


Sodium–Glucose Cotransporter 2 and Glucose Levels Affect Clear Cell Renal Cell Carcinoma Progression

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
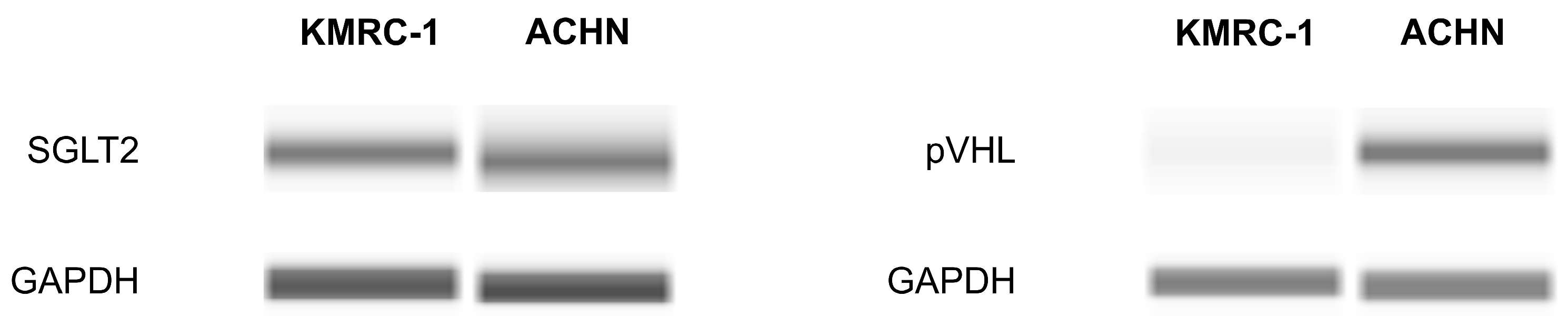
2.1. The Presence of SGLT2 in Human ccRCC Cells
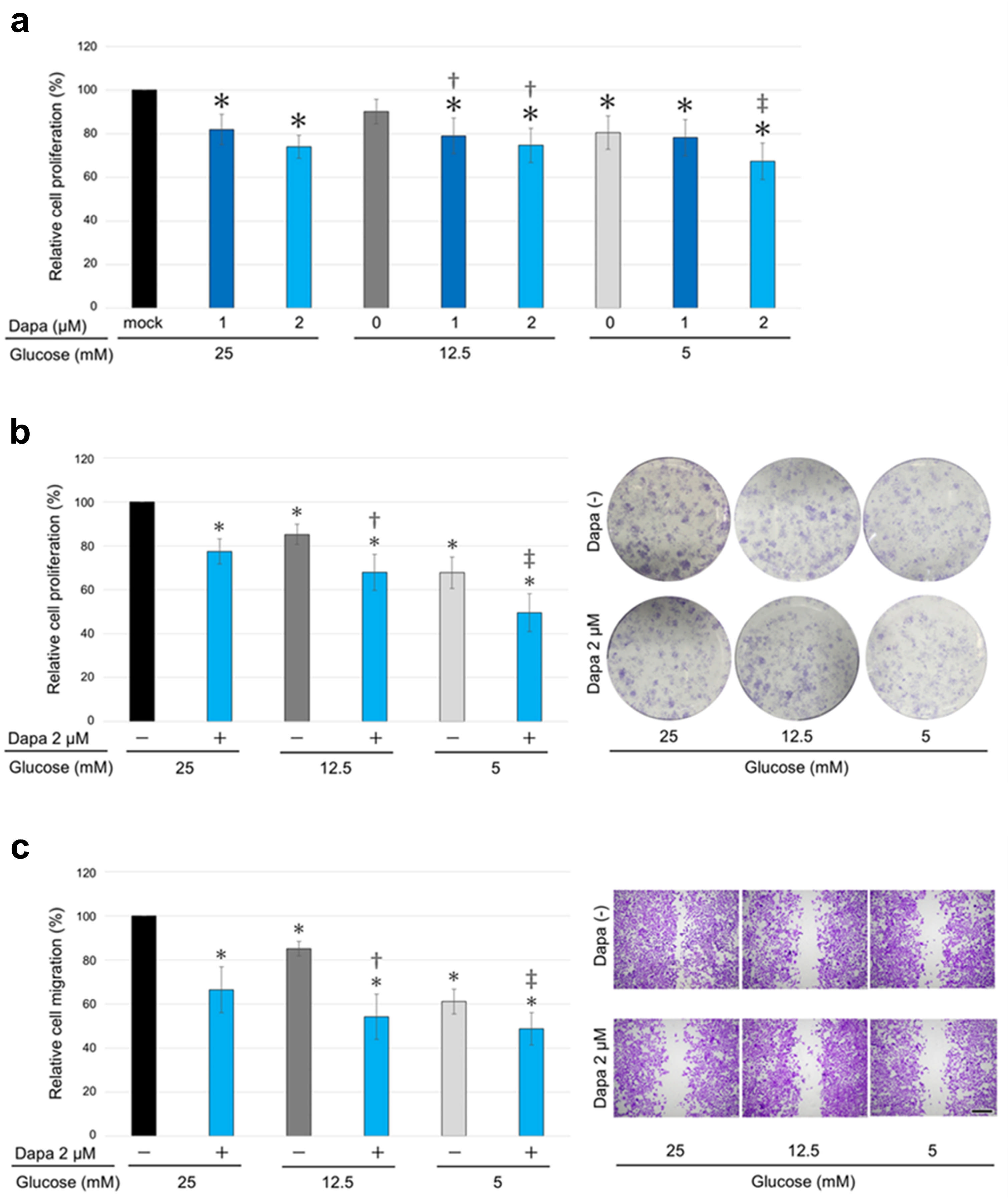
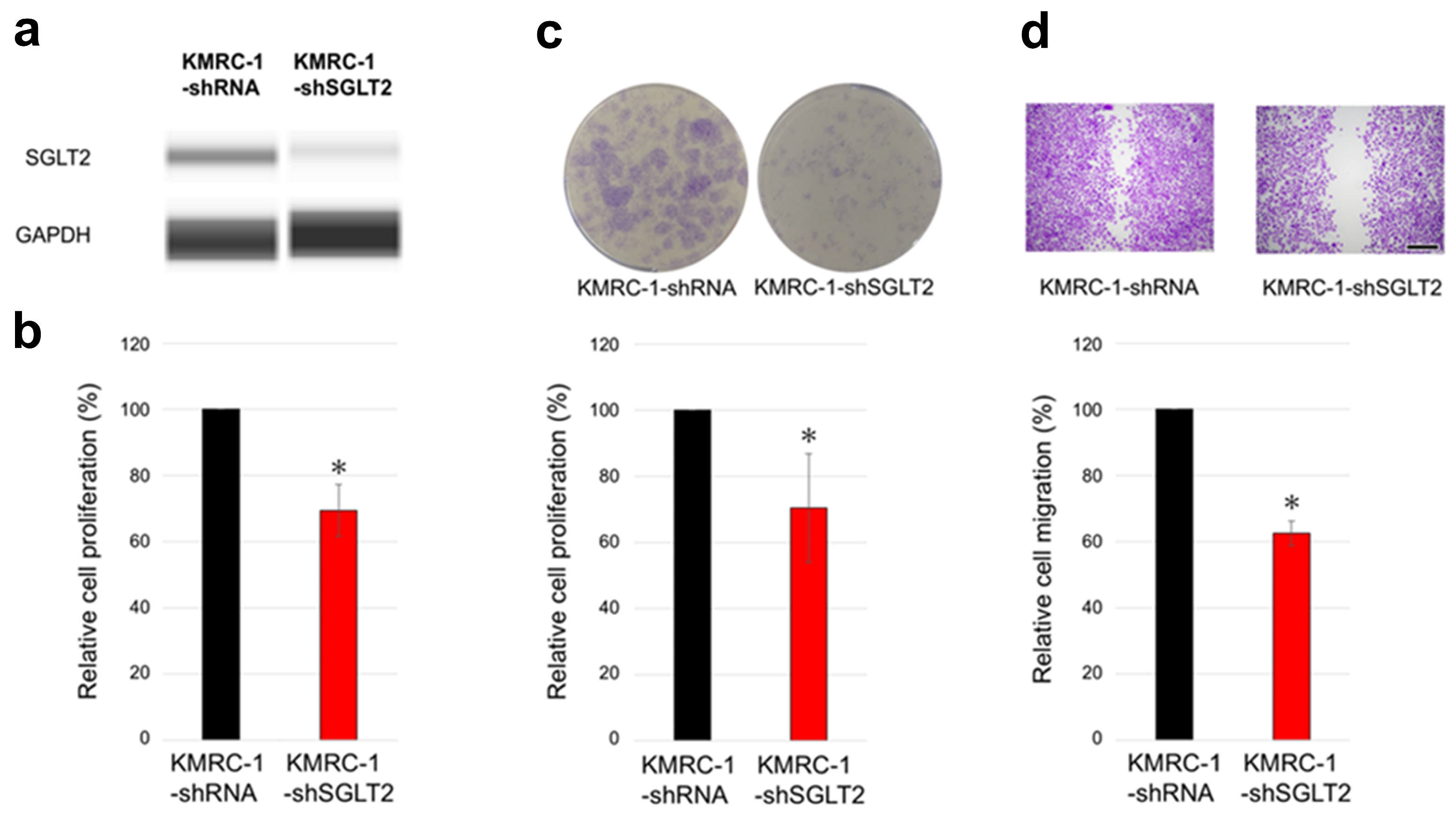
2.2. The Impact of SGLT2 Inactivation and Glucose Concentrations on ccRCC Cell Progression
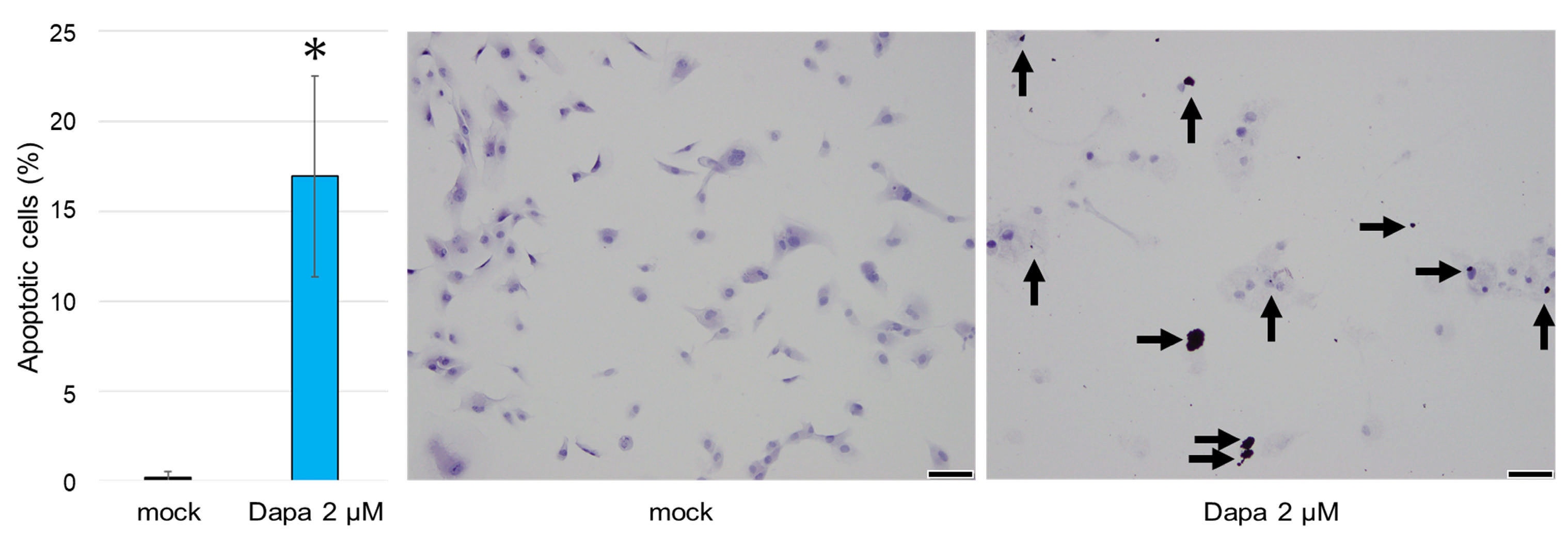
2.3. The Impact of SGLT2 Inhibition on the Apoptosis of ccRCC Cells
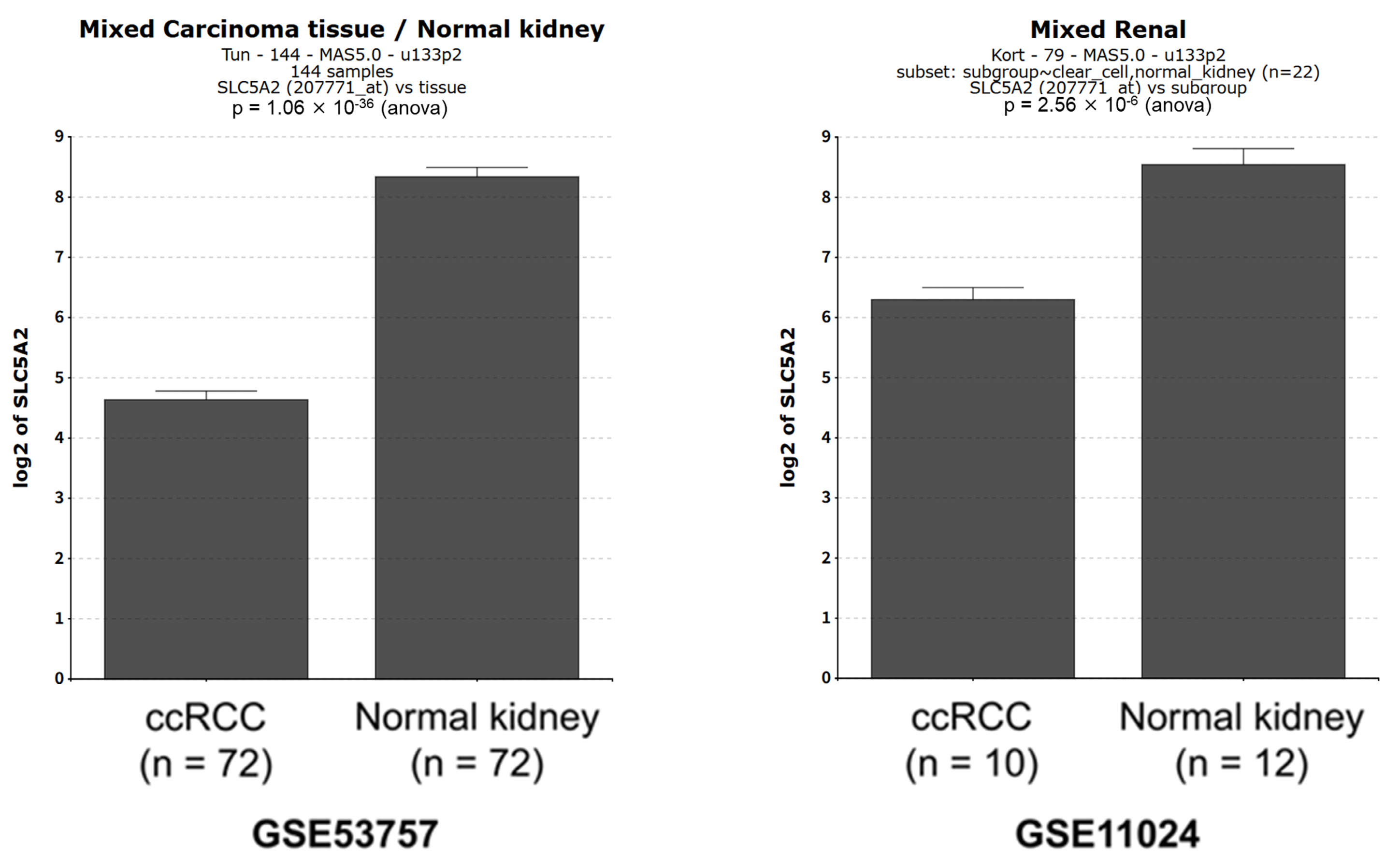
2.4. SLC5A2 Expression in ccRCC Tissues
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Chemicals
4.2. Cell Viability
4.3. Plate Colony Formation
4.4. Cell Migration
4.5. Apoptosis Analysis
4.6. Western Blot
4.7. Public Database Analysis
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SGLT2 | Sodium–glucose cotransporter 2 |
| ccRCC | Clear cell renal cell carcinoma |
| VHL | von Hippel–Lindau |
| SGLT2i | Sodium–glucose cotransporter 2 inhibitor |
| RCC | Renal cell carcinoma |
| HIF | Hypoxia-inducible factor |
| DM | Diabetes mellitus |
| pVHL | von Hippel–Lindau protein |
| FBS | Fetal bovine serum |
| DMSO | Dimethyl sulfoxide |
| shRNA | Short hairpin RNA |
| DMEM | Dulbecco’s modified Eagle’s medium |
References
- WHO. Cancer Today. 2023. Available online: https://gco.iarc.fr/today/home (accessed on 27 February 2025).
