
Transcriptome Analysis Reveals Mechanisms of Stripe Rust Response in Wheat Cultivar Anmai1350

Abstract
1. Introduction
2. Results
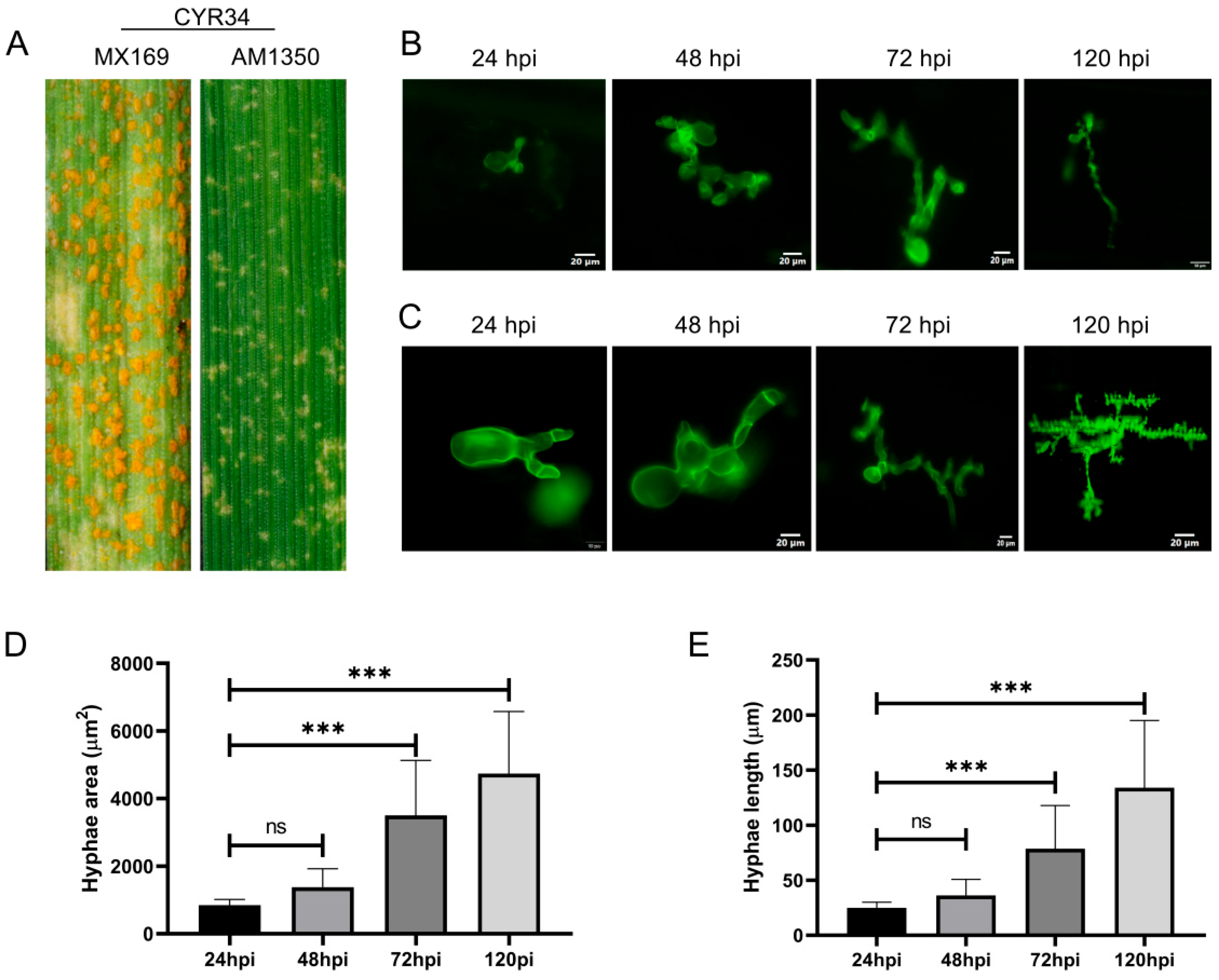
2.1. Changes in Mycelial Length and Area During the Infection Process
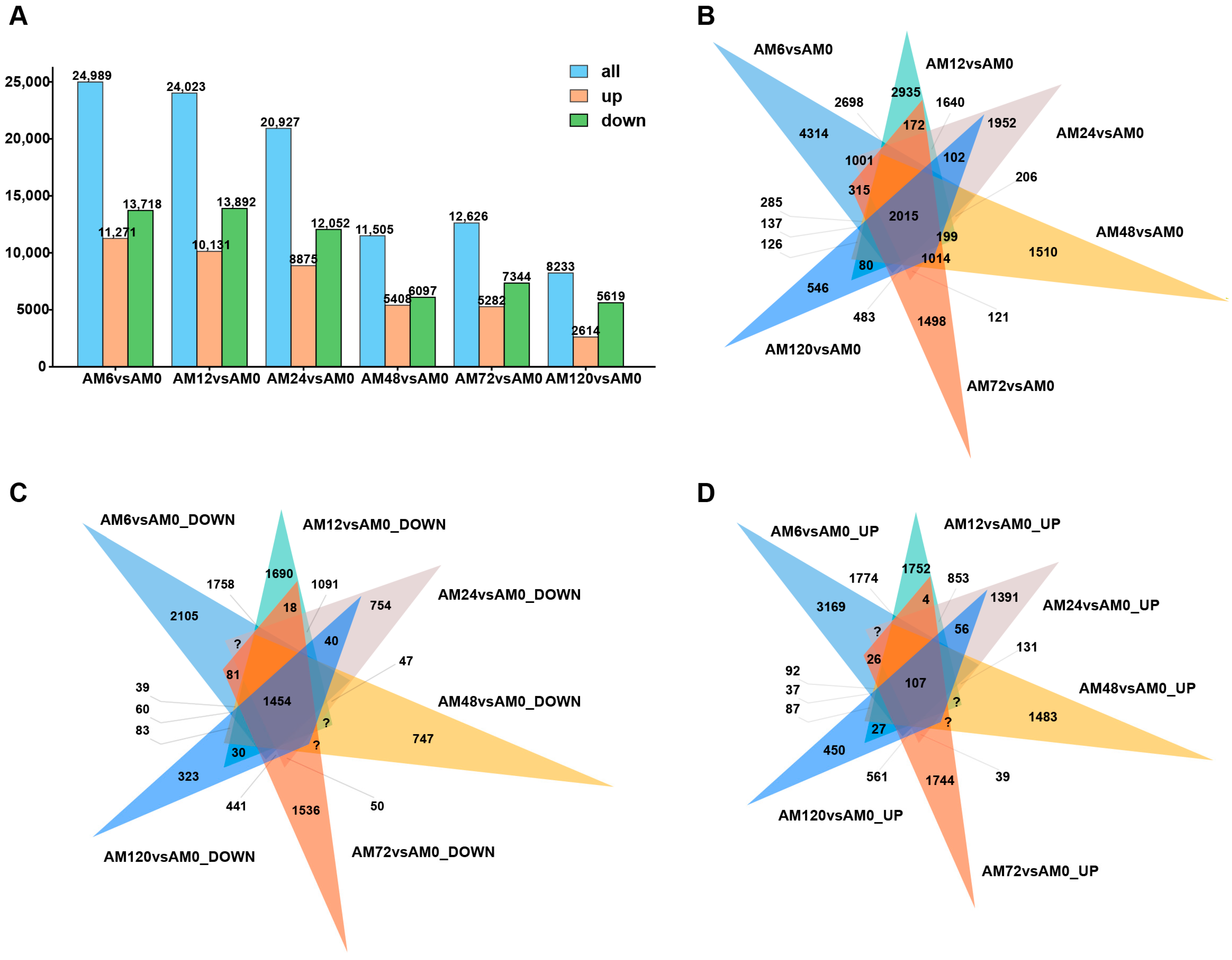
2.2. Statistical Analysis of RNA-seq Results from Different Time Points After Pst Inoculation
2.3. Characterization of DEGs in Response to Pst Infection
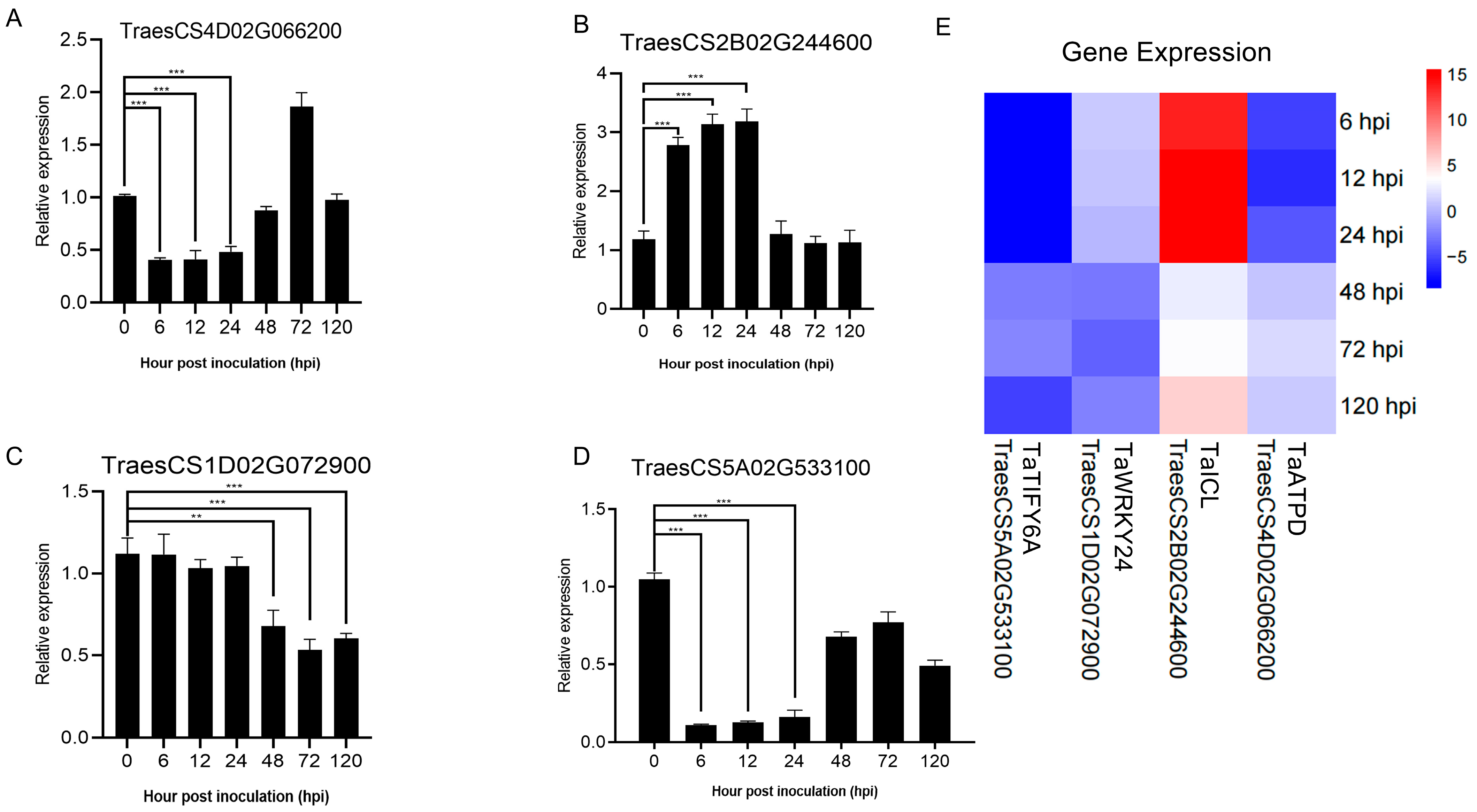
2.4. Validation of RNA-seq Data by qRT-PCR
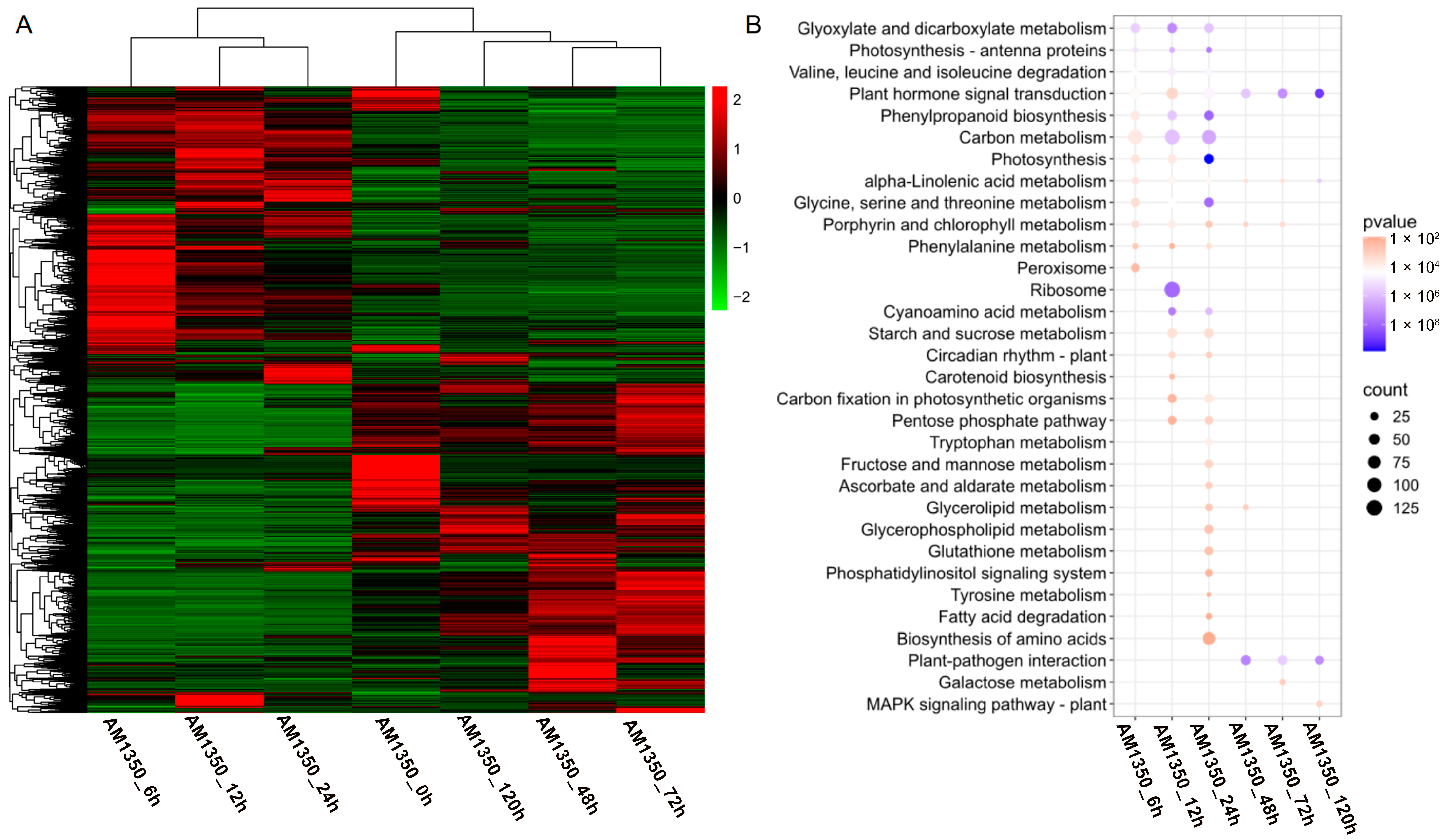
2.5. Cluster Heatmap Group and KEGG Enrichment Analyses of DEGs
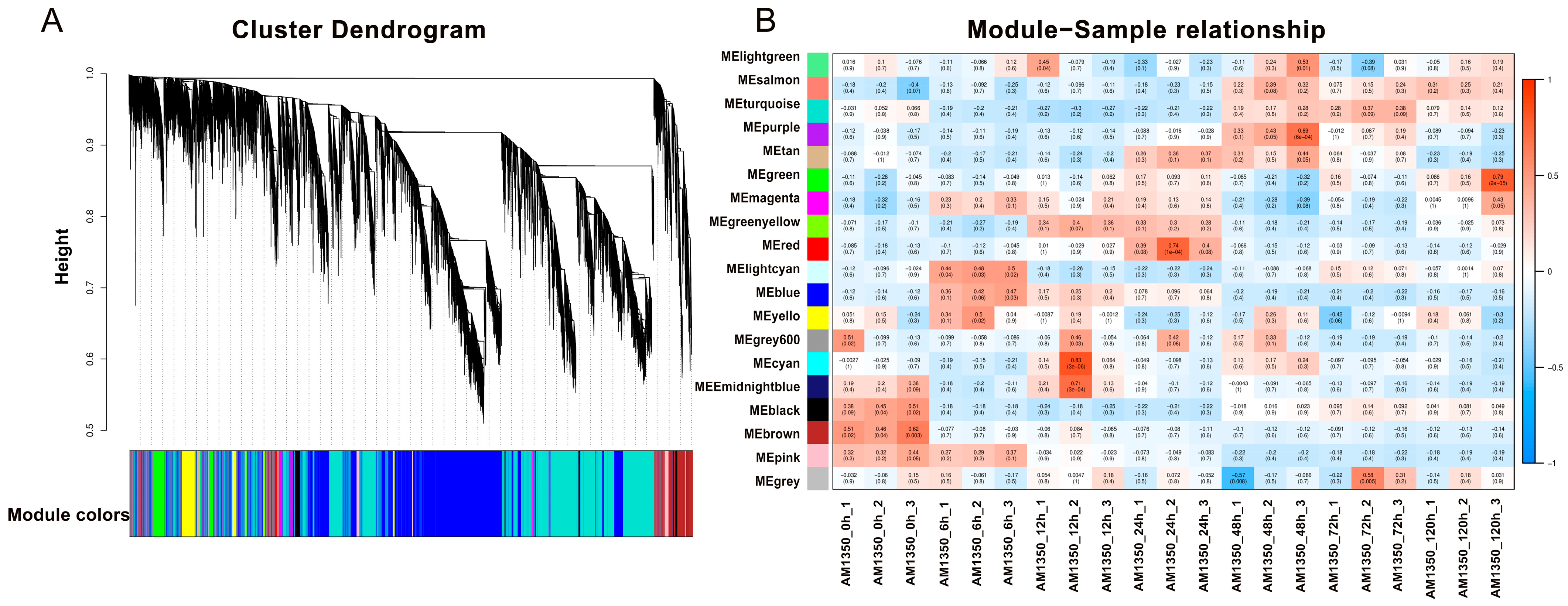
2.6. Weighted Gene Correlation Network Analysis
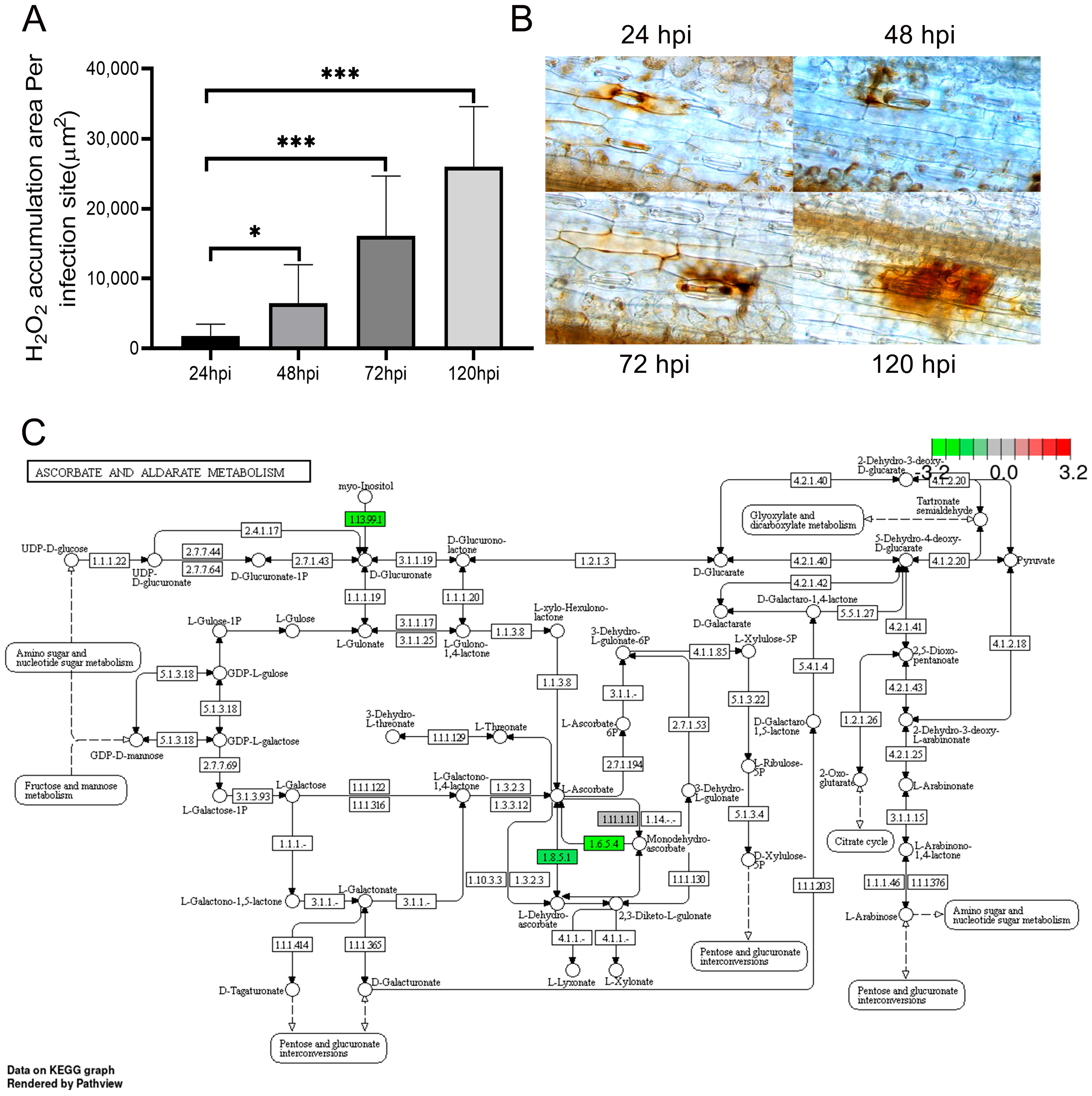
2.7. The Accumulation of ROS
2.8. Metabolic Network Analysis
3. Discussion
3.1. Upregulation and Downregulation of DEGs
3.2. The Role of ROS in the Resistance of AM1350 to Pst
3.3. Analysis of the Reasons for the Changes in Mycelial Length and Area
3.4. Plant–Pathogen Interactions
4. Materials and Methods
4.1. Plant Materials and Pst Inoculation
4.2. RNA Extraction, Library Construction and Sequencing
4.3. Differential Expression Analyses of Genes
4.4. GO Terms and KEGG Enrichment Analyses of DEGs
4.5. Identification and Analysis of Transcription Factors
4.6. Diaminobenzidine Staining and Observation of Samples
4.7. Quantitative Real-Time PCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Reynolds, M.; Foulkes, J.; Furbank, R.; Griffiths, S.; King, J.; Murchie, E.; Parry, M.; Slafer, G. Achieving Yield Gains in Wheat. Plant Cell Environ. 2012, 35, 1799–1823. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Li, P.; Liu, Z.; Cui, Y.; Yao, Z.; Chen, W.; Wang, M.; Yu, C.; Xia, S.; Sun, Y.; et al. Exploring the Molecular Mechanisms by Which Secretory Phospholipase A2 Regulates Lymphatic Endothelial Cell Dysfunction by Activating Macrophages. Int. J. Biol. Macromol. 2025, 294, 139038. [Google Scholar] [CrossRef] [PubMed]
- Schwessinger, B. Fundamental Wheat Stripe Rust Research in the 21st Century. New Phytol. 2017, 213, 1625–1631. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, R.; Mu, J.; Han, D.; Kang, Z. Wheat Stripe Rust Resistance Gene Yr24/Yr26: A Retrospective Review. Crop J. 2018, 6, 321–329. [Google Scholar] [CrossRef]
- Du, Z.; Liu, M.; Sun, M.; Ma, X.; Wang, L.; Kang, Z.; Zhao, J. Virulence and Molecular Characterization Reveal Signs of Sexual Genetic Recombination of Puccinia Striiformis f.sp. Tritici and Puccinia Striiformis f.sp. Hordei in Tibet. Plant Dis. 2024, 108, 2341–2353. [Google Scholar] [CrossRef]
