
Sinhyotaklisan as a Potential Therapeutic for Psoriasis: Network Pharmacology and Experimental Validation

 ,
,
Abstract
1. Introduction
2. Results
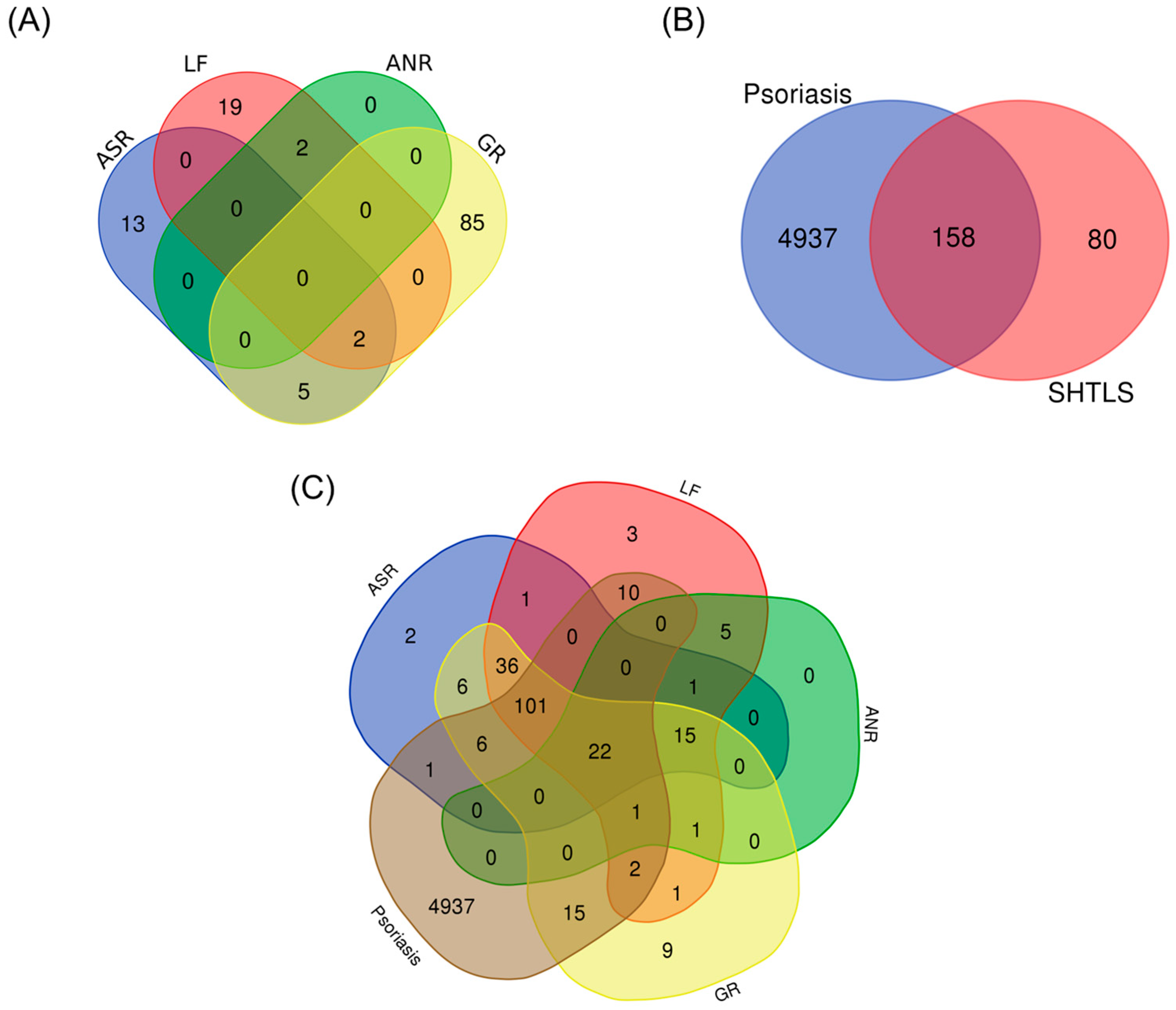
2.1. Screening of Potential Compounds and Key Targets of Sinhyotaklisan (SHTLS) in Psoriasis
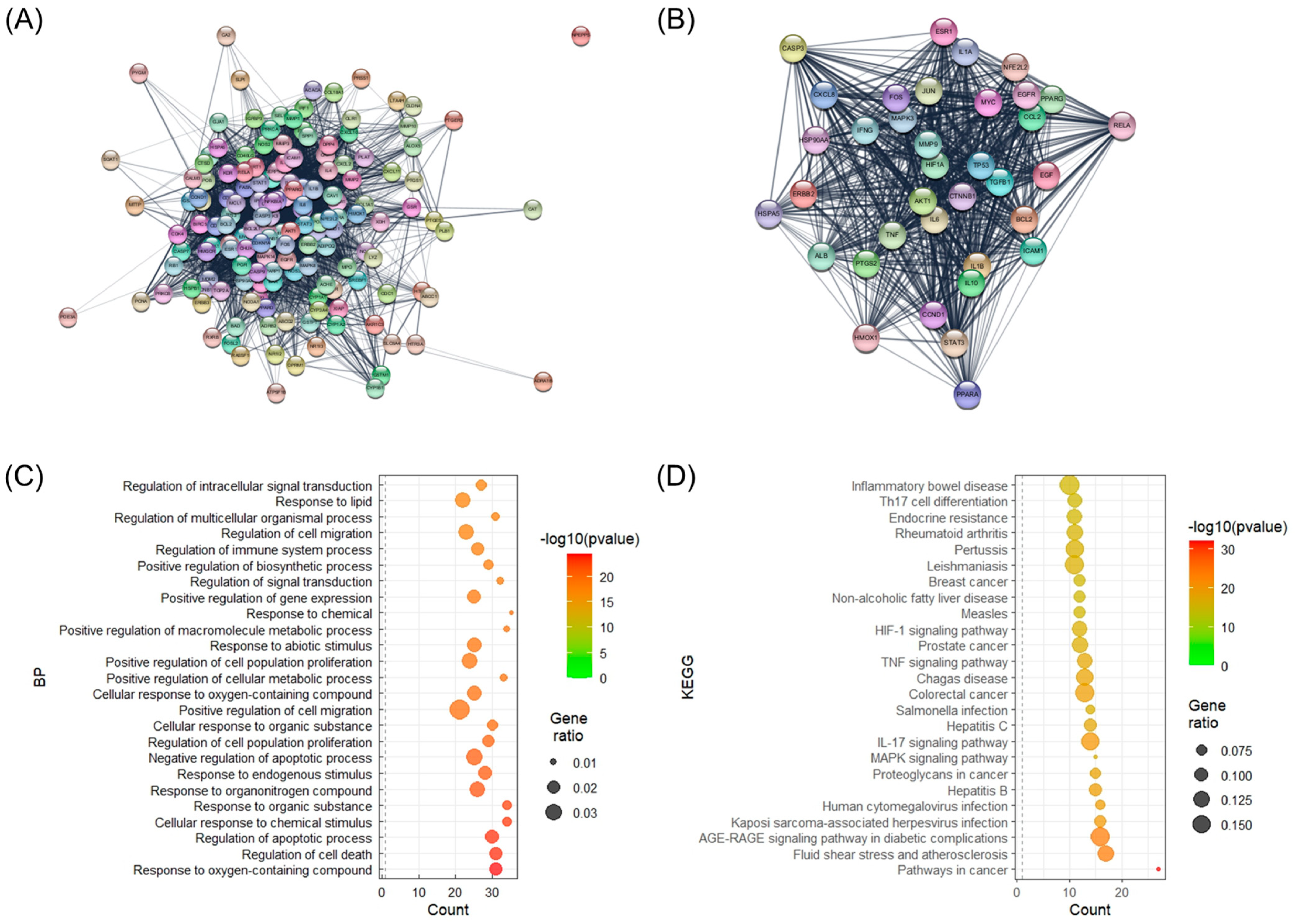
2.2. Analysis of Protein–Protein Interaction (PPI) Network, Biological Process (BP), and Kyoto Encyclopedia of Genes and Genomes (KEGG) Enrichment of Key Target Genes of Sinhyotaklisan (SHTLS)
2.3. Illustration of Key Target Genes in Psoriasis-Related Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathways
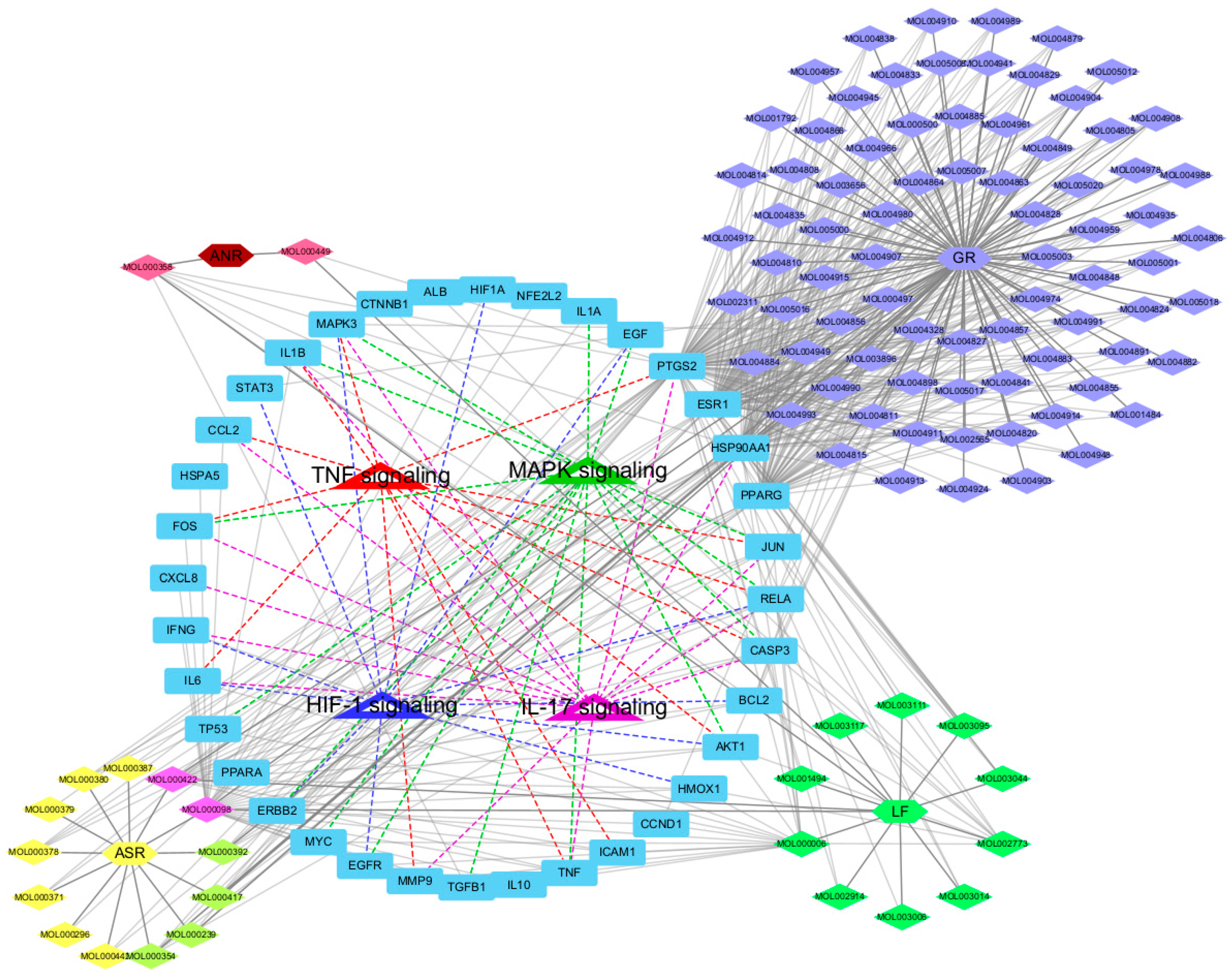
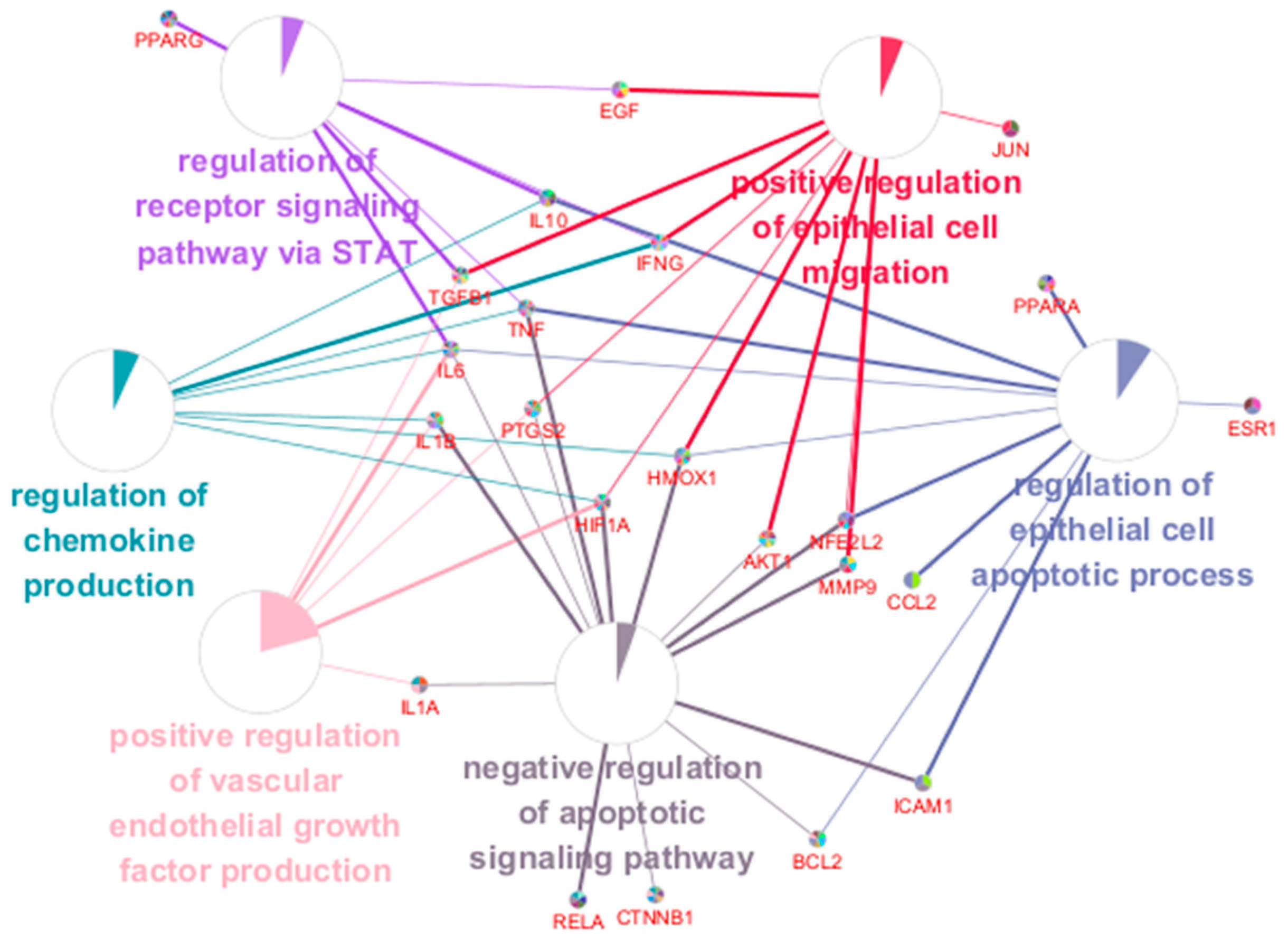
2.4. Visualization of the Herb-Compound-Target-Pathway Network and Enriched Biological Process (BP) Terms Network of Key Targets
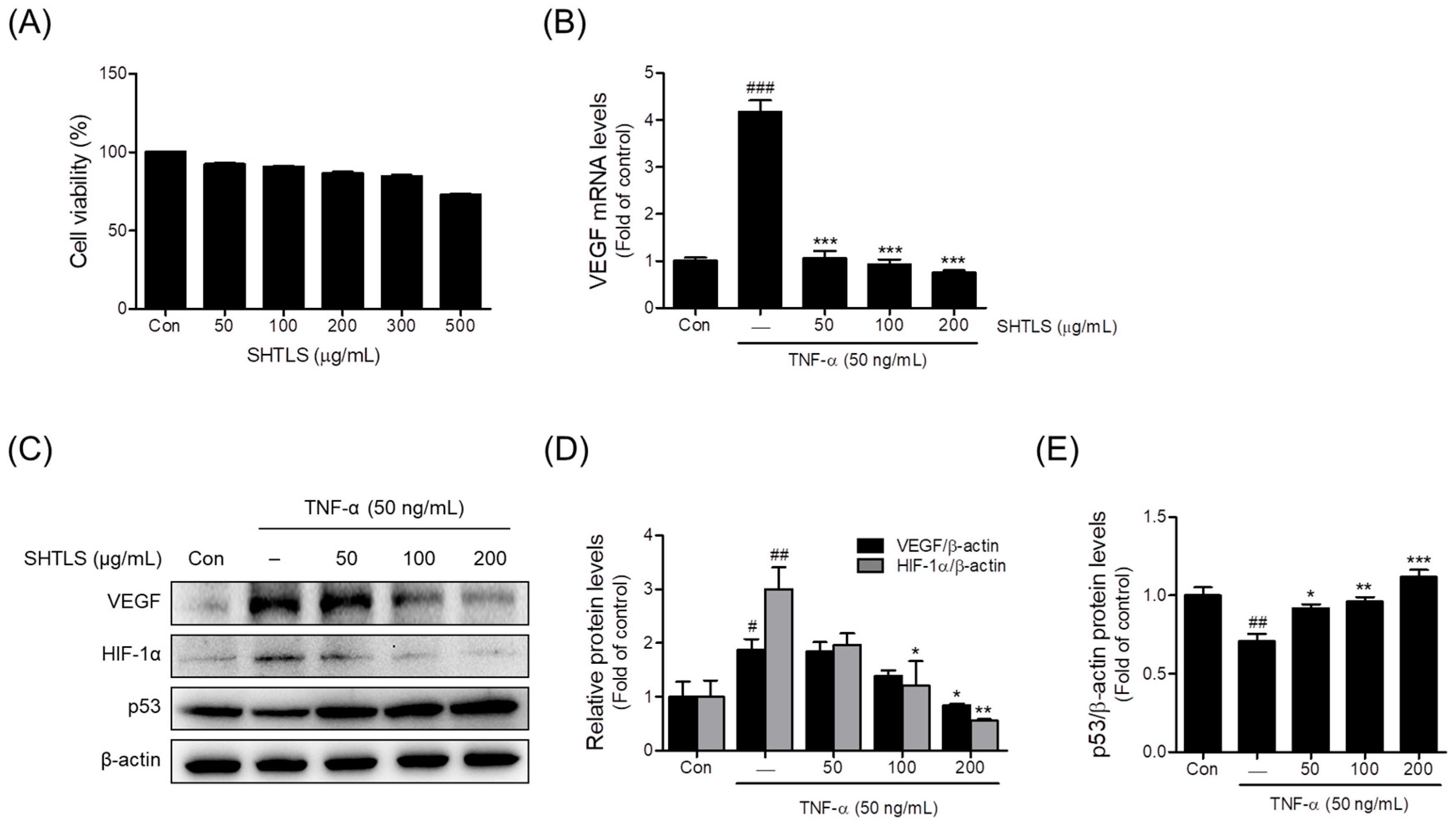
2.5. Effect of Sinhyotaklisan (SHTLS) Treatment on HaCaT Keratinocytes Viability
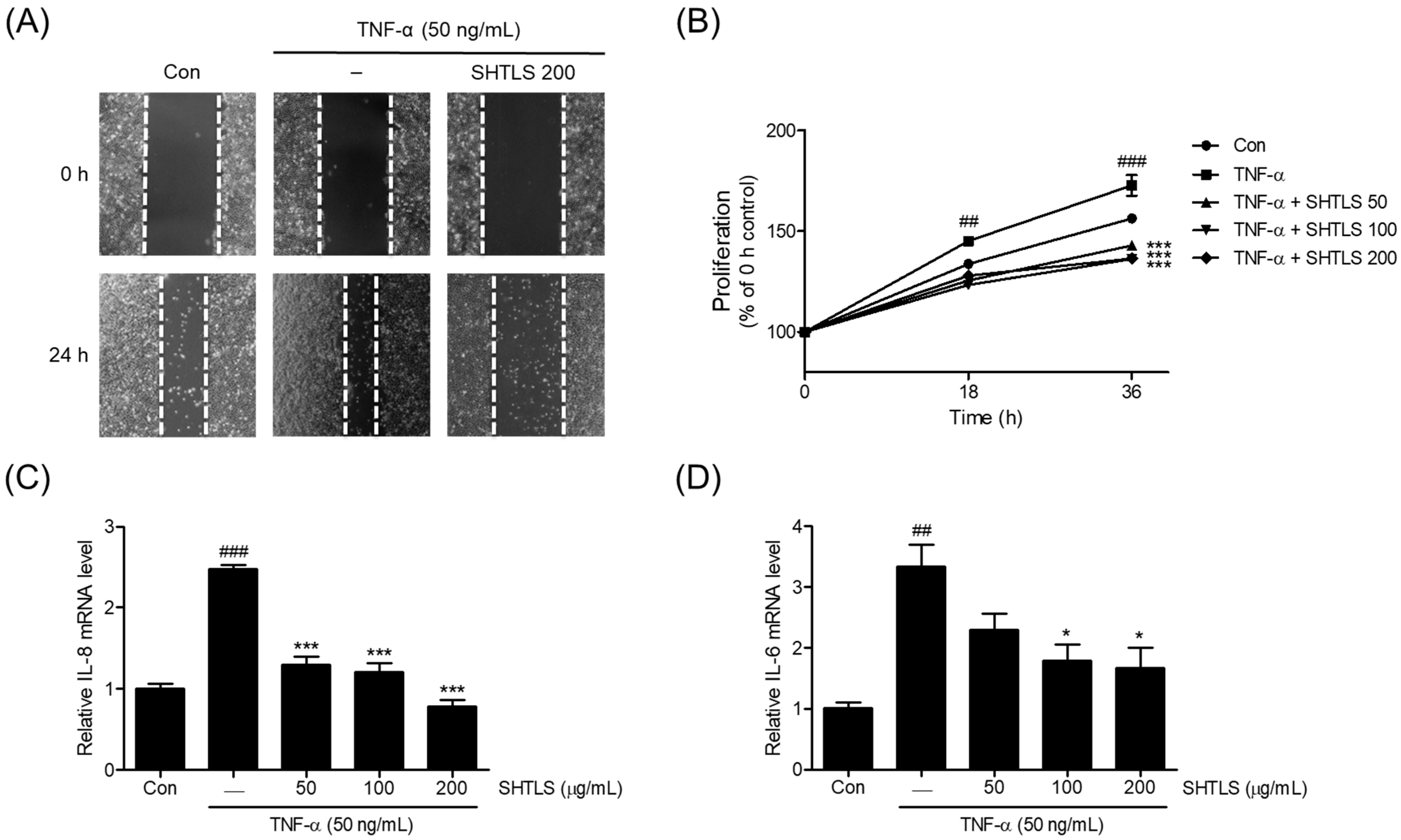
2.6. Anti-Angiogenic Effect of Sinhyotaklisan (SHTLS) in Tumor Necrosis Factor-α (TNF-α)-Stimulated HaCaT Keratinocytes
2.7. Anti-Proliferative Effect of Sinhyotaklisan (SHTLS) in Tumor Necrosis Factor-α (TNF-α)-Stimulated HaCaT Keratinocytes
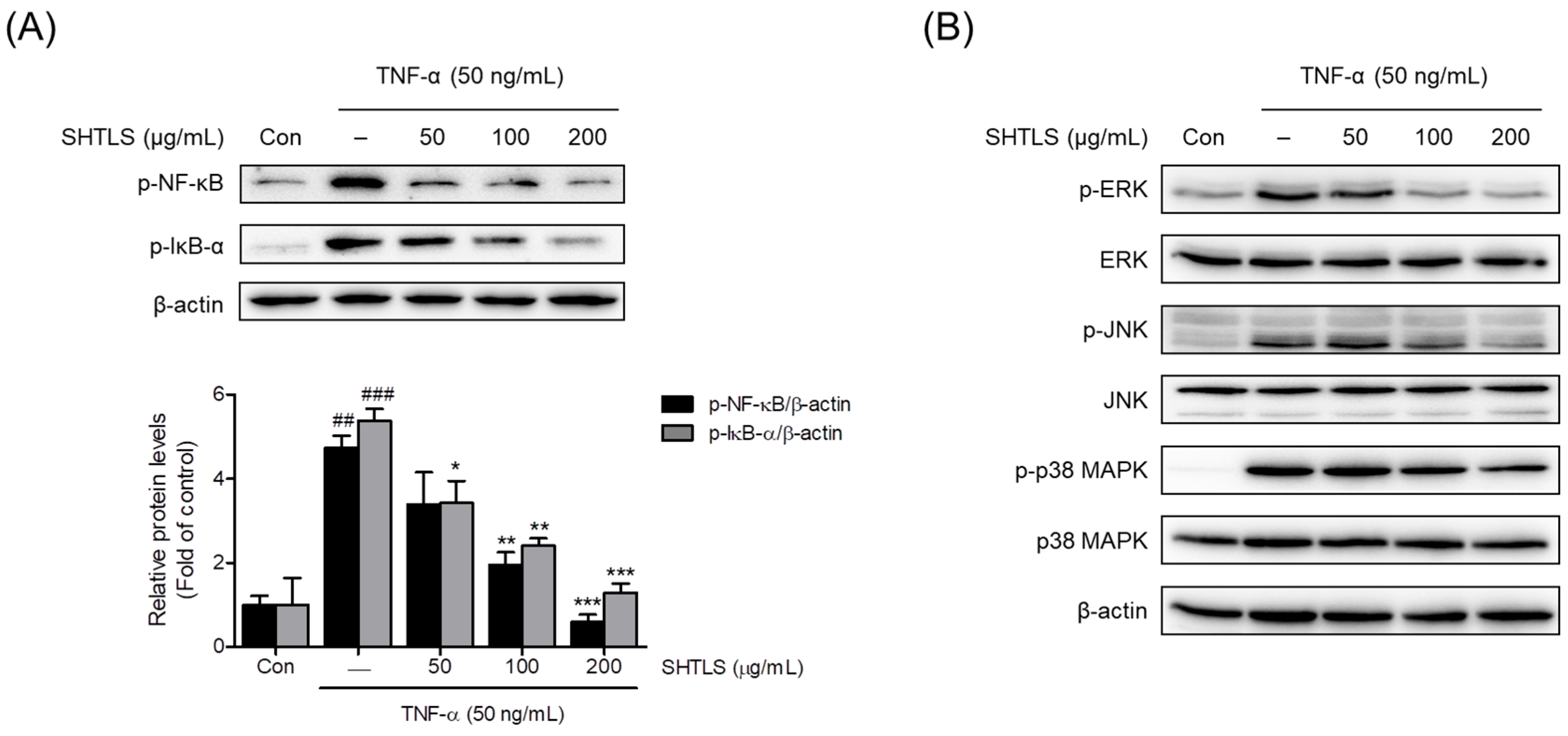
2.8. Modulation of NF-κB and Mitogen-Activated Protein Kinase (MAPK) Signaling Pathways by Sinhyotaklisan (SHTLS) in Tumor Necrosis Factor-α (TNF-α)-Stimulated HaCaT Keratinocytes
2.9. Anti-Psoriatic Effect of Sinhyotaklisan (SHTLS) in an Imiquimod (IMQ)-Induced Psoriasis-like Mouse Model
3. Discussion
4. Materials and Methods
4.1. Acquisition of Active Ingredients and Targets Using Online Databases
4.2. Construction of a Protein–Protein Interaction (PPI) Network and a Compound-Target-Pathway Network
4.3. Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Enrichment Analyses
4.4. ClueGO Biological Process (BP) Term Enrichment Analysis of Key Targets
4.5. Chemicals and Reagents
4.6. Preparation of Sinhyotaklisan (SHTLS) Extract
4.7. Cell Culture
4.8. Cell Cytotoxicity
4.9. Cell Proliferation Assay
4.10. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.11. Western Blot Analysis
4.12. Animals
4.13. Induction of Psoriasis-like Lesions and Efficacy Testing in C57BL/6 Mice
- Control group: Petrolatum cream and PBS (vehicle).
- IMQ group: 5% IMQ cream and PBS (vehicle).
- SHTLS 250 group: 5% IMQ cream and SHTLS 250 mg/kg.
- SHTLS 500 group: 5% IMQ cream and SHTLS 500 mg/kg.
- SHTLS 1000 group: 5% IMQ cream and SHTLS 1000 mg/kg.
4.14. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ADME | Absorption, distribution, metabolism, and excretion |
