
Extracellular Vesicle-Based Drug Delivery Systems in Cancer Therapy


Abstract
1. Introduction
2. Extracellular Vesicles
2.1. Biological Characteristics of Extracellular Vesicles
2.2. EV Isolation Methodologies
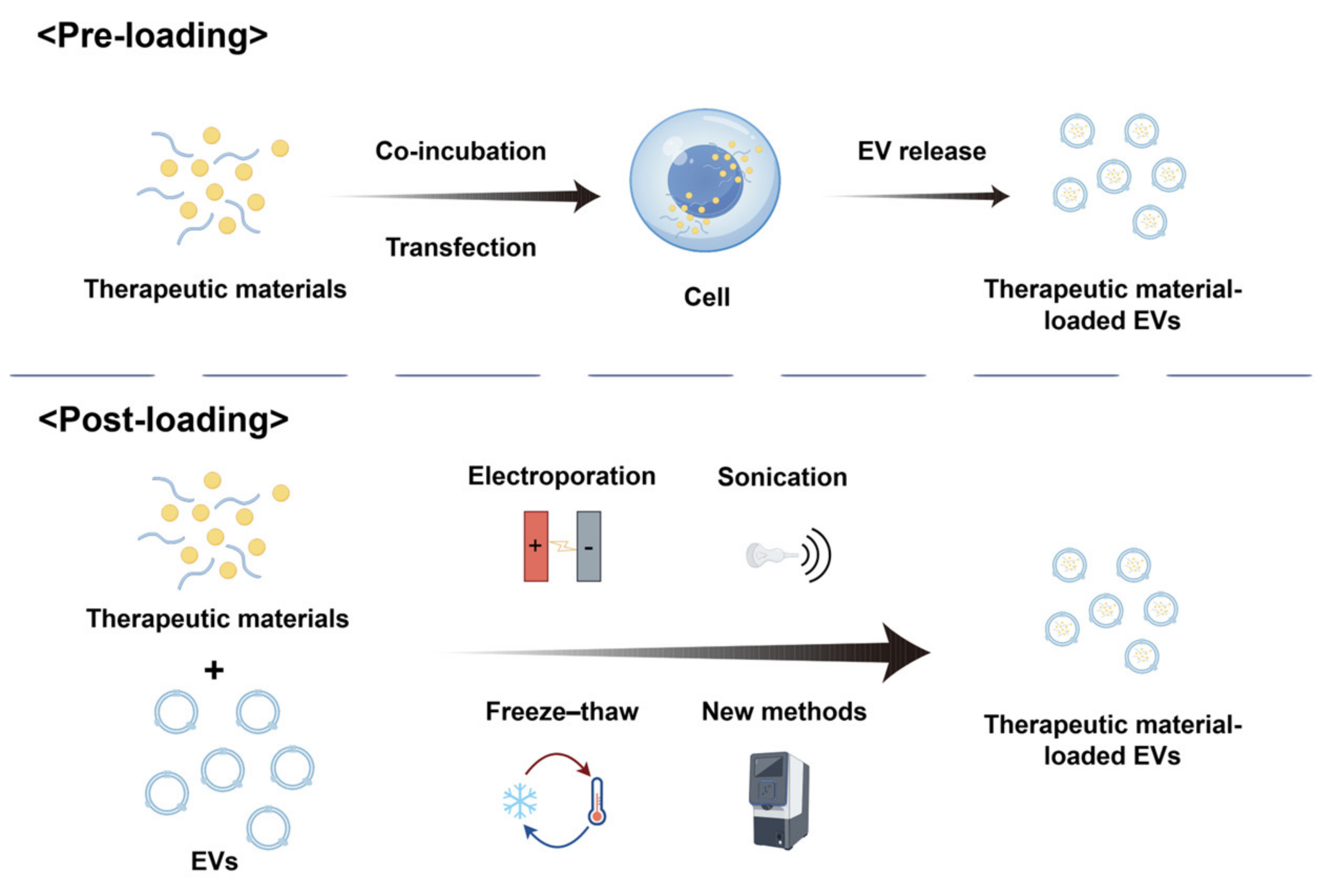
2.3. Drug-Loading Strategies
2.3.1. Pre-Loading
2.3.2. Post-Loading
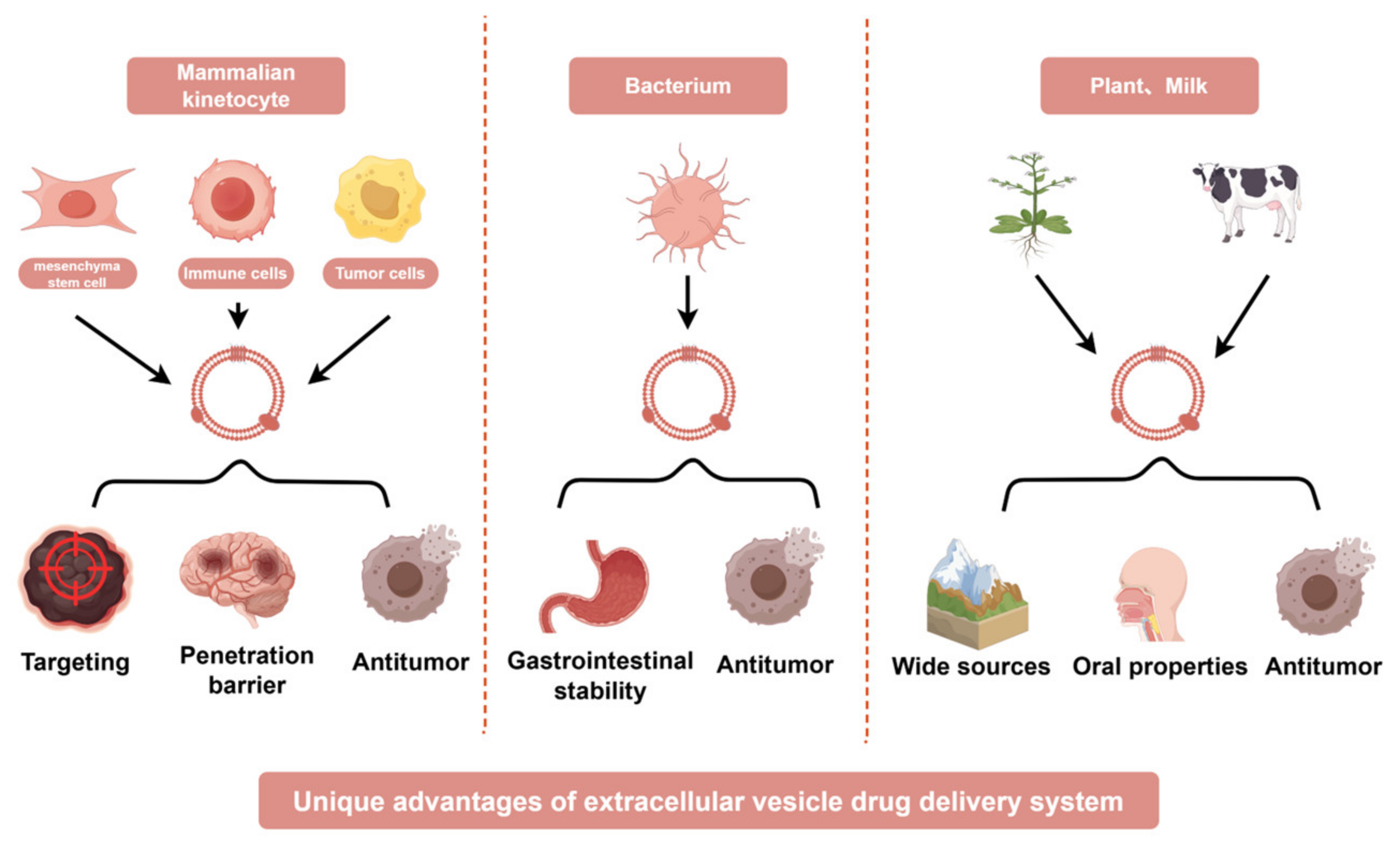
3. Advantages of Extracellular Vesicle-Based Drug Delivery Systems from Diverse Cellular Origins in Cancer Therapy
3.1. Mammalian Cell-Derived EVs
3.1.1. Mesenchymal Stem Cells (MSCs)
3.1.2. Immune Cells
3.1.3. Tumor Cells
3.2. Bacteria-Derived Extracellular Vesicles
3.3. Plant-Derived Extracellular Vesicles
3.4. Milk-Derived Extracellular Vesicles
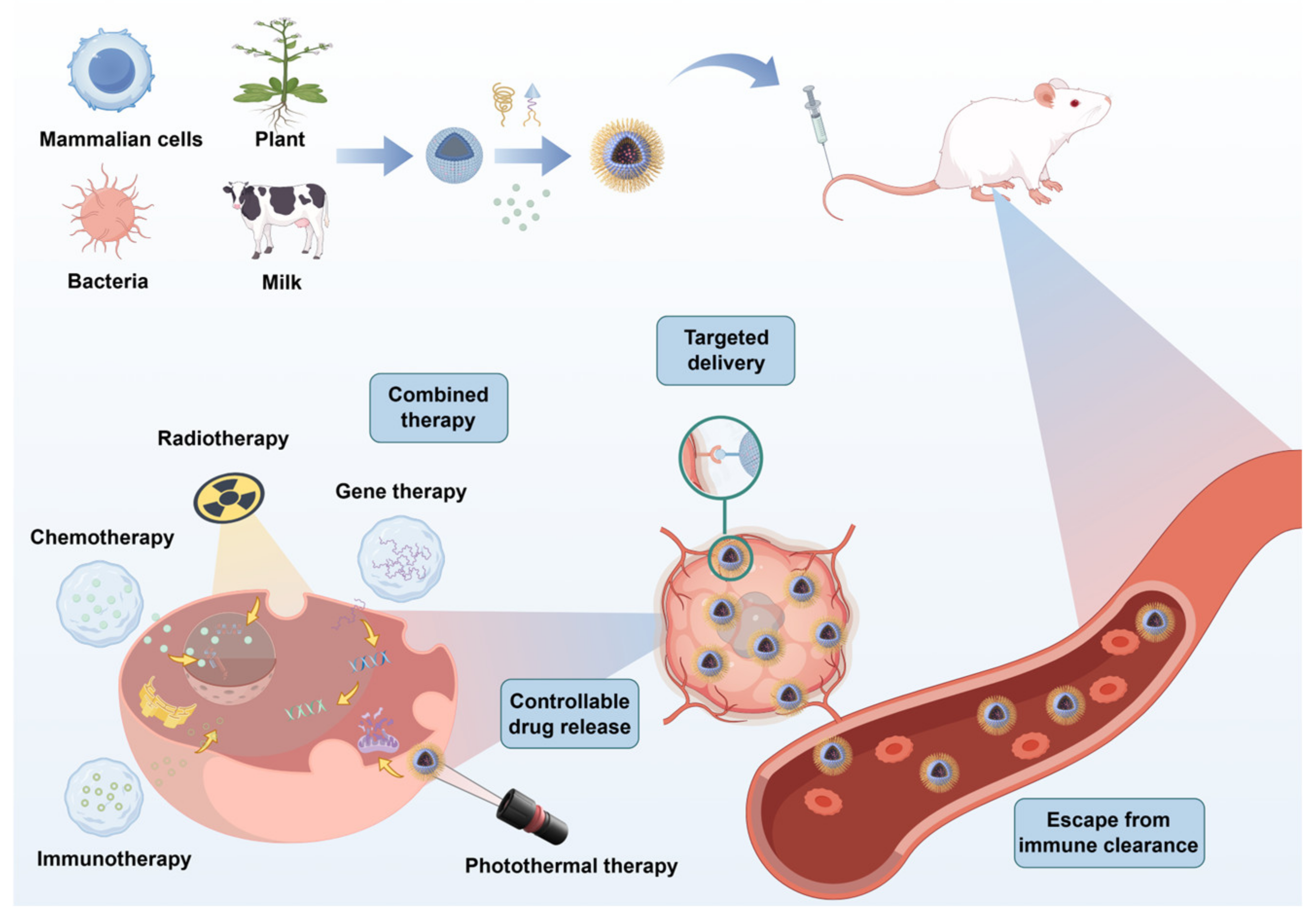
4. Application of Extracellular Vesicle-Based Drug Delivery Systems in Cancer Therapy
4.1. Chemotherapy Applications
4.1.1. Anthracyclines
4.1.2. Plant Alkaloids
4.1.3. Alkylating Agents
4.1.4. Antimetabolites
4.2. Application in Radiotherapy
4.3. Application in Gene Therapy
4.3.1. siRNA Delivery
4.3.2. mRNA Delivery
4.3.3. CRISPR/Cas9
4.4. Application in Immunotherapy
4.5. Application in Photothermal Therapy
4.6. Application in Cancer Vaccines
5. Clinical Advancements in Extracellular Vesicle-Based Drug Delivery Systems
5.1. Completed Clinical Trials
5.2. Ongoing Clinical Trials
5.3. Current Limitations in Clinical Research Progress
6. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| A4F | Asymmetrical flow field-flow fractionation |
| ABCA1 | ATP-binding cassette subfamily A member 1 |
| ACN | Anthocyanins |
| Aex | Ascites-derived exosomes |
| AI | Artificial intelligence |
| AI-TEVs | Attenuated immunogenic TEVs |
| Ang | Angiogenin |
| ARE | Antioxidant response element |
| BBB | Blood–brain barrier |
| BOF | Boronate organic framework |
| BSO | Buthionine sulfoximine |
| caCD8-EVs | Cytokine-activated CD8+ T cell-derived EVs |
| CAFs | Cancer-associated fibroblasts |
| CAR | Chimeric antigen receptor |
| CBP | Carboplatin |
| CD47 | Cluster of differentiation 47 |
| CDEVs | Celery-derived EVs |
| CDKI | Cyclin-dependent kinase inhibitor |
| CLENs | Citron-derived extracellular nanovesicles |
| CNVs | Cell-derived nanovesicles |
| COC | Cyclic olefin copolymer |
| conNK-sEVs | Conventional NK-EVs |
| CPPs | Cell-penetrating peptides |
| CPT | Camptothecin |
| CRC | Colorectal cancer |
| DC | Dendritic cell |
| DEC-205 | Dendritic cell-specific intercellular adhesion molecule-3-grabbing nonintegrin 205 |
| Dex | Dendritic cell-derived exosomes |
| DLTs | Dose-limiting toxicities |
| DOX | Doxorubicin |
| dsDNA | Double-stranded DNA |
| EAL | Esterase-responsive active loading |
| EGFR | Epidermal growth factor receptor |
| EPM | EV-based platform |
| EV-Rapa | Rapamycin-loaded EVs |
| EVs | Extracellular vesicles |
| Fc-EVs | Fc-engineered EVs |
| FeOOH | Iron oxyhydroxide |
| GEM | Gemcitabine |
| G-EVLP | Genetically engineered extracellular vesicle-like particles |
| GEVs | Ginger-derived EVs |
| GM-CSF | Granulocyte-macrophage colony-stimulating factor |
| GNPs | Garlic-derived nanoparticles |
| GQD | Graphene quantum dot |
| GSDMD | Gasdermin D-N-terminal |
| hPMSC-EVs | Human placental MSC-derived EVs |
| Hsp70s | 70 kDa heat-shock proteins |
| hUC-MSCs | Mesenchymal stem cells |
| ICG | Indocyanine green |
| iEDDA | Inverse electron-demand Diels–Alder |
| LEVs | Lactobacillus plantarum-derived EVs |
| LIPUS | Low-intensity pulsed ultrasound |
| LNs | Lymph nodes |
| LpEVs | Lactobacillus paracasei-derived EVs |
| MEVs | Milk-derived EVs |
| MnIO | Manganese-doped iron oxide nanoparticles |
| mNK-sEVs | Memory-like NK cell-derived small EVs |
| MPE | Malignant pleural effusion |
| MSA-2 | Macrophage-stimulating agent 2 |
| MSCs | Mesenchymal stem cells |
| MTD | Maximum tolerated dose |
| MVB | Multivesicular body |
| NSCLC | Non-small-cell lung cancer |
| OMVs | Outer membrane vesicles |
| OMVs | Outer membrane vesicles |
| OVA | Ovalbumin |
| PAMA | Poly(amidoamine) |
| PD-1 | Programmed cell death protein 1 |
| PDAC | Pancreatic ductal adenocarcinoma |
| PDENs | Plant-derived exosome-like nanoparticles |
| PDEs | Plant-derived extracellular vesicles |
| PD-L1 | Programmed death ligand 1 |
| PEG-GNS | Polyethylene glycol-coated gold nanoshells |
| PFC | Perfluorocarbon |
| PFS | Progression-free survival |
| pFUS | Pulsed focused ultrasound |
| PGEVs | Platycodon grandiflorus-derived EVs |
| PTX | Paclitaxel |
| RBCEVs | Red blood cell-derived EVs |
| RELNs | Rhodiola rosea-derived EV-like nanovesicles |
| REV | Regulator of virion expression |
| RGD | Arginine–glycine–aspartic acid |
| RS17 | Research-specific compound 17 |
| sBMEVs | Small bovine milk-derived small EVs |
| SDF-1 | Stromal cell-derived factor-1 |
| SEC | Size-exclusion chromatography |
| SE-FPLC | Size-exclusion fast protein liquid chromatography |
| siHSP47 | SiRNA targeting heat-shock protein 47 |
| STA | Staphylococcal enterotoxin A |
| STING | Stimulator of interferon genes |
| SUR-siRNA | Survivin-targeting siRNA |
| SWEET | Shock wave-mediated extracellular vesicle engineering technology |
| TAAs | Tumor-associated antigens |
| TAMs | Tumor-associated macrophages |
| TC | Tension controlled |
| TDDS | Tumor-targeted drug delivery systems |
| TNBC | Triple-negative breast cancer |
| TPC | Tetraphenylchlorin |
| UCMSCs | Umbilical cord MSCs |
| UDPG | UDP-glucose |
| uPAR | Urokinase-type plasminogen activator receptor |
| μSEC | Polymer-based microfluidic SEC |
References
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2024, 74, 229–263. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Xia, F.; Lin, R. Global Burden of Cancer and Associated Risk Factors in 204 Countries and Territories, 1980–2021: A Systematic Analysis for the GBD 2021. J. Hematol. Oncol. 2024, 17, 119. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zhou, H.; Tan, L.; Siu, K.T.H.; Guan, X.Y. Exploring treatment options in cancer: Tumor treatment strategies. Signal Transduct. Target. Ther. 2024, 9, 175. [Google Scholar] [CrossRef]
- Xia, Y.; Sun, M.; Huang, H.; Jin, W.L. Drug repurposing for cancer therapy. Signal Transduct. Target. Ther. 2024, 9, 92. [Google Scholar] [CrossRef]
- Nurgali, K.; Jagoe, R.T.; Abalo, R. Adverse effects of cancer chemotherapy: Anything new to improve tolerance and reduce sequelae? Front. Pharmacol. 2018, 9, 245. [Google Scholar] [CrossRef]
- Fang, R.H.; Gao, W.; Zhang, L. Targeting drugs to tumours using cell membrane-coated nanoparticles. Nat. Rev. Clin. Oncol. 2023, 20, 33–48. [Google Scholar] [CrossRef]
- Beach, M.A.; Nayanathara, U.; Gao, Y.; Zhang, C.; Xiong, Y.; Wang, Y.; Such, G.K. Polymeric nanoparticles for drug delivery. Chem. Rev. 2024, 124, 5505–5616. [Google Scholar] [CrossRef]
- Herrmann, I.K.; Wood, M.J.A.; Fuhrmann, G. Extracellular vesicles as a next-generation drug delivery platform. Nat. Nanotechnol. 2021, 16, 748–759. [Google Scholar] [CrossRef]
- Brezgin, S.; Danilik, O.; Yudaeva, A.; Kachanov, A.; Kostyusheva, A.; Karandashov, I.; Ponomareva, N.; Zamyatnin, A.A., Jr.; Parodi, A.; Chulanov, V.; et al. Basic Guide for Approaching Drug Delivery with Extracellular Vesicles. Int. J. Mol. Sci. 2024, 25, 10401. [Google Scholar] [CrossRef]
- Ghodasara, A.; Raza, A.; Wolfram, J.; Salomon, C.; Popat, A. Clinical translation of extracellular vesicles. Adv. Healthc. Mater. 2023, 12, e2301010. [Google Scholar] [CrossRef]
- Lorite, P.; Domínguez, J.N.; Palomeque, T.; Torres, M.I. Extracellular Vesicles: Advanced Tools for Disease Diagnosis, Monitoring, and Therapies. Int. J. Mol. Sci. 2024, 26, 189. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xing, K.; Zhang, G.; Li, Z.; Ding, X.; Leong, D.T. Surface Components and Biological Interactions of Extracellular Vesicles. ACS Nano 2025, 19, 8433–8461. [Google Scholar] [CrossRef]
- van Niel, G.; Carter, D.R.F.; Clayton, A.; Lambert, D.W.; Raposo, G.; Vader, P. Challenges and directions in studying cell-cell communication by extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2022, 23, 369–382. [Google Scholar] [CrossRef]
- Liu, X.; Cao, Y.; Wang, S.; Liu, J.; Hao, H. Extracellular vesicles: Powerful candidates in nano-drug delivery systems. Drug Deliv. Transl. Res. 2024, 14, 295–311. [Google Scholar] [CrossRef]
- Rehman, F.U.; Liu, Y.; Zheng, M.; Shi, B. Exosomes based strategies for brain drug delivery. Biomaterials 2023, 293, 121949. [Google Scholar] [CrossRef]
- Mohammadi, A.H.; Ghazvinian, Z.; Bagheri, F.; Harada, M.; Baghaei, K. Modification of extracellular vesicle surfaces: An approach for targeted drug delivery. BioDrugs 2023, 37, 353–374. [Google Scholar] [CrossRef]