- Wang, L.; Tang, X.; Wu, J.; Shen, C.; Dai, M.; Wang, Q.; Zeng, Q.; Kang, Z.; Wu, Y.; Han, D. Stripe Rust Resistance to a Burgeoning Puccinia Striiformis f.sp. Tritici Race CYR34 in Current Chinese Wheat Cultivars for Breeding and Research. Euphytica 2019, 215, 68. [Google Scholar] [CrossRef]
- Liu, R.; Lu, J.; Zhang, L.; Wu, Y. Transcriptomic Insights into the Molecular Mechanism of Wheat Response to Stripe Rust Fungus. Heliyon 2022, 8, e10951. [Google Scholar] [CrossRef]
- Zhang, G.; Liu, W.; Wang, L.; Cheng, X.; Tian, X.; Du, Z.; Kang, Z.; Zhao, J. Evaluation of the Potential Risk of the Emerging Yr5-Virulent Races of Puccinia striiformis f.sp. Tritici to 165 Chinese Wheat Cultivars. Plant Dis. 2022, 106, 1867–1874. [Google Scholar] [CrossRef]
- Shendure, J. The Beginning of the End for Microarrays? Nat. Methods 2008, 5, 585–587. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, Y.; Wang, C.; Liu, M.; Li, H.; Fu, Y.; Wang, Y.; Nie, Y.; Liu, X.; Ji, W. Large-Scale Transcriptome Comparison Reveals Distinct Gene Activations in Wheat Responding to Stripe Rust and Powdery Mildew. BMC Genom. 2014, 15, 898. [Google Scholar] [CrossRef]
- Yadav, I.S.; Sharma, A.; Kaur, S.; Nahar, N.; Bhardwaj, S.C.; Sharma, T.R.; Chhuneja, P. Comparative Temporal Transcriptome Profiling of Wheat near Isogenic Line Carrying Lr 57 under Compatible and Incompatible Interactions. Front. Plant Sci. 2016, 7, 1943. [Google Scholar] [CrossRef]
- Dobon, A.; Bunting, D.C.E.; Cabrera-Quio, L.E.; Uauy, C.; Saunders, D.G.O. The Host-Pathogen Interaction between Wheat and Yellow Rust Induces Temporally Coordinated Waves of Gene Expression. BMC Genom. 2016, 17, 380. [Google Scholar] [CrossRef]
- Wang, Z.; Gerstein, M.; Snyder, M. Rna-Seq: A Revolutionary Tool for Transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef]
- Liu, H.; Wang, Z.; Tian, X. The Role of Photosynthesis in Plant-Pathogen Interactions. Plant Health Med. 2024, 3, 32–41. [Google Scholar]
- Chang, Q.; Liu, J.; Wang, Q.; Han, L.; Liu, J.; Li, M.; Huang, L.; Yang, J.; Kang, Z. The Effect of Puccinia striiformis f.sp. Tritici on the Levels of Water-Soluble Carbohydrates and the Photosynthetic Rate in Wheat Leaves. Physiol. Mol. Plant Pathol. 2013, 84, 131–137. [Google Scholar] [CrossRef]
