
Specific Heat-Killed Lactic Acid Bacteria Enhance Mucosal Aminopeptidase N Activity in the Small Intestine of Aged Mice
 ,
,
Abstract

1. Introduction
2. Results
2.1. Experiment 1
2.1.1. Body Weight, Skeletal Muscle and Adipose Tissue Weights, and Feed Intake
2.1.2. Total Length, Villous Height, and Mucosal APN Activity of Small Intestine
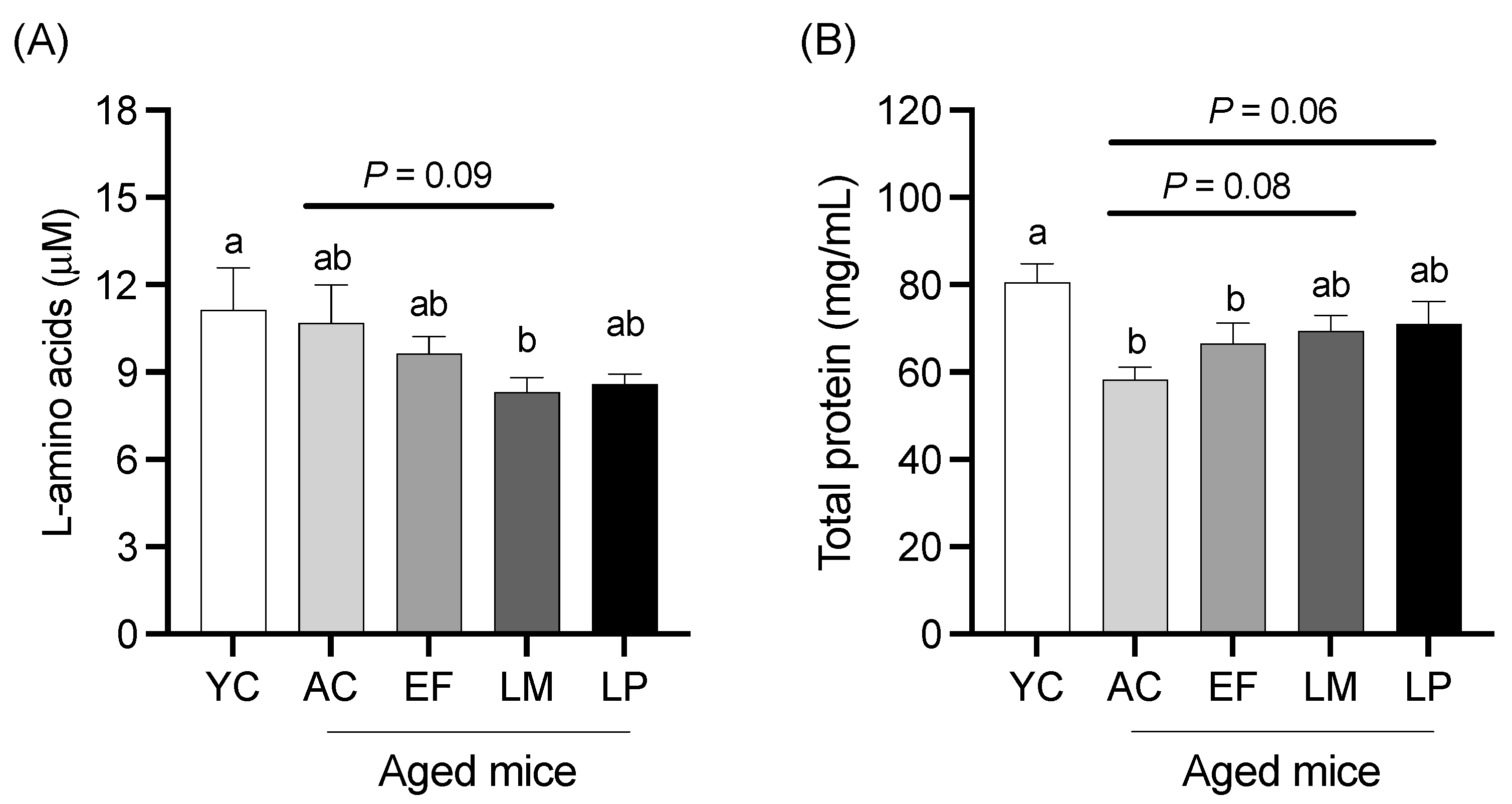
2.1.3. Serum L-Amino Acid and Total Protein Levels
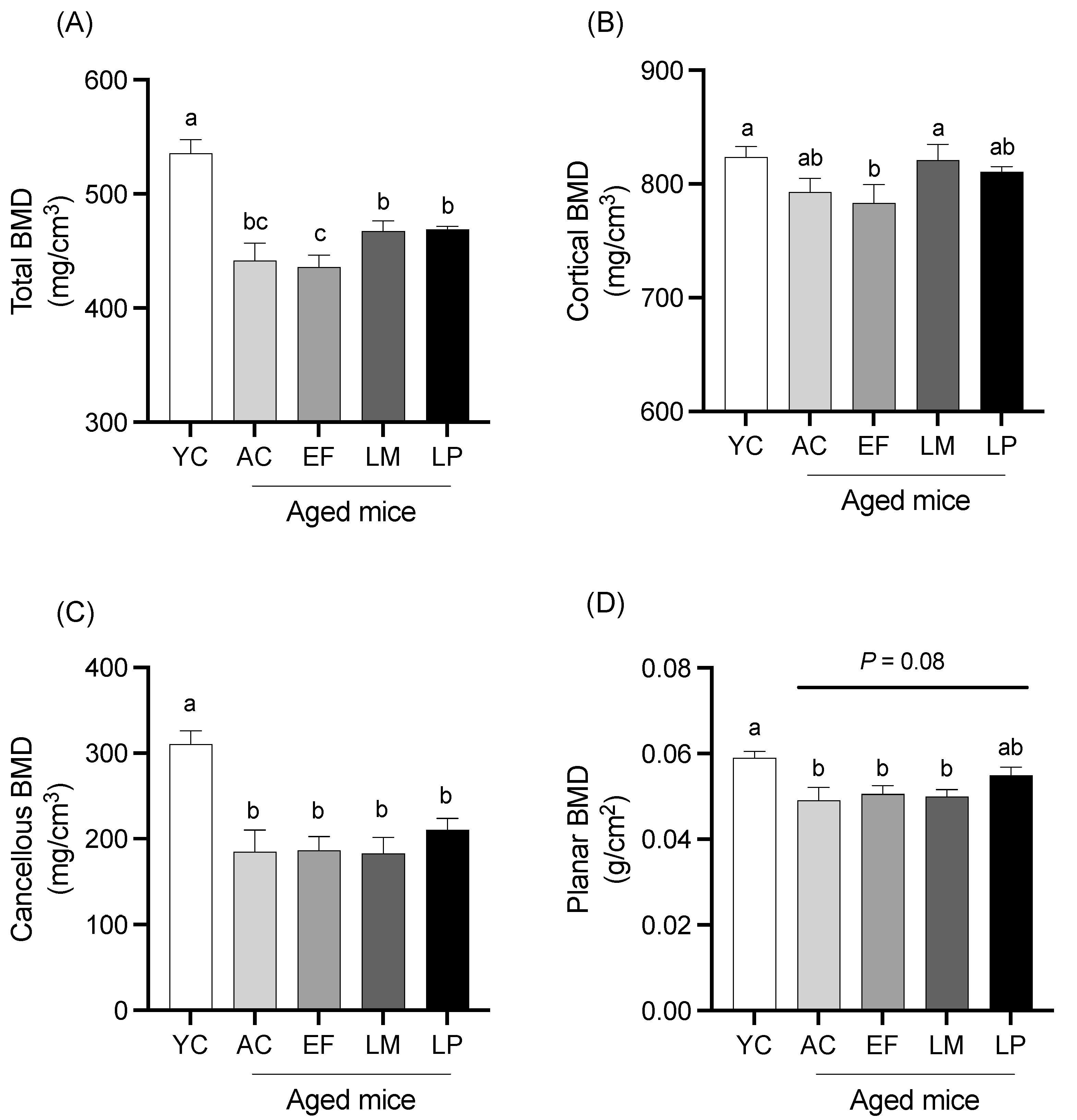
2.1.4. BMD of Femur
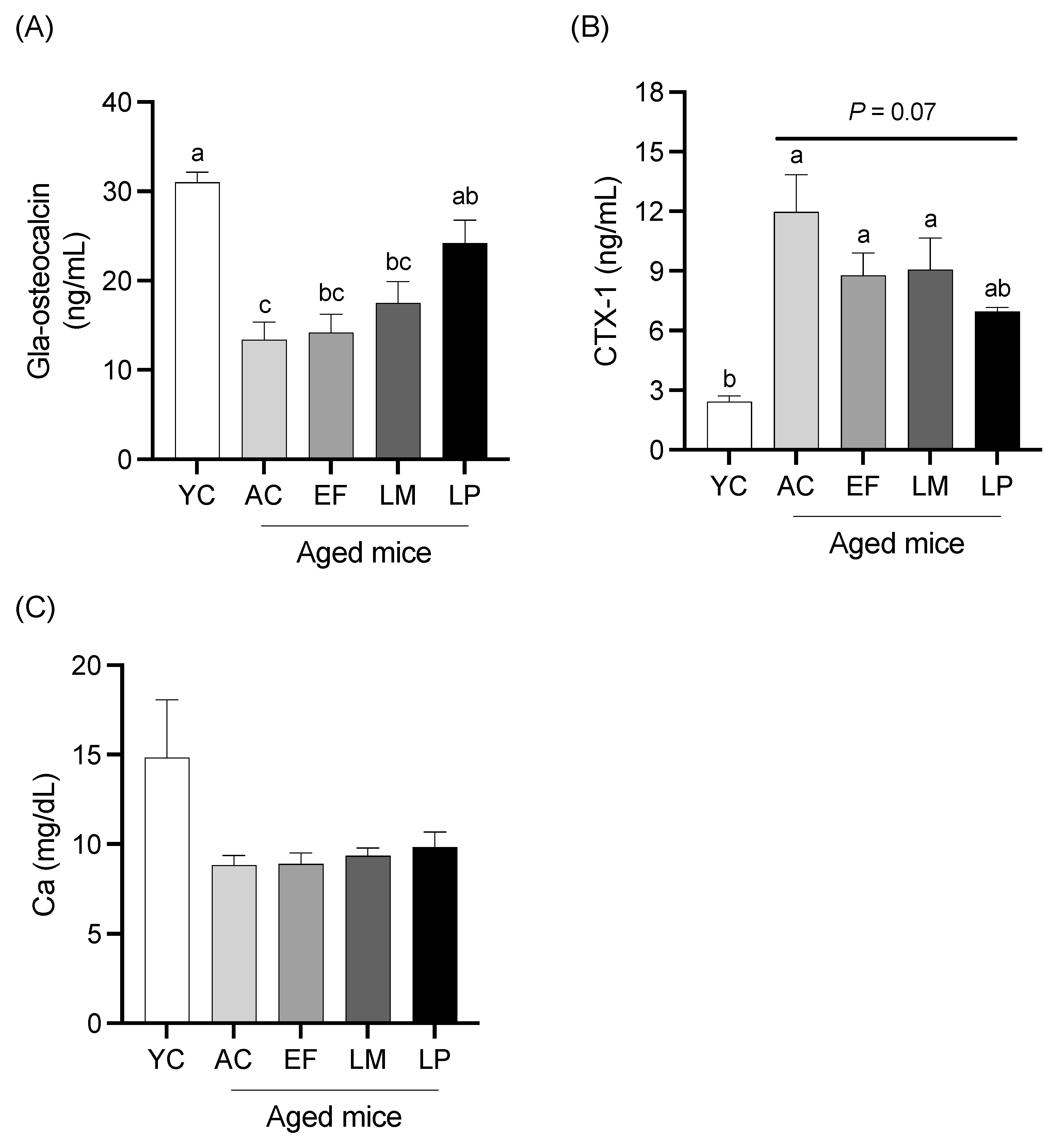
2.1.5. Serum Levels of Bone Metabolism Markers and Calcium
2.2. Experiment 2
2.2.1. APN Activity of IEC-6 Cells Co-Cultured with Heat-Killed LAB
2.2.2. APN Activity in IEC-6 Cells Co-Cultured with RNase-, DNase-, or Lysozyme-Treated LP
2.2.3. APN Activity of IEC-6 Cells Treated with TLR2 Agonists
2.2.4. TLR2 Neutralization Study
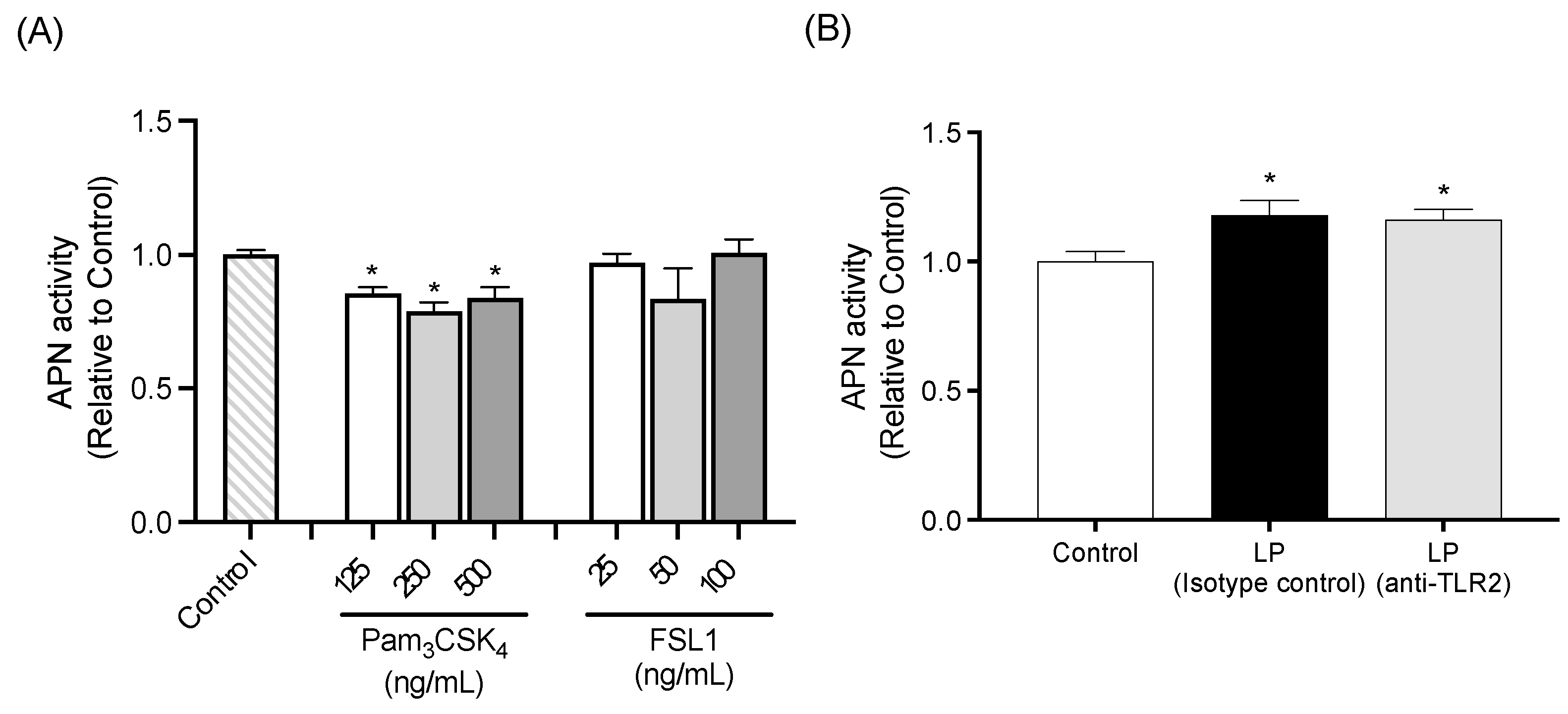
3. Discussion
4. Materials and Methods
4.1. Experiment 1
4.1.1. Preparation of Heat-Killed Lactic Acid Bacteria and Diet
4.1.2. Animal Experiment
4.1.3. APN Activity in Small Intestinal Mucosa
4.1.4. Measurement of Small Intestinal Villus Height
4.1.5. Micro-X-Ray-Computed Tomography Analysis of Femur
4.1.6. Serum Biochemical Analysis
4.2. Experiment 2
4.2.1. Cell Line
4.2.2. APN Activity in IEC-6 Cells Co-Cultured with Heat-Killed LAB
4.2.3. APN Activity in IEC-6 Cells Co-Cultured with RNase-, DNase-, or Lysozyme-Treated LP
4.2.4. APN Activity in IEC-6 Cells Treated with Toll-Like Receptor 2 Agonists
4.2.5. TLR2 Neutralization Study
4.2.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| APN | Aminopeptidase N |
| ANOVA | Analysis of variance |
