
Exploring the Bone–Liver Axis: Impact of Acute Ethanol Intoxication on Post-Traumatic Liver Inflammation and Damage Following Femur Fracture

 , , ,
, , ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
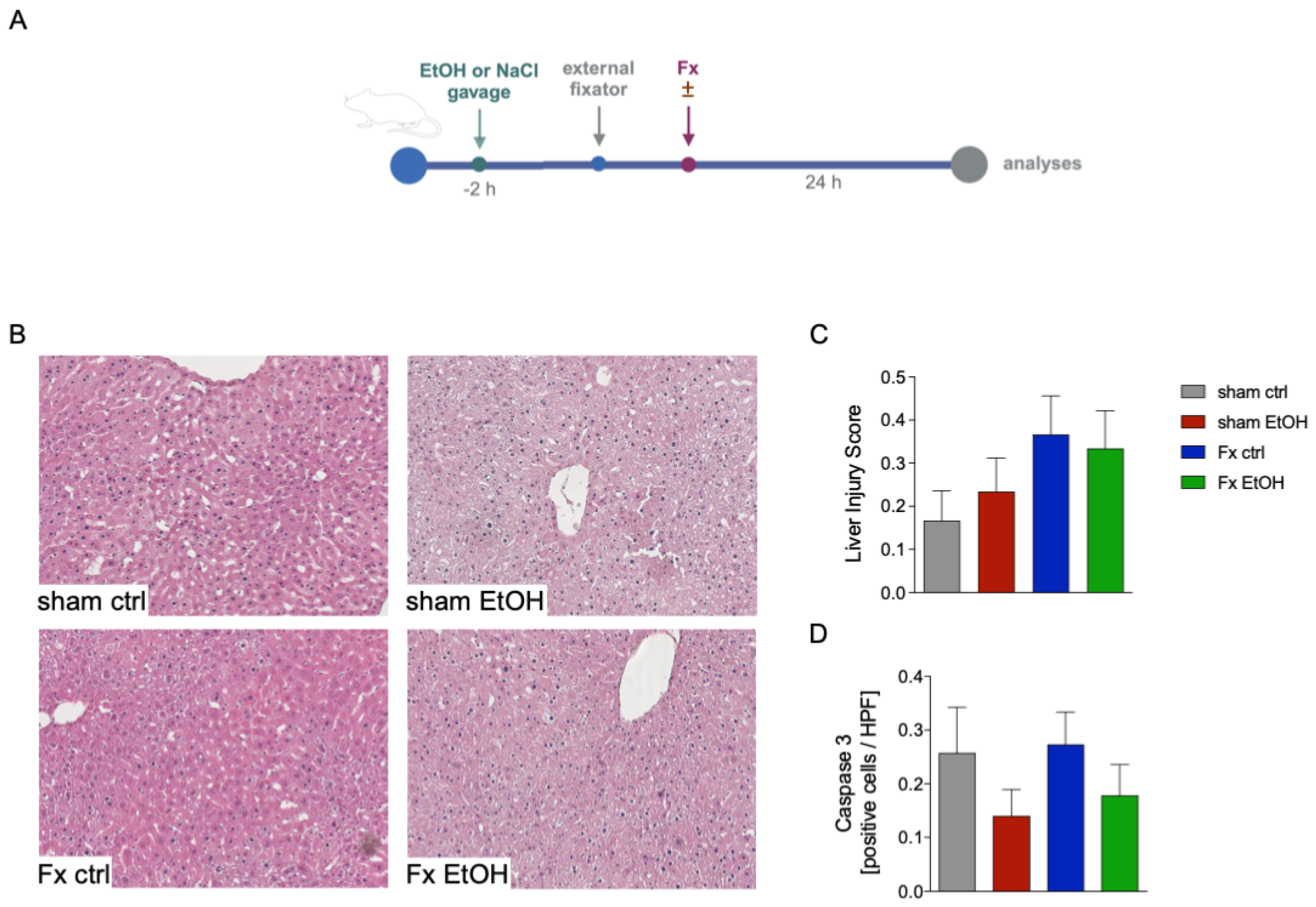
2.1. Fracture and AAI Does Not Lead to Liver Damage
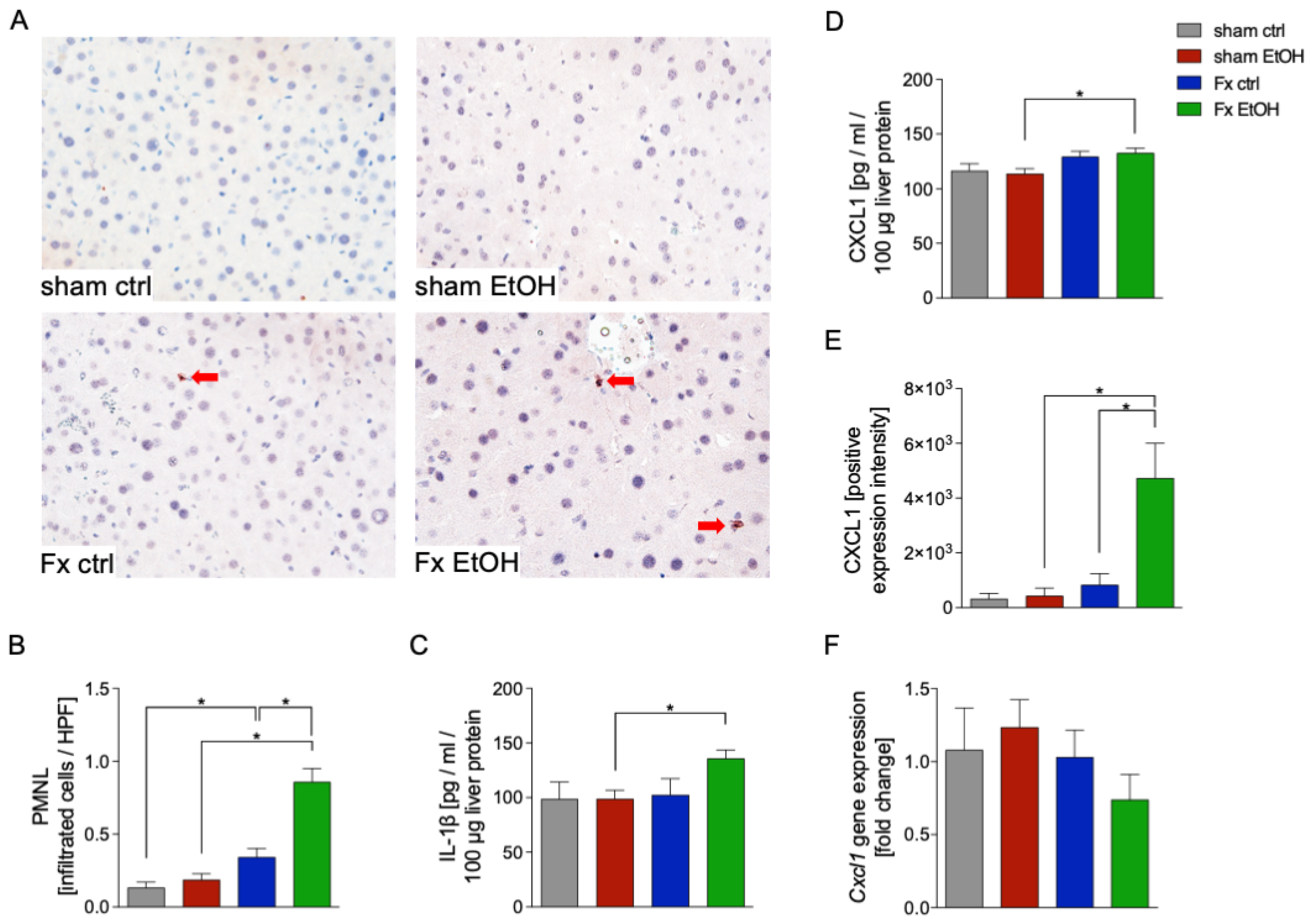
2.2. Increased Neutrophil Infiltration After Fracture and AAI
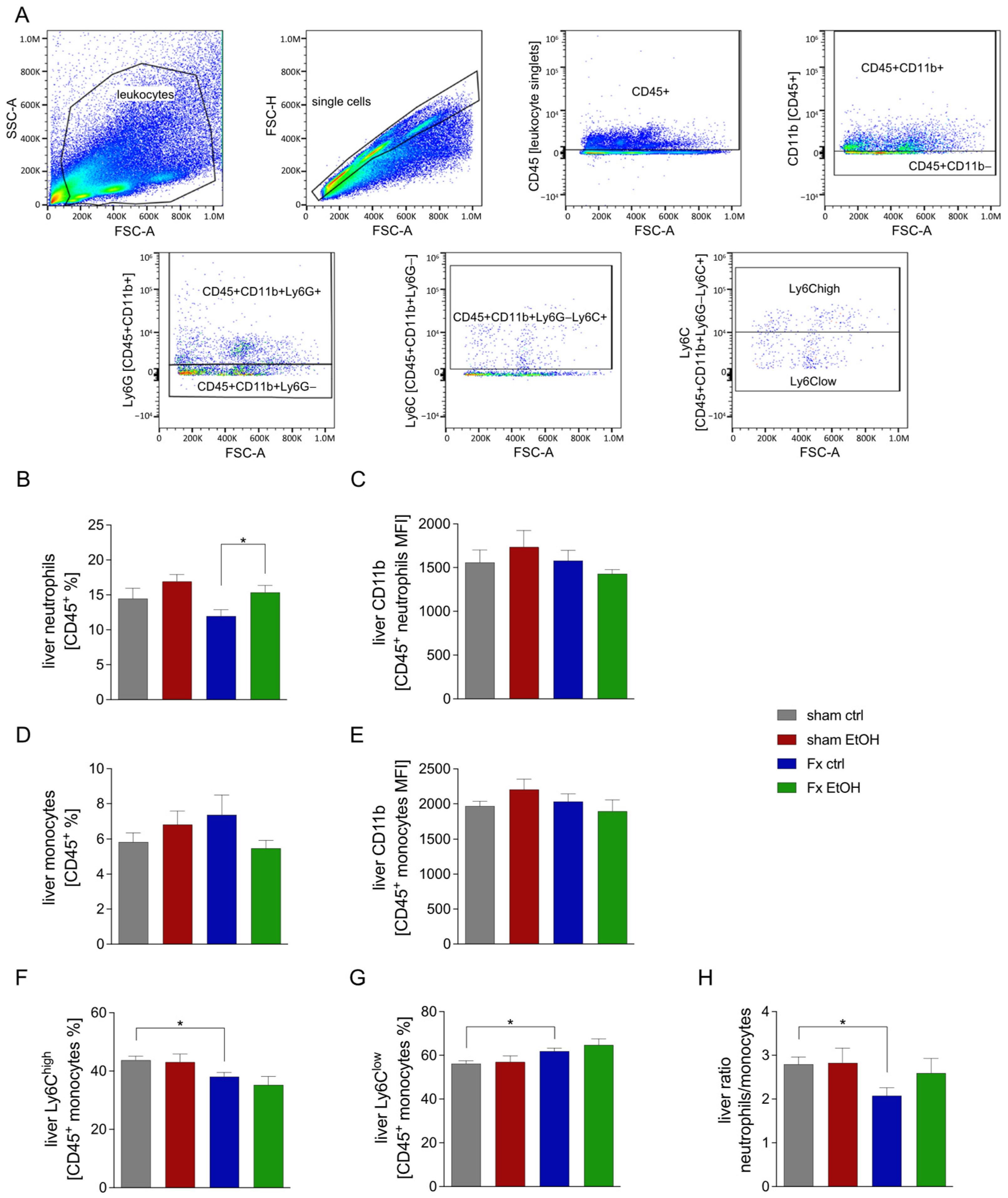
2.3. Fracture Influences Immune Cell Proportion in the Liver
2.4. Fracture and AAI Influence Liver Inflammation
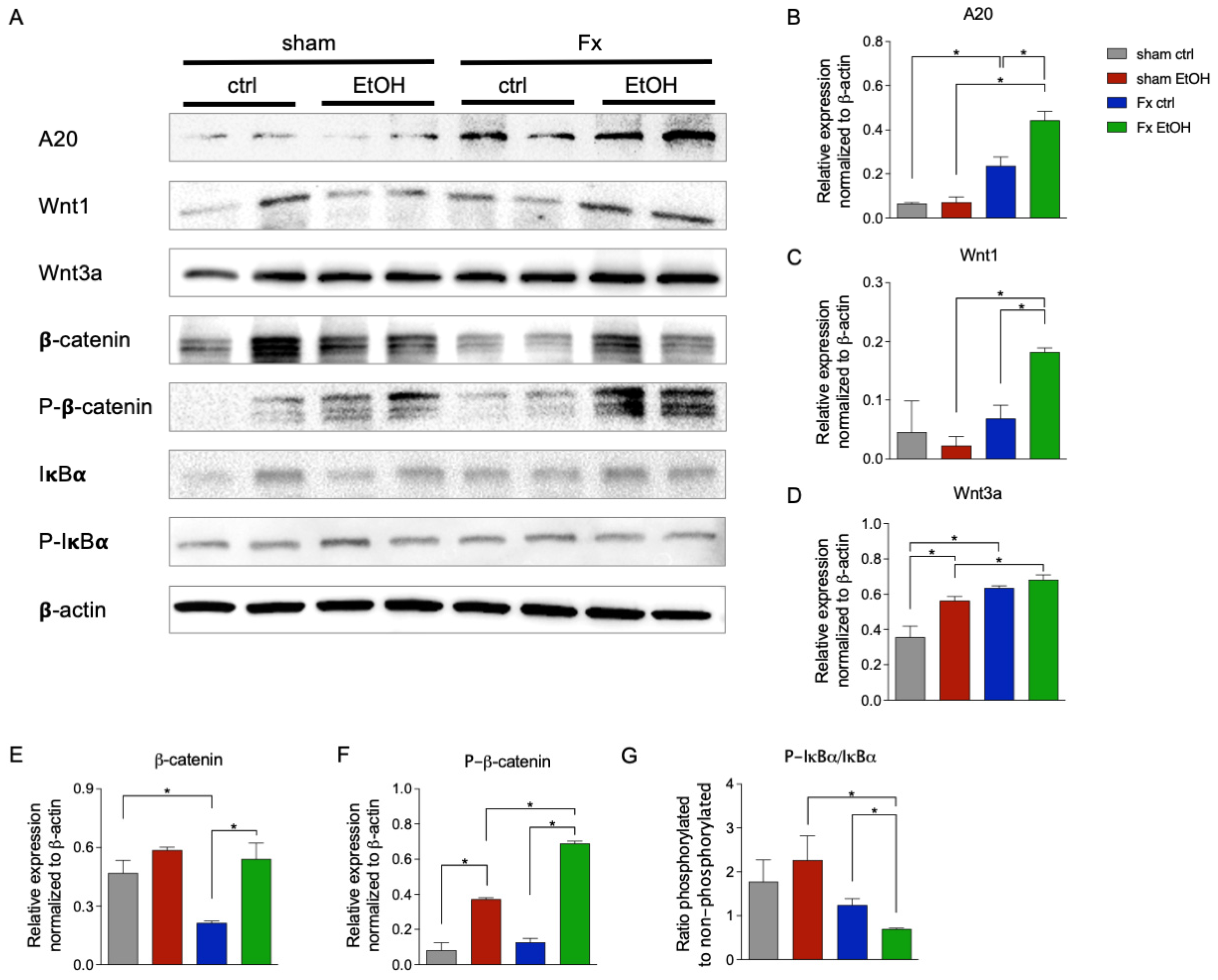
2.5. Fracture and AAI Modulate Wnt and NF-κB Signaling
3. Discussion
4. Materials and Methods
4.1. Animal Care, Husbandry, and Group Organization
4.2. Experimental Model
4.3. Harvesting Procedures
4.4. Examination of Liver Damage
4.5. Immune Histological Staining of CXCL1, Neutrophil Elastase, and Active Caspase-3
4.6. Gene Expression Analysis
4.7. Protein Expression Levels via ELISA
4.8. Flow Cytometry
4.9. Western Blot
4.10. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, A.M.; Bisignano, C.; James, S.L.; Abady, G.G.; Abedi, A.; Abu-Gharbieh, E.; Alhassan, R.K.; Alipour, V.; Arabloo, J.; Asaad, M.; et al. Global, regional, and national burden of bone fractures in 204 countries and territories, 1990–2019: A systematic analysis from the Global Burden of Disease Study 2019. Lancet Healthy Longev. 2021, 2, e580–e592. [Google Scholar] [CrossRef] [PubMed]
- WHO. Fragility Fractures; WHO: Geneva, Switzerland, 2024. [Google Scholar]
- Kolar, P.; Schmidt-Bleek, K.; Schell, H.; Gaber, T.; Toben, D.; Schmidmaier, G.; Perka, C.; Buttgereit, F.; Duda, G.N. The early fracture hematoma and its potential role in fracture healing. Tissue Eng. Part. B Rev. 2010, 16, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Claes, L.; Recknagel, S.; Ignatius, A. Fracture healing under healthy and inflammatory conditions. Nat. Rev. Rheumatol. 2012, 8, 133–143. [Google Scholar] [CrossRef]
- Walters, G.; Pountos, I.; Giannoudis, P.V. The cytokines and micro-environment of fracture haematoma: Current evidence. J. Tissue Eng. Regen. Med. 2018, 12, e1662–e1677. [Google Scholar] [CrossRef] [PubMed]