- Zhu, Z.; Jin, Y.; Zhou, J.; Chen, F.; Chen, M.; Gao, Z.; Hu, L.; Xuan, J.; Li, X.; Song, Z.; et al. PD1/PD-L1 blockade in clear cell renal cell carcinoma: Mechanistic insights, clinical efficacy, and future perspectives. Mol. Cancer 2024, 23, 146. [Google Scholar] [CrossRef] [PubMed]
- Cancer Genome Atlas Research Network. Comprehensive molecular characterization of clear cell renal cell carcinoma. Nature 2013, 499, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Yoshizato, T.; Shiraishi, Y.; Maekawa, S.; Okuno, Y.; Kamura, T.; Shimamura, T.; Sato-Otsubo, A.; Nagae, G.; Suzuki, H.; et al. Integrated molecular analysis of clear-cell renal cell carcinoma. Nat. Genet. 2013, 45, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Wang, X.; Lu, S.; Ou, K. Metabolic reprogramming of clear cell renal cell carcinoma. Front. Endocrinol. 2023, 14, 1195500. [Google Scholar] [CrossRef]
- Chatterjee, S.; Khunti, K.; Davies, M.J. Type 2 diabetes. Lancet 2017, 389, 2239–2251. [Google Scholar] [CrossRef]
- Tsilidis, K.K.; Kasimis, J.C.; Lopez, D.S.; Ntzani, E.E.; Ioannidis, J.P. Type 2 diabetes and cancer: Umbrella review of meta-analyses of observational studies. BMJ 2015, 350, g7607. [Google Scholar] [CrossRef]
- Ling, S.; Brown, K.; Miksza, J.K.; Howells, L.; Morrison, A.; Issa, E.; Yates, T.; Khunti, K.; Davies, M.J.; Zaccardi, F. Association of type 2 diabetes with cancer: A meta-analysis with bias analysis for unmeasured confounding in 151 cohorts comprising 32 million people. Diabetes Care 2020, 43, 2313–2322. [Google Scholar] [CrossRef]