| BCA | Bicinchoninic acid |
| BMD | Bone mineral density |
| CCK-8 | Cell counting kit-8 |
| CTX-1 | C-terminal telopeptides of type I collagen |
| EF | Enterococcus faecalis |
| FBS | Fetal bovine serum |
| FSL1 | Fibroblast-stimulating lipopeptide1 |
| GlcNAc | N-acetylglucosamine |
| HE | Hematoxylin and eosin |
| IEC-6 | Intestinal epithelial cell 6 |
| LAB | Lactic acid bacteria |
| LM | Leuconostoc mesenteroides |
| LP | Lactiplantibacillus plantarum |
| MurNAc | N-acetylmuramic acid |
| NEFA | Non-esterified fatty acids |
| NTX | N-terminal telopeptide of type I collagen |
| Pam3CSK4 | Pam3Cys-Ser-Lys4 |
| PGN | Peptidoglycan |
| pNA | p-nitroaniline |
| STING | Stimulator of interferon genes |
| TLR | Toll-like receptor |
| TNF-α | Tumor necrosis factor-α |
References
- Troen, B.R. The biology of aging. Mt. Sinai J. Med. 2003, 70, 3–22. [Google Scholar] [PubMed]
- Reuter-Lorenz, P.A.; Park, D.C. Human neuroscience and the aging mind: A new look at old problems. J. Gerontol. Ser. B 2010, 65, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Levy, B.R. Mind matters: Cognitive and physical effects of aging self-stereotypes. J. Gerontol. Ser. B Psychol. Sci. Soc. Sci. 2003, 58, P203–P211. [Google Scholar] [CrossRef]
- Höhn, P.; Gabbert, H.; Wagner, R. Differentiation and aging of the rat intestinal mucosa. II. Morphological, enzyme histochemical and disc electrophoretic aspects of the aging of the small intestinal mucosa. Mech. Ageing Dev. 1978, 7, 217–226. [Google Scholar] [CrossRef]
- Holt, P.R.; Yeh, K.-Y.; Kotler, D.P. Altered controls of proliferation in proximal small intestine of the senescent rat. Proc. Natl. Acad. Sci. USA 1988, 85, 2771–2775. [Google Scholar] [CrossRef]