- Bahney, C.S.; Zondervan, R.L.; Allison, P.; Theologis, A.; Ashley, J.W.; Ahn, J.; Miclau, T.; Marcucio, R.S.; Hankenson, K.D. Cellular biology of fracture healing. J. Orthop. Res. 2019, 37, 35–50. [Google Scholar] [CrossRef]
- Gerstenfeld, L.C.; Thiede, M.; Seibert, K.; Mielke, C.; Phippard, D.; Svagr, B.; Cullinane, D.; Einhorn, T.A. Differential inhibition of fracture healing by non-selective and cyclooxygenase-2 selective non-steroidal anti-inflammatory drugs. J. Orthop. Res. 2003, 21, 670–675. [Google Scholar] [CrossRef]
- Ragipoglu, D.; Bülow, J.; Hauff, K.; Voss, M.; Haffner-Luntzer, M.; Dudeck, A.; Ignatius, A.; Fischer, V. Mast Cells Drive Systemic Inflammation and Compromised Bone Repair After Trauma. Front. Immunol. 2022, 13, 883707. [Google Scholar] [CrossRef]
- Li, H.; Liu, J.; Yao, J.; Zhong, J.; Guo, L.; Sun, T. Fracture initiates systemic inflammatory response syndrome through recruiting polymorphonuclear leucocytes. Immunol. Res. 2016, 64, 1053–1059. [Google Scholar] [CrossRef]
- Kobbe, P.; Vodovotz, Y.; Kaczorowski, D.J.; Billiar, T.R.; Pape, H.-C. The role of fracture-associated soft tissue injury in the induction of systemic inflammation and remote organ dysfunction after bilateral femur fracture. J. Orthop. Trauma. 2008, 22, 385–390. [Google Scholar] [CrossRef]
- Mollen, K.P.; Levy, R.M.; Prince, J.M.; A Hoffman, R.; Scott, M.J.; Kaczorowski, D.J.; Vallabhaneni, R.; Vodovotz, Y.; Billiar, T.R. Systemic inflammation and end organ damage following trauma involves functional TLR4 signaling in both bone marrow-derived cells and parenchymal cells. J. Leukoc. Biol. 2008, 83, 80–88. [Google Scholar] [CrossRef]
- Robinson, M.W.; Harmon, C.; O’farrelly, C. Liver immunology and its role in inflammation and homeostasis. Cell Mol. Immunol. 2016, 13, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Racanelli, V.; Rehermann, B. The liver as an immunological organ. Hepatology 2006, 43 (Suppl. 1), S54–S62. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, E.; Baird, A.W.; O’Farrelly, C. Microanatomy of the liver immune system. Semin. Immunopathol. 2009, 31, 333–343. [Google Scholar] [CrossRef]