| BC | Betweenness centrality |
| BP | Biological process |
| DL | Drug-likeness |
| DMEM | Dulbecco’s Modified Eagle’s Medium |
| ERK | Extracellular signal-regulated kinase |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| GO | Gene Ontology |
| H&E | Hematoxylin and eosin |
| HIF-1 | Hypoxia-inducible factor 1 |
| HO-1 | Heme oxygenase-1 |
| HPLC | High-performance liquid chromatography |
| IFN-γ | Interferon gamma |
| IL-6 | Interleukin-6 |
| IL-8 | Interleukin-8 |
| IMQ | Imiquimod |
| iNOS | Inducible nitric oxide synthase |
| JAK | Janus kinase |
| JNK | c-Jun N-terminal kinase |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| LPS | Lipopolysaccharide |
| MAPK | Mitogen-activated protein kinase |
| MMP | Matrix metalloproteinase |
| MTT | 3-[4,5-dimethylthiazole-2-yl]-2,5-diphenyltetrazolium bromide |
| NF-κB | Nuclear factor-kappa B |
| OB | Oral bioavailability |
| PASI | Psoriasis Area and Severity Index |
| PDE4 | Phosphodiesterase 4 |
| PPI | Protein–protein interaction |
| qRT-PCR | Quantitative real-time polymerase chain reaction |
| ROS | Reactive oxygen species |
| SEM | Standard error of the mean |
| SHTLS | Sinhyotaklisan |
| STAT | Signal transducer and activator of transcription |
| TCMSP | Traditional Chinese Medicine Systems Pharmacology Database and Analysis Platform |
| TNF-α | Tumor necrosis factor-α |
| VEGF | Vascular endothelial growth factor |
References
- Smith, C.H.; Barker, J. Psoriasis and its management. BMJ 2006, 333, 380–384. [Google Scholar] [CrossRef]
- Iskandar, I.; Parisi, R.; Griffiths, C.; Ashcroft, D.; Atlas, G.P. Systematic review examining changes over time and variation in the incidence and prevalence of psoriasis by age and gender. Br. J. Dermatol. 2021, 184, 243–258. [Google Scholar] [CrossRef] [PubMed]
- Guillet, C.; Seeli, C.; Nina, M.; Maul, L.V.; Maul, J.-T. The impact of gender and sex in psoriasis: What to be aware of when treating women with psoriasis. Int. J. Women’s Dermatol. 2022, 8, e010. [Google Scholar] [CrossRef]
- Damiani, G.; Bragazzi, N.L.; Karimkhani Aksut, C.; Wu, D.; Alicandro, G.; McGonagle, D.; Guo, C.; Dellavalle, R.; Grada, A.; Wong, P. The global, regional, and national burden of psoriasis: Results and insights from the global burden of disease 2019 study. Front. Med. 2021, 8, 743180. [Google Scholar] [CrossRef]
- Rapalli, V.K.; Singhvi, G.; Dubey, S.K.; Gupta, G.; Chellappan, D.K.; Dua, K. Emerging landscape in psoriasis management: From topical application to targeting biomolecules. Biomed. Pharmacother. 2018, 106, 707–713. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Li, H.; Liu, Y.; Zhu, Z.; Wei, Q. Quercitrin extracted from Tartary buckwheat alleviates imiquimod-induced psoriasis-like dermatitis in mice by inhibiting the Th17 cell response. J. Funct. Foods 2017, 38, 9–19. [Google Scholar] [CrossRef]