- Martin, K.; Kirkwood, T.; Potten, C.S. Age changes in stem cells of murine small intestinal crypts. Exp. Cell Res. 1998, 241, 316–323. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, J.; Li, Q.; Zhang, J.; Duan, X.-L. Morphological changes of cell proliferation and apoptosis in rat jejunal mucosa at different ages. World J. Gastroenterol. 2003, 9, 2060. [Google Scholar] [CrossRef]
- Rosa, E.F.; Silva, A.C.; Ihara, S.S.; Mora, O.A.; Aboulafia, J.; Nouailhetas, V.L. Habitual exercise program protects murine intestinal, skeletal, and cardiac muscles against aging. J. Appl. Physiol. 2005, 99, 1569–1575. [Google Scholar] [CrossRef]
- Hassan, Z.A.; Zauszkiewicz-Pawlak, A.; Abdelrahman, S.A.; Algaidi, S.; Desouky, M.; Shalaby, S.M. Morphological alterations in the jejunal mucosa of aged rats and the possible protective role of green tea. Folia Histochem. Cytobiol. 2017, 55, 124–139. [Google Scholar] [CrossRef]
- Nalapareddy, K.; Nattamai, K.J.; Kumar, R.S.; Karns, R.; Wikenheiser-Brokamp, K.A.; Sampson, L.L.; Mahe, M.M.; Sundaram, N.; Yacyshyn, M.-B.; Yacyshyn, B. Canonical Wnt signaling ameliorates aging of intestinal stem cells. Cell Rep. 2017, 18, 2608–2621. [Google Scholar] [CrossRef]
- Suzuki, T.; Aoki, K.; Shimokobe, K.; Omiya, S.; Funayama, C.; Takahashi, T.; Kato, M. Age-related morphological and functional changes in the small intestine of senescence-accelerated mouse. Exp. Gerontol. 2022, 163, 111795. [Google Scholar] [CrossRef] [PubMed]
- Varljen, J.; Detel, D.; Batičić, L.; Eraković, V.; Štrbo, N.; Ćuk, M.; Milin, Č. Age dependent activity of brush-border enzymes in BALB/c mice. Croat. Chem. Acta 2005, 78, 379–384. [Google Scholar]
