
The Significance of Neuropilins in Gastrointestinal Cancers

 , ,
, ,  and
and 
Abstract
1. Introduction
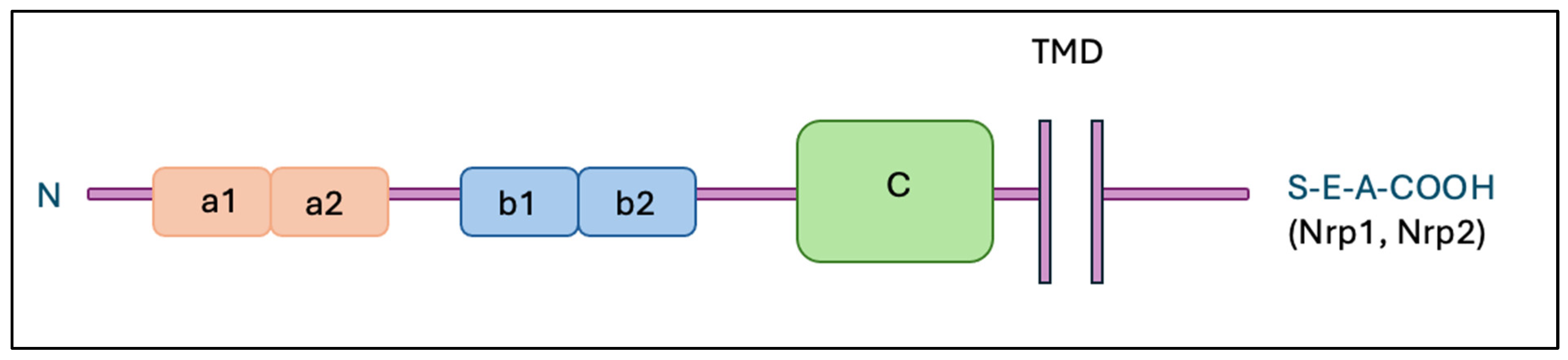
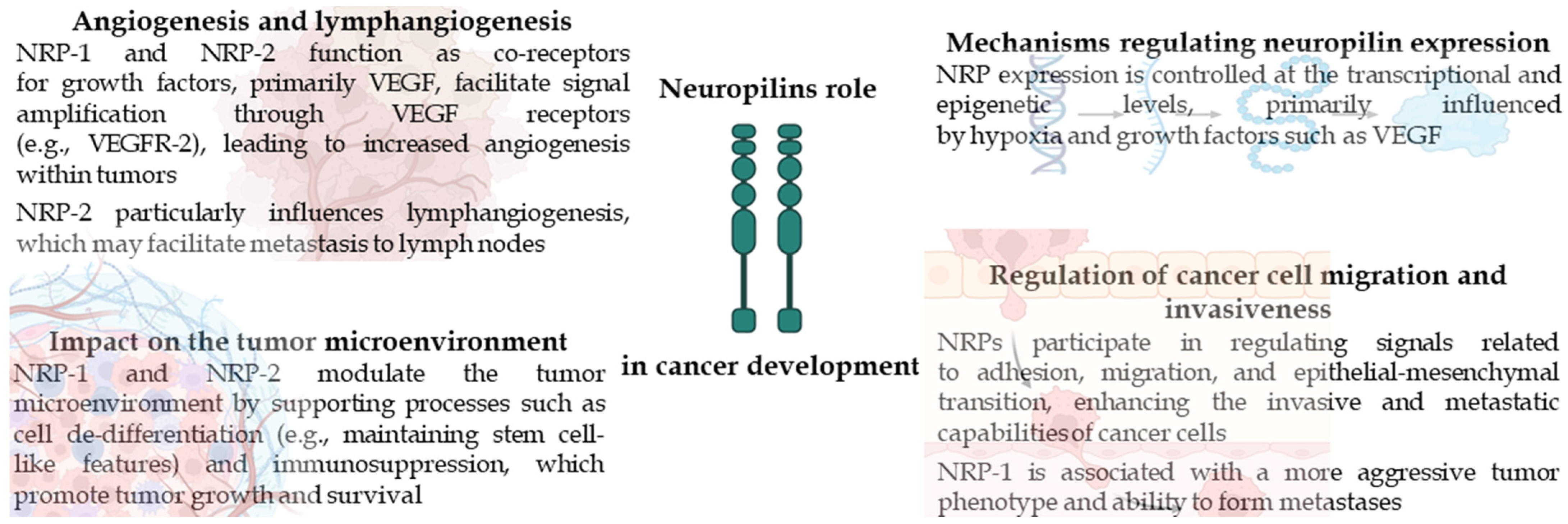
2. Neuropilins
3. Neuropilins in Colorectal Cancer
4. Neuropilins in Gastric Cancer
5. Neuropilins in Pancreatic Cancer
6. Neuropilins in Liver Cancer
7. Non-Coding RNA and NRP
8. Summary
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Song, M.; Garrett, W.S.; Chan, A.T. Nutrients, foods, and colorectal cancer prevention. Gastroenterology 2015, 148, 1244–1260.e16. [Google Scholar] [CrossRef] [PubMed]
- Duffy, M.; Lamerz, R.; Haglund, C.; Nicolini, A.; Kalousová, M.; Holubec, L.; Sturgeon, C. Tumor markers in colorectal cancer, gastric cancer and gastrointestinal stromal cancers: European group on tumor markers 2014 guidelines update. Int. J. Cancer 2014, 134, 2513–2522. [Google Scholar] [CrossRef] [PubMed]
- Ellis, L.M. The role of neuropilins in cancer. Mol. Cancer Ther. 2006, 5, 1099–1107. [Google Scholar] [CrossRef] [PubMed]
- Rizzolio, S.; Tamagnone, L. Multifaceted Role of Neuropilins in Cancer. Curr. Med. Chem. 2011, 18, 3563–3575. [Google Scholar] [CrossRef]
- Pereira, E.R.; Jones, D.; Jung, K.; Padera, T.P. The lymph node microenvironment and its role in the progression of metastatic cancer. Semin. Cell Dev. Biol. 2015, 38, 98–105. [Google Scholar] [CrossRef]
- Grünewald, F.S.; Prota, A.E.; Giese, A.; Ballmer-Hofer, K. Structure–function analysis of VEGF receptor activation and the role of coreceptors in angiogenic signaling. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2010, 1804, 567–580. [Google Scholar] [CrossRef]
- Roth, L.; Nasarre, C.; Dirrig-Grosch, S.; Aunis, D.; Crémel, G.; Hubert, P.; Bagnard, D. Transmembrane Domain Interactions Control Biological Functions of Neuropilin-1. Mol. Biol. Cell 2008, 19, 646–654. [Google Scholar] [CrossRef]
- Bielenberg, D.R.; Zetter, B.R. The Contribution of Angiogenesis to the Process of Metastasis. Cancer J. 2015, 21, 267–273. [Google Scholar] [CrossRef]
- Wang, L.; Dutta, S.K.; Kojima, T.; Xu, X.; Khosravi-Far, R.; Ekker, S.C.; Mukhopadhyay, D. Neuropilin-1 Modulates p53/Caspases Axis to Promote Endothelial Cell Survival. PLoS ONE 2007, 2, e1161. [Google Scholar] [CrossRef]
- Wild, J.R.L.; Staton, C.A.; Chapple, K.; Corfe, B.M. Neuropilins: Expression and roles in the epithelium. Int. J. Exp. Pathol. 2012, 93, 81–103. [Google Scholar] [CrossRef]