- Luedde, T.; Schwabe, R.F. NF-kappaB in the liver-linking injury, fibrosis and hepatocellular carcinoma. Nat. Rev. Gastroenterol. Hepatol. 2011, 8, 108–118. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-kappaB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef]
- Behari, J. The Wnt/beta-catenin signaling pathway in liver biology and disease. Expert. Rev. Gastroenterol. Hepatol. 2010, 4, 745–756. [Google Scholar] [CrossRef]
- Perugorria, M.J.; Olaizola, P.; Labiano, I.; Esparza-Baquer, A.; Marzioni, M.; Marin, J.J.; Bujanda, L.; Banales, J.M. Wnt-beta-catenin signalling in liver development, health and disease. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Shembade, N.; Harhaj, E.W. Regulation of NF-kappaB signaling by the A20 deubiquitinase. Cell Mol. Immunol. 2012, 9, 123–130. [Google Scholar] [CrossRef]
- Nakamura, B.N.; Glazier, A.; Kattah, M.G.; Duong, B.; Jia, Y.; Campo, D.; Shao, L. A20 regulates canonical wnt-signaling through an interaction with RIPK4. PLoS ONE 2018, 13, e0195893. [Google Scholar] [CrossRef]
- WHO. Global Statuse Report on Alcohol and Health 2018; Poznyak, V., Rekve, D., Eds.; WHO: Geneva, Switzerland, 2018; p. 450. [Google Scholar]
- DHS. DHS Jahrbuch Sucht 2024; DHS: Hamm, Germany, 2024. [Google Scholar]
- Ke, Y.; Hu, H.; Zhang, J.; Yuan, L.; Li, T.; Feng, Y.; Wu, Y.; Fu, X.; Wang, M.; Gao, Y.; et al. Alcohol Consumption and Risk of Fractures: A Systematic Review and Dose-Response Meta-Analysis of Prospective Cohort Studies. Adv. Nutr. 2023, 14, 599–611. [Google Scholar] [CrossRef]
- Wagner, N.; Relja, B.; Lustenberger, T.; Leiblein, M.; Wutzler, S.; Lefering, R.; Marzi, I.; TraumaRegister DGU3. The influence of alcohol on the outcome of trauma patients: A matched-pair analysis of the TraumaRegister DGU®. Eur. J. Trauma. Emerg. Surg. 2020, 46, 463–472. [Google Scholar] [CrossRef]
- Relja, B.; Menke, J.; Wagner, N.; Auner, B.; Voth, M.; Nau, C.; Marzi, I. Effects of positive blood alcohol concentration on outcome and systemic interleukin-6 in major trauma patients. Injury 2016, 47, 640–645. [Google Scholar] [CrossRef] [PubMed]
- Eguchi, A.; Franz, N.; Kobayashi, Y.; Iwasa, M.; Wagner, N.; Hildebrand, F.; Takei, Y.; Marzi, I.; Relja, B. Circulating Extracellular Vesicles and Their miR “Barcode” Differentiate Alcohol Drinkers with Liver Injury and Those Without Liver Injury in Severe Trauma Patients. Front. Med. 2019, 6, 30. [Google Scholar] [CrossRef]