- Xian, Y.; Da, P.; Chao, Y.; Hui, X.; Ligang, Y.; Shaokang, W.; Guiju, S. Wheat oligopeptides enhance the intestinal mucosal barrier and alleviate inflammation via the TLR4/Myd88/MAPK signaling pathway in aged mice. Food Nutr. Res. 2022, 66, 10-29219. [Google Scholar] [CrossRef] [PubMed]
- Jardinaud, F.; Banisadr, G.; Noble, F.; Mélik-Parsadaniantz, S.; Chen, H.; Dugave, C.; Laplace, H.; Rostène, W.; Fournié-Zaluski, M.-C.; Roques, B.P. Ontogenic and adult whole body distribution of aminopeptidase N in rat investigated by in vitro autoradiography. Biochimie 2004, 86, 105–113. [Google Scholar] [CrossRef]
- Milan, A.M.; Cameron-Smith, D. Digestion and postprandial metabolism in the elderly. Adv. Food Nutr. Res. 2015, 76, 79–124. [Google Scholar]
- Thoreux, K.; Balas, D.; Bouley, C.; Senegas-Balas, F. Diet supplemented with yoghurt or milk fermented by Lactobacillus casei DN-114 001 stimulates growth and brush-border enzyme activities in mouse small intestine. Digestion 1998, 59, 349–359. [Google Scholar] [CrossRef]
- Makarova, K.S.; Koonin, E.V. Evolutionary genomics of lactic acid bacteria. J. Bacteriol. 2007, 189, 1199–1208. [Google Scholar] [CrossRef]
- Yang, H.; Singh, M.; Kim, S.J.; Schaefer, J. Characterization of the tertiary structure of the peptidoglycan of Enterococcus faecalis. Biochim. Biophys. Acta (BBA)-Biomembr. 2017, 1859, 2171–2180. [Google Scholar] [CrossRef]
- Kleerebezem, M.; Hols, P.; Bernard, E.; Rolain, T.; Zhou, M.; Siezen, R.J.; Bron, P.A. The extracellular biology of the lactobacilli. FEMS Microbiol. Rev. 2010, 34, 199–230. [Google Scholar] [CrossRef]