- Chen, Y.-E.; Cui, J.-M.; Su, Y.-Q.; Yuan, S.; Yuan, M.; Zhang, H.-Y. Influence of Stripe Rust Infection on the Photosynthetic Characteristics and Antioxidant System of Susceptible and Resistant Wheat Cultivars at the Adult Plant Stage. Front. Plant Sci. 2015, 6, 779. [Google Scholar] [CrossRef] [PubMed]
- Zainy, Z.; Fayyaz, M.; Yasmin, T.; Hyder, M.Z.; Haider, W.; Farrakh, S. Antioxidant Enzymes Activity and Gene Expression in Wheat-Stripe Rust Interaction at Seedling Stage. Physiol. Mol. Plant Pathol. 2023, 124, 101960. [Google Scholar] [CrossRef]
- Zheng, S.; Li, Y.; Lu, L.; Liu, Z.; Zhang, C.; Ao, D.; Li, L.; Zhang, C.; Liu, R.; Luo, C.; et al. Evaluating the Contribution of Yr Genes to Stripe Rust Resistance Breeding through Marker-Assisted Detection in Wheat. Euphytica 2017, 213, 50. [Google Scholar] [CrossRef]
- Chen, Y.; Mao, H.; Wu, N.; Ma, J.; Yuan, M.; Zhang, Z.; Yuan, S.; Zhang, H. Effects of Stripe Rust Infection on the Levels of Redox Balance and Photosynthetic Capacities in Wheat. Int. J. Mol. Sci. 2019, 21, 268. [Google Scholar] [CrossRef]
- Kumar, D.; Sharma, S.; Sharma, R.; Pundir, S.; Singh, V.K.; Chaturvedi, D.; Singh, B.; Kumar, S.; Sharma, S. Genome-Wide Association Study in Hexaploid Wheat Identifies Novel Genomic Regions Associated with Resistance to Root Lesion Nematode (Pratylenchus thornei). Sci. Rep. 2021, 11, 3572. [Google Scholar] [CrossRef]
- Zhang, R.; Liu, Z.; Zhao, S.; Zhao, X.; Wang, S.; Li, X.; Lin, D.; Li, C.; Xiao, J.; Wang, X.; et al. TaEF1a Is Involved in Low Phosphorus Stress Responses and Affects Root Development. Plant Growth Regul. 2024, 102, 227–236. [Google Scholar] [CrossRef]
- Liu, N.; Li, Q.; Shan, Q. Combined Prokaryotic Transcriptomics and Proteomics Analysis of Clinical Trueperella Pyogenes Isolates with Distinctive Cytotoxicity. Int. J. Mol. Sci. 2025, 26, 1490. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Chen, L.; Liu, J.; Zhou, B.; Sha, Y.; Hu, G.; Peng, J. Transcriptomic Insights into the Molecular Mechanism for Response of Wild Emmer Wheat to Stripe Rust Fungus. Front. Plant Sci. 2024, 14, 1320976. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Cao, F.; Gao, Q.; Liang, K.; Tang, Y. Improving Diagnosis Accuracy of Non-Small Cell Lung Carcinoma on Noisy Data by Adaptive Group Lasso Regularized Multinomial Regression. Biomed. Signal Process. Control 2023, 79, 104148. [Google Scholar] [CrossRef]
- Qi, F.; Li, J.; Ai, Y.; Shangguan, K.; Li, P.; Lin, F.; Liang, Y. Dgk5β-Derived Phosphatidic Acid Regulates Ros Production in Plant Immunity by Stabilizing Nadph Oxidase. Cell Host Microbe 2024, 32, 425–440.e7. [Google Scholar] [CrossRef]