- Wagner, N.; Dieteren, S.; Franz, N.; Köhler, K.; Perl, M.; Marzi, I.; Relja, B. Alcohol-induced attenuation of post-traumatic inflammation is not necessarily liver-protective following trauma/hemorrhage. Int. J. Mol. Med. 2019, 44, 1127–1138. [Google Scholar] [CrossRef] [PubMed]
- Relja, B.; Höhn, C.; Bormann, F.; Seyboth, K.; Henrich, D.; Marzi, I.; Lehnert, M. Acute alcohol intoxication reduces mortality, inflammatory responses and hepatic injury after haemorrhage and resuscitation in vivo. Br. J. Pharmacol. 2012, 165, 1188–1199. [Google Scholar] [CrossRef]
- Franz, N.; Dieteren, S.; Köhler, K.; Mörs, K.; Sturm, R.; Marzi, I.; Perl, M.; Relja, B.; Wagner, N. Alcohol Binge Reduces Systemic Leukocyte Activation and Pulmonary PMN Infiltration After Blunt Chest Trauma and Hemorrhagic Shock. Inflammation 2019, 42, 690–701. [Google Scholar] [CrossRef] [PubMed]
- Jridi, I.; Canté-Barrett, K.; Pike-Overzet, K.; Staal, F.J.T. Inflammation and Wnt Signaling: Target for Immunomodulatory Therapy? Front. Cell Dev. Biol. 2020, 8, 615131. [Google Scholar] [CrossRef]
- Li, Z.; Wen, X.; Li, N.; Zhong, C.; Chen, L.; Zhang, F.; Zhang, G.; Lyu, A.; Liu, J. The roles of hepatokine and osteokine in liver-bone crosstalk: Advance in basic and clinical aspects. Front. Endocrinol. 2023, 14, 1149233. [Google Scholar] [CrossRef]
- Lupu, F.; Terwilliger, J.D.; Lee, K.; Segre, G.V.; Efstratiadis, A. Roles of growth hormone and insulin-like growth factor 1 in mouse postnatal growth. Dev. Biol. 2001, 229, 141–162. [Google Scholar] [CrossRef]
- Kurland, E.S.; Chan, F.K.; Rosen, C.J.; Bilezikian, J.P. Normal growth hormone secretory reserve in men with idiopathic osteoporosis and reduced circulating levels of insulin-like growth factor-I. J. Clin. Endocrinol. Metab. 1998, 83, 2576–2579. [Google Scholar]
- Liang, S.; Cheng, X.; Hu, Y.; Song, R.; Li, G. Insulin-like growth factor 1 and metabolic parameters are associated with nonalcoholic fatty liver disease in obese children and adolescents. Acta Paediatr. 2017, 106, 298–303. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Lei, H.; Wang, T.; Xiong, X. Liver-bone crosstalk in non-alcoholic fatty liver disease: Clinical implications and underlying pathophysiology. Front. Endocrinol. 2023, 14, 1161402. [Google Scholar] [CrossRef]