- Billot-Klein, D.; Gutmann, L.; Sablé, S.; Guittet, E.; Van Heijenoort, J. Modification of peptidoglycan precursors is a common feature of the low-level vancomycin-resistant VANB-type Enterococcus D366 and of the naturally glycopeptide-resistant species Lactobacillus casei, Pediococcus pentosaceus, Leuconostoc mesenteroides, and. J. Bacteriol. 1994, 176, 2398–2405. [Google Scholar] [CrossRef]
- Castellino, P.; Luzi, L.; Simonson, D.C.; Haymond, M.; Defronzo, R.A. Effect of insulin and plasma amino acid concentrations on leucine metabolism in man. Role of substrate availability on estimates of whole body protein synthesis. J. Clin. Investig. 1987, 80, 1784–1793. [Google Scholar] [CrossRef] [PubMed]
- Islamoglu, A.H.; Garipagaoglu, M.; Bicer, H.S.; Kurtulus, D.; Ozturk, M.; Gunes, F.E. The effects of dietary changes on bone markers in postmenopausal vertebral osteopenia. Clin. Nutr. 2020, 39, 3744–3749. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Shin, C.S.; Yoon, J.-W.; Jeon, S.-M.; Bang, J.; Ku, S.K.; Jegal, K.H. 2′-Fucosyllactose improved muscle health in middle-aged mice performing forced swimming exercise. J. Funct. Foods 2024, 123, 106567. [Google Scholar] [CrossRef]
- Wackerhage, H.; Schoenfeld, B.J.; Hamilton, D.L.; Lehti, M.; Hulmi, J.J. Stimuli and sensors that initiate skeletal muscle hypertrophy following resistance exercise. J. Appl. Physiol. 2019, 126, 30–43. [Google Scholar] [CrossRef]
- Callewaert, L.; Michiels, C.W. Lysozymes in the animal kingdom. J. Biosci. 2010, 35, 127–160. [Google Scholar] [CrossRef]