- Pearson-Stuttard, J.; Papadimitriou, N.; Markozannes, G.; Cividini, S.; Kakourou, A.; Gill, D.; Rizos, E.C.; Monori, G.; Ward, H.A.; Kyrgiou, M.; et al. Type 2 diabetes and cancer: An umbrella review of observational and Mendelian randomization studies. Cancer Epidemiol. Biomarkers Prev. 2021, 30, 1218–1228. [Google Scholar] [CrossRef]
- Bao, C.; Yang, X.; Xu, W.; Luo, H.; Xu, Z.; Su, C.; Qi, X. Diabetes mellitus and incidence and mortality of kidney cancer: A meta-analysis. J. Diabetes Complicat. 2013, 27, 357–364. [Google Scholar] [CrossRef]
- Psutka, S.P.; Stewart, S.B.; Boorjian, S.A.; Lohse, C.M.; Tollefson, M.K.; Cheville, J.C.; Leibovich, B.C.; Thompson, R.H. Diabetes mellitus is independently associated with an increased risk of mortality in patients with clear cell renal cell carcinoma. J. Urol. 2014, 192, 1620–1627. [Google Scholar] [CrossRef]
- Lee, H.; Kwak, C.; Kim, H.H.; Byun, S.S.; Lee, S.E.; Hong, S.K. Diabetes mellitus as an independent predictor of survival of patients surgically treated for renal cell carcinoma: A propensity score matching study. J. Urol. 2015, 194, 1554–1560. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.H.; Wang, W.Y.; Chen, H.Y.; Liao, P.L.; Jong, G.P.; Yang, T.Y. Decreased risk of renal cell carcinoma in patients with type 2 diabetes treated with sodium glucose cotransporter-2 inhibitors. Cancer Sci. 2024, 115, 2059–2066. [Google Scholar] [CrossRef] [PubMed]
