
The Anatomical and Evolutionary Impact of Pain, Pleasure, Motivation, and Cognition: Integrating Energy Metabolism and the Mind–Body BERN (Behavior, Exercise, Relaxation, and Nutrition) Framework



{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
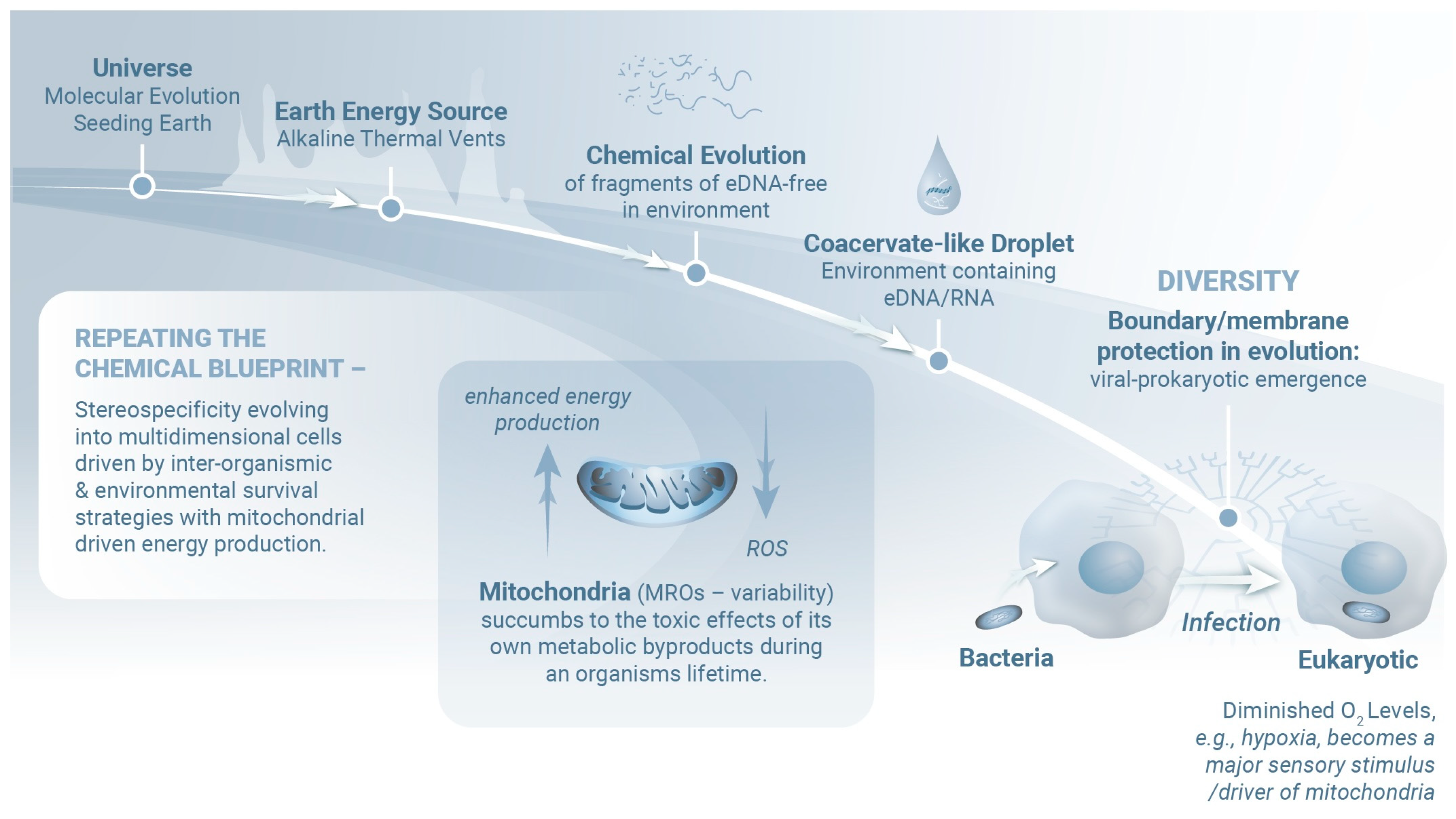
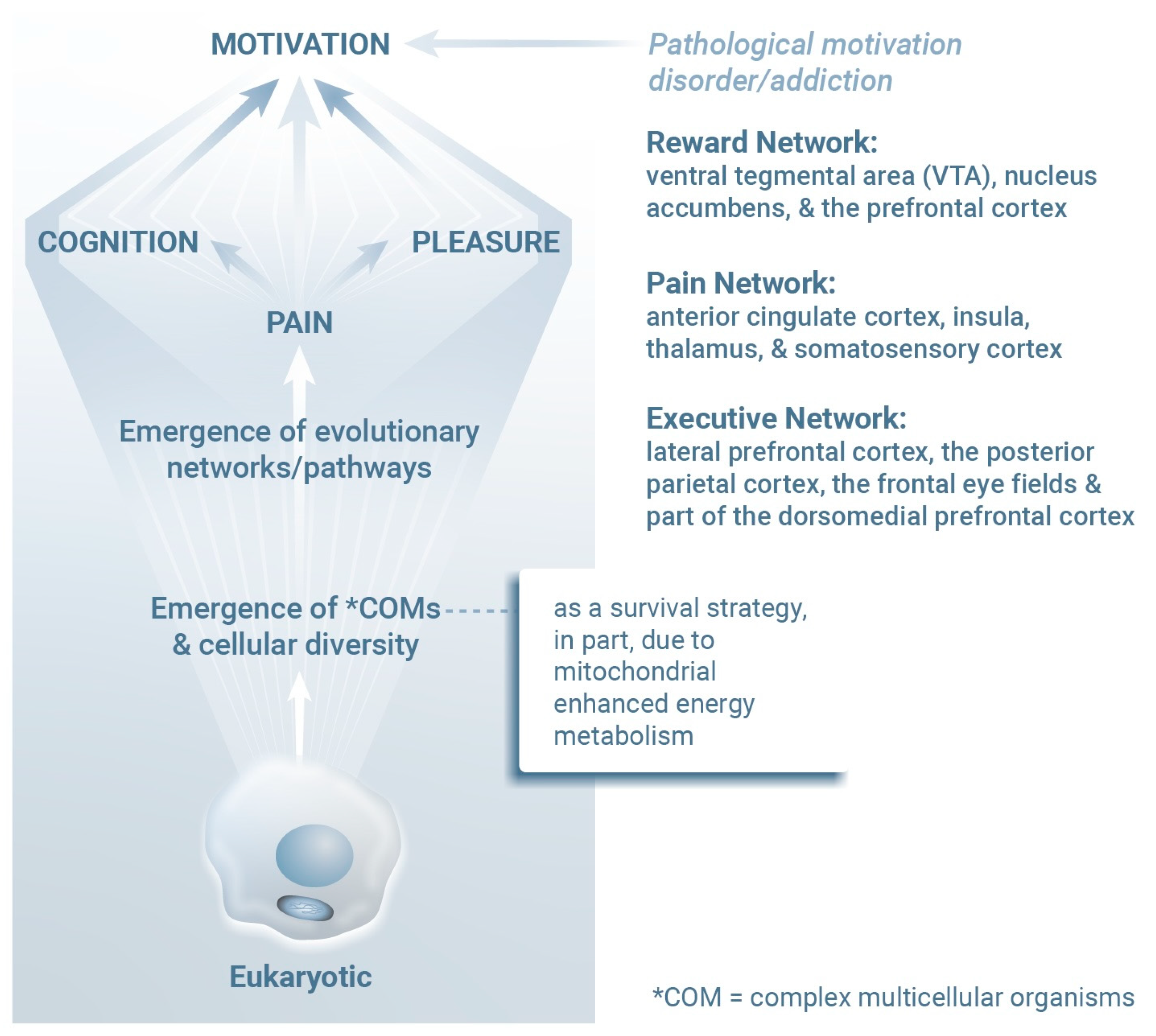
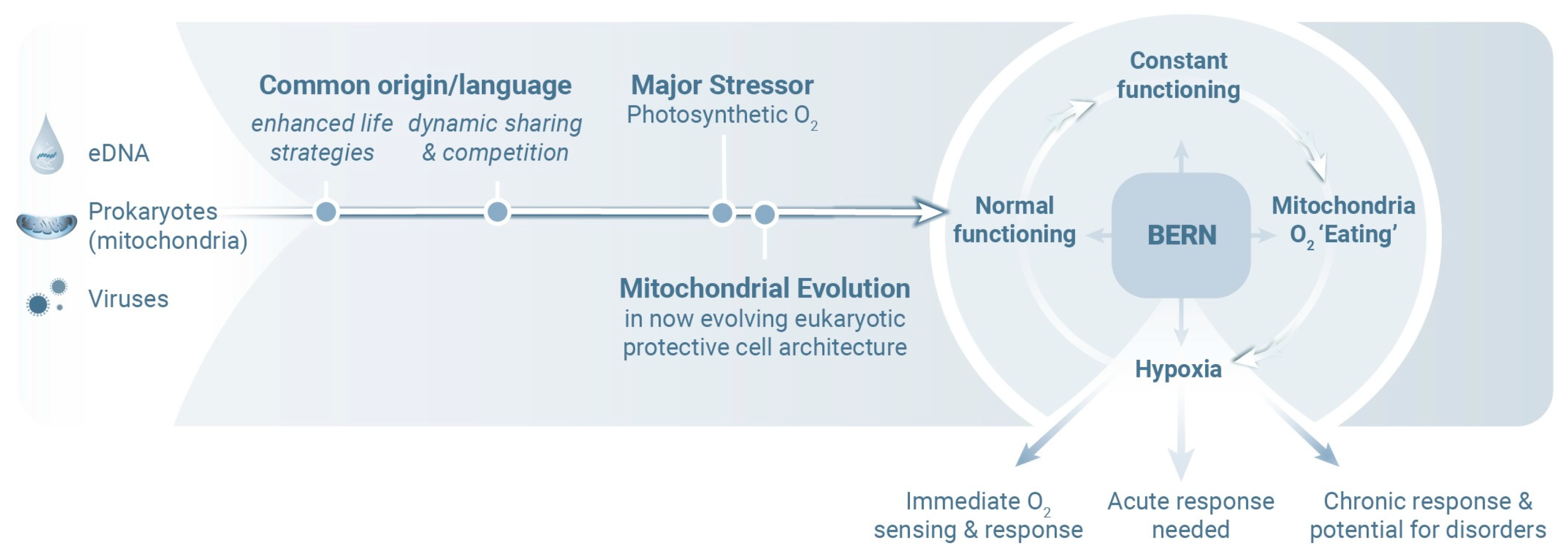
2. Molecular Mind–Body Platform
3. Psychiatric Ramifications
4. Evolutionary Efficiency
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ADP | adenosine diphosphate |
| ATP | adenosine triphosphate |
| BERN | Behavior, Exercise, Relaxation, and Nutrition |
| DNA | deoxyribonucleic acid |
| eDNARNA | environmental DNAribonucleic acid |
| MROs | mitochondrion-related organelles |
| CNS | central nervous system |
| VTA | ventral tegmental area |
| NAc | nucleus accumbens |
| PFC | prefrontal cortex |
| ACC | anterior cingulate cortex |
| LPFC | lateral prefrontal cortex |
| PCC | posterior cingulate cortex |
| PPC | posterior parietal cortex |
| dmPFC | dorsomedial prefrontal cortex |
| HIF | hypoxia-inducible factor |
| ROSs | reactive oxygen species |
References
- Stefano, G.B.; Kream, R.M. Hypoxia defined as a common culprit/initiation factor in mitochondrial-mediated proinflammatory processes. Med. Sci. Monit. 2015, 21, 1478–1484. [Google Scholar] [CrossRef] [PubMed]
- McCoy, T.J.; Russell, S.S.; Zega, T.J.; Thomas-Keprta, K.L.; Singerling, S.A.; Brenker, F.E.; Timms, N.E.; Rickard, W.D.A.; Barnes, J.J.; Libourel, G.; et al. An evaporite sequence from ancient brine recorded in Bennu samples. Nature 2025, 637, 1072–1077. [Google Scholar] [CrossRef] [PubMed]
- Glavin, D.P.; Dworkin, J.P.; Alexander, C.M.O.D.; Aponte, J.C.; Baczynski, A.A.; Barnes, J.J.; Bechtel, H.A.; Berger, E.L.; Burton, A.S.; Caselli, P.; et al. Abundant ammonia and nitrogen-rich soluble organic matter in samples from asteroid (101955) Bennu. Nat. Astron. 2025, 9, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Marchi, S.; Guilbaud, E.; Tait, S.W.G.; Yamazaki, T.; Galluzzi, L. Mitochondrial control of inflammation. Nat. Rev. Immunol. 2022, 23, 159–173. [Google Scholar] [CrossRef] [PubMed]
- López-Armada, M.J.; Riveiro-Naveira, R.R.; Vaamonde-García, C.; Valcárcel-Ares, M.N. Mitochondrial dysfunction and the inflammatory response. Mitochondrion 2013, 13, 106–118. [Google Scholar] [CrossRef]
- Van Horssen, J.; Van Schaik, P.; Witte, M. Inflammation and mitochondrial dysfunction: A vicious circle in neurodegenerative disorders? Neurosci. Lett. 2017, 710, 132931. [Google Scholar] [CrossRef]
- Stefano, G.B.; Esch, T.; Kream, R.M. Behaviorally-mediated entrainment of whole-body metabolic processes: Conservation and evolutionary development of mitochondrial respiratory complexes. Med. Sci. Monit. 2019, 25, 9306–9309. [Google Scholar] [CrossRef]
- Stefano, G.B.; Esch, T.; Kream, R.M. Augmentation of Whole-Body Metabolic Status by Mind-Body Training: Synchronous Integration of Tissue- and Organ-Specific Mitochondrial Function. Med. Sci. Monit. Basic Res. 2019, 25, 8–14. [Google Scholar] [CrossRef]
- Eltzschig, H.K.; Carmeliet, P. Hypoxia and inflammation. N. Engl. J. Med. 2011, 364, 656–665. [Google Scholar] [CrossRef]