- Yu, L.; Zhu, G.; Zhang, Z.; Yu, Y.; Zeng, L.; Xu, Z.; Weng, J.; Xia, J.; Li, J.; Pathak, J.L. Apoptotic bodies: Bioactive treasure left behind by the dying cells with robust diagnostic and therapeutic application potentials. J. Nanobiotechnol. 2023, 21, 218. [Google Scholar] [CrossRef]
- Jeppesen, D.K.; Fenix, A.M.; Franklin, J.L.; Higginbotham, J.N.; Zhang, Q.; Zimmerman, L.J.; Liebler, D.C.; Ping, J.; Liu, Q.; Evans, R.; et al. Reassessment of exosome composition. Cell 2019, 177, 428–445. [Google Scholar] [CrossRef]
- Zhou, M.; Li, Y.J.; Tang, Y.C.; Hao, X.Y.; Xu, W.J.; Xiang, D.X.; Wu, J.Y. Apoptotic bodies for advanced drug delivery and therapy. J. Control. Release 2022, 351, 394–406. [Google Scholar] [CrossRef]
- Dixson, A.C.; Dawson, T.R.; Di Vizio, D.; Weaver, A.M. Context-specific regulation of extracellular vesicle biogenesis and cargo selection. Nat. Rev. Mol. Cell Biol. 2023, 24, 454–476. [Google Scholar] [CrossRef]
- Yáñez-Mó, M.; Siljander, P.R.; Andreu, Z.; Zavec, A.B.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef]
- Clancy, J.W.; D’Souza-Schorey, C. Tumor-derived extracellular vesicles: Multifunctional entities in the tumor microenvironment. Annu. Rev. Pathol. 2023, 18, 205–229. [Google Scholar] [CrossRef] [PubMed]
- Kuang, L.; Wu, L.; Li, Y. Extracellular vesicles in tumor immunity: Mechanisms and novel insights. Mol. Cancer 2025, 24, 45. [Google Scholar] [CrossRef] [PubMed]
- Mortezaee, K.; Majidpoor, J. The impact of hypoxia on extracellular vesicle secretome profile of cancer. Med. Oncol. 2023, 40, 128. [Google Scholar] [CrossRef] [PubMed]
- Boussadia, Z.; Gambardella, A.R.; Mattei, F.; Parolini, I. Acidic and hypoxic microenvironment in melanoma: Impact of tumour exosomes on disease progression. Cells 2021, 10, 3311. [Google Scholar] [CrossRef]
- Wang, Z.; Zhou, X.; Kong, Q.; He, H.; Sun, J.; Qiu, W.; Zhang, L.; Yang, M. Extracellular vesicle preparation and analysis: A state-of-the-art review. Adv. Sci. 2024, 11, e2401069. [Google Scholar] [CrossRef]
- Zhu, L.; Sun, H.T.; Wang, S.; Huang, S.L.; Zheng, Y.; Wang, C.Q.; Hu, B.Y.; Qin, W.; Zou, T.T.; Fu, Y.; et al. Isolation and characterization of exosomes for cancer research. J. Hematol. Oncol. 2020, 13, 152. [Google Scholar] [CrossRef]
- Zhang, Q.; Jeppesen, D.K.; Higginbotham, J.N.; Franklin, J.L.; Coffey, R.J. Comprehensive Isolation of Extracellular Vesicles and Nanoparticles. Nat. Protoc. 2023, 18, 1462–1487. [Google Scholar] [CrossRef]
- Tiwari, S.; Kumar, V.; Randhawa, S.; Verma, S.K. Preparation and Characterization of Extracellular Vesicles. Am. J. Reprod. Immunol. 2021, 85, e13367. [Google Scholar] [CrossRef]
- Guo, J.; Wu, C.; Lin, X.; Zhou, J.; Zhang, J.; Zheng, W.; Wang, T.; Cui, Y. Establishment of a simplified dichotomic size-exclusion chromatography for isolating extracellular vesicles toward clinical applications. J. Extracell. Vesicles 2021, 10, e12145. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, Y.; Wei, S.; Zhou, C.; Yu, J.; Wang, G.; Wang, W.; Zhao, L. Extracellular vesicles isolated by size-exclusion chromatography present suitability for RNomics analysis in plasma. J. Transl. Med. 2021, 19, 104. [Google Scholar] [CrossRef]
- Kapoor, K.S.; Harris, K.; Arian, K.A.; Ma, L.; Schueng Zancanela, B.; Church, K.A.; McAndrews, K.M.; Kalluri, R. High throughput and rapid isolation of extracellular vesicles and exosomes with purity using size exclusion liquid chromatography. Bioact. Mater. 2024, 40, 683–695. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Wang, X.; Chen, Y.; Yuan, J.; Zhang, H.; Jin, X.; Jiang, Y.; Cao, J.; Wang, Z.; Yang, S.; et al. Distinct molecular properties and functions of small EV subpopulations isolated from human umbilical cord MSCs using tangential flow filtration combined with size exclusion chromatography. J. Extracell. Vesicles 2025, 14, e70029. [Google Scholar] [CrossRef] [PubMed]
- Brambilla, D.; Sola, L.; Ferretti, A.M.; Chiodi, E.; Zarovni, N.; Fortunato, D.; Criscuoli, M.; Dolo, V.; Giusti, I.; Murdica, V.; et al. EV separation: Release of intact extracellular vesicles immunocaptured on magnetic particles. Anal. Chem. 2021, 93, 5476–5483. [Google Scholar] [CrossRef]
- Di, K.; Fan, B.; Gu, X.; Huang, R.; Khan, A.; Liu, C.; Shen, H.; Li, Z. Highly efficient and automated isolation technology for extracellular vesicles microRNA. Front. Bioeng. Biotechnol. 2022, 10, 948757. [Google Scholar] [CrossRef]
- Wang, Z.; Dai, J.; He, H.; Si, T.; Ng, K.; Zheng, S.; Zhou, X.; Zhou, Z.; Yuan, H.; Yang, M. Cellulose nanofibrils of high immunoaffinity for efficient enrichment of small extracellular vesicles. Small Methods 2024, 8, e2400426. [Google Scholar] [CrossRef]
- Bajo-Santos, C.; Priedols, M.; Kaukis, P.; Paidere, G.; Gerulis-Bergmanis, R.; Mozolevskis, G.; Abols, A.; Rimsa, R. Extracellular vesicles isolation from large volume samples using a polydimethylsiloxane-free microfluidic device. Int. J. Mol. Sci. 2023, 24, 7971. [Google Scholar] [CrossRef]
- Leong, S.Y.; Ong, H.B.; Tay, H.M.; Kong, F.; Upadya, M.; Gong, L.; Dao, M.; Dalan, R.; Hou, H.W. Microfluidic size exclusion chromatography (μSEC) for extracellular vesicles and plasma protein separation. Small 2022, 18, e2104470. [Google Scholar] [CrossRef]
- Xin, F.; Ren, X.; Lin, X.; Ma, W.; Ran, B.; Teng, Y.; Gao, P.; Wang, C.; Wu, L.; Cun, D.; et al. Rapid isolation of extracellular vesicles using covalent organic frameworks combined with microfluidic technique. J. Pharm. Biomed. Anal. 2024, 245, 116153. [Google Scholar] [CrossRef]
- Lau, H.C.; Han, D.W.; Park, J.; Lehner, E.; Kals, C.; Arzt, C.; Bayer, E.; Auer, D.; Schally, T.; Grasmann, E.; et al. GMP-compliant manufacturing of biologically active cell-derived vesicles produced by extrusion technology. J. Extracell. Biol. 2022, 1, e70. [Google Scholar] [CrossRef]
- Li, X.; Zhong, Y.; Zhou, W.; Song, Y.; Li, W.; Jin, Q.; Gao, T.; Zhang, L.; Xie, M. Low-intensity pulsed ultrasound (LIPUS) enhances the anti-inflammatory effects of bone marrow mesenchymal stem cells (BMSCs)-derived extracellular vesicles. Cell. Mol. Biol. Lett. 2023, 28, 9. [Google Scholar] [CrossRef] [PubMed]
- Ullah, M.; Liu, D.D.; Rai, S.; Razavi, M.; Concepcion, W.; Thakor, A.S. Pulsed focused ultrasound enhances the therapeutic effect of mesenchymal stromal cell-derived extracellular vesicles in acute kidney injury. Stem Cell Res. Ther. 2020, 11, 398. [Google Scholar] [CrossRef] [PubMed]
- Zmievskaya, E.A.; Mukhametshin, S.A.; Ganeeva, I.A.; Gilyazova, E.M.; Siraeva, E.T.; Kutyreva, M.P.; Khannanov, A.A.; Yuan, Y.; Bulatov, E.R. Artificial extracellular vesicles generated from T cells using different induction techniques. Biomedicines 2024, 12, 919. [Google Scholar] [CrossRef] [PubMed]
- Boban, Z.; Mardešić, I.; Subczynski, W.K.; Raguz, M. Giant Unilamellar Vesicle Electroformation: What to Use, What to Avoid, and How to Quantify the Results. Membranes 2021, 11, 860. [Google Scholar] [CrossRef]
- Nair, K.S.; Bajaj, H. Advances in Giant Unilamellar Vesicle Preparation Techniques and Applications. Adv. Colloid Interface Sci. 2023, 318, 102935. [Google Scholar] [CrossRef]
- Boban, Z.; Mardešić, I.; Jozić, S.P.; Šumanovac, J.; Subczynski, W.K.; Raguz, M. Electroformation of Giant Unilamellar Vesicles from Damp Lipid Films Formed by Vesicle Fusion. Membranes 2023, 13, 352. [Google Scholar] [CrossRef]
- Waeterschoot, J.; Gosselé, W.; Alizadeh Zeinabad, H.; Lammertyn, J.; Koos, E.; Casadevall, I.; Solvas, X. Formation of Giant Unilamellar Vesicles Assisted by Fluorinated Nanoparticles. Adv. Sci. 2023, 10, e2302461. [Google Scholar] [CrossRef]
- Ernits, M.; Reinsalu, O.; Yandrapalli, N.; Kopanchuk, S.; Moradpur-Tari, E.; Sanka, I.; Scheler, O.; Rinken, A.; Kurg, R.; Kyritsakis, A.; et al. Microfluidic Production, Stability and Loading of Synthetic Giant Unilamellar Vesicles. Sci. Rep. 2024, 14, 14071. [Google Scholar] [CrossRef]
- Zeng, H.; Guo, S.; Ren, X.; Wu, Z.; Liu, S.; Yao, X. Current Strategies for Exosome Cargo Loading and Targeting Delivery. Cells 2023, 12, 1416. [Google Scholar] [CrossRef]
- Bahadorani, M.; Nasiri, M.; Dellinger, K.; Aravamudhan, S.; Zadegan, R. Engineering Exosomes for Therapeutic Applications: Decoding Biogenesis, Content Modification, and Cargo Loading Strategies. Int. J. Nanomed. 2024, 19, 7137–7164. [Google Scholar] [CrossRef]
- Gu, W.; Luozhong, S.; Cai, S.; Londhe, K.; Elkasri, N.; Hawkins, R.; Yuan, Z.; Su-Greene, K.; Yin, Y.; Cruz, M.; et al. Extracellular vesicles incorporating retrovirus-like capsids for the enhanced packaging and systemic delivery of mRNA into neurons. Nat. Biomed. Eng. 2024, 8, 415–426. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Li, S.; Ou, H.; Zhang, Y.; Zhu, G.; Li, S.; Lei, L. Exosome-based delivery strategies for tumor therapy: An update on modification, loading, and clinical application. J. Nanobiotechnol. 2024, 22, 41. [Google Scholar] [CrossRef] [PubMed]