- Basak, D.; Gamez, D.; Deb, S. SGLT2 inhibitors as potential anticancer agents. Biomedicines 2023, 11, 1867. [Google Scholar] [CrossRef]
- Kuang, H.; Liao, L.; Chen, H.; Kang, Q.; Shu, X.; Wang, Y. Therapeutic effect of sodium glucose co-transporter 2 inhibitor dapagliflozin on renal cell carcinoma. Med. Sci. Monit. 2017, 23, 3737–3745. [Google Scholar] [CrossRef]
- Kobayashi, M.; Uematsu, T.; Tokura, Y.; Takei, K.; Sakamoto, K.; Narimatsu, T.; Nukui, A.; Kamai, T. Immunohistochemical expressionof sodium-dependent glucose transporter-2 (SGLT-2) in clear cell renal carcinoma: Possible prognostic implications. Int. Braz. J. Urol. 2019, 45, 169–178. [Google Scholar] [CrossRef]
- Von Roemeling, C.A.; Radisky, D.C.; Marlow, L.A.; Cooper, S.J.; Grebe, S.K.; Anastasiadis, P.Z.; Tun, H.W.; Copland, J.A. Neuronal pentraxin 2 supports clear cell renal cell carcinoma by activating the AMPA-selective glutamate receptor-4. Cancer Res. 2014, 74, 4796–4810. [Google Scholar] [CrossRef]
- Kort, E.J.; Farber, L.; Tretiakova, M.; Petillo, D.; Furge, K.A.; Yang, X.J.; Cornelius, A.; Teh, B.T. The E2F3-Oncomir-1 axis is activated in Wilms’ tumor. Cancer Res. 2008, 68, 4034–4038. [Google Scholar] [CrossRef]
- Pandey, A.; Alcaraz, M., Jr.; Saggese, P.; Soto, A.; Gomez, E.; Jaldu, S.; Yanagawa, J.; Scafoglio, C. Exploring the role of SGLT2 inhibitors in cancer: Mechanisms of action and therapeutic opportunities. Cancers 2025, 17, 466. [Google Scholar] [CrossRef]