- Hartmann, G.; Tschöp, M.; Fischer, R.; Bidlingmaier, C.; Riepl, R.; Tschöp, K.; Hautmann, H.; Endres, S.; Toepfer, M. High Altitude Increases Circulating Interleukin-6, Interleukin-1 Receptor Antagonist and C-Reactive Protein. Cytokine 2000, 12, 246–252. [Google Scholar] [CrossRef]
- Palazon, A.; Goldrath, A.W.; Nizet, V.; Johnson, R.S. HIF transcription factors, inflammation, and immunity. Immunity 2014, 41, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.-T.; Huang, L.-T.; Chen, C.-C.; Chen, C.-M. Molecular mechanisms underlying hyperoxia-induced lung fibrosis. Pediatr. Neonatol. 2022, 63, 109–116. [Google Scholar] [CrossRef]
- Bakare, A.B.; Lesnefsky, E.J.; Iyer, S. Leigh Syndrome: A tale of two genomes. Front. Physiol. 2021, 12, 693734. [Google Scholar] [CrossRef]
- van de Wal, M.A.E.; Adjobo-Hermans, M.J.W.; Keijer, J.; Schirris, T.J.J.; Homberg, J.R.; Wieckowski, M.R.; Grefte, S.; Van Schothorst, E.M.; Van Karnebeek, C.; Quintana, A.; et al. Ndufs4 knockout mouse models of Leigh syndrome: Pathophysiology and intervention. Brain 2021, 145, 45–63. [Google Scholar] [CrossRef]
- Jain, I.H.; Zazzeron, L.; Goli, R.; Alexa, K.; Schatzman-Bone, S.; Dhillon, H.; Goldberger, O.; Peng, J.; Shalem, O.; Sanjana, N.E.; et al. Hypoxia as a therapy for mitochondrial disease. Science 2016, 352, 54–61. [Google Scholar] [CrossRef]
- Ferrari, M.; Jain, I.H.; Goldberger, O.; Rezoagli, E.; Thoonen, R.; Cheng, K.-H.; Sosnovik, D.E.; Scherrer-Crosbie, M.; Mootha, V.K.; Zapol, W.M. Hypoxia treatment reverses neurodegenerative disease in a mouse model of Leigh syndrome. Proc. Natl. Acad. Sci. USA 2017, 114, E4241–E4250. [Google Scholar] [CrossRef]
- Finsterer, J.; Zarrouk-Mahjoub, S. Psychosis in Leigh syndrome. Asian J. Psychiatry 2019, 41, 76–77. [Google Scholar] [CrossRef] [PubMed]
- Jaballah, F.; Nouira, R.B.S.; Mallouli, S.; Boussaid, H.; Younes, S.; Zarrouk, L.; Younes, S. Schizophrenia-Like psychotic symptoms associated to Leigh Syndrome. Case Rep. Psychiatry 2023, 2023, 8886555. [Google Scholar] [CrossRef] [PubMed]
- Stefano, G.B.; Kream, R.M. Primordial biochemicals within coacervate-Like droplets and the origins of life. Viruses 2025, 17, 146. [Google Scholar] [CrossRef]
- Záhonová, K.; Treitli, S.C.; Le, T.; Škodová-Sveráková, I.; Hanousková, P.; Čepička, I.; Tachezy, J.; Hampl, V. Anaerobic derivates of mitochondria and peroxisomes in the free-living amoeba Pelomyxa schiedti revealed by single-cell genomics. BMC Biol. 2022, 20, 56. [Google Scholar] [CrossRef]
- Schavemaker, P.E.; Muñoz-Gómez, S.A. The role of mitochondrial energetics in the origin and diversification of eukaryotes. Nat. Ecol. Evol. 2022, 6, 1307–1317. [Google Scholar] [CrossRef]
- Muñoz-Gómez, S.A.; Susko, E.; Williamson, K.; Eme, L.; Slamovits, C.H.; Moreira, D.; López-García, P.; Roger, A.J. Site-and-branch-heterogeneous analyses of an expanded dataset favour mitochondria as sister to known Alphaproteobacteria. Nat. Ecol. Evol. 2022, 6, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Stephens, O.R.; Grant, D.; Frimel, M.; Wanner, N.; Yin, M.; Willard, B.; Erzurum, S.C.; Asosingh, K. Characterization and origins of cell-free mitochondria in healthy murine and human blood. Mitochondrion 2020, 54, 102–112. [Google Scholar] [CrossRef]
- Miliotis, S.; Nicolalde, B.; Ortega, M.; Yepez, J.; Caicedo, A. Forms of extracellular mitochondria and their impact in health. Mitochondrion 2019, 48, 16–30. [Google Scholar] [CrossRef] [PubMed]