- Zhu, B.; Jing, M.; Yu, Q.; Ge, X.; Yuan, F.; Shi, L. Treatments in psoriasis: From standard pharmacotherapy to nanotechnology therapy. Adv. Dermatol. Allergol. Postępy Dermatol. I Alergol. 2022, 39, 460–471. [Google Scholar] [CrossRef]
- Schön, M.P. Adaptive and innate immunity in psoriasis and other inflammatory disorders. Front. Immunol. 2019, 10, 1764. [Google Scholar] [CrossRef]
- Xu, F.; Xu, J.; Xiong, X.; Deng, Y. Salidroside inhibits MAPK, NF-κB, and STAT3 pathways in psoriasis-associated oxidative stress via SIRT1 activation. Redox Rep. 2019, 24, 70–74. [Google Scholar] [CrossRef]
- Zhou, Q.; Mrowietz, U.; Rostami-Yazdi, M. Oxidative stress in the pathogenesis of psoriasis. Free Radic. Biol. Med. 2009, 47, 891–905. [Google Scholar] [CrossRef]
- Marcus, E.; Demmler, D.; Rudolph, A.; Fischer, M. Bacterial colonization of psoriasis plaques. Is it relevant? Dermatol. Rep. 2011, 3, e14. [Google Scholar] [CrossRef] [PubMed]
- Afonina, I.S.; Van Nuffel, E.; Beyaert, R. Immune responses and therapeutic options in psoriasis. Cell. Mol. Life Sci. 2021, 78, 2709–2727. [Google Scholar] [CrossRef] [PubMed]
- Alenzi, F.Q. Links between apoptosis, proliferation and the cell cycle. Br. J. Biomed. Sci. 2004, 61, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Han, J.; Zhao, Y.; Zhu, Q.; Hu, J. Curcumin induces apoptosis in tumor necrosis factor-alpha-treated HaCaT cells. Int. Immunopharmacol. 2012, 13, 170–174. [Google Scholar] [CrossRef]
- Tse, W.-P.; Che, C.-T.; Liu, K.; Lin, Z.-X. Evaluation of the anti-proliferative properties of selected psoriasis-treating Chinese medicines on cultured HaCaT cells. J. Ethnopharmacol. 2006, 108, 133–141. [Google Scholar] [CrossRef]
- Takematsu, H.; Tagami, H. Mode of release of interleukin-8 from proliferating human epidermal keratinocytes in vitro. Exp. Dermatol. 1993, 2, 121–124. [Google Scholar] [CrossRef]
- Steude, J.; Kulke, R.; Christophers, E. Interleukin-1-stimulated secretion of interleukin-8 and growth-related oncogene-α demonstrates greatly enhanced keratinocyte growth in human raft cultured epidermis. J. Investig. Dermatol. 2002, 119, 1254–1260. [Google Scholar] [CrossRef]
- Varricchi, G.; Granata, F.; Loffredo, S.; Genovese, A.; Marone, G. Angiogenesis and lymphangiogenesis in inflammatory skin disorders. J. Am. Acad. Dermatol. 2015, 73, 144–153. [Google Scholar] [CrossRef]