- Saito, T.; Okada, S.; Yamada, E.; Shimoda, Y.; Osaki, A.; Tagaya, Y.; Shibusawa, R.; Okada, J.; Yamada, M. Effect of dapagliflozin on colon cancer cell [Rapid Communication]. Endocr. J. 2015, 62, 1133–1137. [Google Scholar] [CrossRef]
- Villani, L.A.; Smith, B.K.; Marcinko, K.; Ford, R.J.; Broadfield, L.A.; Green, A.E.; Houde, V.P.; Muti, P.; Tsakiridis, T.; Steinberg, G.R. The diabetes medication canagliflozin reduces cancer cell proliferation by inhibiting mitochondrial complex-I supported respiration. Mol. Metab. 2016, 5, 1048–1056. [Google Scholar] [CrossRef] [PubMed]
- Kaji, K.; Nishimura, N.; Seki, K.; Sato, S.; Saikawa, S.; Nakanishi, K.; Furukawa, M.; Kawaratani, H.; Kitade, M.; Moriya, K.; et al. Sodium glucose cotransporter 2 inhibitor canagliflozin attenuates liver cancer cell growth and angiogenic activity by inhibiting glucose uptake. Int. J. Cancer 2018, 142, 1712–1722. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, S.; Nomiyama, T.; Numata, T.; Kawanami, T.; Hamaguchi, Y.; Iwaya, C.; Horikawa, T.; Fujimura-Tanaka, Y.; Hamanoue, N.; Motonaga, R.; et al. SGLT2 inhibitor ipragliflozin attenuates breast cancer cell proliferation. Endocr. J. 2020, 67, 99–106. [Google Scholar] [CrossRef]
- Zhou, J.; Zhu, J.; Yu, S.J.; Ma, H.L.; Chen, J.; Ding, X.F.; Chen, G.; Liang, Y.; Zhang, Q. Sodium-glucose co-transporter-2 (SGLT-2) inhibition reduces glucose uptake to induce breast cancer cell growth arrest through AMPK/mTOR pathway. Biomed. Pharmacother. 2020, 132, 110821. [Google Scholar] [CrossRef]