- Wang, N.; Fan, X.; He, M.; Hu, Z.; Tang, C.; Zhang, S.; Lin, D.; Gan, P.; Wang, J.; Huang, X.; et al. Transcriptional Repression of Tanox10 by Tawrky19 Compromises Ros Generation and Enhances Wheat Susceptibility to Stripe Rust. Plant Cell 2022, 34, 1784–1803. [Google Scholar] [CrossRef]
- Wang, C.-F.; Huang, L.-L.; Buchenauer, H.; Han, Q.-M.; Zhang, H.-C.; Kang, Z.-S. Histochemical Studies on the Accumulation of Reactive Oxygen Species (O2− and H2O2) in the Incompatible and Compatible Interaction of Wheat—Puccinia striiformis f.sp. tritici. Physiol. Mol. Plant Pathol. 2007, 71, 230–239. [Google Scholar] [CrossRef]
- Yang, X.; Guo, T.; Du, Z.; Qin, X.; Wang, K.; Kebreab, E.; Wang, D.; Lyu, L. Protective Effects of Mnq against Lipopolysaccharide-Induced Inflammatory Damage in Bovine Ovarian Follicular Granulosa Cells in Vitro. J. Steroid Biochem. Mol. Biol. 2023, 230, 106274. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Kawashima, M.; Ishiguro-Watanabe, M. Kegg for Taxonomy-Based Analysis of Pathways and Genomes. Nucleic Acids Res. 2023, 51, D587–D592. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. Kegg: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Njus, D.; Kelley, P.M.; Tu, Y.-J.; Schlegel, H.B. Ascorbic Acid: The Chemistry Underlying Its Antioxidant Properties. Free Radic. Biol. Med. 2020, 159, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Arrigoni, O.; Tullio, M.C.D. Ascorbic Acid: Much More Than Just an Antioxidant. Biochim. Et Biophys. Acta (BBA)-Gen. Subj. 2002, 1569, 1–9. [Google Scholar] [CrossRef]
- Ivanov, K.; Blöchl, E.C.; Tenhaken, R. The Myo-Inositol Pathway Does Not Contribute to Ascorbic Acid Synthesis. Plant Biol. 2019, 21, 95–102. [Google Scholar] [CrossRef]
- Munir, S.; Mumtaz, M.A.; Ahiakpa, J.K.; Liu, G.; Chen, W.; Zhou, G.; Zheng, W.; Ye, Z.; Zhang, Y. Genome-Wide Analysis of Myo-Inositol Oxygenase Gene Family in Tomato Reveals Their Involvement in Ascorbic Acid Accumulation. BMC Genom. 2020, 21, 284. [Google Scholar] [CrossRef]
- Hong, G.; Zhang, W.; Li, H.; Shen, X.; Guo, Z. Separate Enrichment Analysis of Pathways for up-and Downregulated Genes. J. R. Soc. Interface 2014, 11, 20130950. [Google Scholar] [CrossRef]
- Sakai, Y.; Honda, M.; Fujinaga, H.; Tatsumi, I.; Mizukoshi, E.; Nakamoto, Y.; Kaneko, S. Common Transcriptional Signature of Tumor-Infiltrating Mononuclear Inflammatory Cells and Peripheral Blood Mononuclear Cells in Hepatocellular Carcinoma Patients. Cancer Res. 2008, 68, 10267–10279. [Google Scholar] [CrossRef] [PubMed]