- Duffy, A.M.; Bouchier-Hayes, D.J.; Harmey, J.H. Vascular endothelial growth factor (VEGF) and its role in non-endothelial cells: Autocrine signalling by VEGF. In Madame Curie Bioscience Database [Internet]; Landes Bioscience: Austin, TX, USA, 2013. [Google Scholar]
- Marina, M.E.; Roman, I.I.; Constantin, A.-M.; Mihu, C.M.; Tătaru, A.D. VEGF involvement in psoriasis. Clujul Med. 2015, 88, 247. [Google Scholar] [CrossRef]
- Canavese, M.; Altruda, F.; Ruzicka, T.; Schauber, J. Vascular endothelial growth factor (VEGF) in the pathogenesis of psoriasis—A possible target for novel therapies? J. Dermatol. Sci. 2010, 58, 171–176. [Google Scholar] [CrossRef]
- Federman, D.G.; Froelich, C.W.; Kirsner, R.S. Topical psoriasis therapy. Am. Fam. Physician 1999, 59, 957–962. [Google Scholar] [PubMed]
- Abma, E.; Blanken, R.; De Heide, L. Cushing’s syndrome caused by topical steroid therapy for psoriasis. Neth. J. Med. 2002, 60, 148–150. [Google Scholar]
- Kim, D.H.; Jung, J.M. Topical therapy for psoriasis. J. Korean Soc. Psoriasis 2024, 21, 5–10. [Google Scholar] [CrossRef]
- Heo, J. Dongeuibogam; Namsandang: Seoul, Republic of Korea, 1966. [Google Scholar]
- Ahn, J.-Y.; Lim, D.-W.; Park, S.Y.; Lee, J.-H. Sinhyotaklisan alleviates inflammation in LPS-activated macrophages by modulating the heme oxygenase pathway. J. Ethnopharmacol. 2025, 344, 119548. [Google Scholar] [CrossRef] [PubMed]
- Sahu, N.; Madan, S.; Walia, R.; Tyagi, R.; Fantoukh, O.I.; Hawwal, M.F.; Akhtar, A.; Almarabi, I.; Alam, P.; Saxena, S. Multi-target mechanism of Solanum xanthocarpum for treatment of psoriasis based on network pharmacology and molecular docking. Saudi Pharm. J. 2023, 31, 101788. [Google Scholar] [CrossRef]
- Kuai, L.; Song, J.-k.; Zhang, R.-x.; Xing, M.; Luo, Y.; Ru, Y.; Ding, X.-j.; Liu, L.; Lu, Y.; Sun, X.-y. Uncovering the mechanism of Jueyin granules in the treatment of psoriasis using network pharmacology. J. Ethnopharmacol. 2020, 262, 113214. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Liu, Z.; Li, M.; Du, L.; Zhu, X.; Leng, Y.; Han, C.; Xu, Q.; Zhang, C. Based on UPLC-Q-TOF/MS and Network Pharmacology to Explore the Mechanism of Qingre Lishi Decoction in the Treatment of Psoriasis. Drug Des. Devel. Ther. 2024, 18, 3871–3889. [Google Scholar] [CrossRef]
- Guo, J.; Zhang, H.; Lin, W.; Lu, L.; Su, J.; Chen, X. Signaling pathways and targeted therapies for psoriasis. Signal Transduct. Target. Ther. 2023, 8, 437. [Google Scholar] [CrossRef]
- Zhou, X.; Chen, Y.; Cui, L.; Shi, Y.; Guo, C. Advances in the pathogenesis of psoriasis: From keratinocyte perspective. Cell Death Dis. 2022, 13, 81. [Google Scholar] [CrossRef]