- Wei, W.; Dutchak, P.A.; Wang, X.; Ding, X.; Wang, X.; Bookout, A.L.; Goetz, R.; Mohammadi, M.; Gerard, R.D.; Dechow, P.C.; et al. Fibroblast growth factor 21 promotes bone loss by potentiating the effects of peroxisome proliferator-activated receptor gamma. Proc. Natl. Acad. Sci. USA 2012, 109, 3143–3148. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wei, W.; Krzeszinski, J.Y.; Wang, Y.; Wan, Y. A Liver-Bone Endocrine Relay by IGFBP1 Promotes Osteoclastogenesis and Mediates FGF21-Induced Bone Resorption. Cell Metab. 2015, 22, 811–824. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Grabner, A.; Yanucil, C.; Schramm, K.; Czaya, B.; Krick, S.; Czaja, M.J.; Bartz, R.; Abraham, R.; Di Marco, G.S.; et al. Fibroblast growth factor 23 directly targets hepatocytes to promote inflammation in chronic kidney disease. Kidney Int. 2016, 90, 985–996. [Google Scholar] [CrossRef]
- Kumar, P.; Liu, Y.; Shen, Y.; Maher, J.J.; Cingolani, F.; Czaja, M.J. Mouse liver injury induces hepatic macrophage FGF23 production. PLoS ONE 2022, 17, e0264743. [Google Scholar] [CrossRef]
- Yoshida, K.; Murata, M.; Yamaguchi, T.; Zaki, K.M. TGF-beta/Smad signaling during hepatic fibro-carcinogenesis (review). Int. J. Oncol. 2014, 45, 1363–1371. [Google Scholar] [CrossRef]
- Störmann, P.; Wagner, N.; Köhler, K.; Auner, B.; Simon, T.-P.; Pfeifer, R.; Horst, K.; Pape, H.-C.; Hildebrand, F.; Wutzler, S.; et al. Monotrauma is associated with enhanced remote inflammatory response and organ damage, while polytrauma intensifies both in porcine trauma model. Eur. J. Trauma. Emerg. Surg. 2020, 46, 31–42. [Google Scholar] [CrossRef]
- Meng, F.; Zhou, Y.; Wagner, A.; Bülow, J.M.; Köhler, K.; Neunaber, C.; Bundkirchen, K.; Relja, B. Impact of age on liver damage, inflammation, and molecular signaling pathways in response to femoral fracture and hemorrhage. Front. Immunol. 2023, 14, 1239145. [Google Scholar] [CrossRef]
- Schneider, C.P.; Schwacha, M.G.; Chaudry, I.H. The role of interleukin-10 in the regulation of the systemic inflammatory response following trauma-hemorrhage. Biochim. Biophys. Acta 2004, 1689, 22–32. [Google Scholar] [CrossRef]