- Sathiyaseelan, A.; Wang, M.-H. Preparation and characterization of dopamine-loaded bacterial cargo for anti-inflammatory applications. J. Drug Deliv. Sci. Technol. 2023, 89, 105079. [Google Scholar] [CrossRef]
- Shahbazi, R.; Akbari, M.E.; Hashemian, M.; Abbasi, M.; Jalali, S.; Homayounfar, R.; Davoodi, S.H. High body mass index and young age are not associated with post-mastectomy pain syndrome in breast cancer survivors: A case-control study. Iran. J. Cancer Prev. 2015, 8, 29. [Google Scholar]
- Melton III, L.J.; Chrischilles, E.A.; Cooper, C.; Lane, A.W.; Riggs, B.L. Perspective how many women have osteoporosis? J. Bone Miner. Res. 1992, 7, 1005–1010. [Google Scholar] [CrossRef]
- Zhang, Y.-W.; Cao, M.-M.; Li, Y.-J.; Dai, G.-C.; Lu, P.-P.; Zhang, M.; Bai, L.-Y.; Chen, X.-X.; Shi, L.; Zhang, C. Dietary protein intake in relation to the risk of osteoporosis in middle-aged and older individuals: A cross-sectional study. J. Nutr. Health Aging 2022, 26, 252–258. [Google Scholar] [CrossRef]
- Blais, A.; Rochefort, G.Y.; Moreau, M.; Calvez, J.; Wu, X.; Matsumoto, H.; Blachier, F. Monosodium Glutamate Supplementation Improves Bone Status in Mice Under Moderate Protein Restriction. JBMR Plus 2019, 3, e10224. [Google Scholar] [CrossRef]
- Panpetch, W.; Chancharoenthana, W.; Bootdee, K.; Nilgate, S.; Finkelman, M.; Tumwasorn, S.; Leelahavanichkul, A. Lactobacillus rhamnosus L34 Attenuates Gut Translocation-Induced Bacterial Sepsis in Murine Models of Leaky Gut. Infect. Immun. 2018, 86, 10-1128. [Google Scholar] [CrossRef] [PubMed]
- Sashihara, T.; Sueki, N.; Ikegami, S. An Analysis of the Effectiveness of Heat-Killed Lactic Acid Bacteria in Alleviating Allergic Diseases. J. Dairy Sci. 2006, 89, 2846–2855. [Google Scholar] [CrossRef] [PubMed]