- Lizzul, P.F.; Aphale, A.; Malaviya, R.; Sun, Y.; Masud, S.; Dombrovskiy, V.; Gottlieb, A.B. Differential expression of phosphorylated NF-κB/RelA in normal and psoriatic epidermis and downregulation of NF-κB in response to treatment with etanercept. J. Investig. Dermatol. 2005, 124, 1275–1283. [Google Scholar] [CrossRef]
- Akhtar, T.; Wani, W.Y.; Kamal, M.A.; Kaur, R. Role of angiogenic growth factors in psoriasis: A review. Curr. Drug Metab. 2018, 19, 910–916. [Google Scholar] [CrossRef] [PubMed]
- Schön, M.P.; Wilsmann-Theis, D. Current developments and perspectives in psoriasis. JDDG J. Der Dtsch. Dermatol. Ges. 2023, 21, 363–372. [Google Scholar] [CrossRef]
- Gordon, K.B.; Foley, P.; Krueger, J.G.; Pinter, A.; Reich, K.; Vender, R.; Vanvoorden, V.; Madden, C.; White, K.; Cioffi, C. Bimekizumab efficacy and safety in moderate to severe plaque psoriasis (BE READY): A multicentre, double-blind, placebo-controlled, randomised withdrawal phase 3 trial. Lancet 2021, 397, 475–486. [Google Scholar] [CrossRef]
- Ibezim, A.; Onah, E.; Dim, E.N.; Ntie-Kang, F. A computational multi-targeting approach for drug repositioning for psoriasis treatment. BMC Complement. Med. Ther. 2021, 21, 193. [Google Scholar] [CrossRef] [PubMed]
- Brotas, A.M.; Cunha, J.M.T.; Lago, E.H.J.; Machado, C.C.N.; Carneiro, S.C.d.S. Tumor necrosis factor-alpha and the cytokine network in psoriasis. An. Bras. Dermatol. 2012, 87, 673–683. [Google Scholar] [CrossRef]
- Roy, B.; Simard, M.; Lorthois, I.; Bélanger, A.; Maheux, M.; Duque-Fernandez, A.; Rioux, G.; Simard, P.; Deslauriers, M.; Masson, L.-C. In vitro models of psoriasis. In Skin Tissue Models; Elsevier: Amsterdam, The Netherlands, 2018; pp. 103–128. [Google Scholar]
- Luo, T.; Ma, Y.; Wei, W. Murine models of psoriasis and its applications in drug development. J. Pharmacol. Toxicol. Methods 2020, 101, 106657. [Google Scholar] [CrossRef]
- Ghoreschi, K.; Balato, A.; Enerbäck, C.; Sabat, R. Therapeutics targeting the IL-23 and IL-17 pathway in psoriasis. Lancet 2021, 397, 754–766. [Google Scholar] [CrossRef]
- Madonna, S.; Girolomoni, G.; Dinarello, C.A.; Albanesi, C. The significance of IL-36 hyperactivation and IL-36R targeting in psoriasis. Int. J. Mol. Sci. 2019, 20, 3318. [Google Scholar] [CrossRef] [PubMed]
- Chiricozzi, A.; Guttman-Yassky, E.; Suárez-Farinas, M.; Nograles, K.E.; Tian, S.; Cardinale, I.; Chimenti, S.; Krueger, J.G. Integrative responses to IL-17 and TNF-α in human keratinocytes account for key inflammatory pathogenic circuits in psoriasis. J. Investig. Dermatol. 2011, 131, 677–687. [Google Scholar] [CrossRef]
- Locksley, R.M.; Killeen, N.; Lenardo, M.J. The TNF and TNF receptor superfamilies: Integrating mammalian biology. Cell 2001, 104, 487–501. [Google Scholar] [CrossRef]