- Korbecki, J.; Gąssowska-Dobrowolska, M.; Wójcik, J.; Szatkowska, I.; Barczak, K.; Chlubek, M.; Baranowska-Bosiacka, I. The Importance of CXCL1 in Physiology and Noncancerous Diseases of Bone, Bone Marrow, Muscle and the Nervous System. Int. J. Mol. Sci. 2022, 23, 4205. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, Q.; Liu, T.; Zheng, Q.; Xu, X.E.; Liu, X.; Gao, W.; Li, Z.; Bai, X. Early plasma monocyte chemoattractant protein 1 predicts the development of sepsis in trauma patients: A prospective observational study. Medicine 2018, 97, e0356. [Google Scholar] [CrossRef]
- Sawant, K.V.; Poluri, K.M.; Dutta, A.K.; Sepuru, K.M.; Troshkina, A.; Garofalo, R.P.; Rajarathnam, K. Chemokine CXCL1 mediated neutrophil recruitment: Role of glycosaminoglycan interactions. Sci. Rep. 2016, 6, 33123. [Google Scholar] [CrossRef] [PubMed]
- Roth, K.; Strickland, J.; Pant, A.; Freeborn, R.; Kennedy, R.; Rockwell, C.E.; Luyendyk, J.P.; Copple, B.L. Interleukin-10 disrupts liver repair in acetaminophen-induced acute liver failure. Front. Immunol. 2023, 14, 1303921. [Google Scholar] [CrossRef]
- Mandrekar, P.; Ambade, A.; Lim, A.; Szabo, G.; Catalano, D. An essential role for monocyte chemoattractant protein-1 in alcoholic liver injury: Regulation of proinflammatory cytokines and hepatic steatosis in mice. Hepatology 2011, 54, 2185–2197. [Google Scholar] [CrossRef] [PubMed]
- Deshmane, S.L.; Kremlev, S.; Amini, S.; Sawaya, B.E. Monocyte chemoattractant protein-1 (MCP-1): An overview. J. Interferon Cytokine Res. 2009, 29, 313–326. [Google Scholar] [CrossRef]
- Spiller, K.L.; Koh, T.J. Macrophage-based therapeutic strategies in regenerative medicine. Adv. Drug Deliv. Rev. 2017, 122, 74–83. [Google Scholar] [CrossRef]
- Lee, E.G.; Boone, D.L.; Chai, S.; Libby, S.L.; Chien, M.; Lodolce, J.P.; Ma, A. Failure to regulate TNF-induced NF-kappaB and cell death responses in A20-deficient mice. Science 2000, 289, 2350–2354. [Google Scholar] [CrossRef]
- Yang, W.; Zheng, Y.; He, H.; Zhang, H.; Wan, L.; Zhang, Q.; Yu, Z. Inhibition of trauma-associated inflammatory liver damage by blocking NF-kappaB activity. Hepatogastroenterology 2012, 59, 1174–1178. [Google Scholar]
