
Genome-Wide Identification of UGT Gene Family and Functional Analysis of PgUGT29 in Platycodon grandiflorus

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Sequence Acquisition and Physicochemical Characterisation of PgUGT Proteins
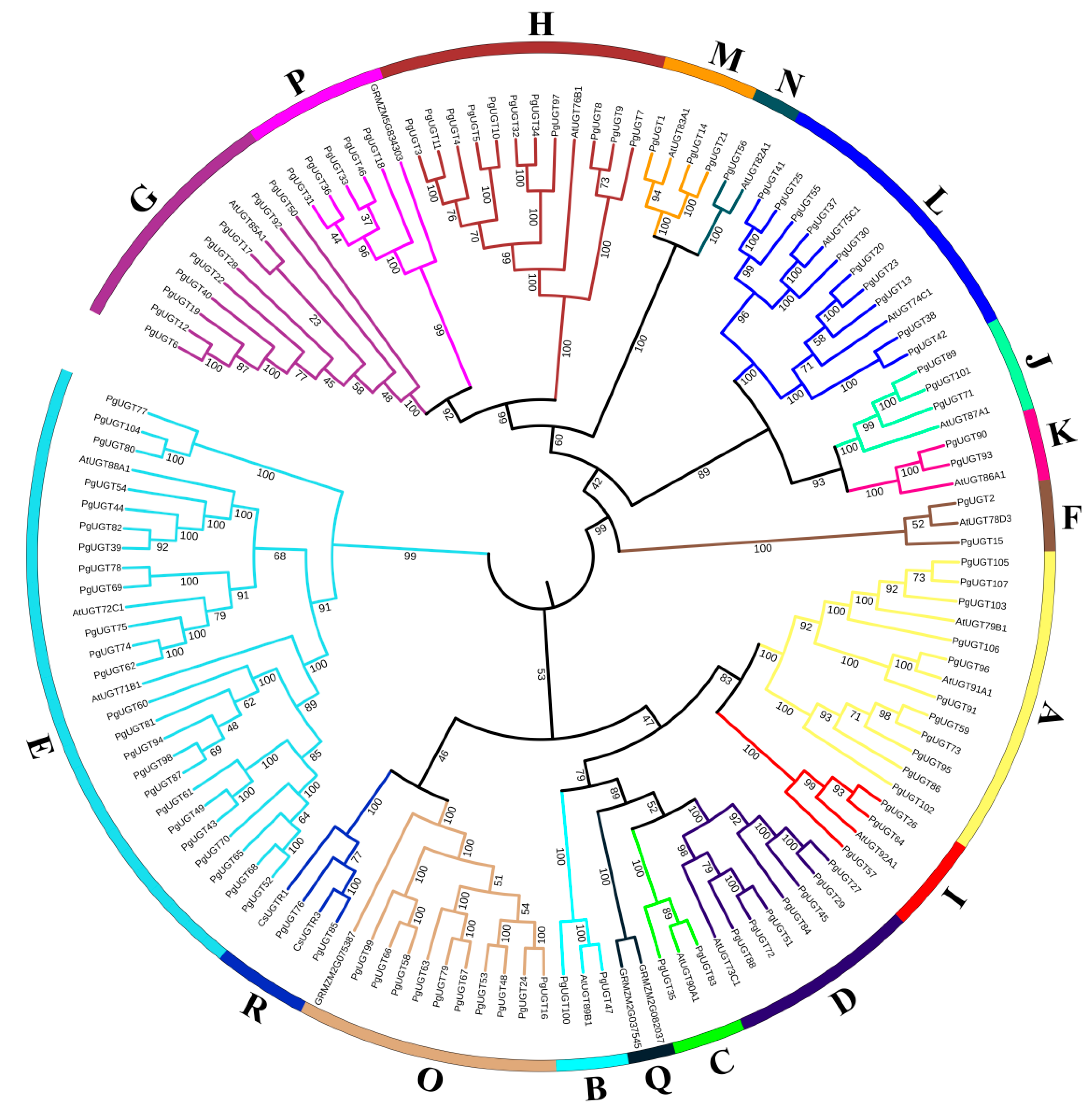
2.2. Phylogenetic Tree Analysis of PgUGT Proteins
2.3. Chromosomal Localisation and Collinearity Analyses of PgUGTs
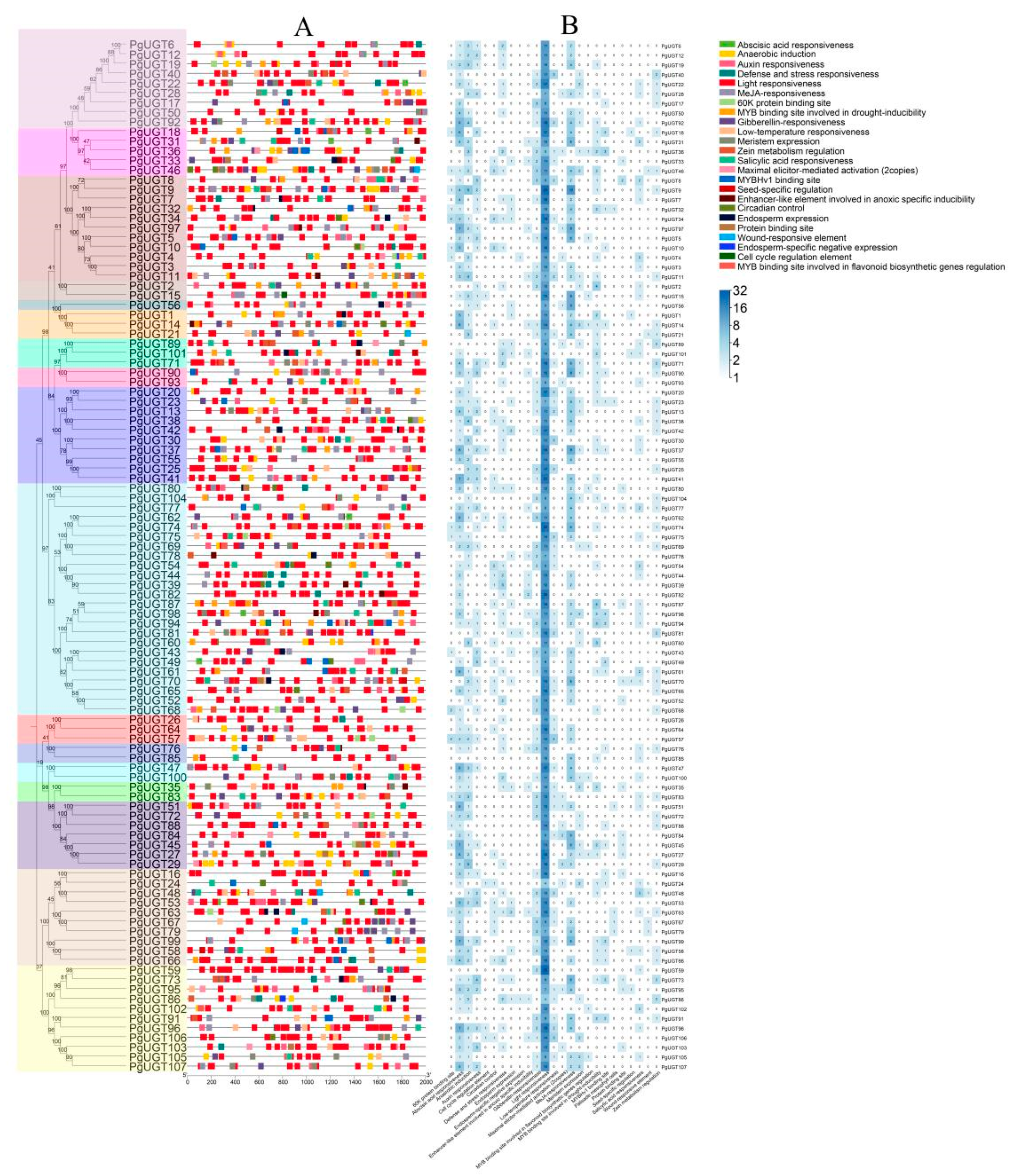
2.4. Cis-Regulatory Elements in PgUGT Promoters
2.5. Conserved Motifs, Domains, and Gene Structures of PgUGTs
2.6. Screening of Candidate PgUGTs
2.7. Protein Expression and Purification
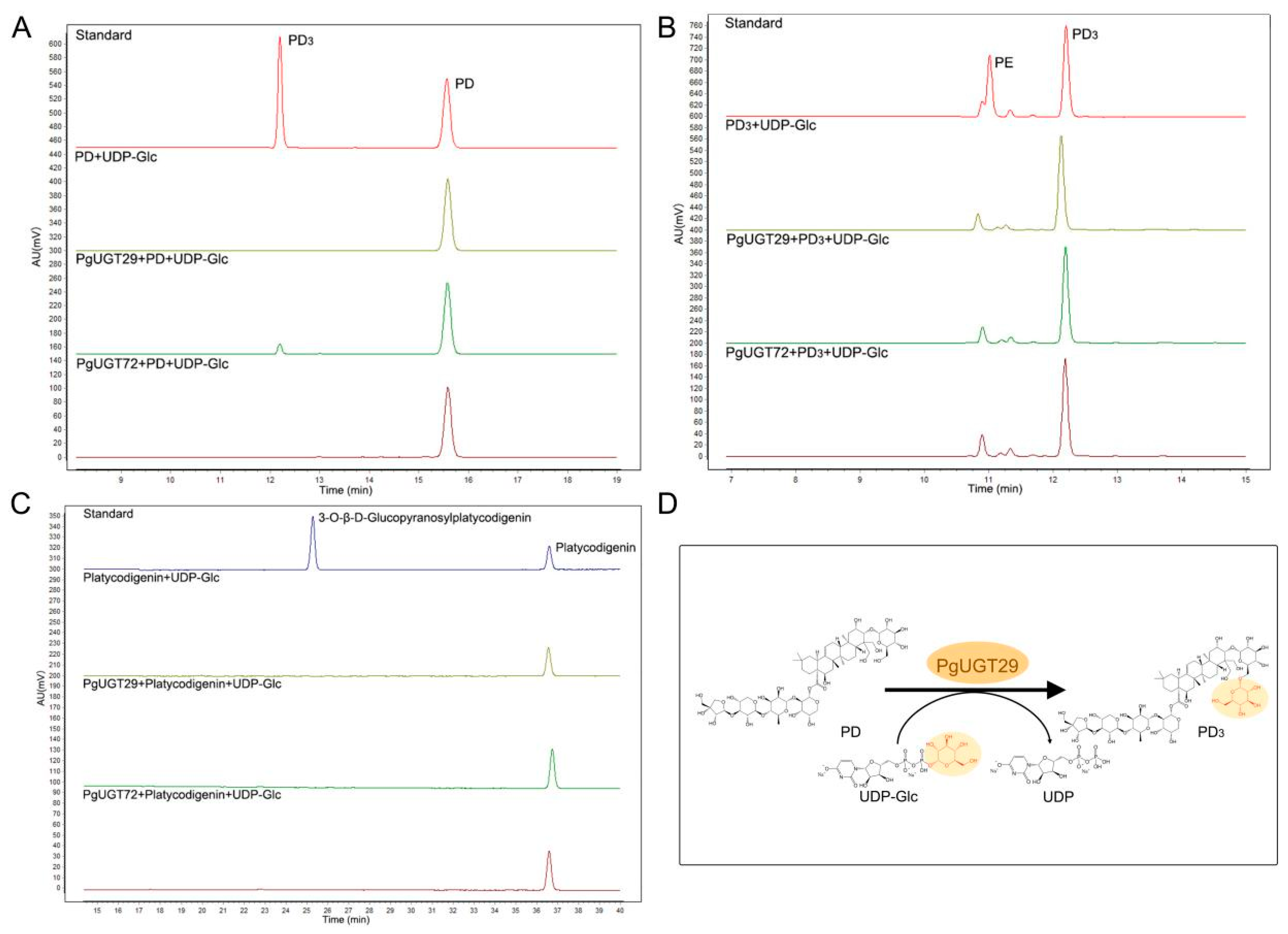