- Lennaárd, A.J.; Mamand, D.R.; Wiklander, R.J.; El Andaloussi, S.; Wiklander, O.P.B. Optimised electroporation for loading of extracellular vesicles with doxorubicin. Pharmaceutics 2021, 14, 38. [Google Scholar] [CrossRef] [PubMed]
- Lamichhane, T.N.; Raiker, R.S.; Jay, S.M. Exogenous DNA loading into extracellular vesicles via electroporation is size-dependent and enables limited gene delivery. Mol. Pharm. 2015, 12, 3650–3657. [Google Scholar] [CrossRef]
- Torabi, C.; Choi, S.E.; Pisanic, T.R.; Paulaitis, M.; Hur, S.C. Streamlined miRNA loading of surface protein-specific extracellular vesicle subpopulations through electroporation. Biomed. Eng. Online 2024, 23, 116. [Google Scholar] [CrossRef]
- Yuan, R.; Mu, Z.; Zhang, H.; Guo, J.; Tian, Y.; Xin, Q.; Zhu, X.; Dong, Z.; Wang, H.; Shi, Y. Ultrasonic microfluidic method used for siHSP47 loaded in human embryonic kidney cell-derived exosomes for inhibiting TGF-β1 induced fibroblast differentiation and migration. Int. J. Mol. Sci. 2025, 26, 382. [Google Scholar] [CrossRef]
- Lowe, N.M.; Mizenko, R.R.; Nguyen, B.B.; Chiu, K.L.; Arun, V.; Panitch, A.; Carney, R.P. Orthogonal analysis reveals inconsistencies in cargo loading of extracellular vesicles. J. Extracell. Biol. 2024, 3, e70003. [Google Scholar] [CrossRef]
- Jiang, Q.; Sun, Y.; Si, X.; Cui, H.; Li, J.; Bao, Y.; Wang, L.; Li, B. Anthocyanin-loaded milk-derived extracellular vesicles nano-delivery system: Stability, mucus layer penetration, and pro-oxidant effect on HepG2 cells. Food Chem. 2024, 458, 140152. [Google Scholar] [CrossRef]
- Hettich, B.F.; Bader, J.J.; Leroux, J.C. Encapsulation of hydrophilic compounds in small extracellular vesicles: Loading capacity and impact on vesicle functions. Adv. Healthc. Mater. 2022, 11, e2100047. [Google Scholar] [CrossRef]
- Lee, C.; Kumar, S.; Park, J.; Choi, Y.; Clarissa, E.M.; Cho, Y.K. Tonicity-induced cargo loading into extracellular vesicles. Lab Chip 2024, 24, 2069–2079. [Google Scholar] [CrossRef]
- Zhang, Y.; Kim, G.; Zhu, Y.; Wang, C.; Zhu, R.; Lu, X.; Chang, H.C.; Wang, Y. Chiral graphene quantum dots enhanced drug loading into small extracellular vesicles. ACS Nano 2023, 17, 10191–10205. [Google Scholar] [CrossRef] [PubMed]
- Man, F.; Xing, H.; Wang, H.; Wang, J.; Lu, R. Engineered small extracellular vesicles as a versatile platform to efficiently load ferulic acid via an “esterase-responsive active loading” strategy. Front. Bioeng. Biotechnol. 2022, 10, 1043130. [Google Scholar] [CrossRef] [PubMed]
- Kronstadt, S.M.; Jay, S.M.; Jeyaram, A. Extracellular vesicle loading via pH-gradient modification. Methods Mol. Biol. 2022, 2504, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Su, N.; Liu, W.; Li, M.; Zheng, H.; Li, B.; Jin, X.; Gao, M.; Zhang, X. An effective cell-penetrating peptide-based loading method to extracellular vesicles and enhancement in cellular delivery of drugs. Anal. Bioanal. Chem. 2025, 415, 12345. [Google Scholar] [CrossRef]
- Bao, Y.R.; Chen, Y.J.; Deng, X.F.; Wang, Y.K.; Zhang, Y.X.; Xu, L.L.; Huang, W.H.; Cheng, S.B.; Zhang, H.B.; Xie, M. Boron clusters escort doxorubicin squashing into exosomes and overcome drug resistance. Adv. Sci. 2025, 12, e2412501. [Google Scholar] [CrossRef]
- Harrell, C.R.; Volarevic, A.; Djonov, V.; Volarevic, V. Mesenchymal stem-cell-derived exosomes as novel drug carriers in anti-cancer treatment: A myth or reality? Cells 2025, 14, 202. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, G.; Zhang, K.; Cao, Q.; Liu, T.; Li, J. Mesenchymal stem cells-derived exosomes for drug delivery. Stem Cell Res. Ther. 2021, 12, 561. [Google Scholar] [CrossRef]
- Liu, T.; Li, T.; Zheng, Y.; Xu, X.; Sun, R.; Zhan, S.; Guo, X.; Zhao, Z.; Zhu, W.; Feng, B.; et al. Evaluating adipose-derived stem cell exosomes as miRNA drug delivery systems for the treatment of bladder cancer. Cancer Med. 2022, 11, 3687–3699. [Google Scholar] [CrossRef]
- Wei, H.; Chen, F.; Chen, J.; Lin, H.; Wang, S.; Wang, Y.; Wu, C.; Lin, J.; Zhong, G. Mesenchymal stem cell derived exosomes as nanodrug carrier of doxorubicin for targeted osteosarcoma therapy via SDF1-CXCR4 axis. Int. J. Nanomed. 2022, 17, 3483–3495. [Google Scholar] [CrossRef]
- Song, L.L.; Tang, Y.P.; Qu, Y.Q.; Yun, Y.X.; Zhang, R.L.; Wang, C.R.; Wong, V.K.W.; Wang, H.M.; Liu, M.H.; Qu, L.Q.; et al. Exosomal delivery of rapamycin modulates blood-brain barrier penetration and VEGF axis in glioblastoma. J. Control. Release 2025, 381, 113605. [Google Scholar] [CrossRef]
- Zhou, M.; Li, H.; Zhao, J.; Zhang, Q.; Han, Z.; Han, Z.C.; Zhu, L.; Wang, H.; Li, Z. Extracellular vesicles derived from mesenchymal stem cells suppress breast cancer progression by inhibiting angiogenesis. Mol. Med. Rep. 2024, 30, 192. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Wang, M.; Jin, C.; Li, L.; Tang, Y.; Wang, Z.; Wang, X.; Xu, W.; Qian, H. Highly Efficient Delivery of Novel MiR-13896 by Human Umbilical Cord Mesenchymal Stem Cell-Derived Small Extracellular Vesicles Inhibits Gastric Cancer Progression by Targeting ATG2A-Mediated Autophagy. Biomater. Res. 2024, 28, 0119. [Google Scholar] [CrossRef] [PubMed]
- Cui, F.; Chen, Y.; Wu, X.; Zhao, W. Mesenchymal Stem Cell-Derived Exosomes Carrying miR-486-5p Inhibit Glycolysis and Cell Stemness in Colorectal Cancer by Targeting NEK2. BMC Cancer 2024, 24, 1356. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Liu, B.; Dong, B.; Xu, J.; Zhou, H.; Na, S.; Liu, Y.; Pan, Y.; Chen, F.; Li, L.; et al. Exosome-based delivery of natural products in cancer therapy. Front. Cell Dev. Biol. 2021, 9, 650426. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, G.; Wan, Y.; Xia, B.; Ni, Q.; Shan, S.; Hu, Z.; Liang, X.J. Immune cell-derived exosomes as promising tools for cancer therapy. J. Control. Release 2023, 364, 508–528. [Google Scholar] [CrossRef]
- Zhang, L.; Meng, Y.; An, Y.; Yang, X.; Wei, F.; Ren, X. The antitumor effect of extracellular vesicles derived from cytokine-activated CD8+ T cells. J. Leukoc. Biol. 2024, 116, 1033–1044. [Google Scholar] [CrossRef]
- Kim, H.Y.; Min, H.K.; Song, H.W.; Yoo, A.; Lee, S.; Kim, K.P.; Park, J.O.; Choi, Y.H.; Choi, E. Delivery of human natural killer cell-derived exosomes for liver cancer therapy: An in vivo study in subcutaneous and orthotopic animal models. Drug Deliv. 2022, 29, 2897–2911. [Google Scholar] [CrossRef]
- Shi, Y.; Chen, Y.; Wang, Y.; Mo, D.; Ai, H.; Zhang, J.; Guo, M.; Qian, H. Therapeutic effect of small extracellular vesicles from cytokine-induced memory-like natural killer cells on solid tumors. J. Nanobiotechnol. 2024, 22, 447. [Google Scholar] [CrossRef]
- Haney, M.J.; Zhao, Y.; Jin, Y.S.; Li, S.M.; Bago, J.R.; Klyachko, N.L.; Kabanov, A.V.; Batrakova, E.V. Macrophage-derived extracellular vesicles as drug delivery systems for triple negative breast cancer (TNBC) therapy. J. Neuroimmune Pharmacol. 2020, 15, 487–500. [Google Scholar] [CrossRef]
- Gambardella, A.R.; Antonucci, C.; Zanetti, C.; Noto, F.; Andreone, S.; Vacca, D.; Pellerito, V.; Sicignano, C.; Parrottino, G.; Tirelli, V.; et al. IL-33 stimulates the anticancer activities of eosinophils through extracellular vesicle-driven reprogramming of tumor cells. J. Exp. Clin. Cancer Res. 2024, 43, 209. [Google Scholar] [CrossRef]
- Villa, A.; De Mitri, Z.; Vincenti, S.; Crippa, E.; Castiglioni, L.; Gelosa, P.; Rebecchi, M.; Tosi, D.; Brunialti, E.; Oevermann, A.; et al. Canine glioblastoma-derived extracellular vesicles as precise carriers for glioblastoma imaging: Targeting across the blood-brain barrier. Biomed. Pharmacother. 2024, 172, 116201. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Sun, L.; Chen, Y.; Lu, Y. The homologous tumor-derived-exosomes loaded with miR-1270 selectively enhanced the suppression effect for colorectal cancer cells. Cancer Med. 2024, 13, e6936. [Google Scholar] [CrossRef]
- Sulthana, S.; Shrestha, D.; Aryal, S. Maximizing liposome tumor delivery by hybridizing with tumor-derived extracellular vesicles. Nanoscale 2024, 16, 16652–16663. [Google Scholar] [CrossRef]
- Sancho-Albero, M.; Decio, A.; Akpinar, R.; De Luigi, A.; Giavazzi, R.; Terracciano, L.M.; De Cola, L. Melanoma extracellular vesicles membrane coated nanoparticles as targeted delivery carriers for tumor and lungs. Mater. Today Bio 2024, 30, 101433. [Google Scholar] [CrossRef]