- Dache, Z.A.A.; Thierry, A.R. Mitochondria-derived cell-to-cell communication. Cell Rep. 2023, 42, 112728. [Google Scholar] [CrossRef] [PubMed]
- Dache, Z.A.A.; Otandault, A.; Tanos, R.; Pastor, B.; Meddeb, R.; Sanchez, C.; Arena, G.; Lasorsa, L.; Bennett, A.; Grange, T.; et al. Blood contains circulating cell-free respiratory competent mitochondria. FASEB J. 2020, 34, 3616–3630. [Google Scholar] [CrossRef]
- Park, J.-H.; Hayakawa, K. Extracellular mitochondria signals in CNS disorders. Front. Cell Dev. Biol. 2021, 9, 642853. [Google Scholar] [CrossRef]
- Raval, P.K.; Garg, S.G.; Gould, S.B. Endosymbiotic selective pressure at the origin of eukaryotic cell biology. eLife 2022, 11, e81033. [Google Scholar] [CrossRef]
- Mattson, M.P.; Gleichmann, M.; Cheng, A. Mitochondria in neuroplasticity and neurological disorders. Neuron 2008, 60, 748–766. [Google Scholar] [CrossRef]
- Johri, A.; Beal, M.F. Mitochondrial dysfunction in neurodegenerative diseases. J. Pharmacol. Exp. Ther. 2012, 342, 619–630. [Google Scholar] [CrossRef]
- Harris, J.J.; Jolivet, R.; Attwell, D. Synaptic energy use and supply. Neuron 2012, 75, 762–777. [Google Scholar] [CrossRef] [PubMed]
- Picard, M.; McEwen, B.S.; Epel, E.S.; Sandi, C. An energetic view of stress: Focus on mitochondria. Front. Neuroendocrinol. 2018, 49, 72–85. [Google Scholar] [CrossRef] [PubMed]
- Ni, P.; Ma, Y.; Chung, S. Mitochondrial dysfunction in psychiatric disorders. Schizophr. Res. 2024, 273, 62–77. [Google Scholar] [CrossRef] [PubMed]
- Pei, L.; Wallace, D.C. Mitochondrial etiology of Neuropsychiatric disorders. Biol. Psychiatry 2017, 83, 722–730. [Google Scholar] [CrossRef]
- Wei, W.; Cheng, B.; Zhao, Y.; He, D.; Chu, X.; Qin, X.; Zhang, N.; Shi, S.; Cai, Q.; Hui, J.; et al. Exploring the Interplay between Mitochondrial DNA and Lifestyle Factors in the Pathogenesis of Psychiatric Disorders. Depress. Anxiety 2024, 2024, 4914777. [Google Scholar] [CrossRef]
- Esch, T.; Fricchione, G.L.; Stefano, G.B. The therapeutic use of the relaxation response in stress-related diseases. Med. Sci. Monit. 2003, 9, RA23–RA34. [Google Scholar]
- Balban, M.Y.; Neri, E.; Kogon, M.M.; Weed, L.; Nouriani, B.; Jo, B.; Holl, G.; Zeitzer, J.M.; Spiegel, D.; Huberman, A.D. Brief structured respiration practices enhance mood and reduce physiological arousal. Cell Rep. Med. 2023, 4, 100895. [Google Scholar] [CrossRef]
- Cavanagh, M.; Cope, T.; Smith, D.; Tolley, I.; Orrock, P.; Vaughan, B. The effectiveness of an osteopathic manual technique compared with a breathing exercise on vagal tone as indicated by heart rate variability, a crossover study. J. Bodyw. Mov. Ther. 2024, 38, 449–453. [Google Scholar] [CrossRef]
- Herawati, I.; Ludin, A.F.M.; M, M.; Ishak, I.; Farah, N.M.F. Breathing exercise for hypertensive patients: A scoping review. Front. Physiol. 2023, 14, 1048338. [Google Scholar] [CrossRef]
- Leknes, S.; Tracey, I. A common neurobiology for pain and pleasure. Nat. Rev. Neurosci. 2008, 9, 314–320. [Google Scholar] [CrossRef]
- Nicholls, D.G.; Ferguson, S.J. Bioenergetics, 4th ed.; University of Oxford: Oxford, UK, 2013. [Google Scholar] [CrossRef]
- Cheng, A.; Hou, Y.; Mattson, M.P. Mitochondria and neuroplasticity. ASN Neuro 2010, 2, e00045. [Google Scholar] [CrossRef] [PubMed]