- Trompezinski, S.; Denis, A.; Schmitt, D.; Viac, J. Comparative effects of polyphenols from green tea (EGCG) and soybean (genistein) on VEGF and IL-8 release from normal human keratinocytes stimulated with the proinflammatory cytokine TNFα. Arch. Dermatol. Res. 2003, 295, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Weng, Z.; Patel, A.B.; Vasiadi, M.; Therianou, A.; Theoharides, T.C. Luteolin inhibits human keratinocyte activation and decreases NF-κB induction that is increased in psoriatic skin. PLoS ONE 2014, 9, e90739. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Tsirka, S.E. Neuroprotection by inhibition of matrix metalloproteinases in a mouse model of intracerebral haemorrhage. Brain 2005, 128, 1622–1633. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Jackson, C.J. Autocrine actions of matrix metalloproteinase (MMP)-2 counter the effects of MMP-9 to promote survival and prevent terminal differentiation of cultured human keratinocytes. J. Investig. Dermatol. 2008, 128, 2676–2685. [Google Scholar] [CrossRef]
- Manna, S.K.; Ramesh, G.T. Interleukin-8 induces nuclear transcription factor-κB through a TRAF6-dependent pathway. J. Biol. Chem. 2005, 280, 7010–7021. [Google Scholar] [CrossRef]
- Yang, J.-H.; Do, H.J.; Lee, E.; Yim, N.-H.; Cho, W.-K.; Park, K.-I.; Ma, J.Y. Jageum-Jung improves 2, 4-dinitrochlorobenzene-induced atopic dermatitis-like skin lesions in mice and suppresses pro-inflammatory chemokine production by inhibiting TNF-α/IFN-γ-induced STAT-1 and NFκB signaling in HaCaT cells. J. Ethnopharmacol. 2018, 221, 48–55. [Google Scholar] [CrossRef]
- Yang, X.-H.; Man, X.-Y.; Cai, S.-Q.; Yao, Y.-G.; Bu, Z.-Y.; Zheng, M. Expression of VEGFR-2 on HaCaT cells is regulated by VEGF and plays an active role in mediating VEGF induced effects. Biochem. Biophys. Res. Commun. 2006, 349, 31–38. [Google Scholar] [CrossRef]
- Kwon, Y.-W.; Kwon, K.-S.; Moon, H.-E.; Park, J.A.; Choi, K.-S.; Kim, Y.-S.; Jang, H.-S.; Oh, C.-K.; Lee, Y.-M.; Kwon, Y.-G. Insulin-like growth factor-II regulates the expression of vascular endothelial growth factor by the human keratinocyte cell line HaCaT. J. Investig. Dermatol. 2004, 123, 152–158. [Google Scholar] [CrossRef]
- Rezvani, H.R.; Ali, N.; Nissen, L.J.; Harfouche, G.; De Verneuil, H.; Taïeb, A.; Mazurier, F. HIF-1α in epidermis: Oxygen sensing, cutaneous angiogenesis, cancer, and non-cancer disorders. J. Investig. Dermatol. 2011, 131, 1793–1805. [Google Scholar] [CrossRef]
