Mechanism | Ref. | |
|---|---|---|---|---|---|---|---|---|---|
| ↑ | ↓ | ||||||||
| OA | DMM and ACLT | BMSC | hsa-miR-181c-5p, hsa-miR-18a-3p | Hypoxic conditions | Intraarticular injections, 100 µg protein per 100 µL PBS | Chondrocyte proliferation and migration | Apoptosis in IL-1β-treated chondrocytes OARSI scores | miRNA-18-3P/JAK/STAT or miRNA-181c-5p/MAPK pathways | [212] |
| OA | MLI | BMSC | miR-135b | Stimulation with TGF-β1 | Intraarticular injections, 1 × 1011 exosome particles/mL 100 µL PBS | EV-BMSC: Arg-1 and iNOS in macrophages | EV-BMSC-TGF-β1: IL-1β, PGE2, COX-2, COX-1, and NO in serum | EV-BMSC-TGF-β1: overexpression of miR-135b, M2 macrophage polarization via the MAPK6 pathway | [223] |
| RA | CIA, mice | BMSC | miR-205-5p | Chondrogenic differentiation | Tail vein injections, 200 µL PBS | - | TNF-α, IL-6, MMP1, and MMP13 in IL-1β-treated FLSs. MDM2, IL-1β, IL-6, and TNF-α in the serum | miR-205-5 expression, MAPK, and NF-κB pathways inhibition through MDM2 | [184] |
| OA | CIOA | BMSC | miR-92a-3p | Chondrogenic differentiation | Intraarticular injections, 15 µL of EV in PBS (500 µg/mL). | COL2A1, aggrecan | MMP13 | miR-92a-3p by targeting WNT5A | [265] |
| OA | DMM | BMSC | NEAT1 | Transduction with a lentivirus for NEAT1 overexpression | Intraarticular injections, 10 µg protein per dose | Chondrocyte proliferation and autophagy | Apoptosis | Binding miR-122-5p, activating the Sesn2/Nrf2 axis | [225] |
| OA | Intraarticular injections of cold water (4 °C) | SMSC | miR-155-5p | Transfection of miR-155-5p using Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc., Carlsbad, CA, USA). | Intraarticular injections, 30 μL; 1011 exo particles/mL PBS | Proliferation, migration, and ECM secretion in chondrocytes; number of chondrocytes | OARSI scores | By targeting Runx2 | [248] |
| OA | ACLT | BMSC | miR-206 | Transfection of miR-206 using Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc., Carlsbad, CA, USA) | Intraarticular injections, 1011 EV/mL, 100 µL PBS | OCN and BMP2, alkaline phosphatase activity, calcium deposition | Apoptosis in OA osteoblasts | miR-206-overexpression by downregulating the E74-like factor 3 | [250] |
| OA | MIA | UCMSC | miR-223 | (1) Transduction with a lentivirus encoding CTP-Lamp2b (2) exosome loading with miR-223 by electroporation | Intraarticular injections, 1010 particles/mL, 50 µL PBS | Col II and Sox9 in chondrocytes | MMP13, NLRP3, IL-1β, MMP2, IL-1β, TNF-α, and PGE2 | via the miR-223/NLRP3/pyroptosis axis | [238] |
| OA | DMM | BMSC | circRNA_0001236 | Transfection of a plasmid encoding circRNA_0001236 | Intraarticular injections, 500 µg/mL EV circRNA_0001236, 10 µL PBS | Col2a1 and Sox9 in human chondrocyte cartilage tissues | MMP13 | circRNA_0001236 through the miR-3677-3p/Sox9 axis | [256] |
| OA | ACLT | (iPSC) derived MSC | - | Surface charge by polyethylene distearoyl phosphatidylethanolamine incorporation | Intraarticular injections, modified and unmodified EV at a particle concentration of (1 × 109 particles/mL; 1 × 1010 particles/mL) | Chondrocyte uptake, Col2, aggrecan | IL-1β-induced apoptosis MMP13 and ADAMTS5 | - | [266] |
| RA | CIA | BMSC | FGL-1 | Transfection with an FGL1 plasmid using Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc., Carlsbad, CA, USA) | - | - | Apoptosis in FLSs. MMP-9, IL-8, IL-1β, and IL-17 levels in serum | By inhibiting the NF-κB transduction | [185] |
| OA | DMM | IPFP | miR-100-5p | - | Intraarticular injections, 10 µL MSC-IPFP-EV (1010 particles/mL) | Col2 | ADAMTS5 and MMP13 IL-1β-induced apoptosis in chondrocytes | miR-100-5p inhibits the mTOR-autophagy pathway | [267] |