- Liang, K.; Sun, Y.; Xie, L.; Liu, Y.; You, Y.; Xu, J.; Ma, F.; Huang, Y.; Song, Q.; Xiao, W.; et al. Biologically self-assembled tumor cell-derived cancer nanovaccines as an all-in-one platform for cancer immunotherapy. ACS Nano 2024, 18, 6702–6717. [Google Scholar] [CrossRef]
- Wang, L.; Wang, W.; Hu, D.; Liang, Y.; Liu, Z.; Zhong, T.; Wang, X. Tumor-derived extracellular vesicles regulate macrophage polarization: Role and therapeutic perspectives. Front. Immunol. 2024, 15, 1346587. [Google Scholar] [CrossRef]
- Zheng, M.; Chavda, V.P.; Vaghela, D.A.; Bezbaruah, R.; Gogoi, N.R.; Patel, K.; Kulkarni, M.; Shen, B.; Singla, R.K. Plant-derived exosomes in therapeutic nanomedicine, paving the path toward precision medicine. Phytomedicine 2024, 135, 156087. [Google Scholar] [CrossRef]
- Luo, Z.; Cheng, X.; Feng, B.; Fan, D.; Liu, X.; Xie, R.; Luo, T.; Wegner, S.V.; Ma, D.; Chen, F.; et al. Engineering versatile bacteria-derived outer membrane vesicles: An adaptable platform for advancing cancer immunotherapy. Adv. Sci. 2024, 11, e2400049. [Google Scholar] [CrossRef]
- Yi, S.; Jung, E.; Kim, H.; Choi, J.; Kim, S.; Lim, E.K.; Kim, K.S.; Kang, T.; Jung, J. Harnessing Lactobacillus reuteri-derived extracellular vesicles for multifaceted cancer treatment. Small 2024, 21, e2406094. [Google Scholar] [CrossRef]
- Shi, Y.; Zhang, C.; Cao, W.; Li, L.; Liu, K.; Zhu, H.; Balcha, F.; Fang, Y. Extracellular vesicles from Lacticaseibacillus paracasei PC-H1 inhibit HIF-1α-mediated glycolysis of colon cancer. Future Microbiol. 2024, 19, 227–239. [Google Scholar] [CrossRef]
- Zhang, J.; Huang, X.; Zhang, T.; Gu, C.; Zuo, W.; Fu, L.; Dong, Y.; Liu, H. Antitumorigenic potential of Lactobacillus-derived extracellular vesicles: p53 succinylation and glycolytic reprogramming in intestinal epithelial cells via SIRT5 modulation. Cell Biol. Toxicol. 2024, 40, 66. [Google Scholar] [CrossRef]
- Fan, Y.L.; Jin, J.X.; Zhu, J.; Ruan, H.B.; Huang, J.Q. Extracellular vesicles of Bifidobacterium longum reverse the acquired carboplatin resistance in ovarian cancer cells via p53 phosphorylation on Ser15. Kaohsiung J. Med. Sci. 2024, 40, 530–541. [Google Scholar] [CrossRef]
- Won, S.; Lee, C.; Bae, S.; Lee, J.; Choi, D.; Kim, M.G.; Song, S.; Lee, J.; Kim, E.; Shin, H.; et al. Mass-produced gram-negative bacterial outer membrane vesicles activate cancer antigen-specific stem-like CD8+ T cells which enables an effective combination immunotherapy with anti-PD-1. J. Extracell. Vesicles 2023, 12, e12357. [Google Scholar] [CrossRef]
- Xiao, T.; Ma, Y.; Zhang, Z.; Zhang, Y.; Zhao, Y.; Zhou, X.; Wang, X.; Ge, K.; Guo, J.; Zhang, J.; et al. Tailoring therapeutics via a systematic beneficial elements comparison between photosynthetic bacteria-derived OMVs and extruded nanovesicles. Bioact. Mater. 2024, 36, 48–61. [Google Scholar] [CrossRef]
- Kim, J.; Li, S.; Zhang, S.; Wang, J. Plant-derived exosome-like nanoparticles and their therapeutic activities. Asian J. Pharm. Sci. 2022, 17, 53–69. [Google Scholar] [CrossRef]
- Kim, D.K.; Kang, S.J.; Rhee, W.J. Perilla-leaf-derived extracellular vesicles selectively inhibit breast cancer cell proliferation and invasion. Int. J. Mol. Sci. 2023, 24, 15633. [Google Scholar] [CrossRef]
- Lu, X.; Yu, Z.; Wang, J.; Tian, A.; Wu, T.; Cheng, Y.; Han, Q.; Li, F.; Xia, W. The unexpected PD-L1 suppression function of celery-derived extracellular vesicles improves lung cancer chemotherapy efficacy. Extracell. Vesicles Circ. Nucl. Acids 2024, 5, 697–713. [Google Scholar] [CrossRef]
- Yang, M.; Guo, J.; Li, J.; Wang, S.; Sun, Y.; Liu, Y.; Peng, Y. Platycodon grandiflorum-derived extracellular vesicles suppress triple-negative breast cancer growth by reversing the immunosuppressive tumor microenvironment and modulating the gut microbiota. J. Nanobiotechnol. 2025, 23, 92. [Google Scholar] [CrossRef]
- Ünsal, N.; Koçak Denizci, P.; Yilmaz, H.; Şahin, F.; Yildirim Canpolat, M. The apoptotic effect of garlic (Allium sativum) derived SEVs on different types of cancer cell lines in vitro. Turk. J. Biol. 2024, 48, 182–191. [Google Scholar] [CrossRef]
- Xu, J.; Yu, Y.; Zhang, Y.; Dai, H.; Yang, Q.; Wang, B.; Ma, Q.; Chen, Y.; Xu, F.; Shi, X.; et al. Oral administration of garlic-derived nanoparticles improves cancer immunotherapy by inducing intestinal IFNγ-producing γδ T cells. Nat. Nanotechnol. 2024, 19, 1569–1578. [Google Scholar] [CrossRef] [PubMed]
- Ou, X.; Wang, H.; Tie, H.; Liao, J.; Luo, Y.; Huang, W.; Yu, R.; Song, L.; Zhu, J. Novel plant-derived exosome-like nanovesicles from Catharanthus roseus: Preparation, characterization, and immunostimulatory effect via TNF-α/NF-κB/PU.1 axis. J. Nanobiotechnol. 2023, 21, 160. [Google Scholar] [CrossRef]
- Yi, C.; Lu, L.; Li, Z.; Guo, Q.; Ou, L.; Wang, R.; Tian, X. Plant-derived exosome-like nanoparticles for microRNA delivery in cancer treatment. Drug Deliv. Transl. Res. 2025, 15, 84–101. [Google Scholar] [CrossRef]
- Cui, L.; Perini, G.; Augello, A.; Palmieri, V.; De Spirito, M.; Papi, M. Plant-derived extracellular nanovesicles: A promising biomedical approach for effective targeting of triple negative breast cancer cells. Front. Bioeng. Biotechnol. 2024, 12, 1390708. [Google Scholar] [CrossRef]
- Wang, F.; Li, L.; Deng, J.; Ai, J.; Mo, S.; Ding, D.; Xiao, Y.; Hu, S.; Zhu, D.; Li, Q.; et al. Lipidomic analysis of plant-derived extracellular vesicles for guidance of potential anti-cancer therapy. Bioact. Mater. 2024, 46, 82–96. [Google Scholar] [CrossRef]
- Chintapula, U.; Oh, D.; Perez, C.; Davis, S.; Ko, J. Anti-cancer bioactivity of sweet basil leaf derived extracellular vesicles on pancreatic cancer cells. J. Extracell. Biol. 2024, 3, e142. [Google Scholar] [CrossRef]
- Li, S.; Tang, Y.; Dou, Y. The potential of milk-derived exosomes for drug delivery. Curr. Drug Deliv. 2021, 18, 688–699. [Google Scholar] [CrossRef]
- Cui, Z.; Amevor, F.K.; Zhao, X.; Mou, C.; Pang, J.; Peng, X.; Liu, A.; Lan, X.; Liu, L. Potential therapeutic effects of milk-derived exosomes on intestinal diseases. J. Nanobiotechnol. 2023, 21, 496. [Google Scholar] [CrossRef]
- González-Sarrías, A.; Iglesias-Aguirre, C.E.; Cortés-Martín, A.; Vallejo, F.; Cattivelli, A.; Del Pozo-Acebo, L.; Del Saz, A.; López de Las Hazas, M.C.; Dávalos, A.; Espín, J.C. Milk-derived exosomes as nanocarriers to deliver curcumin and resveratrol in breast tissue and enhance their anticancer activity. Int. J. Mol. Sci. 2022, 23, 2860. [Google Scholar] [CrossRef]
- Reif, S.; Birimberg-Schwartz, L.; Grunewald, M.; Duran, D.; Sebbag-Sznajder, N.; Toledano, T.; Musseri, M.; Golan-Gerstl, R. The effect of milk-derived extracellular vesicles on intestinal epithelial cell proliferation. Int. J. Mol. Sci. 2024, 25, 13519. [Google Scholar] [CrossRef]
- Ahmed, F.; Tamma, M.; Pathigadapa, U.; Reddanna, P.; Yenuganti, V.R. Drug loading and functional efficacy of cow, buffalo, and goat milk-derived exosomes: A comparative study. Mol. Pharm. 2022, 19, 763–774. [Google Scholar] [CrossRef]
- Nathani, A.; Aare, M.; Sun, L.; Bagde, A.; Li, Y.; Rishi, A.; Singh, M. Unlocking the potential of camel milk-derived exosomes as novel delivery systems: Enhanced bioavailability of ARV-825 PROTAC for cancer therapy. Pharmaceutics 2024, 16, 1070. [Google Scholar] [CrossRef]
- Winkler, R.; Ciria, M.; Ahmad, M.; Plank, H.; Marcuello, C. A Review of the Current State of Magnetic Force Microscopy to Unravel the Magnetic Properties of Nanomaterials Applied in Biological Systems and Future Directions for Quantum Technologies. Nanomaterials 2023, 13, 2585. [Google Scholar] [CrossRef]
- Wang, S.; Mao, Y.; Rong, S.; Liu, G.; Cao, Y.; Yang, Z.; Yu, H.; Zhang, X.; Fang, H.; Cai, Z.; et al. Engineering Magnetic Extracellular Vesicles Mimetics for Enhanced Targeting Chemodynamic Therapy to Overcome Ovary Cancer. ACS Appl. Mater. Interfaces 2024, 16, 39021–39034. [Google Scholar] [CrossRef]
- Ye, C.; Yan, C.; Bian, S.J.; Li, X.R.; Li, Y.; Wang, K.X.; Zhu, Y.H.; Wang, L.; Wang, Y.C.; Wang, Y.Y.; et al. Momordica charantia L.-derived exosome-like nanovesicles stabilize p62 expression to ameliorate doxorubicin cardiotoxicity. J. Nanobiotechnol. 2024, 22, 464. [Google Scholar] [CrossRef]
- Shamshiripour, P.; Rahnama, M.; Nikoobakht, M.; Rad, V.F.; Moradi, A.R.; Ahmadvand, D. Extracellular vesicles derived from dendritic cells loaded with VEGF-A siRNA and doxorubicin reduce glioma angiogenesis in vitro. J. Control. Release 2024, 369, 128–145. [Google Scholar] [CrossRef]
- Wiklander, O.P.B.; Mamand, D.R.; Mohammad, D.K.; Zheng, W.; Jawad Wiklander, R.; Sych, T.; Zickler, A.M.; Liang, X.; Sharma, H.; Lavado, A.; et al. Antibody-displaying extracellular vesicles for targeted cancer therapy. Nat. Biomed. Eng. 2024, 8, 1453–1468. [Google Scholar] [CrossRef]