- Draghici, S.; Khatri, P.; Tarca, A.L.; Amin, K.; Done, A.; Voichita, C.; Georgescu, C.; Romero, R. A Systems Biology Approach for Pathway Level Analysis. Genome Res. 2007, 17, 1537–1545. [Google Scholar] [CrossRef] [PubMed]
- Doke, N. Involvement of Superoxide Anion Generation in the Hypersensitive Response of Potato Tuber Tissues to Infection with an Incompatible Race of Phytophthora Infestans and to the Hyphal Wall Components. Physiol. Plant Pathol. 1983, 23, 345–357. [Google Scholar] [CrossRef]
- Segal, A.W.; Abo, A. The Biochemical Basis of the Nadph Oxidase of Phagocytes. Trends Biochem. Sci. 1993, 18, 43–47. [Google Scholar] [CrossRef]
- Yin, X.; Gao, Y.; Song, S.; Hassani, D.; Lu, J. Identification, Characterization and Functional Analysis of Grape (Vitis vinifera L.) Mitochondrial Transcription Termination Factor (Mterf) Genes in Responding to Biotic Stress and Exogenous Phytohormone. BMC Genom. 2021, 22, 136. [Google Scholar] [CrossRef]
- Wang, P.; Liu, W.; Han, C.; Wang, S.; Bai, M.; Song, C. Reactive Oxygen Species: Multidimensional Regulators of Plant Adaptation to Abiotic Stress and Development. J. Integr. Plant Biol. 2024, 66, 330–367. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, S.; Serrano, M.; L’Haridon, F.; Tjamos, S.E.; Métraux, J.-P. Reactive Oxygen Species and Plant Resistance to Fungal Pathogens. Phytochemistry 2015, 112, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhao, Y.; Zhang, Y.; Niu, L.; Li, W.; Lu, W.; Li, J.; Schäfer, P.; Meng, Y.; Shan, W. A Mitochondrial Rna Processing Protein Mediates Plant Immunity to a Broad Spectrum of Pathogens by Modulating the Mitochondrial Oxidative Burst. Plant Cell 2022, 34, 2343–2363. [Google Scholar] [CrossRef]
- Fang, N.; Jia, C.; Chen, R.; An, J.; Kang, Z.; Liu, J. The Wheat Cc-Nbs-Lrr Protein Targa3 Confers Resistance to Stripe Rust by Suppressing Ascorbate Peroxidase 6 Activity. Plant Physiol. 2025, 197, kiae603. [Google Scholar] [CrossRef]
- Wojtaszek, P. Mechanisms for the Generation of Reactive Oxygen Species in Plant Defence Response. Acta Physiol. Plant. 1997, 19, 581–589. [Google Scholar] [CrossRef]
- Liu, X.; Williams, C.E.; Nemacheck, J.A.; Wang, H.; Subramanyam, S.; Zheng, C.; Chen, M.-S. Reactive Oxygen Species Are Involved in Plant Defense against a Gall Midge. Plant Physiol. 2010, 152, 985–999. [Google Scholar] [CrossRef]