- Manji, H.; Kato, T.; Di Prospero, N.A.; Ness, S.; Beal, M.F.; Krams, M.; Chen, G. Impaired mitochondrial function in psychiatric disorders. Nat. Rev. Neurosci. 2012, 13, 293–307. [Google Scholar] [CrossRef] [PubMed]
- Wallace, D.C. A mitochondrial paradigm of metabolic and degenerative diseases, aging, and Cancer: A Dawn for Evolutionary Medicine. Annu. Rev. Genet. 2005, 39, 359–407. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.I.; Neumeister, M.W. Pain: Pathways and Physiology. Clin. Plast. Surg. 2020, 47, 173–180. [Google Scholar] [CrossRef]
- Geuter, S.; Reynolds Losin, E.A.; Roy, M.; Atlas, L.Y.; Schmidt, L.; Krishnan, A.; Koban, L.; Wager, T.D.; Lindquist, M.A. Multiple Brain Networks Mediating Stimulus-Pain Relationships in Humans. Cereb. Cortex 2020, 30, 4204–4219. [Google Scholar] [CrossRef]
- Lewis, R.G.; Florio, E.; Punzo, D.; Borrelli, E. The Brain’s Reward System in Health and Disease. In Circadian Clock in Brain Health and Disease; Engmann, O., Brancaccio, M., Eds.; Springer International Publishing: Cham, Switzerland, 2021; Volume 1344, pp. 57–69. [Google Scholar]
- Park, H.J.; Friston, K. Structural and functional brain networks: From connections to cognition. Science 2013, 342, 1238411. [Google Scholar] [CrossRef]
- Cohen, J.R.; D’Esposito, M. The Segregation and Integration of Distinct Brain Networks and Their Relationship to Cognition. J. Neurosci. 2016, 36, 12083–12094. [Google Scholar] [CrossRef]
- Baik, J.-H. Stress and the dopaminergic reward system. Exp. Mol. Med. 2020, 52, 1879–1890. [Google Scholar] [CrossRef]
- Seeley, W.W. Behavioral Variant Frontotemporal Dementia. Continuum 2019, 25, 76–100. [Google Scholar] [CrossRef]
- Lewis, R.G.; Florio, E.; Punzo, D.; Borrelli, E. The brain’s reward system in health and disease. Adv. Exp. Med. Biol. 2021, 1344, 57–69. [Google Scholar] [CrossRef]
- Horsburgh, A.; Summers, S.J.; Lewis, A.; Keegan, R.J.; Flood, A. The Relationship Between Pain and Interoception: A Systematic Review and Meta-Analysis. J. Pain. 2024, 25, 104476. [Google Scholar] [CrossRef] [PubMed]
- Bushnell, M.C.; Čeko, M.; Low, L.A. Cognitive and emotional control of pain and its disruption in chronic pain. Nat. Rev. Neurosci. 2013, 14, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Kummer, K.; Sheets, P.L. Targeting PFC dysfunction in pain. J. Pharmacol. Exp. Ther. 2024, 389, 268–276. [Google Scholar] [CrossRef]
- Yun, A.J.; Lee, P.Y.; Doux, J.D.; Conley, B.R. A general theory of evolution based on energy efficiency: Its implications for diseases. Med. Hypotheses 2005, 66, 664–670. [Google Scholar] [CrossRef] [PubMed]
- Lynch, M. The bioenergetic cost of building a metazoan. Proc. Natl. Acad. Sci. USA 2024, 121, e2414742121. [Google Scholar] [CrossRef]
- Attwell, D.; Laughlin, S.B. An energy budget for signaling in the grey matter of the brain. J. Cereb. Blood Flow Metab. 2001, 21, 1133–1145. [Google Scholar] [CrossRef]
- Magistretti, P.J.; Allaman, I. A cellular perspective on brain energy metabolism and functional imaging. Neuron 2015, 86, 883–901. [Google Scholar] [CrossRef]
- Surmeier, D.J.; Obeso, J.A.; Halliday, G.M. Selective neuronal vulnerability in Parkinson disease. Nat. Rev. Neurosci. 2017, 18, 101–113. [Google Scholar] [CrossRef]
- Lennie, P. The cost of cortical computation. Curr. Biol. 2003, 13, 493–497. [Google Scholar] [CrossRef]