- Xu, M.; Duan, M.; Chen, M.; Mahal, A.; Yang, L.; Meng, C.; Zhang, Z.; Ren, J.; Obaidullah, A.J.; Li, S.; et al. Study on the activity of targeted delivery of DOX against melanoma by exosome-like nanovesicles of Rhodiola rosea. Biochim. Biophys. Acta Gen. Subj. 2025, 1869, 130776. [Google Scholar] [CrossRef]
- Paschoal Barbosa, L.M.; Gomes, E.R.; Barros, A.L.B.; Cassali, G.D.; Carvalho, A.T.; Silva, J.O.; Pádua, A.L.; Oliveira, M.C. Hybrid nanosystem formed by DOX-loaded liposomes and extracellular vesicles from MDA-MB-231 is effective against breast cancer cells with different molecular profiles. Pharmaceutics 2024, 16, 739. [Google Scholar] [CrossRef]
- Zheng, W.; Zhu, T.; Tang, L.; Li, Z.; Jiang, G.; Huang, X. Inhalable CAR-T cell-derived exosomes as paclitaxel carriers for treating lung cancer. J. Transl. Med. 2023, 21, 383. [Google Scholar] [CrossRef]
- Creeden, J.F.; Sevier, J.; Zhang, J.T.; Lapitsky, Y.; Brunicardi, F.C.; Jin, G.; Nemunaitis, J.; Liu, J.Y.; Kalinoski, A.; Rao, D.; et al. Smart exosomes enhance PDAC targeted therapy. J. Control. Release 2024, 368, 413–429. [Google Scholar] [CrossRef]
- Chen, C.; Pan, X.; Sun, M.; Wang, J.; Su, X.; Zhang, T.; Chen, Y.; Wu, D.; Li, J.; Wu, S.; et al. Phospholipid-anchored ligand conjugation on extracellular vesicles for enhanced cancer targeting. Small 2024, 20, e2310712. [Google Scholar] [CrossRef] [PubMed]
- Talatapeh, S.P.; Rezaie, J.; Nejati, V. Extracellular vesicle-based delivery of paclitaxel to lung cancer cells: Uptake, anticancer effects, autophagy and mitophagy pathways. Arch. Med. Res. 2025, 56, 103194. [Google Scholar] [CrossRef] [PubMed]
- Bhullar, A.S.; Jin, K.; Shi, H.; Jones, A.; Hironaka, D.; Xiong, G.; Xu, R.; Guo, P.; Binzel, D.W.; Shu, D. Engineered extracellular vesicles for combinatorial TNBC therapy: SR-SIM-guided design achieves substantial drug dosage reduction. Mol. Ther. 2024, 32, 4467–4481. [Google Scholar] [CrossRef]
- Hass, R.; von der Ohe, J.; Luo, T. Human mesenchymal stroma/stem-like cell-derived taxol-loaded EVs/exosomes transfer anti-tumor microRNA signatures and express enhanced SDF-1-mediated tumor tropism. Cell Commun. Signal. 2024, 22, 506. [Google Scholar] [CrossRef]
- Sağraç, D.; Kırbaş, O.K.; Öztürkoğlu, D.; Süt, P.A.; Taşlı, P.N.; Şahin, F. Exploring the protective potential of NRF2 overexpressed neural extracellular vesicles against cisplatin-induced neurotoxicity via NRF2/ARE pathway. Toxicology 2024, 508, 153934. [Google Scholar] [CrossRef]
- Nathani, A.; Sun, L.; Khan, I.; Aare, M.; Bagde, A.; Li, Y.; Singh, M. Combined role of interleukin-15 stimulated natural killer cell-derived extracellular vesicles and carboplatin in osimertinib-resistant H1975 lung cancer cells with EGFR mutations. Pharmaceutics 2024, 16, 83. [Google Scholar] [CrossRef]
- Chandler, K.; Millar, J.; Ward, G.; Boyall, C.; White, T.; Ready, J.D.; Maani, R.; Chapple, K.; Tempest, R.; Brealey, J.; et al. Imaging of light-enhanced extracellular vesicle-mediated delivery of oxaliplatin to colorectal cancer cells via laser ablation, inductively coupled plasma mass spectrometry. Cells 2023, 13, 24. [Google Scholar] [CrossRef]
- Tang, Z.G.; Chen, T.M.; Lu, Y.; Wang, Z.; Wang, X.C.; Kong, Y. Human bone marrow mesenchymal stem cell-derived exosomes loaded with gemcitabine inhibit pancreatic cancer cell proliferation by enhancing apoptosis. World J. Gastrointest. Oncol. 2024, 16, 4006–4013. [Google Scholar] [CrossRef]
- Kim, B.; Park, H.; Liu, H.; Kim, S.; Lee, Y.K.; Kim, Y.C. Hybrid nanoparticles of extracellular vesicles and gemcitabine prodrug-loaded liposomes with enhanced targeting ability for effective PDAC treatment. ACS Appl. Bio Mater. 2024, 7, 6025–6033. [Google Scholar] [CrossRef]
- Gaurav, I.; Thakur, A.; Zhang, K.; Thakur, S.; Hu, X.; Xu, Z.; Kumar, G.; Jaganathan, R.; Iyaswamy, A.; Li, M.; et al. Peptide-conjugated vascular endothelial extracellular vesicles encapsulating vinorelbine for lung cancer targeted therapeutics. Nanomaterials 2024, 14, 1669. [Google Scholar] [CrossRef]
- Zhao, M.; Li, Q.; Chai, Y.; Rong, R.; He, L.; Zhang, Y.; Cui, H.; Xu, H.; Zhang, X.; Wang, Z.; et al. An anti-CD19-exosome delivery system navigates the blood-brain barrier for targeting of central nervous system lymphoma. J. Nanobiotechnol. 2025, 23, 173. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Hou, L.; Li, M.; Zhang, J.; Wang, L.; Liu, C.; Luo, T.; Yan, L.; Zheng, L. Broccoli extracellular vesicles enhance the therapeutic effects and restore the chemosensitivity of 5-fluorouracil on colon cancer. Food Chem. Toxicol. 2024, 186, 114563. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Hu, F.; Xiang, J.; Zhou, X.; Wu, B.; Fan, B.; Tang, H.; Liu, B.; Chen, L. Mesoporous microneedles enabled localized controllable delivery of stimulator of interferon gene agonist nanoexosomes for FLASH radioimmunotherapy against breast cancer. ACS Appl. Mater. Interfaces 2024, 16, 58180–58190. [Google Scholar] [CrossRef]
- Yang, Q.; Gao, W.; Li, X.; Li, X.; Zhou, X.; Li, W.; Zhou, C.; Luo, A.; Liu, Z. Targeting ABCA1 via extracellular vesicle-encapsulated staurosporine as a therapeutic strategy to enhance radiosensitivity. Adv. Healthc. Mater. 2024, 13, e2400381. [Google Scholar] [CrossRef]
- Zhang, F.; Zhang, Z.; Yang, W.; Peng, Z.; Sun, J.; Li, G.; Wei, Y.; Wang, X.; Zhao, L.; Xie, W. Engineering autologous cell-derived exosomes to boost melanoma-targeted radio-immunotherapy by cascade cGAS-STING pathway activation. Small 2025, 21, e2408769. [Google Scholar] [CrossRef]
- Oshchepkova, A.; Zenkova, M.; Vlassov, V. Extracellular vesicles for therapeutic nucleic acid delivery: Loading strategies and challenges. Int. J. Mol. Sci. 2023, 24, 7287. [Google Scholar] [CrossRef]
- Huang, H.; Yi, X.; Wei, Q.; Li, M.; Cai, X.; Lv, Y.; Weng, L.; Mao, Y.; Fan, W.; Zhao, M.; et al. Edible and cation-free kiwi fruit derived vesicles mediated EGFR-targeted siRNA delivery to inhibit multidrug resistant lung cancer. J. Nanobiotechnol. 2023, 21, 41. [Google Scholar] [CrossRef]
- Rabienezhad Ganji, N.; Urzì, O.; Tinnirello, V.; Costanzo, E.; Polito, G.; Palumbo Piccionello, A.; Manno, M.; Raccosta, S.; Gallo, A.; Lo Pinto, M.; et al. Proof-of-concept study on the use of tangerine-derived nanovesicles as siRNA delivery vehicles toward colorectal cancer cell line SW480. Int. J. Mol. Sci. 2023, 25, 546. [Google Scholar] [CrossRef]
- Jiang, J.; Lu, Y.; Chu, J.; Zhang, X.; Xu, C.; Liu, S.; Wan, Z.; Wang, J.; Zhang, L.; Liu, K.; et al. Anti-EGFR ScFv functionalized exosomes delivering LPCAT1 specific siRNAs for inhibition of lung cancer brain metastases. J. Nanobiotechnol. 2024, 22, 159. [Google Scholar] [CrossRef]
- Kim, G.; Zhu, R.; Yu, S.; Fan, B.; Jeon, H.; Leon, J.; Webber, M.J.; Wang, Y. Enhancing gene delivery to breast cancer with highly efficient siRNA loading and pH-responsive small extracellular vesicles. ACS Biomater. Sci. Eng. 2025, 11, 213–227. [Google Scholar] [CrossRef]
- Rahmani, R.; Kiani, J.; Tong, W.Y.; Soleimani, M.; Voelcker, N.H.; Arefian, E. Engineered anti-EGFRvIII targeted exosomes induce apoptosis in glioblastoma multiforme. J. Drug Target. 2023, 31, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Gong, L.; Chen, Z.; Feng, K.; Luo, L.; Zhang, J.; Yuan, J.; Ren, Y.; Wang, Y.; Zheng, X.; Li, Q. A versatile engineered extracellular vesicle platform simultaneously targeting and eliminating senescent stromal cells and tumor cells to promote tumor regression. J. Nanobiotechnol. 2024, 22, 105. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Choi, Y.; Kim, K.H.; Byun, Y.; Kim, T.H.; Kim, J.H.; An, S.H.; Bae, D.; Choi, M.K.; Lee, M.; et al. Scalable production of siRNA-encapsulated extracellular vesicles for the inhibition of KRAS-mutant cancer using acoustic shock waves. J. Extracell. Vesicles 2024, 13, e12508. [Google Scholar] [CrossRef] [PubMed]
- Taghavi-Farahabadi, M.; Mahmoudi, M.; Mojtabavi, N.; Noorbakhsh, F.; Ghanbarian, H.; Koochaki, A.; Hashemi, S.M.; Rezaei, N. Enhancing the anti-tumor activity and reprogramming M2 macrophages by delivering siRNAs against SIRPα and STAT6 via M1 exosomes and combining with anti-PD-L1. Life Sci. 2025, 361, 123311. [Google Scholar] [CrossRef]
- Liu, M.; Hu, S.; Yan, N.; Popowski, K.D.; Cheng, K. Inhalable extracellular vesicle delivery of IL-12 mRNA to treat lung cancer and promote systemic immunity. Nat. Nanotechnol. 2024, 19, 565–575. [Google Scholar] [CrossRef]
- Xing, Y.; Zhang, F.; Ji, P.; Wei, M.; Yin, C.; Yang, A.; Yang, G.; Zhao, J. Efficient delivery of GSDMD-N mRNA by engineered extracellular vesicles induces pyroptosis for enhanced immunotherapy. Small 2023, 19, e2204031. [Google Scholar] [CrossRef]