- Ma, X.; Zhang, Z.; Deng, R.; Liu, N.; Jiang, H.; Kang, Z.; Liu, J. Secreted Xylanase Pstxyn1 Contributes to Stripe Rust Infection Possibly by Overcoming Cell Wall Barrier and Suppressing Defense Responses in Wheat. J. Agric. Food Chem. 2024, 73, 380–392. [Google Scholar] [CrossRef]
- Enkerli, K.; Mims, C.W.; Hahn, M.G. Ultrastructure of Compatible and Incompatible Interactions of Soybean Roots Infected with the Plant Pathogenic Oomycete Phytophthora Sojae. Can. J. Bot. 1997, 75, 1493–1508. [Google Scholar] [CrossRef]
- Chen, H.-L.; Wang, Y.-Q.; Chu, C.-C.; Li, P. Plant Non-Host Resistance: Current Progress and Future Prospect. Yi Chuan Hered. 2008, 30, 977–982. [Google Scholar] [CrossRef]
- Dolatabadian, A.; Fernando, W.G.D. Genomic Variations and Mutational Events Associated with Plant–Pathogen Interactions. Biology 2022, 11, 421. [Google Scholar] [CrossRef]
- Duba, A.; Goriewa-Duba, K.; Wachowska, U. A Review of the Interactions between Wheat and Wheat Pathogens: Zymoseptoria tritici, Fusarium Spp. and Parastagonospora nodorum. Int. J. Mol. Sci. 2018, 19, 1138. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Moreno-Pérez, A.; Coaker, G. Understanding Plant Pathogen Interactions Using Spatial and Single-Cell Technologies. Commun. Biol. 2023, 6, 814. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Huang, L.; Luo, W.; Jin, Y.; Gong, F.; He, J.; Liu, D.; Zheng, Y.; Wu, B. Transcriptome Analysis Provides Insights into the Mechanisms Underlying Wheat Cultivar Shumai126 Responding to Stripe Rust. Gene 2021, 768, 145290. [Google Scholar] [CrossRef]
- L’Haridon, F.; Besson-Bard, A.; Binda, M.; Serrano, M.; Abou-Mansour, E.; Balet, F.; Schoonbeek, H.-J.; Hess, S.; Mir, R.; Léon, J.; et al. A Permeable Cuticle Is Associated with the Release of Reactive Oxygen Species and Induction of Innate Immunity. PLoS Pathog. 2011, 7, e1002148. [Google Scholar] [CrossRef]
- Gogoi, K.; Gogoi, H.; Borgohain, M.; Saikia, R.; Chikkaputtaiah, C.; Hiremath, S.; Basu, U. The Molecular Dynamics between Reactive Oxygen Species (ROS), Reactive Nitrogen Species (RNS) and Phytohormones in Plant’s Response to Biotic Stress. Plant Cell Rep. 2024, 43, 263. [Google Scholar] [CrossRef]
- Qin, W.-M.; Lan, W.-Z.; Yang, X. Involvement of Nadph Oxidase in Hydrogen Peroxide Accumulation by Aspergillus Niger Elicitor-Induced Taxus Chinensis Cell Cultures. J. Plant Physiol. 2004, 161, 355–361. [Google Scholar] [CrossRef]
- Wang, Q.; Tang, Y.; Li, Y.; Ren, J.; Zuo, H.; Cheng, P.; Li, Q.; Wang, B. Abscisic Acid-, Stress-, Ripening-Induced 2 Like Protein, Taasr2l, Promotes Wheat Resistance to Stripe Rust. Mol. Plant Pathol. 2024, 25, e70028. [Google Scholar] [CrossRef]