- Erecińska, M.; Silver, I.A. Tissue oxygen tension and brain sensitivity to hypoxia. Respir. Physiol. 2001, 128, 263–276. [Google Scholar] [CrossRef]
- Esch, T.; Stefano, G.B.; Michaelsen, M.M. The foundations of mind-body medicine: Love, good relationships, and happiness modulate stress and promote health. Stress Health 2024, 40, e3387. [Google Scholar] [CrossRef] [PubMed]
- Komaroff, A.L. Mind-Body Medicine: A Special Health Report; Harvard Health Publications: Boston, MA, USA, 2001. [Google Scholar]
- Esch, T. The Neurobiology of Meditation and Mindfulness. In Meditation—Neuroscientific Approaches and Philosophical Implications. Studies in Neuroscience, Consciousness and Spirituality; Schmidt, S., Walach, H., Eds.; Springer International Publishing: Cham, Switzerland, 2014; Volume 2, pp. 153–173. [Google Scholar] [CrossRef]
- Michaelsen, M.M.; Esch, T. Motivation and reward mechanisms in health behavior change processes. Brain Res. 2021, 1757, 147309. [Google Scholar] [CrossRef] [PubMed]
- Sternberg, E.M. Neural regulation of innate immunity: A coordinated nonspecific host response to pathogens. Nat. Rev. Immunol. 2006, 6, 318–328. [Google Scholar] [CrossRef]
- Pavlov, V.A.; Chavan, S.S.; Tracey, K.J. Molecular and functional neuroscience in immunity. Annu. Rev. Immunol. 2018, 36, 783–812. [Google Scholar] [CrossRef] [PubMed]
- Kumar, H.; Choi, D.-K. Hypoxia Inducible factor pathway and physiological adaptation: A cell survival pathway? Mediat. Inflamm. 2015, 2015, 584758. [Google Scholar] [CrossRef]
- Burtscher, J.; Hohenauer, E.; Burtscher, M.; Millet, G.P.; Egg, M. Environmental and behavioral regulation of HIF-mitochondria crosstalk. Free. Radic. Biol. Med. 2023, 206, 63–73. [Google Scholar] [CrossRef]
- Esch, T.; Stefano, G.B. The BERN Framework of Mind-Body Medicine: Integrating Self-Care, Health Promotion, Resilience, and Applied Neuroscience. Front. Integr. Neurosci. 2022, 16, 913573. [Google Scholar] [CrossRef]
- Esch, T. Integrating OpenNotes and Promoting Self-Management in Primary Care in Germany: The Witten Model. The BMJ (Opinion) 2021. Available online: https://blogs.bmj.com/bmj/2021/04/01/tobias-esch-integrating-opennotes-and-promoting-self-management-in-primary-care-in-germany-the-witten-model (accessed on 1 June 2025).
- Esch, T. Der Nutzen von Selbstheilungspotenzialen in der professionellen Gesundheitsfürsorge am Beispiel der Mind-Body-Medizin. Bundesgesundheitsblatt–Gesundheitsforschung–Gesundheitsschutz 2020, 63, 577–585. [Google Scholar] [CrossRef]
- Esch, T.; Kream, R.M.; Stefano, G.B. Chromosomal processes in mind-body medicine: Chronic stress, cell aging, and telomere length. Med. Sci. Monit. Basic Res. 2018, 24, 134–140. [Google Scholar] [CrossRef]
- Esch, T.; Stefano, G.B. The neurobiology of stress management. Neuro Endocrinol. Lett. 2010, 31, 19–39. [Google Scholar]
- Esch, T. Gesund im Stress: Der Wandel des Stresskonzeptes und seine Bedeutung für Prävention, Gesundheit und Lebensstil. Das. Gesundheitswesen 2002, 64, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Uhl, J.; Schönfeld, S.; Meyer, L.; Reus, A.; Neumann, C.; Langer, L.; Michaelsen, M.M.; Esch, T. (Digital) Mind-Body Intervention to Promote Health and Subjective Well-Being of Residents in Nursing Homes: A Cluster-Randomized Controlled Pilot Study. Das. Gesundheitswesen 2025. [Google Scholar] [CrossRef] [PubMed]