- Mendes-Da-Cruz, D.A.; Linhares-Lacerda, L.; Smaniotto, S.; Dardenne, M.; Savino, W. Semaphorins and neuropilins: New players in the neuroendocrine control of the intrathymic T-cell migration in humans. Exp. Physiol. 2012, 97, 1146–1150. [Google Scholar] [CrossRef] [PubMed]
- Grage-Griebenow, E.; Löseke, S.; Kauth, M.; Gehlhar, K.; Zawatzky, R.; Bufe, A. Anti-BDCA-4 (neuropilin-1) antibody can suppress virus-induced IFN-alpha production of plasmacytoid dendritic cells. Immunol. Cell Biol. 2007, 85, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Solomon, B.D.; Mueller, C.; Chae, W.-J.; Alabanza, L.M.; Bynoe, M.S. Neuropilin-1 attenuates autoreactivity in experimental autoimmune encephalomyelitis. Proc. Natl. Acad. Sci. USA 2011, 108, 2040–2045. [Google Scholar] [CrossRef] [PubMed]
- Pan, Q.; Chanthery, Y.; Liang, W.-C.; Stawicki, S.; Mak, J.; Rathore, N.; Tong, R.K.; Kowalski, J.; Yee, S.F.; Pacheco, G.; et al. Blocking Neuropilin-1 Function Has an Additive Effect with Anti-VEGF to Inhibit Tumor Growth. Cancer Cell 2007, 11, 53–67. [Google Scholar] [CrossRef]
- Appleton, B.A.; Wu, P.; Maloney, J.; Yin, J.; Liang, W.-C.; Stawicki, S.; Mortara, K.; Bowman, K.K.; Elliott, J.M.; Desmarais, W.; et al. Structural studies of neuropilin/antibody complexes provide insights into semaphorin and VEGF binding. EMBO J. 2007, 26, 4902–4912. [Google Scholar] [CrossRef]
- Yaqoob, U.; Cao, S.; Shergill, U.; Jagavelu, K.; Geng, Z.; Yin, M.; De Assuncao, T.M.; Cao, Y.; Szabolcs, A.; Thorgeirsson, S.; et al. Neuropilin-1 stimulates tumor growth by increasing fibronectin fibril assembly in the tumor microenvironment. Cancer Res. 2012, 72, 4047–4059. [Google Scholar] [CrossRef]
- Sugahara, K.N.; Teesalu, T.; Prakash Karmali, P.; Ramana Kotamraju, V.; Agemy, L.; Greenwald, D.R.; Ruoslahti, E. Coadministration of a Tumor-Penetrating Peptide Enhances the Efficacy of Cancer Drugs. Science 2010, 328, 1031–1035. [Google Scholar] [CrossRef]
- Parikh, A.A.; Fan, F.; Liu, W.B.; Ahmad, S.A.; Stoeltzing, O.; Reinmuth, N.; Bielenberg, D.; Bucana, C.D.; Klagsbrun, M.; Ellis, L.M. Neuropilin-1 in human colon cancer: Expression, regulation, and role in induction of angiogenesis. Am. J. Pathol. 2004, 164, 2139–2151. [Google Scholar] [CrossRef]
- Fernández-Palanca, P.; Payo-Serafín, T.; Fondevila, F.; Méndez-Blanco, C.; San-Miguel, B.; Romero, M.R.; Tuñón, M.J.; Marin, J.J.G.; González-Gallego, J.; Mauriz, J.L. Neuropilin-1 as a Potential Biomarker of Prognosis and Invasive-Related Parameters in Liver and Colorectal Cancer: A Systematic Review and Meta-Analysis of Human Studies. Cancers 2022, 14, 3455. [Google Scholar] [CrossRef]