- Bai, X.; Zhan, G.; Tian, S.; Peng, H.; Cui, X.; Islam, M.A.; Goher, F.; Ma, Y.; Kang, Z.; Xu, Z.-S.; et al. Transcription Factor Bzr2 Activates Chitinase Cht20. 2 Transcription to Confer Resistance to Wheat Stripe Rust. Plant Physiol. 2021, 187, 2749–2762. [Google Scholar] [CrossRef]
- Guo, S.; Zhang, F.; Du, X.; Zhang, X.; Huang, X.; Li, Z.; Zhang, Y.; Gan, P.; Li, H.; Li, M.; et al. Taank-Tpr1 Enhances Wheat Resistance against Stripe Rust Via Controlling Gene Expression and Protein Activity of Nlr Protein Tarpp13l1. Dev. Cell 2025. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Li, X.-M.; Chen, Y.; Wu, T.-T.; Ding, C.-H.; Zhang, M.-Q.; Guo, Y.-T.; Wang, C.-Y.; Zhang, J.; Zhang, X.; et al. Phosphorylation of Kat-2b by Wks1/Yr36 Redirects the Lipid Flux to Jasmonates to Enhance Resistance against Wheat Stripe Rust. J. Genet. Genom. 2023, 50, 872–882. [Google Scholar] [CrossRef]
- Mittler, R.; Zandalinas, S.I.; Fichman, Y.; Van Breusegem, F. Reactive Oxygen Species Signalling in Plant Stress Responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 663–679. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhang, S. Mitogen-Activated Protein Kinase Cascades in Plant Signaling. J. Integr. Plant Biol. 2022, 64, 301–341. [Google Scholar] [CrossRef] [PubMed]
- Stewart, P.; Globig, S. Phytopathology in Plants; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene Ontology Analysis for RNA-Seq: Accounting for Selection Bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef]
- Hu, P.; Luo, S.; Qu, G.; Luo, Q.; Tian, Y.; Huang, K.; Sun, T. Identification and Validation of Feature Genes Associated with M1 Macrophages in Preeclampsia. Aging 2023, 15, 13822. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Clean_Bases | Total_Reads | Error_Rate (%) | Q20 (%) | Q30 (%) | GC_pct (%) | Total_Map | Mapping Rate (%) |
|---|---|---|---|---|---|---|---|---|
| AM1350_0h_1 | 6.94G | 46,272,540 | 0.03 | 97.61 | 93.42 | 55.37 | 43,222,298 | 93.41 |
| AM1350_0h_2 | 6.92G | 46,119,588 | 0.03 | 97.54 | 93.22 | 56.27 | 43,144,542 | 93.55 |
| AM1350_0h_3 | 6.68G | 44,541,752 | 0.03 | 97.68 | 93.57 | 53.80 | 41,576,571 | 93.34 |
| AM1350_6h_1 | 6.93G | 46,229,954 | 0.03 | 97.48 | 93.13 | 55.80 | 42,768,413 | 92.51 |
| AM1350_6h_2 | 6.73G | 44,852,278 | 0.03 | 97.56 | 93.29 | 55.82 | 41,577,526 | 92.70 |
| AM1350_6h_3 | 6.89G | 45,964,126 | 0.03 | 97.56 | 93.27 | 54.73 | 42,404,143 | 92.25 |