- Liu, M.; Zhang, Y.; He, J.; Liu, W.; Li, Z.; Zhang, Y.; Gu, A.; Zhao, M.; Liu, M.; Liu, X. Fusion with ARRDC1 or CD63: A strategy to enhance p53 loading into extracellular vesicles for tumor suppression. Biomolecules 2024, 14, 591. [Google Scholar] [CrossRef]
- Zhou, L.; Zhang, X.; Wang, Z.; Li, D.; Zhou, G.; Liu, H. Extracellular vesicle-mediated delivery of miR-766-3p from bone marrow stromal cells as a therapeutic strategy against colorectal cancer. Cancer Cell Int. 2024, 24, 330. [Google Scholar] [CrossRef]
- Zhou, P.; Du, X.; Jia, W.; Feng, K.; Zhang, Y. Engineered extracellular vesicles for targeted reprogramming of cancer-associated fibroblasts to potentiate therapy of pancreatic cancer. Signal Transduct. Target. Ther. 2024, 9, 151. [Google Scholar] [CrossRef]
- Liu, X.; Cao, Z.; Wang, W.; Zou, C.; Wang, Y.; Pan, L.; Jia, B.; Zhang, K.; Zhang, W.; Li, W.; et al. Engineered extracellular vesicle-delivered CRISPR/Cas9 for radiotherapy sensitization of glioblastoma. ACS Nano 2023, 17, 16432–16447. [Google Scholar] [CrossRef]
- Liu, X.; Liu, X.; Luo, X.; Zhu, M.; Liu, N.; Li, J.; Zhang, Q.; Zou, C.; Wu, Y.; Cao, Z.; et al. Synergistic strategies for glioblastoma treatment: CRISPR-based multigene editing combined with immune checkpoint blockade. J. Nanobiotechnol. 2025, 23, 94. [Google Scholar] [CrossRef] [PubMed]
- Qian, X.; Zhang, Z.; Zhao, L.; Qin, Y.; Cai, H.; Geng, Z.; Zhu, X.; Zhang, W.; Zhang, Y.; Tan, J.; et al. Tropism-facilitated delivery of CRISPR/Cas9 system with chimeric antigen receptor-extracellular vesicles against B-cell malignancies. J. Control. Release 2020, 326, 455–467. [Google Scholar] [CrossRef]
- Yue, M.; Hu, S.; Sun, H.; Tuo, B.; Jia, B.; Chen, C.; Wang, W.; Liu, J.; Liu, Y.; Sun, Z.; et al. Extracellular vesicles remodel tumor environment for cancer immunotherapy. Mol. Cancer 2023, 22, 203. [Google Scholar] [CrossRef]
- Peng, Z.; Zhao, T.; Gao, P.; Zhang, G.; Wu, X.; Tian, H.; Qu, M.; Tan, X.; Zhang, Y.; Zhao, X.; et al. Tumor-derived extracellular vesicles enable tumor tropism chemo-genetherapy for local immune activation in triple-negative breast cancer. ACS Nano 2024, 18, 30943–30956. [Google Scholar] [CrossRef]
- Li, J.; Yang, R.; Dong, F.; Qiu, Q.; Jiang, Z.; Ren, H.; Zhang, C.; Liu, G.; Lovell, J.F.; Zhang, Y. Enzyme-dynamic extracellular vesicles for metalloimmunotherapy of malignant pleural effusions. ACS Nano 2024, 18, 21855–21872. [Google Scholar] [CrossRef]
- Song, H.; Chen, L.; Pan, X.; Shen, Y.; Ye, M.; Wang, G.; Cui, C.; Zhou, Q.; Tseng, Y.; Gong, Z.; et al. Targeting tumor monocyte-intrinsic PD-L1 by rewiring STING signaling and enhancing STING agonist therapy. Cancer Cell 2025, 43, 503–518. [Google Scholar] [CrossRef]
- Ma, D.; Xie, A.; Lv, J.; Min, X.; Zhang, X.; Zhou, Q.; Gao, D.; Wang, E.; Gao, L.; Cheng, L.; et al. Engineered extracellular vesicles enable high-efficient delivery of intracellular therapeutic proteins. Protein Cell 2024, 15, 724–743. [Google Scholar] [CrossRef]
- Qian, L.; Zhang, Z.; Zhang, R.; Zheng, X.; Xiao, B.; Zhang, X.; Wu, Y.; Chen, Y.; Zhang, X.; Zhou, P.; et al. Activated STING-containing R-EVs from iPSC-derived MSCs promote antitumor immunity. Cancer Lett. 2024, 597, 217081. [Google Scholar] [CrossRef]
- Zhang, J.; Wan, S.; Zhou, H.; Du, J.; Li, Y.; Zhu, H.; Weng, L.; Ding, X.; Wang, L. Programmed nanocloak of commensal bacteria-derived nanovesicles amplify strong immunoreactivity against tumor growth and metastatic progression. ACS Nano 2024, 18, 9613–9626. [Google Scholar] [CrossRef]
- Wei, X.; Sun, L.; Deng, J.; Yang, Q.; Zhao, J.; Zhou, S. A multifunctional exosome with dual homeostasis disruption augments cGAS-STING-mediated tumor immunotherapy by boosting ferroptosis. Nano Lett. 2024, 24, 14263–14272. [Google Scholar] [CrossRef]
- Jayasinghe, M.K.; Lay, Y.S.; Liu, D.X.T.; Lee, C.Y.; Gao, C.; Yeo, B.Z.; How, F.Y.X.; Prajogo, R.C.; Hoang, D.V.; Le, H.A.; et al. Extracellular vesicle surface display enhances the therapeutic efficacy and safety profile of cancer immunotherapy. Mol. Ther. 2024, 32, 3558–3579. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Xi, W.; Ni, J.; Jiang, J.; Lei, Y.; Li, L.; Chu, J.; Li, R.; An, Y.; Ouyang, Y.; et al. Genetically modified extracellular vesicles loaded with activated gasdermin D potentially inhibit prostate-specific membrane antigen-positive prostate carcinoma growth and enhance immunotherapy. Biomaterials 2025, 315, 122894. [Google Scholar] [CrossRef] [PubMed]
- Garaeva, L.; Komarova, E.; Emelianova, S.; Putevich, E.; Konevega, A.L.; Margulis, B.; Guzhova, I.; Shtam, T. Grapefruit-derived vesicles loaded with recombinant HSP70 activate antitumor immunity in colon cancer in vitro and in vivo. Biomedicines 2024, 12, 2759. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Lv, T.; Li, Z.; Wei, X.; Yang, C.; Li, W.; Hou, X.; Wang, Z.; Qian, R. Acidity-activatable dynamic hybrid nanoplatforms derived from extracellular vesicles of M1 macrophages enhance cancer immunotherapy through synergistic triple immunotherapy. J. Nanobiotechnol. 2024, 22, 430. [Google Scholar] [CrossRef]
- Sato, K.; Ogawa, K.; Tagami, T.; Ozeki, T. Photothermal therapeutic effect by gold nanostars/extracellular vesicles nanocomplex on melanoma cells. J. Pharm. Sci. 2025, 114, 1391–1397. [Google Scholar] [CrossRef]
- Bi, Y.; Chen, J.; Li, Q.; Li, Y.; Zhang, L.; Liu, Z.; Yuan, F.; Zhang, R. Tumor-derived extracellular vesicle drug delivery system for chemo-photothermal-immune combination cancer treatment. iScience 2024, 27, 108833. [Google Scholar] [CrossRef]
- Lu, M.; Xing, H.; Shao, W.; Wu, P.; Fan, Y.; He, H.; Barth, S.; Zheng, A.; Liang, X.J.; Huang, Y. Antitumor synergism between PAK4 silencing and immunogenic phototherapy of engineered extracellular vesicles. Acta Pharm. Sin. B 2023, 13, 3945–3955. [Google Scholar] [CrossRef]
- Lee, Y.H.; Huang, C.Y. Engineered perfluorochemical cancer-derived exosomes loaded with indocyanine green and camptothecin provide targeted photochemotherapy for effective cancer treatment. Int. J. Nanomed. 2025, 20, 327–342. [Google Scholar] [CrossRef]
- Dang, X.T.T.; Phung, C.D.; Lim, C.M.H.; Jayasinghe, M.K.; Ang, J.; Tran, T.; Schwarz, H.; Le, M.T.N. Dendritic cell-targeted delivery of antigens using extracellular vesicles for anti-cancer immunotherapy. Cell Prolif. 2024, 57, e13622. [Google Scholar] [CrossRef]
- Wang, H.; Mu, J.; Chen, Y.; Liu, Y.; Li, X.; Li, H.; Cao, P. Hybrid ginseng-derived extracellular vesicles-like particles with autologous tumor cell membrane for personalized vaccination to inhibit tumor recurrence and metastasis. Adv. Sci. 2024, 11, e2308235. [Google Scholar] [CrossRef]
- Han, J.; Kim, S.; Hwang, Y.H.; Kim, S.A.; Lee, Y.; Kim, J.; Cho, S.; Woo, J.; Jeong, C.; Kwon, M.; et al. Novel personalized cancer vaccine using tumor extracellular vesicles with attenuated tumorigenicity and enhanced immunogenicity. Adv. Sci. 2024, 11, e2308662. [Google Scholar] [CrossRef] [PubMed]
- Ding, X.; Zhang, J.; Wan, S.; Wang, X.; Wang, Z.; Pu, K.; Wang, M.; Cao, Y.; Weng, L.; Zhu, H.; et al. Non-discriminating engineered masking of immuno-evasive ligands on tumour-derived extracellular vesicles enhances tumour vaccination outcomes. Nat. Nanotechnol. 2025, 20, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Chen, H.; Ming, R.; Wang, W.; Liu, S.; Jing, Y.; Yan, Z.; Lu, G.; Huang, L.L. A self-adjuvant multiantigenic nanovaccines simultaneously activate the antiviral and antitumor immunity for the treatment of cancers. J. Nanobiotechnol. 2025, 23, 150. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liu, S.; Ming, R.; Wang, W.; Wang, C.; Li, C.; Yang, J.; Zhang, F.; Lu, G.; Mei, L.; et al. Engineered virus-mimicking nanovaccine with lymph node-tumor dual-targeting and STING-activating capacity for robust cancer immunotherapy. J. Control. Release 2025, 378, 416–427. [Google Scholar] [CrossRef]
- Molina, E.; Tejero, M.; Duzenli, O.F.; Kuoch, H.; Caine, C.; Krotova, K.; Paulaitis, M.; Aslanidi, G. Insights in AAV-mediated antigen-specific immunity and a strategy for AAV vaccine dose reduction through AAV-extracellular vesicle association. Mol. Ther. Methods Clin. Dev. 2024, 32, 101358. [Google Scholar] [CrossRef]
- Mathlouthi, S.; Kuryk, L.; Prygiel, M.; Lupo, M.G.; Zasada, A.A.; Pesce, C.; Ferri, N.; Rinner, B.; Salmaso, S.; Garofalo, M. Extracellular vesicles powered cancer immunotherapy: Targeted delivery of adenovirus-based cancer vaccine in humanized melanoma model. J. Control. Release 2024, 376, 777–793. [Google Scholar] [CrossRef]
- Peng, B.; Yang, Y.; Wu, Z.; Tan, R.; Pham, T.T.; Yeo, E.Y.M.; Pirisinu, M.; Jayasinghe, M.K.; Pham, T.C.; Liang, K.; et al. Red blood cell extracellular vesicles deliver therapeutic siRNAs to skeletal muscles for treatment of cancer cachexia. Mol. Ther. 2023, 31, 1418–1436. [Google Scholar] [CrossRef]