- Ren, D.; Sun, Y.; Zhang, D.; Li, D.; Liu, Z.; Jin, X.; Wu, H. SGLT2 promotes pancreatic cancer progression by activating the Hippo signaling pathway via the hnRNPK-YAP1 axis. Cancer Lett. 2021, 519, 277–288. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, L.; Mao, L.; Zhang, L.; Zhu, Y.; Xu, Y.; Cheng, Y.; Sun, R.; Zhang, Y.; Ke, J.; et al. SGLT2 inhibition restrains thyroid cancer growth via G1/S phase transition arrest and apoptosis mediated by DNA damage response signaling pathways. Cancer Cell Int. 2022, 22, 74. [Google Scholar] [CrossRef]
- Wu, W.; Zhang, Z.; Jing, D.; Huang, X.; Ren, D.; Shao, Z.; Zhang, Z. SGLT2 inhibitor activates the STING/IRF3/IFN-β pathway and induces immune infiltration in osteosarcoma. Cell Death Dis. 2022, 13, 523. [Google Scholar] [CrossRef]
- Miao, X.; Zhang, J.; Huang, W.; Wang, Y.; Jin, A.; Cao, J.; Zhao, Z. Research progress of SGLT2 inhibitors in cancer treatment. Drug Des. Dev. Ther. 2025, 19, 505–514. [Google Scholar] [CrossRef]
- Jang, J.H.; Lee, T.J.; Sung, E.G.; Song, I.H.; Kim, J.Y. Dapagliflozin induces apoptosis by downregulating cFILPL and increasing cFILPS instability in Caki-1 cells. Oncol. Lett. 2022, 24, 401. [Google Scholar] [CrossRef]
- Lin, T.K.; Wang, W.Y.; Yang, T.Y.; Jong, G.P. Type disparity in sodium-glucose Cotransporter-2 inhibitors in incidences of renal cell carcinoma: A propensity-score-matched cohort study. Cancers 2024, 16, 2145. [Google Scholar] [CrossRef]