| AM1350_12h_1 | 6.79G | 45,272,470 | 0.03 | 97.55 | 93.26 | 54.45 | 41,741,226 | 92.20 |
| AM1350_12h_2 | 6.36G | 42,380,874 | 0.03 | 97.48 | 93.16 | 54.81 | 39,218,703 | 92.54 |
| AM1350_12h_3 | 6.18G | 41,192,308 | 0.03 | 97.54 | 93.30 | 54.97 | 38,155,268 | 92.63 |
| AM1350_24h_1 | 6.19G | 41,263,874 | 0.03 | 97.49 | 93.23 | 54.52 | 37,915,463 | 91.89 |
| AM1350_24h_2 | 7.13G | 47,543,002 | 0.03 | 97.64 | 93.53 | 55.62 | 43,969,645 | 92.48 |
| AM1350_24h_3 | 7.44G | 49,590,532 | 0.03 | 97.22 | 92.49 | 54.68 | 45,738,645 | 92.23 |
| AM1350_48h_1 | 7.24G | 48,254,104 | 0.03 | 97.61 | 93.43 | 54.48 | 44,718,819 | 92.67 |
| AM1350_48h_2 | 6.63G | 44,228,828 | 0.03 | 97.46 | 93.08 | 55.82 | 40,993,286 | 92.68 |
| AM1350_48h_3 | 6.56G | 43,727,762 | 0.03 | 97.78 | 93.73 | 56.35 | 40,682,044 | 93.03 |
| AM1350_72h_1 | 6.46G | 43,076,234 | 0.03 | 97.56 | 93.25 | 53.00 | 38,587,764 | 89.58 |
| AM1350_72h_2 | 6.18G | 41,231,804 | 0.03 | 97.74 | 93.69 | 55.43 | 38,759,419 | 94.00 |
| AM1350_72h_3 | 6.43G | 42,840,026 | 0.03 | 97.70 | 93.63 | 55.47 | 40,041,500 | 93.47 |
| AM1350_120h_1 | 6.27G | 41,795,472 | 0.03 | 97.40 | 92.94 | 55.82 | 38,686,028 | 92.56 |
| AM1350_120h_2 | 6.35G | 42,331,432 | 0.03 | 97.65 | 93.49 | 55.76 | 39,374,360 | 93.01 |
| AM1350_120h_3 | 6.89G | 45,923,630 | 0.03 | 97.31 | 92.80 | 53.31 | 40,694,489 | 88.61 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, F.; Zhu, J.; Xue, X.; Chen, H.; Nong, X.; Yang, C.; Shen, W.; Gan, P. Transcriptome Analysis Reveals Mechanisms of Stripe Rust Response in Wheat Cultivar Anmai1350. Int. J. Mol. Sci. 2025, 26, 5538. https://doi.org/10.3390/ijms26125538
Gao F, Zhu J, Xue X, Chen H, Nong X, Yang C, Shen W, Gan P. Transcriptome Analysis Reveals Mechanisms of Stripe Rust Response in Wheat Cultivar Anmai1350. International Journal of Molecular Sciences. 2025; 26(12):5538. https://doi.org/10.3390/ijms26125538
Chicago/Turabian StyleGao, Feng, Jingyi Zhu, Xin Xue, Hongqi Chen, Xiaojin Nong, Chunling Yang, Weimin Shen, and Pengfei Gan. 2025. "Transcriptome Analysis Reveals Mechanisms of Stripe Rust Response in Wheat Cultivar Anmai1350" International Journal of Molecular Sciences 26, no. 12: 5538. https://doi.org/10.3390/ijms26125538
APA StyleGao, F., Zhu, J., Xue, X., Chen, H., Nong, X., Yang, C., Shen, W., & Gan, P. (2025). Transcriptome Analysis Reveals Mechanisms of Stripe Rust Response in Wheat Cultivar Anmai1350. International Journal of Molecular Sciences, 26(12), 5538. https://doi.org/10.3390/ijms26125538