- Sonoyama, K.; Kiriyama, S.; Niki, R. Effect of dietary protein level on intestinal aminopeptidase activity and mRNA level in rats. J. Nutr. Biochem. 1994, 5, 291–297. [Google Scholar] [CrossRef]
- Quaroni, A.; Wands, J.; Trelstad, R.L.; Isselbacher, K.J. Epithelioid cell cultures from rat small intestine. Characterization by morphologic and immunologic criteria. J. Cell Biol. 1979, 80, 248–265. [Google Scholar] [CrossRef]
- Murray, R.G.E.; Raymond, N.; Doetsch, R.N.; Robinow, C.F. Determinative and cytological light microscopy. In Methods for General and Molecular Bacteriology; American Society for Microbiology: Washington, DC, USA, 1994; pp. 21–41. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| YC | Aged Mice | ||||
|---|---|---|---|---|---|
| AC | EF | LM | LP | ||
| Final body weight (g) | 41.17 ± 1.42 | 46.68 ± 3.23 | 45.63 ± 2.40 | 46.87 ± 2.85 | 45.48 ± 1.94 |
| Gastrocnemius muscle | |||||
| Weight (g) | 0.18 ± 0.00 | 0.15 ± 0.03 | 0.18 ± 0.00 | 0.17 ± 0.01 | 0.16 ± 0.01 |
| Ratio in BW (%) | 0.44 ± 0.01 a | 0.31 ± 0.06 b | 0.40 ± 0.02 ab | 0.36 ± 0.01 b | 0.35 ± 0.02 b |
| Epididymal adipose tissue | |||||
| Weight (g) | 1.07 ± 0.07 | 1.25 ± 0.20 | 1.38 ± 0.15 | 1.23 ± 0.15 | 1.30 ± 0.07 |
| Ratio in BW (%) | 2.61 ± 0.14 | 2.60 ± 0.31 | 3.01 ± 0.28 | 2.58 ± 0.24 | 2.88 ± 0.20 |
| Feed intake (g/mouse/day) | 2.26 ± 0.19 | 2.14 ± 0.23 | 2.27 ± 0.17 | 2.56 ± 0.24 | 2.51 ± 0.21 |
| Ingredients (g/kg Diet) | Control Diet | LAB-Supplemented Diet |
|---|---|---|
| Maize starch | 519.486 | 517.486 |
| α- Maize starch | 10 | 10 |
| Sucrose | 100 | 100 |
| Casein | 200 | 200 |
| Soybean oil | 70 | 70 |