- Grandclement, C.; Pallandre, J.R.; Valmary Degano, S.; Viel, E.; Bouard, A.; Balland, J.; Remy-Martin, J.P.; Simon, B.; Rouleau, A.; Boireau, W.; et al. Neuropilin-2 expression promotes TGF-β1-mediated epithelial to mesenchymal transition in colorectal cancer cells. PLoS ONE 2011, 6, e20444. [Google Scholar] [CrossRef]
- Geretti, E.; van Meeteren, L.A.; Shimizu, A.; Dudley, A.C.; Claesson-Welsh, L.; Klagsbrun, M. A mutated soluble neuropilin-2 B domain antagonizes vascular endothelial growth factor bioactivity and inhibits tumor progression. Mol. Cancer Res. 2010, 8, 1063–1073. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Li, Y.; Huang, L.; Bai, B.; Xu, Z. Clinicopathological Significance of Neuropilin 1 Expression in Gastric Cancer: A Meta-Analysis. Dis. Markers 2020, 2020, 4763492. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Zhou, Y.; Wang, L.; Lv, M.; Sun, J.; Luo, Z.; He, J. Sema3A Alleviates the Malignant Behaviors of Gastric Cancer Cells by Inhibiting NRP-1. Curr. Mol. Med. 2024, 24, 931–939. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Zeng, M.-H.; Liao, G.; Qian, K.; Li, H. Neuropilin-1 Interacts with Fibronectin-1 to Promote Epithelial–Mesenchymal Transition Progress in Gastric Cancer. OncoTargets Ther. 2020, 13, 10677–10687. [Google Scholar] [CrossRef]
- Seo, H.S.; Hyeon, J.; Song, I.H.; Lee, H.H. Relationship between neuropilin-1 expression and prognosis, according to gastric cancer histology. Histochem. J. 2020, 51, 199–208. [Google Scholar] [CrossRef]
- Jin, Q.; Ren, Q.; Chang, X.; Yu, H.; Jin, X.; Lu, X.; He, N.; Wang, G. Neuropilin-1 predicts poor prognosis and promotes tumor metastasis through epithelial-mesenchymal transition in gastric cancer. J. Cancer 2021, 12, 3648–3659. [Google Scholar] [CrossRef]
- Ding, Y.; Zhou, J.; Wang, S.; Li, Y.; Mi, Y.; Gao, S.; Xu, Y.; Chen, Y.; Yan, J. Anti-neuropilin-1 monoclonal antibody suppresses the migration and invasion of human gastric cancer cells via Akt dephosphorylation. Exp. Ther. Med. 2018, 16, 537–546. [Google Scholar] [CrossRef]
- Islam, R.; Mishra, J.; Bodas, S.; Bhattacharya, S.; Batra, S.K.; Dutta, S.; Datta, K. Role of Neuropilin-2-mediated signaling axis in cancer progression and therapy resistance. Cancer Metastasis Rev. 2022, 41, 771–787. [Google Scholar] [CrossRef]
- Roy, S.; Bag, A.K.; Dutta, S.; Polavaram, N.S.; Islam, R.; Schellenburg, S.; Banwait, J.; Guda, C.; Ran, S.; Hollingsworth, M.A.; et al. Macrophage-Derived Neuropilin-2 Exhibits Novel Tumor-Promoting Functions. Cancer Res. 2018, 78, 5600–5617. [Google Scholar] [CrossRef]