- Kohls, N.; Esch, T.; Gerber, L.; Adrian, L.; Wittmann, M. Mindfulness meditation and fantasy relaxation in a group setting leads to a diminished sense of self and an increased present orientation. Behav. Sci. 2019, 9, 87. [Google Scholar] [CrossRef] [PubMed]
- Möltner, H.; Leve, J.; Esch, T. Burnout-Prävention und mobile Achtsamkeit: Evaluation eines appbasierten Gesundheitstrainings bei Berufstätigen. Das. Gesundheitswesen 2017, 57, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Gimpel, C.; Von Scheidt, C.; Jose, G.; Sonntag, U.; Stefano, G.B.; Michalsen, A.; Esch, T. Changes and interactions of flourishing, mindfulness, sense of coherence, and quality of life in patients of a mind-body medicine outpatient clinic. Complement. Med. Res. 2014, 21, 154–162. [Google Scholar] [CrossRef]
- Schnieder, S.; Stappert, S.; Takahashi, M.; Fricchione, G.L.; Esch, T.; Krajewski, J. Sustainable reduction of sleepiness through salutogenic self-care procedure in lunch breaks: A pilot study. Evid.-Based Complement. Altern. Med. 2013, 2013, 387356. [Google Scholar] [CrossRef]
- Esch, T.; Jose, G.; Gimpel, C.; Von Scheidt, C.; Michalsen, A. Die Flourishing Scale (FS) von Diener et al. liegt jetzt in einer autorisierten deutschen Fassung (FS-D) vor: Einsatz bei einer Mind-Body-medizinischen Fragestellung. Complement. Med. Res. 2013, 20, 267–275. [Google Scholar] [CrossRef]
- Esch, T.; Sonntag, U.; Esch, S.M.; Thees, S. Stress Management and Mind-Body Medicine: A randomized controlled longitudinal evaluation of students’ health and effects of a behavioral group intervention at a middle-size German university (SM-MESH). Complement. Med. Res. 2013, 20, 129–137. [Google Scholar] [CrossRef]
- Feicht, T.; Wittmann, M.; Jose, G.; Mock, A.; Von Hirschhausen, E.; Esch, T. Evaluation of a Seven-Week Web-Based Happiness Training to Improve Psychological Well-Being, Reduce Stress, and Enhance Mindfulness and Flourishing: A Randomized Controlled Occupational Health Study. Evid.-Based Complement. Altern. Med. 2013, 2013, 676953. [Google Scholar] [CrossRef]
- Esch, T.; Duckstein, J.; Welke, J.; Braun, V. Mind/body techniques for physiological and psychological stress reduction: Stress management via Tai Chi training—A pilot study. Med. Sci. Monit. 2007, 13, CR488–CR497. [Google Scholar]
- Michalsen, A.; Grossman, P.; Acil, A.; Langhorst, J.; Lüdtke, R.; Esch, T.; Stefano, G.B.; Dobos, G.J. Rapid stress reduction and anxiolysis among distressed women as a consequence of a three-month intensive yoga program. Med. Sci. Monit. 2005, 11, CR555–CR561. [Google Scholar]
- Picard, M.; McEwen, B.S. Psychological Stress and Mitochondria: A Systematic Review. Psychosom. Med. 2018, 80, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Chandel, N.S. Mitochondria as signaling organelles. BMC Biol. 2014, 12, 34. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, G. The mitochondrial integrated stress response: A novel approach to anti-aging and pro-longevity. Ageing Res. Rev. 2025, 103, 102603. [Google Scholar] [CrossRef] [PubMed]
- Zong, Y.; Li, H.; Liao, P.; Chen, L.; Pan, Y.; Zheng, Y.; Zhang, C.; Liu, D.; Zheng, M.; Gao, J. Mitochondrial dysfunction: Mechanisms and advances in therapy. Signal Transduct. Target. Ther. 2024, 9, 124. [Google Scholar] [CrossRef]