| OA | ACLT | BMSC | miR-361-5p | miR-NC and miR-361-5p by electroporation | Intraarticular injections, Exo-miR-NC and Exo-miR-361-5p concentration of 250 ng/5 µL of PBS | - | iNOS, MMP-3, MMP-13, IL-18, IL-6, and TNF-α | miR-361-5p inhibits NF-κB signaling by suppressing DDX20 | [268] |
| RA | CIA | DC | Indolamine-2,3-dioxygenase IDO | Transduction with adenovirus expressing IDO | Intravenous injections, DC-Exo/DC-Exo-IDO concentration 1 μg protein in 20 μL of PBS | - | The arthritis index | The costimulatory molecules B7 | [234] |
| RA | CIA | BMMSC | miR-34a | Transfection with miR-34a inhibitor using Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc., Carlsbad, CA, USA) | Intravenous injections, BMMSC-EV 75 µg/mL of PBS | RA-FLS apoptosis | TNF-α, IL-6, and IL-8 mRNA in synovial tissue and synovial fluid. RA-FLS proliferation and increased | By inhibiting the Cyclin I/ATM/ATR/p53 signaling pathway | [269] |
| RA | CIA | GMDSC | miR-29a-3p and miR-93-5p | Transfection with miR-29a-3p and miR-93-5p with Entranster-R (Engreen Biosystem Co., Ltd., Beijing, China) | Intravenous injections, GMDSC-EV 100 μg/mouse | - | the mean arthritis index IFN-γ and IL-17A in the serum Th1 and Th17 differentiation in vitro | - | [197] |
| RA | CIA | BMMSC | miR-320a | Transfection with miR-320a plasmids using Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc., Carlsbad, CA, USA) | Intravenous injections, BMMSC-EV 100 µg/day | - | IL-1β, IL-6, and IL-8 in RA-FLSs | Overexpressed miR-320a by suppressing CXCL9 expression | [270] |
| OA | ACLT/MCLT | BMMSC | miR-9-5p | Transfection with mimic and inhibitors miR-9-5p using Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc., Carlsbad, CA, USA) | Intraarticular injections, BMMSC -EV, or liposomes containing miR-9-5p mimic/inhibitor | - | IL-1, IL-6, TNF-α, and CRP oxidative stress indicators (NO, iNOS, COX2, and SOD) MMP-13 and OCN in synovial fluid | Regulation of the expression of syndecan-1 | [271] |
| OA | ACLT | M2-Mφ | miR-21-5p | - | Intraarticular injections, M1-AB and M2-AB 10 µg/10 µL | M2-AB: IL-4 and IL-10 in M1 macrophages M1-ABs: CD-86 in synovial tissue, cartilage thickness | M2-Ab: IL-1β, IL-6, IL-17, TNF-α, IFN-γ, and MCP-1 CD206 in synovial tissue | - | [201] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klyucherev, T.O.; Peshkova, M.A.; Yurkanova, M.D.; Kosheleva, N.V.; Svistunov, A.A.; Liang, X.-J.; Timashev, P.S. Advances in Regenerative Therapies for Inflammatory Arthritis: Exploring the Potential of Mesenchymal Stem Cells and Extracellular Vesicles. Int. J. Mol. Sci. 2025, 26, 5766. https://doi.org/10.3390/ijms26125766
Klyucherev TO, Peshkova MA, Yurkanova MD, Kosheleva NV, Svistunov AA, Liang X-J, Timashev PS. Advances in Regenerative Therapies for Inflammatory Arthritis: Exploring the Potential of Mesenchymal Stem Cells and Extracellular Vesicles. International Journal of Molecular Sciences. 2025; 26(12):5766. https://doi.org/10.3390/ijms26125766
Chicago/Turabian StyleKlyucherev, Timofey O., Maria A. Peshkova, Maria D. Yurkanova, Nastasia V. Kosheleva, Andrey A. Svistunov, Xing-Jie Liang, and Peter S. Timashev. 2025. "Advances in Regenerative Therapies for Inflammatory Arthritis: Exploring the Potential of Mesenchymal Stem Cells and Extracellular Vesicles" International Journal of Molecular Sciences 26, no. 12: 5766. https://doi.org/10.3390/ijms26125766
APA StyleKlyucherev, T. O., Peshkova, M. A., Yurkanova, M. D., Kosheleva, N. V., Svistunov, A. A., Liang, X.-J., & Timashev, P. S. (2025). Advances in Regenerative Therapies for Inflammatory Arthritis: Exploring the Potential of Mesenchymal Stem Cells and Extracellular Vesicles. International Journal of Molecular Sciences, 26(12), 5766. https://doi.org/10.3390/ijms26125766