- Ilic, M.; Ilic, I. Epidemiology of pancreatic cancer. World J. Gastroenterol. 2016, 22, 9694–9705. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- McDonald, O.G. The biology of pancreatic cancer morphology. Pathology 2021, 54, 236–247. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Meng, S.; Hara, T.; Sato, H.; Tatekawa, S.; Tsuji, Y.; Saito, Y.; Hamano, Y.; Arao, Y.; Gotoh, N.; Ogawa, K.; et al. Revealing neuropilin expression patterns in pancreatic cancer: From single-cell to therapeutic opportunities (Review). Oncol. Lett. 2022, 27, 113. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Piera-Velazquez, S.; Jimenez, S.A. Endothelial to Mesenchymal Transition: Role in Physiology and in the Pathogenesis of Human Diseases. Physiol. Rev. 2019, 99, 1281–1324. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Borchardt, H.; Schulz, A.; Datta, K.; Muders, M.H.; Aigner, A. Silencing of Neuropilins and GIPC1 in pancreatic ductal adenocarcinoma exerts multiple cellular and molecular antitumor effects. Sci. Rep. 2019, 9, 15471. [Google Scholar] [CrossRef]
- Zhou, R.; Curry, J.M.; Roy, L.D.; Grover, P.; Haider, J.; Moore, L.J.; Wu, S.-T.; Kamesh, A.; Yazdanifar, M.; Ahrens, W.A.; et al. A novel association of neuropilin-1 and MUC1 in pancreatic ductal adenocarcinoma: Role in induction of VEGF signaling and angiogenesis. Oncogene 2016, 35, 5608–5618. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Matkar, P.N.; Jong, E.D.; Ariyagunarajah, R.; Prud’Homme, G.J.; Singh, K.K.; Leong-Poi, H. Jack of many trades: Multifaceted role of neuropilins in pancreatic cancer. Cancer Med. 2018, 7, 5036–5046. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Geretti, E.; Klagsbrun, M. Neuropilins. Cell Adhes. Migr. 2007, 1, 56–61. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Czaplicka, A.; Lachota, M.; Pączek, L.; Zagożdżon, R.; Kaleta, B. Chimeric Antigen Receptor T Cell Therapy for Pancreatic Cancer: A Review of Current Evidence. Cells 2024, 13, 101. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- El-Serag, H.B. Hepatocellular carcinoma. N. Engl. J. Med. 2011, 365, 1118–1127. [Google Scholar] [CrossRef] [PubMed]
- Wittmann, P.; Grubinger, M.; Gröger, C.; Huber, H.; Sieghart, W.; Peck-Radosavljevic, M.; Mikulits, W. Neuropilin-2 induced by transforming growth factor-β augments migration of hepatocellular carcinoma cells. BMC Cancer 2015, 15, 909. [Google Scholar] [CrossRef] [PubMed]