- Brodaczewska, K.K.; Szczylik, C.; Fiedorowicz, M.; Porta, C.; Czarnecka, A.M. Choosing the right cell line for renal cell cancer research. Mol. Cancer 2016, 15, 83. [Google Scholar] [CrossRef] [PubMed]
- Faubert, B.; Solmonson, A.; DeBerardinis, R.J. Metabolic reprogramming and cancer progression. Science 2020, 368, eaaw5473. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, B.; Truong, L.D. Renal epithelial neoplasms: The diagnostic implications of electron microscopic study in 55 cases. Hum. Pathol. 2002, 33, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Kondo, K.; Klco, J.; Nakamura, E.; Lechpammer, M.; Kaelin, W.G., Jr. Inhibition of HIF is necessary for tumor suppression by the von Hippel–Lindau protein. Cancer Cell 2002, 1, 237–246. [Google Scholar] [CrossRef]
- Isono, T.; Chano, T.; Yonese, J.; Yuasa, T. Therapeutic inhibition of mitochondrial function induces cell death in starvation-resistant renal cell carcinomas. Sci. Rep. 2016, 6, 25669. [Google Scholar] [CrossRef]
- Isono, T.; Chano, T.; Yoshida, T.; Kageyama, S.; Kawauchi, A.; Suzaki, M.; Yuasa, T. Hydroxyl-HIF2-alpha is potential therapeutic target for renal cell carcinomas. Am. J. Cancer Res. 2016, 6, 2263–2276. [Google Scholar]
- Nagata, Y.; Goto, T.; Teramoto, Y.; Matsukawa, T.; Fujimoto, N.; Miyamoto, H. Mineralocorticoid receptor signaling inhibits bladder cancer progression. Am. J. Cancer Res. 2024, 14, 696–708. [Google Scholar] [CrossRef]