- Zhang, X.; Taylor, H.; Valdivia, A.; Dasari, R.; Buckley, A.; Bonacquisti, E.; Nguyen, J.; Kanchi, K.; Corcoran, D.L.; Herring, L.E.; et al. Auto-loaded TRAIL-exosomes derived from induced neural stem cells for brain cancer therapy. J. Control. Release 2024, 372, 433–445. [Google Scholar] [CrossRef]
- Duong, A.; Parmar, G.; Kirkham, A.M.; Burger, D.; Allan, D.S. Registered clinical trials investigating treatment with cell-derived extracellular vesicles: A scoping review. Cytotherapy 2023, 25, 939–945. [Google Scholar] [CrossRef]
- Besse, B.; Charrier, M.; Lapierre, V.; Dansin, E.; Lantz, O.; Planchard, D.; Le Chevalier, T.; Livartoski, A.; Barlesi, F.; Laplanche, A.; et al. Dendritic cell-derived exosomes as maintenance immunotherapy after first line chemotherapy in NSCLC. Oncoimmunology 2015, 5, e1071008. [Google Scholar] [CrossRef]
- Dai, S.; Wei, D.; Wu, Z.; Zhou, X.; Wei, X.; Huang, H.; Li, G. Phase I clinical trial of autologous ascites-derived exosomes combined with GM-CSF for colorectal cancer. Mol. Ther. 2008, 16, 782–790. [Google Scholar] [CrossRef] [PubMed]
- Escudier, B.; Dorval, T.; Chaput, N.; André, F.; Caby, M.P.; Novault, S.; Flament, C.; Leboulaire, C.; Borg, C.; Amigorena, S.; et al. Vaccination of metastatic melanoma patients with autologous dendritic cell (DC) derived-exosomes: Results of the first phase I clinical trial. J. Transl. Med. 2005, 3, 10. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Wu, F.; Hu, G.; Chen, L.; Xu, J.; Xu, P.; Wang, X.; Li, Y.; Liu, S.; Zhang, S.; et al. Autologous Tumor Cell-Derived Microparticle-Based Targeted Chemotherapy in Lung Cancer Patients with Malignant Pleural Effusion. Sci. Transl. Med. 2019, 11, eaat5690. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Zhang, H.; Zhou, N.; Xu, P.; Wang, J.; Gao, Y.; Jin, X.; Liang, X.; Lv, J.; Zhang, Y.; et al. Methotrexate-Loaded Tumour-Cell-Derived Microvesicles Can Relieve Biliary Obstruction in Patients with Extrahepatic Cholangiocarcinoma. Nat. Biomed. Eng. 2020, 4, 743–753. [Google Scholar] [CrossRef]
- ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT03608631 (accessed on 28 February 2025).
- ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT01294072 (accessed on 28 February 2025).
- ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT01159288 (accessed on 28 February 2025).
- ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT05375604 (accessed on 28 February 2025).
- Kim, H.I.; Park, J.; Zhu, Y.; Wang, X.; Han, Y.; Zhang, D. Recent Advances in Extracellular Vesicles for Therapeutic Cargo Delivery. Exp. Mol. Med. 2024, 56, 836–849. [Google Scholar] [CrossRef]
- Burnouf, T.; Agrahari, V.; Agrahari, V. Extracellular Vesicles As Nanomedicine: Hopes And Hurdles In Clinical Translation. Int J Nanomedicine 2019, 14, 8847–8859. [Google Scholar] [CrossRef]
- Greenberg, Z.F.; Graim, K.S.; He, M. Towards artificial intelligence-enabled extracellular vesicle precision drug delivery. Adv. Drug Deliv. Rev. 2023, 199, 114974. [Google Scholar] [CrossRef]
- Ma, Y.; Dong, S.; Grippin, A.J.; Teng, L.; Lee, A.S.; Kim, B.Y.S.; Jiang, W. Engineering Therapeutical Extracellular Vesicles for Clinical Translation. Trends Biotechnol. 2025, 43, 61–82. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EV Source | EV Type | Key Advantages/Mechanisms | Cancer Model | Ref. |
|---|---|---|---|---|
| Rat adipose-derived MSCs | EVs | Specific targeting and penetration into bladder cancer tissues for RNA agent delivery | Mouse bladder cancer model | [68] |
| Mouse bone marrow MSCs | EVs | Osteosarcoma targeting via SDF1-CXCR4 axis; reduced chemotherapy-induced toxicity | Mouse osteosarcoma model | [69] |
| Human placental MSCs | EVs | Anti-angiogenic effects; inhibition of tumor cell proliferation and migration | Mouse breast cancer model | [71] |
| Human NK cells | EVs | Induced tumor apoptosis via serine/threonine kinase dephosphorylation; active liver cancer targeting | Mouse hepatocellular carcinoma model | [77] |
| CD8+ T cells | EVs | Delivered granzyme B, perforin, and IFNγ to induce tumor apoptosis | Breast cancer cells | [76] |
| Human memory-like NK cells | EVs | Triggered caspase-dependent apoptosis | Mouse NSCLC and gastric cancer models | [72] |
| Rat adipose-derived MSCs | EVs | Specific targeting and penetration into bladder cancer tissues for RNA agent delivery | Mouse bladder cancer model | [68] |
| Mouse/human eosinophils | EVs | Arrested tumor cell cycle at G0/G1 phase; suppressed proliferation and tumor spheroid formation | Mouse melanoma model | [80] |
| Human colorectal cancer cells | EVs | Tumor-specific delivery of miRNA therapeutics | Mouse colorectal cancer model | [81] |
| Mouse breast cancer cells | EVs | Intrinsic breast cancer tissue targeting | Mouse breast cancer model | [83] |
| Canine glioma cells | EVs | Penetrated blood–brain barrier (BBB); selective accumulation in brain tumors | Mouse glioma model | [85] |
| Melanoma cells | EVs | Targeted primary melanoma and lung metastatic nodules | Mouse melanoma model | [84] |
| Lactobacillus reuteri | EVs | Modulated apoptotic signaling; oral administration with gastrointestinal stability | Mouse NSCLC model | [89] |
| Lactobacillus paracasei | EVs | Inhibited HIF-1α-mediated glycolysis | Mouse colon cancer model | [90] |
| Lactobacillus plantarum | EVs | Impaired p53 desuccinylation, suppressing glycolysis and proliferation | Mouse colon cancer model | [91] |
| Bifidobacterium longum | EVs | Enhanced p53 Ser15 phosphorylation; sensitized ovarian cancer cells to carboplatin | Ovarian cancer cell model | [92] |
| Escherichia coli | OMVs | Activated CD8+ T-cell infiltration and antitumor immunity | Mouse bladder/breast cancer models | [93] |
| Photosynthetic bacteria | OMVs | Polarized macrophages to M1 phenotype; promoted pro-inflammatory cytokine release | Mouse breast cancer model | [94] |
| Catharanthus roseus (plant) | PELNVs | Mitigated chemotherapy-induced immunosuppression by modulating TNF-α/NF-κB/Spi-B signaling | In vitro | [101] |
| Perilla leaves | EVs | Selectively inhibited breast cancer cell proliferation and invasion | Human breast cancer cell model | [96] |
| Celery | EVs | Blocked PD-L1/PD-1 interaction; reversed T-cell suppression | Mouse lung cancer model | [97] |
| Platycodon root | EVs | Induced ROS-mediated apoptosis; repolarized TAMs to M1 phenotype; tumor targeting | Mouse breast cancer model | [98] |
| Lemon | EVs | Inhibited PI3K/AKT and MAPK/ERK pathways; suppressed proliferation and metastasis | Human breast cancer cell model | [103] |
| Ginger | EVs | Delivered cytotoxic gingerols/shogaols; induced cell cycle arrest and p53-mediated apoptosis | Mouse melanoma model | [104] |
| Basil plant | EVs | Efficient tumor uptake; promoted apoptosis | Human pancreatic cancer cells | [105] |
| Garlic | EVs | Activated caspase-dependent apoptosis in multiple cancer types | Human liver, neuroblastoma, pancreatic, glioblastoma, prostate cancers; HUVECs | [99] |
| Cow milk | EVs | Protected polyphenols from degradation; enhanced therapeutic efficacy | Human breast cancer cells | [108] |
| Method | Drug | EV Source | EV Type | Therapeutic Effects | Cancer Type | Ref. |
|---|---|---|---|---|---|---|
| Chemotherapy | DOX | Bitter melon | EVs | Suppressed tumor growth and alleviated cardiotoxicity | Breast cancer | [114] |
| DOX | Dendritic cells | EVs | Reduced tumor angiogenesis and induced apoptosis | Glioma | [115] | |
| DOX | FreeStyle 293F cells | PD-L1 antibody-engineered Fc-EVs | Enhanced tumor suppression with reduced systemic toxicity | Melanoma | [116] | |
| DOX | Rhodiola rosea | pYEEIE-modified exosome-like vesicles | Tumor-targeted delivery and growth inhibition | Melanoma | [117] | |
| DOX | Human breast cancer cells | EVs | Synergistic tumor suppression and toxicity mitigation | Breast cancer | [118] | |
| PTX | T cells | CAR-modified EVs | Targeted tumor inhibition and reduced adverse effects | Lung cancer | [119] | |
| PTX | HEK293 cells | RGD- and CD47-engineered EVs | Tumor-specific targeting and growth suppression | Pancreatic ductal adenocarcinoma | [120] | |
| PTX | Cow milk | Transferrin-conjugated EVs | Enhanced drug delivery and tumor suppression | Liver cancer | [121] | |
| PTX | Lung cancer cells | Large EVs | Inhibited autophagy pathways and induced apoptosis | Lung cancer | [122] | |
| PTX + SUR-siRNA | HEK293T cells | CD44-modified EVs | Dual suppression of tumor growth and chemoresistance | Breast cancer | [123] | |