- Coulouarn, C.; Corlu, A.; Glaise, D.; Guénon, I.; Thorgeirsson, S.S.; Clément, B. Hepatocyte–Stellate Cell Cross-Talk in the Liver Engenders a Permissive Inflammatory Microenvironment That Drives Progression in Hepatocellular Carcinoma. Cancer Res. 2012, 72, 2533–2542. [Google Scholar] [CrossRef]
- Yan, X.-L.; Fu, C.-J.; Chen, L.; Qin, J.-H.; Zeng, Q.; Yuan, H.-F.; Nan, X.; Chen, H.-X.; Zhou, J.-N.; Lin, Y.-L.; et al. Mesenchymal stem cells from primary breast cancer tissue promote cancer proliferation and enhance mammosphere formation partially via EGF/EGFR/Akt pathway. Breast Cancer Res. Treat. 2012, 132, 153–164. [Google Scholar] [CrossRef]
- Cao, S.; Yaqoob, U.; Das, A.; Shergill, U.; Jagavelu, K.; Huebert, R.C.; Routray, C.; Abdelmoneim, S.; Vasdev, M.; Leof, E.; et al. Neuropilin-1 promotes cirrhosis of the rodent and human liver by enhancing PDGF/TGF-β signaling in hepatic stellate cells. J. Clin. Investig. 2010, 120, 2379–2394. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.B.; Zheng, K.W.; Hu, Y.Y.; Salameen, H.; Zhu, Z.Y.; Wu, F.F.; Ding, X. VEGF/Nrp1/HIF-1α promotes proliferation of hepatocellular carcinoma through a positive feedback loop. Med. Oncol. 2023, 40, 339. [Google Scholar] [CrossRef]
- Xu, Z.C.; Shen, H.X.; Chen, C.; Ma, L.; Li, W.Z.; Wang, L.; Geng, Z.M. Neuropilin-1 promotes primary liver cancer progression by potentiating the activity of hepatic stellate cells. Oncol. Lett. 2018, 15, 2245–2251. [Google Scholar] [CrossRef]
- Elpek, G.Ö. Neuropilins and liver. World J. Gastroenterol. 2015, 21, 7065–7073. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ao, J.; Qiang, N.; Kanzaki, H.; Nakamura, M.; Kakiuchi, R.; Zhang, J.; Kojima, R.; Koroki, K.; Inoue, M.; Kanogawa, N.; et al. Dual effects of targeting neuropilin-1 in lenvatinib-resistant hepatocellular carcinoma: Inhibition of tumor growth and angiogenesis. Am. J. Physiol. Physiol. 2024, 327, C1150–C1161. [Google Scholar] [CrossRef]
- Wu, Q.; Pan, C.; Zhou, Y.; Wang, S.; Xie, L.; Zhou, W.; Ding, L.; Chen, T.; Qian, J.; Su, R.; et al. Targeting neuropilin-1 abolishes anti-PD-1-upregulated regulatory T cells and synergizes with 4-1BB agonist for liver cancer treatment. Hepatology 2023, 78, 1402–1417. [Google Scholar] [CrossRef]
- Dong, X.; Guo, W.; Zhang, S.; Wu, T.; Sun, Z.; Yan, S.; Zheng, S. Elevated expression of neuropilin-2 associated with unfavorable prognosis in hepatocellular carcinoma. OncoTargets Ther. 2017, 10, 3827–3833. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Gene Expression Profiling Interactive Analysis. Available online: http://gepia2.cancer-pku.cn/ (accessed on 15 May 2025).