- Sawant, K.V.; Sepuru, K.M.; Lowry, E.; Penaranda, B.; Frevert, C.W.; Garofalo, R.P.; Rajarathnam, K. Neutrophil recruitment by chemokines Cxcl1/KC and Cxcl2/MIP2: Role of Cxcr2 activation and glycosaminoglycan interactions. J. Leukoc. Biol. 2021, 109, 777–791. [Google Scholar] [CrossRef]
- Liu, J.; Xiao, Q.; Xiao, J.; Niu, C.; Li, Y.; Zhang, X.; Zhou, Z.; Shu, G.; Yin, G. Wnt/beta-catenin signalling: Function, biological mechanisms, and therapeutic opportunities. Signal Transduct. Target. Ther. 2022, 7, 3. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Han, W.; Park, T.; Kim, E.J.; Bang, I.; Lee, H.S.; Jeong, Y.; Roh, K.; Kim, J.; Kim, J.S.; et al. Sclerostin inhibits Wnt signaling through tandem interaction with two LRP6 ectodomains. Nat. Commun. 2020, 11, 5357. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.O.; Monga, S.P. Wnt/beta-Catenin Signaling in Liver Development, Homeostasis, and Pathobiology. Annu. Rev. Pathol. 2018, 13, 351–378. [Google Scholar] [CrossRef]
- Liu, K.; Wang, F.S.; Xu, R. Neutrophils in liver diseases: Pathogenesis and therapeutic targets. Cell Mol. Immunol. 2021, 18, 38–44. [Google Scholar] [CrossRef]
- Cai, J.; Zhang, X.J.; Li, H. The Role of Innate Immune Cells in Nonalcoholic Steatohepatitis. Hepatology 2019, 70, 1026–1037. [Google Scholar] [CrossRef]
- Sturm, R.; Haag, F.; Bergmann, C.B.; Marzi, I.; Relja, B. Alcohol drinking leads to sex-dependent differentiation of T cells. Eur. J. Trauma. Emerg. Surg. 2025, 51, 87. [Google Scholar] [CrossRef] [PubMed]
- Mörs, K.; Braun, O.; Wagner, N.; Auner, B.; Voth, M.; Störmann, P.; Wutzler, S.; Marzi, I.; Relja, B. Influence of gender on systemic IL-6 levels, complication rates and outcome after major trauma. Immunobiology 2016, 221, 904–910. [Google Scholar] [CrossRef]
- Bundkirchen, K.; Ye, W.; Nowak, A.J.; Lienenklaus, S.; Welke, B.; Relja, B.; Neunaber, C. Fracture Healing in Elderly Mice and the Effect of an Additional Severe Blood Loss: A Radiographic and Biomechanical Murine Study. Bioengineering 2023, 10, 70. [Google Scholar] [CrossRef]