- Daniels, T.E.; Olsen, E.M.; Tyrka, A.R. Stress and Psychiatric Disorders: The role of mitochondria. Annu. Rev. Clin. Psychol. 2020, 16, 165–186. [Google Scholar] [CrossRef]
- Andreazza, A.C.; Shao, L.; Wang, J.-F.; Young, L.T. Mitochondrial complex I activity and oxidative damage to mitochondrial proteins in the prefrontal cortex of patients with bipolar disorder. Arch. Gen. Psychiatry 2010, 67, 360–368. [Google Scholar] [CrossRef]
- Scaini, G.; Mason, B.L.; Diaz, A.P.; Jha, M.K.; Soares, J.C.; Trivedi, M.H.; Quevedo, J. Dysregulation of mitochondrial dynamics, mitophagy and apoptosis in major depressive disorder: Does inflammation play a role? Mol. Psychiatry 2021, 27, 1095–1102. [Google Scholar] [CrossRef]
- Ye, J.; Duan, C.; Han, J.; Chen, J.; Sun, N.; Li, Y.; Yuan, T.; Peng, D. Peripheral mitochondrial DNA as a neuroinflammatory biomarker for major depressive disorder. Neural Regen. Res. 2024, 20, 1541–1554. [Google Scholar] [CrossRef]
- Roberts, R.C. Mitochondrial dysfunction in schizophrenia: With a focus on postmortem studies. Mitochondrion 2020, 56, 91–101. [Google Scholar] [CrossRef]
- Nunnari, J.; Suomalainen, A. Mitochondria: In sickness and in health. Cell 2012, 148, 1145–1159. [Google Scholar] [CrossRef] [PubMed]
- Kann, O.; Papageorgiou, I.E.; Draguhn, A. Highly energized inhibitory interneurons are a central element for information processing in cortical networks. J. Cereb. Blood Flow Metab. 2014, 34, 1270–1282. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Sun, P.; Li, J.; Yang, Q.; Tian, Q.; Yuan, S.; Zhang, X.; Chen, P.; Li, C.; Zhang, X. Exploring genetic associations and drug targets for mitochondrial proteins and schizophrenia risk. Schizophrenia 2025, 11, 10. [Google Scholar] [CrossRef]
- Friston, K. The free-energy principle: A unified brain theory? Nat. Rev. Neurosci. 2010, 11, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Friston, K.; Kiebel, S. Predictive coding under the free-energy principle. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 1211–1221. [Google Scholar] [CrossRef]
- Barlow, H.B. Possible principles underlying the transformation of sensory messages. In Sensory Communication; MIT Press: Cambridge, MA, USA, 1961; pp. 217–234. [Google Scholar]
- Rao, R.P.N.; Ballard, D.H. Predictive coding in the visual cortex: A functional interpretation of some extra-classical receptive-field effects. Nat. Neurosci. 1999, 2, 79–87. [Google Scholar] [CrossRef]
- Keller, G.B.; Bonhoeffer, T.; Hübener, M. Sensorimotor mismatch signals in primary visual cortex of the behaving mouse. Neuron 2012, 74, 809–815. [Google Scholar] [CrossRef]
- Miall, R.C.; Wolpert, D.M. Forward models for physiological motor control. Neural Netw. 1996, 9, 1265–1279. [Google Scholar] [CrossRef]
- Shipp, S.; Adams, R.A.; Friston, K.J. Reflections on agranular architecture: Predictive coding in the motor cortex. Trends Neurosci. 2013, 36, 706–716. [Google Scholar] [CrossRef]
- Schultz, W.; Dayan, P.; Montague, P.R. A neural substrate of prediction and reward. Science 1997, 275, 1593–1599. [Google Scholar] [CrossRef] [PubMed]