- Nagata, Y.; Matsukawa, T.; Goto, T.; Teramoto, Y.; Jiang, G.; Fujimoto, N.; Miyamoto, H. Protective role of mineralocorticoid receptor signaling in urothelial tumorigenesis. Am. J. Cancer Res. 2023, 13, 408–418. [Google Scholar]
- Nguyen, U.; Squaglia, N.; Boge, A.; Fung, P.A. The Simple Western™: A gel-free, blot-free, hands-free Western blotting reinvention. Nat. Methods 2011, 8, v–vi. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagata, Y.; Tomisaki, I.; Aono, H.; Quynh, N.T.; Kashiwagi, E.; Fujimoto, N. Sodium–Glucose Cotransporter 2 and Glucose Levels Affect Clear Cell Renal Cell Carcinoma Progression. Int. J. Mol. Sci. 2025, 26, 5501. https://doi.org/10.3390/ijms26125501
Nagata Y, Tomisaki I, Aono H, Quynh NT, Kashiwagi E, Fujimoto N. Sodium–Glucose Cotransporter 2 and Glucose Levels Affect Clear Cell Renal Cell Carcinoma Progression. International Journal of Molecular Sciences. 2025; 26(12):5501. https://doi.org/10.3390/ijms26125501
Chicago/Turabian StyleNagata, Yujiro, Ikko Tomisaki, Hisami Aono, Nguyen Thu Quynh, Eiji Kashiwagi, and Naohiro Fujimoto. 2025. "Sodium–Glucose Cotransporter 2 and Glucose Levels Affect Clear Cell Renal Cell Carcinoma Progression" International Journal of Molecular Sciences 26, no. 12: 5501. https://doi.org/10.3390/ijms26125501
APA StyleNagata, Y., Tomisaki, I., Aono, H., Quynh, N. T., Kashiwagi, E., & Fujimoto, N. (2025). Sodium–Glucose Cotransporter 2 and Glucose Levels Affect Clear Cell Renal Cell Carcinoma Progression. International Journal of Molecular Sciences, 26(12), 5501. https://doi.org/10.3390/ijms26125501