- Koukourakis, M.I.; Giatromanolaki, A.; Sivridis, E.; Simopoulos, C.; Turley, H.; Talks, K.; Gatter, K.C.; Harris, A.L.; The Tumour and Angiogenesis Research Group. Hypoxia-inducible factor (HIF1A and HIF2A), angiogenesis, and chemoradiotherapy outcome of squamous cell head-and-neck cancer. Int. J. Radiat. Oncol. Biol. Phys. 2002, 53, 1192–1202. [Google Scholar] [CrossRef]
- Jia, Z.; Wang, X.; Wang, X.; Wei, P.; Li, L.; Wu, P.; Hong, M. Calycosin alleviates allergic contact dermatitis by repairing epithelial tight junctions via down-regulating HIF-1α. J. Cell. Mol. Med. 2018, 22, 4507–4521. [Google Scholar] [CrossRef] [PubMed]
- Leire, E.; Olson, J.; Isaacs, H.; Nizet, V.; Hollands, A. Role of hypoxia inducible factor-1 in keratinocyte inflammatory response and neutrophil recruitment. J. Inflamm. 2013, 10, 28. [Google Scholar] [CrossRef] [PubMed]
- Son, D.J.; Jung, Y.Y.; Park, M.H.; Lee, H.L.; Song, M.J.; Yoo, H.-S.; Hwang, D.Y.; Han, S.B.; Hong, J.T. Activated natural killer cells mediate the suppressive effect of interleukin-4 on tumor development via STAT6 activation in an atopic condition melanoma model. Neoplasia 2017, 19, 537–548. [Google Scholar] [CrossRef]
- Hamadeh, H.K.; Vargas, M.; Lee, E.; Menzel, D.B. Arsenic disrupts cellular levels of p53 and mdm2: A potential mechanism of carcinogenesis. Biochem. Biophys. Res. Commun. 1999, 263, 446–449. [Google Scholar] [CrossRef]
- Colombo, I.; Sangiovanni, E.; Maggio, R.; Mattozzi, C.; Zava, S.; Corbett, Y.; Fumagalli, M.; Carlino, C.; Corsetto, P.A.; Scaccabarozzi, D. HaCaT cells as a reliable in vitro differentiation model to dissect the inflammatory/repair response of human keratinocytes. Mediat. Inflamm. 2017, 2017, 7435621. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.B.; Jacob, S. A simple practice guide for dose conversion between animals and human. J. Basic Clin. Pharm. 2016, 7, 27–31. [Google Scholar] [CrossRef]
- Shinno-Hashimoto, H.; Eguchi, A.; Sakamoto, A.; Wan, X.; Hashimoto, Y.; Fujita, Y.; Mori, C.; Hatano, M.; Matsue, H.; Hashimoto, K. Effects of splenectomy on skin inflammation and psoriasis-like phenotype of imiquimod-treated mice. Sci. Rep. 2022, 12, 14738. [Google Scholar] [CrossRef]
- Ru, J.; Li, P.; Wang, J.; Zhou, W.; Li, B.; Huang, C.; Li, P.; Guo, Z.; Tao, W.; Yang, Y. TCMSP: A database of systems pharmacology for drug discovery from herbal medicines. J. Cheminform. 2014, 6, 13. [Google Scholar] [CrossRef]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.-H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef]
- Kim, C.H.; Yoo, J.K.; Jeon, S.H.; Lim, C.Y.; Lee, J.H.; Koo, D.B.; Park, M.Y. Anti-psoriatic effect of myeloid-derived suppressor cells on imiquimod-induced skin inflammation in mice. Scand. J. Immunol. 2019, 89, e12742. [Google Scholar] [CrossRef]