| PTX | Human MSCs | EVs | Tumor growth inhibition | Breast cancer | [124] | |
| Carboplatin | NK cells | EVs | Enhanced antitumor efficacy | Lung cancer | [126] | |
| Oxaliplatin | Human colorectal cancer cells | Porphyrin-modified EVs | Tumor suppression with reduced toxicity | Colorectal cancer | [127] | |
| Gemcitabine | Human MSCs | EVs | Promoted apoptosis and reduced off-target effects | Pancreatic cancer | [128] | |
| Gemcitabine | Human pancreatic cancer cells | Exosome–liposome hybrid nanoparticles | Precision targeting and tumor growth inhibition | Pancreatic ductal adenocarcinoma | [129] | |
| MTX | hAMSCs | CD19-modified exosomes | Blood–brain barrier (BBB) penetration and tumor-specific delivery | CNS lymphoma (CNSL) | [131] | |
| Vinorelbine | Human umbilical endothelial cells | GE11 peptide-modified EVs | Tumor-targeted delivery with minimal toxicity | Lung cancer | [130] | |
| 5-FU | Broccoli | EVs | Enhanced chemosensitivity and tumor suppression | Colorectal cancer | [132] | |
| Radiotherapy | MSA-2 | Human breast cancer cells | EVs | Reduced radioresistance and optimized FLASH radiotherapy | Breast cancer | [133] |
| STS | Human esophageal cancer cells | EVs | Improved radiosensitivity and therapeutic efficacy | Esophageal cancer | [134] | |
| Mn2+,2′,3′-cGAMP | Melanoma cells | EVs | Overcame immunosuppression and enhanced radioimmunotherapy | Melanoma | [135] | |
| Gene Therapy | siSTAT3 | Kiwifruit | EVs | Tumor growth inhibition | Non-small cell lung cancer (NSCLC) | [177] |
| DDHD1-siRNA | Orange juice | EVs | Suppressed tumor proliferation | Colorectal cancer | [138] | |
| Mstn-siRNA | Red blood cells | EVs | Reduced cachexia-associated toxicity | Cancer cachexia | [177] | |
| TRAIL | Neural stem cells | EVs | Selective tumor targeting and growth suppression | Brain cancer | [178] | |
| siSIRPα + siSTAT6 | M1 macrophages | EVs | Dual inhibition of tumor growth and immunosuppression | Breast cancer | [144] | |
| siRNA (KRASG12C) | Cow milk | EVs | KRAS oncogene silencing and tumor suppression | NSCLC | [143] | |
| LPCAT1-siRNA | HEK293T cells | EGFR scFv-modified EVs | BBB penetration and tumor-specific delivery with low toxicity | Lung cancer brain metastasis | [138] | |
| siR-DG | Human breast cancer cells | Chiral graphene quantum dot/pH-responsive peptide-modified EVs | Targeted tumor cell apoptosis | Breast cancer | [137] | |
| miR-34a + CDAmiR | Human MSCs | EGFRvIII antibody-modified EVs | Precision targeting and apoptosis induction | Glioblastoma | [141] | |
| Src siRNA | Human MSCs | uPA peptide-modified EVs | Tumor cell apoptosis and metastasis inhibition | Breast cancer | [142] | |
| IL-12 mRNA | — | EVs | Enhanced antitumor immunity with reduced toxicity | Lung cancer | [139] | |
| GSDMD-N mRNA | — | EVs | Induced pyroptosis and tumor suppression | Breast cancer | [146] | |
| p53 mRNA | HEK293T cells | ARRDC1- and CD63-engineered EVs | Restored p53 function and tumor apoptosis | Lung cancer | [147] | |
| miR-13896 | Human MSCs | EVs | Autophagy inhibition and tumor growth suppression | Gastric cancer | [72] | |
| miR-766-3p | Human MSCs | EVs | Suppressed proliferation and promoted apoptosis | Colorectal cancer | [148] | |
| Cas9/sgRNA | HEK293T cells | Angiopep-2- and TAT peptide-modified EVs | BBB penetration and tumor growth inhibition | Glioblastoma | [147] | |
| Cas9/sgRNA | HEK293T cells | PD-1- and angiopep-2-modified EVs | Anti-angiogenic effects and tumor apoptosis | Glioblastoma | [151] | |
| Cas9/sgRNA | HEK293T cells | CAR-modified EVs | Reduced proliferation and enhanced apoptosis in B-cell malignancies | B-cell malignancies | [152] | |
| Immunotherapy | siSTAT3 + DOX | Mouse breast cancer cells | EVs | Reversed immunosuppressive TME and suppressed tumor growth | Breast cancer | [154] |
| SOD + diABZI-2 + siSTAT3 | M1 macrophages | EVs | Alleviated immunosuppressive TME and inhibited tumor progression | Malignant pleural effusion | [155] | |
| cGAS | HEK293T cells | EVs | Activated cGAS-STING pathway to remodel TME and suppress tumorigenesis | Colon cancer | [157] | |
| Activated STING | MSCs | EVs | Induced IFNβ expression and enhanced antitumor immunity | Breast cancer | [158] | |
| — | Bacteroides fragilis | Responsive nanocloak-coated EVs | Stimulated cGAS-STING signaling and inhibited tumor growth | Breast cancer | [159] | |
| MnIO + GW4869 + BSO | Mouse breast cancer cells | EVs | Promoted tumor immunogenicity and ferroptosis | Breast cancer | [160] | |
| N-GSDMD | HEK293T | PAMAscFv-modified EVs | Induced pyroptosis and suppressed tumor proliferation | Prostate cancer | [162] | |
| Photothermal Therapy | HSP70 | Grapefruit | EVs | Elicited antitumor immunity and suppressed tumor growth | Colon cancer | [163] |
| REV + SR780Fe | M1 macrophages | RS17 peptide-engineered EVs | Enhanced T-lymphocyte infiltration and tumor suppression | Breast cancer | [164] | |
| — | HEK293T cells | PEG-GNS-modified EVs | Generated cytotoxic effects under laser irradiation | Melanoma | [165] | |
| Melanin + paclitaxel albumin (PA) | Mouse breast cancer cells | EVs | Near-infrared (NIR)-activated CD8+ T-cell infiltration and antitumor efficacy | Breast cancer | [166] | |
| Cancer Vaccines | — | M1 macrophages | siPAK4- and TPC-modified EVs | Induced PAK4 silencing and immunogenic phototherapy | Melanoma | [167] |
| ICG + CPT + PFC | Melanoma cells | EVs | NIR-triggered tumor suppression without systemic toxicity | Melanoma | [168] | |
| OVA | Mouse red blood cells | αDECab-engineered EVs | Enhanced DC phagocytosis of tumor antigens via TLR4-mediated maturation, enabling durable tumor protection | — | [169] | |
| Tumor antigens + G-EVLP | Ginseng | EV-like hybrid membrane particles | Boosted adaptive immunity to inhibit tumor recurrence and metastasis | Melanoma, breast cancer | [170] | |
| — | Mouse thymoma cells | EVs | Induced tumor-specific, long-lasting immunity against recurrence | Thymoma | [171] | |
| — | Mouse breast cancer cells | FeOOH-modified EVs | Activated DC and macrophage responses, prolonging survival and delaying tumor growth | Breast cancer | [172] | |
| — | VSV-infected mouse breast cancer cells | EVs | Accumulated in tumors and lymph nodes, triggering innate/adaptive immune responses | Breast cancer | [173] | |
| OVA, TRP-1 | Melanoma cells | EVs | Activated antigen-specific immunity and improved survival | Melanoma | [175] | |
| Oncolytic virus | Melanoma cells | EVs | Enhanced lymphocyte infiltration and antitumor efficacy | Melanoma | [176] |
| NCT Number | EV Type | Disease Type | Intervention/ Treatment | Sponsor | Status | Ref. |
|---|---|---|---|---|---|---|
| NCT01159288 | Dendritic cell-derived exosomes loaded with antigen | Non-Small-Cell Lung Cancer | Biological: Dex2 | Gustave Roussy, Cancer Campus, Grand Paris | Completed; Phase 2 | [187] |
| NCT03608631 | Mesenchymal–stromal cell-derived exosomes loaded with siRNA against KrasG12D | Metastatic Pancreatic Adenocarcinoma; Pancreatic Ductal Adenocarcinoma | Drug: Mesenchymal–stromal cell-derived exosomes with KRAS G12D siRNA | M.D. Anderson Cancer Center | Active not recruiting; Phase 1 | [185] |
| NCT01294072 | Plant exosomes loaded with curcumin | Colon Cancer | Dietary supplement: curcumin Dietary supplement: curcumin conjugated with plant exosomes Other: no intervention | University of Louisville | Recruiting; NA | [186] |
| NCT05375604 | Cell-derived exosomes loaded with a synthetic lipid-tagged oligonucleotide | Advanced Hepatocellular Carcinoma (HCC) Gastric Cancer Metastatic to Liver Colorectal Cancer Metastatic to Liver | Drug: CDK-004 | Codiak BioSciences | Terminated; Phase 1 | [188] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, J.; Jin, Z.; Fu, T.; Qian, Y.; Bian, X.; Zhang, X.; Zhang, J. Extracellular Vesicle-Based Drug Delivery Systems in Cancer Therapy. Int. J. Mol. Sci. 2025, 26, 4835. https://doi.org/10.3390/ijms26104835
Wu J, Jin Z, Fu T, Qian Y, Bian X, Zhang X, Zhang J. Extracellular Vesicle-Based Drug Delivery Systems in Cancer Therapy. International Journal of Molecular Sciences. 2025; 26(10):4835. https://doi.org/10.3390/ijms26104835
Chicago/Turabian StyleWu, Jiahao, Zhesi Jin, Tingyu Fu, Yu Qian, Xinyue Bian, Xu Zhang, and Jiahui Zhang. 2025. "Extracellular Vesicle-Based Drug Delivery Systems in Cancer Therapy" International Journal of Molecular Sciences 26, no. 10: 4835. https://doi.org/10.3390/ijms26104835
APA StyleWu, J., Jin, Z., Fu, T., Qian, Y., Bian, X., Zhang, X., & Zhang, J. (2025). Extracellular Vesicle-Based Drug Delivery Systems in Cancer Therapy. International Journal of Molecular Sciences, 26(10), 4835. https://doi.org/10.3390/ijms26104835