- Chanes, L.; Barrett, L.F. Redefining the role of limbic areas in cortical processing. Trends Cogn. Sci. 2015, 20, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Hoogland, M.; Ploeger, A. Two different mismatches: Integrating the developmental and the Evolutionary-Mismatch hypothesis. Perspect. Psychol. Sci. 2022, 17, 1737–1745. [Google Scholar] [CrossRef]
- Bakhtiari, S. Energy efficiency as a normative account for predictive coding. Patterns 2022, 3, 100661. [Google Scholar] [CrossRef]
- Ficco, L.; Mancuso, L.; Manuello, J.; Teneggi, A.; Liloia, D.; Duca, S.; Costa, T.; Kovacs, G.Z.; Cauda, F. Disentangling predictive processing in the brain: A meta-analytic study in favour of a predictive network. Sci. Rep. 2021, 11, 16258. [Google Scholar] [CrossRef]
- Esch, T. The ABC model of happiness-neurobiological aspects of motivation and positive mood, and their dynamic changes through practice, the course of life. Biology 2022, 11, 843. [Google Scholar] [CrossRef]
- Mukandala, G.; Tynan, R.; Lanigan, S.; O’Connor, J. The effects of hypoxia and inflammation on synaptic signaling in the CNS. Brain Sci. 2016, 6, 6. [Google Scholar] [CrossRef]
- Fiskum, V.; Sandvig, A.; Sandvig, I. Silencing of Activity During Hypoxia Improves Functional Outcomes in Motor Neuron Networks in vitro. Front. Integr. Neurosci. 2021, 15, 792863. [Google Scholar] [CrossRef] [PubMed]
- Hencz, A.; Magony, A.; Thomas, C.; Kovacs, K.; Szilagyi, G.; Pal, J.; Sik, A. Mild hypoxia-induced structural and functional changes of the hippocampal network. Front. Cell. Neurosci. 2023, 17, 1277375. [Google Scholar] [CrossRef]
- Bustamante-Barrientos, F.A.; Luque-Campos, N.; Araya, M.J.; Lara-Barba, E.; De Solminihac, J.; Pradenas, C.; Molina, L.; Herrera-Luna, Y.; Utreras-Mendoza, Y.; Elizondo-Vega, R.; et al. Mitochondrial dysfunction in neurodegenerative disorders: Potential therapeutic application of mitochondrial transfer to central nervous system-residing cells. J. Transl. Med. 2023, 21, 613. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stefano, G.B.; Buttiker, P.; Michaelsen, M.M.; Esch, T. The Anatomical and Evolutionary Impact of Pain, Pleasure, Motivation, and Cognition: Integrating Energy Metabolism and the Mind–Body BERN (Behavior, Exercise, Relaxation, and Nutrition) Framework. Int. J. Mol. Sci. 2025, 26, 5491. https://doi.org/10.3390/ijms26125491
Stefano GB, Buttiker P, Michaelsen MM, Esch T. The Anatomical and Evolutionary Impact of Pain, Pleasure, Motivation, and Cognition: Integrating Energy Metabolism and the Mind–Body BERN (Behavior, Exercise, Relaxation, and Nutrition) Framework. International Journal of Molecular Sciences. 2025; 26(12):5491. https://doi.org/10.3390/ijms26125491
Chicago/Turabian StyleStefano, George B., Pascal Buttiker, Maren M. Michaelsen, and Tobias Esch. 2025. "The Anatomical and Evolutionary Impact of Pain, Pleasure, Motivation, and Cognition: Integrating Energy Metabolism and the Mind–Body BERN (Behavior, Exercise, Relaxation, and Nutrition) Framework" International Journal of Molecular Sciences 26, no. 12: 5491. https://doi.org/10.3390/ijms26125491
APA StyleStefano, G. B., Buttiker, P., Michaelsen, M. M., & Esch, T. (2025). The Anatomical and Evolutionary Impact of Pain, Pleasure, Motivation, and Cognition: Integrating Energy Metabolism and the Mind–Body BERN (Behavior, Exercise, Relaxation, and Nutrition) Framework. International Journal of Molecular Sciences, 26(12), 5491. https://doi.org/10.3390/ijms26125491