- Noack, L.; Bundkirchen, K.; Xu, B.; Gylstorff, S.; Zhou, Y.; Köhler, K.; Jantaree, P.; Neunaber, C.; Nowak, A.J.; Relja, B. Acute Intoxication with Alcohol Reduces Trauma-Induced Proinflammatory Response and Barrier Breakdown in the Lung via the Wnt/beta-Catenin Signaling Pathway. Front. Immunol. 2022, 13, 866925. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Chandrasekar, A.; Heuvel, F.O.; Powerski, M.; Nowak, A.; Noack, L.; Omari, J.; Huber-Lang, M.; Roselli, F.; Relja, B. Ethanol Intoxication Alleviates the Inflammatory Response of Remote Organs to Experimental Traumatic Brain Injury. Int. J. Mol. Sci. 2020, 21, 8181. [Google Scholar] [CrossRef]
- Zhang, T.; Neunaber, C.; Ye, W.; Wagner, A.; Bülow, J.M.; Relja, B.; Bundkirchen, K. Aging Influences Fracture Healing on the Cellular Level and Alters Systemic RANKL and OPG Concentrations in a Murine Model. Adv. Biol. 2024, 8, e2300653. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bülow, J.M.; Rinderknecht, H.; Becker, N.; Köhler, K.; Wagner, A.; Yang, Y.; Bundkirchen, K.; Neunaber, C.; Relja, B. Exploring the Bone–Liver Axis: Impact of Acute Ethanol Intoxication on Post-Traumatic Liver Inflammation and Damage Following Femur Fracture. Int. J. Mol. Sci. 2025, 26, 4923. https://doi.org/10.3390/ijms26104923
Bülow JM, Rinderknecht H, Becker N, Köhler K, Wagner A, Yang Y, Bundkirchen K, Neunaber C, Relja B. Exploring the Bone–Liver Axis: Impact of Acute Ethanol Intoxication on Post-Traumatic Liver Inflammation and Damage Following Femur Fracture. International Journal of Molecular Sciences. 2025; 26(10):4923. https://doi.org/10.3390/ijms26104923
Chicago/Turabian StyleBülow, Jasmin Maria, Helen Rinderknecht, Nils Becker, Kernt Köhler, Alessa Wagner, Yuntao Yang, Katrin Bundkirchen, Claudia Neunaber, and Borna Relja. 2025. "Exploring the Bone–Liver Axis: Impact of Acute Ethanol Intoxication on Post-Traumatic Liver Inflammation and Damage Following Femur Fracture" International Journal of Molecular Sciences 26, no. 10: 4923. https://doi.org/10.3390/ijms26104923
APA StyleBülow, J. M., Rinderknecht, H., Becker, N., Köhler, K., Wagner, A., Yang, Y., Bundkirchen, K., Neunaber, C., & Relja, B. (2025). Exploring the Bone–Liver Axis: Impact of Acute Ethanol Intoxication on Post-Traumatic Liver Inflammation and Damage Following Femur Fracture. International Journal of Molecular Sciences, 26(10), 4923. https://doi.org/10.3390/ijms26104923