- Tang, Z.; Kang, B.; Li, C.; Chen, T.; Zhang, Z. GEPIA2: An enhanced web server for large-scale expression profiling and interactive analysis. Nucleic Acids Res. 2019, 47, W556–W560. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Guo, S.; Tang, J.; Wen, J.; Wang, H.; Hu, X.; Gu, Q. MicroRNA-19b-3p suppresses gastric cancer development by negatively regulating neuropilin-1. Cancer Cell Int. 2020, 20, 193. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, L. miR-124-3p inhibits CRC proliferation, migration, and invasion by targeting ITGB1. Discov. Oncol. 2025, 16, 158. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, R.; Wang, M. miR-331-3p suppresses cell invasion and migration in colorectal carcinoma by directly targeting NRP2. Oncol. Lett. 2019, 18, 6501–6508. [Google Scholar] [CrossRef]
- Jin, Y.; Che, X.; Qu, X.; Li, X.; Lu, W.; Wu, J.; Wang, Y.; Hou, K.; Li, C.; Zhang, X.; et al. CircHIPK3 Promotes Metastasis of Gastric Cancer via miR-653-5p/miR-338-3p-NRP1 Axis Under a Long-Term Hypoxic Microenvironment. Front. Oncol. 2020, 10, 1612. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Ligand | NRP-1 | NRP-2 |
|---|---|---|
| VEGF family proteins | ||
| VEGF-A165 | + | + |
| VEGF-A121 | + | |
| VEGF-B167 | + | |
| VEGF-C | + | + |
| VEGF-D | + | + |
| PlGF-2 | + | + |
| VEGFR (R1/R2/R3) | + | + |
| Semaphorin family proteins | ||
| SEMA3A | + | |
| SEMA3B, C, D, F | + | + |
| SEMA3G | + | |
| Plexin-A1 to A4, D1 | + | + |
| Other growth factors | ||
| TGF-β1 and LAP | + | + |
| TGF-β receptors (TbRI, TbRII) | + | + |
| FGF-1, 2, 4, 7 | + | + |
| FGFR-1 | + | |
| Other molecules | ||
| Heparin | + | |
| HGF and c-MET | + | + |
| PDGF and PDGFR | + | |
| Integrins (α5β1, αvβ3) | + | + |
| Fibronectin | + |
| Neuropilin (NRP) | Cancer Type | Activity | Effect |
|---|---|---|---|
| NRP-1 | Gastric cancer | Activation of VEGF and TGF-β, regulation of EMT | Increased angiogenesis, invasiveness and metastasis |
| Colorectal cancer | Promotion of tumour cell migration via VEGF and PI3K/AKT signalling | Increased aggressiveness and resistance to treatment | |
| Pancreatic cancer | Involvement in the activation of tumour-associated fibroblasts (CAFs), regulation of the cancer microenvironment | Increased desmoplasia and cancer progression | |
| Liver cancer | Facilitates EMT by promoting the migration of cancer cells and enhancing their metastatic potential | Increased expression, associated with lower patient survival, higher risk of vascular invasion, and activation of HSC | |
| NRP-2 | Colorectal cancer | Regulation of VEGF-C/D signalling and lymphangiogenesis | Greater propensity for lymph node metastasis |
| Pancreatic cancer | Promotes immunosuppression through effects on Treg cells | Bypassing the immune response and cancer progression | |
| Gastric cancer | Supports PI3K/AKT and MAPK pathways | Increased proliferation and resistance to treatment |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Królikowska, K.; Kurman, N.; Błaszczak, K.; Ławicki, S.; Gudowska-Sawczuk, M.; Zajkowska, M. The Significance of Neuropilins in Gastrointestinal Cancers. Int. J. Mol. Sci. 2025, 26, 4937. https://doi.org/10.3390/ijms26104937
Królikowska K, Kurman N, Błaszczak K, Ławicki S, Gudowska-Sawczuk M, Zajkowska M. The Significance of Neuropilins in Gastrointestinal Cancers. International Journal of Molecular Sciences. 2025; 26(10):4937. https://doi.org/10.3390/ijms26104937
Chicago/Turabian StyleKrólikowska, Kinga, Natalia Kurman, Katarzyna Błaszczak, Sławomir Ławicki, Monika Gudowska-Sawczuk, and Monika Zajkowska. 2025. "The Significance of Neuropilins in Gastrointestinal Cancers" International Journal of Molecular Sciences 26, no. 10: 4937. https://doi.org/10.3390/ijms26104937
APA StyleKrólikowska, K., Kurman, N., Błaszczak, K., Ławicki, S., Gudowska-Sawczuk, M., & Zajkowska, M. (2025). The Significance of Neuropilins in Gastrointestinal Cancers. International Journal of Molecular Sciences, 26(10), 4937. https://doi.org/10.3390/ijms26104937