- Yoo, J.K.; Choo, Y.-K.; Kwak, D.H.; Lee, J.M.; Lim, C.-Y.; Lee, J.-H.; Park, M.-Y.; Kim, C.-H. Protective effects of agonistic anti-4-1BB antibody on the development of imiquimod-induced psoriasis-like dermatitis in mice. Immunol. Lett. 2016, 178, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Morris, S.A.; Ananthapadmanabhan, K.P.; Kasting, G.B. Anionic surfactant–induced changes in skin permeability. J. Pharm. Sci. 2019, 108, 3640–3648. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Degree | Betweenness Centrality |
|---|---|---|
| AKT1 | 126 | 0.038798 |
| TNF | 125 | 0.036107 |
| IL-6 | 122 | 0.028189 |
| TP53 | 120 | 0.026442 |
| ALB | 119 | 0.034818 |
| IL1B | 115 | 0.023147 |
| CASP3 | 110 | 0.012303 |
| PTGS2 | 110 | 0.02313 |
| ESR1 | 109 | 0.022367 |
| JUN | 109 | 0.015067 |
| MMP9 | 108 | 0.016962 |
| HIF1A | 107 | 0.011356 |
| BCL2 | 106 | 0.010885 |
| EGFR | 106 | 0.02012 |
| STAT3 | 105 | 0.010239 |
| PPARG | 103 | 0.01913 |
| MYC | 100 | 0.013551 |
| CTNNB1 | 97 | 0.010247 |
| MAPK3 | 97 | 0.01432 |
| TGFB1 | 95 | 0.00717 |
| FOS | 92 | 0.014389 |
| IFNG | 92 | 0.007102 |
| CCL2 | 92 | 0.008057 |
| HSP90AA1 | 90 | 0.01548 |
| CCND1 | 89 | 0.007204 |
| IL-10 | 88 | 0.007228 |
| CXCL8 (IL8) | 87 | 0.006118 |
| ERBB2 | 86 | 0.007181 |
| EGF | 86 | 0.004684 |
| ICAM1 | 82 | 0.013478 |
| IL1A | 82 | 0.006435 |
| HMOX1 | 79 | 0.004625 |
| RELA | 77 | 0.005654 |
| NFE2L2 | 73 | 0.005323 |
| PPARA | 70 | 0.007071 |
| HSPA5 | 66 | 0.006927 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahn, J.-Y.; Lim, D.-W.; Kim, J.-H.; Park, S.-Y.; Park, S.-D.; Lee, J.-H. Sinhyotaklisan as a Potential Therapeutic for Psoriasis: Network Pharmacology and Experimental Validation. Int. J. Mol. Sci. 2025, 26, 5082. https://doi.org/10.3390/ijms26115082
Ahn J-Y, Lim D-W, Kim J-H, Park S-Y, Park S-D, Lee J-H. Sinhyotaklisan as a Potential Therapeutic for Psoriasis: Network Pharmacology and Experimental Validation. International Journal of Molecular Sciences. 2025; 26(11):5082. https://doi.org/10.3390/ijms26115082
Chicago/Turabian StyleAhn, Jung-Yun, Dong-Woo Lim, Jin-Hee Kim, Sung-Yun Park, Sun-Dong Park, and Ju-Hee Lee. 2025. "Sinhyotaklisan as a Potential Therapeutic for Psoriasis: Network Pharmacology and Experimental Validation" International Journal of Molecular Sciences 26, no. 11: 5082. https://doi.org/10.3390/ijms26115082
APA StyleAhn, J.-Y., Lim, D.-W., Kim, J.-H., Park, S.-Y., Park, S.-D., & Lee, J.-H. (2025). Sinhyotaklisan as a Potential Therapeutic for Psoriasis: Network Pharmacology and Experimental Validation. International Journal of Molecular Sciences, 26(11), 5082. https://doi.org/10.3390/ijms26115082