| Cellulose | 50 | 50 |
| AIN-93 Vitamin mix | 10 | 10 |
| AIN-93 Mineral mix | 35 | 35 |
| L-Cystine | 3 | 3 |
| Choline bitartrate | 2.5 | 2.5 |
| Tert-butylhydroquinone | 0.014 | 0.014 |
| Heat-killed LAB | - | 2 |
| Total | 1000 | 1000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsuruta, T.; Wakisaka, M.; Watanabe, T.; Nishijima, A.; Ikeda, A.; Teraoka, M.; Wang, T.; Chen, K.; Nishino, N. Specific Heat-Killed Lactic Acid Bacteria Enhance Mucosal Aminopeptidase N Activity in the Small Intestine of Aged Mice. Int. J. Mol. Sci. 2025, 26, 5742. https://doi.org/10.3390/ijms26125742
Tsuruta T, Wakisaka M, Watanabe T, Nishijima A, Ikeda A, Teraoka M, Wang T, Chen K, Nishino N. Specific Heat-Killed Lactic Acid Bacteria Enhance Mucosal Aminopeptidase N Activity in the Small Intestine of Aged Mice. International Journal of Molecular Sciences. 2025; 26(12):5742. https://doi.org/10.3390/ijms26125742
Chicago/Turabian StyleTsuruta, Takeshi, Mami Wakisaka, Takumi Watanabe, Aoi Nishijima, Akihito Ikeda, Mao Teraoka, Tianyang Wang, Kuiyi Chen, and Naoki Nishino. 2025. "Specific Heat-Killed Lactic Acid Bacteria Enhance Mucosal Aminopeptidase N Activity in the Small Intestine of Aged Mice" International Journal of Molecular Sciences 26, no. 12: 5742. https://doi.org/10.3390/ijms26125742
APA StyleTsuruta, T., Wakisaka, M., Watanabe, T., Nishijima, A., Ikeda, A., Teraoka, M., Wang, T., Chen, K., & Nishino, N. (2025). Specific Heat-Killed Lactic Acid Bacteria Enhance Mucosal Aminopeptidase N Activity in the Small Intestine of Aged Mice. International Journal of Molecular Sciences, 26(12), 5742. https://doi.org/10.3390/ijms26125742