2.8. Functional Validation and Molecular Docking
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Genome-Wide Identification of PgUGTs and Construction of Phylogenetic Tree
4.3. Chromosomal Localisation and Collinearity Analysis
4.4. Cis-Regulatory Elements Analysis
4.5. Motif, Domain, and Gene Structure Analysis
4.6. Total RNA Extraction, cDNA Synthesis, and RT-qPCR
4.7. Heterologous Expression and Purification of PgUGTs in Escherichia coli BL21 (DE3)
4.8. SDS-PAGE and Western Blot Analyses
4.9. Enzymatic Assays of UGTs In Vitro
4.10. Molecular Docking
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, S.; Chai, X.; Hou, G.; Zhao, F.; Meng, Q. Platycodon grandiflorum (Jacq.) A. DC.: A review of phytochemistry, pharmacology, toxicology and traditional use. Phytomedicine 2022, 106, 154422. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Han, Y.; Kim, Y.A.; Jin, S.W.; Lee, G.H.; Jeong, H.M.; Lee, H.S.; Chung, Y.C.; Lee, Y.C.; Kim, E.J.; et al. Platycodin D Inhibits Osteoclastogenesis by Repressing the NFATc1 and MAPK Signaling Pathway. J. Cell. Biochem. 2017, 118, 860–868. [Google Scholar] [CrossRef]
- Huang, W.; Zhou, H.; Yuan, M.; Lan, L.; Hou, A.; Ji, S. Comprehensive characterization of the chemical constituents in Platycodon grandiflorum by an integrated liquid chromatography-mass spectrometry strategy. J. Chromatogr. A 2021, 1654, 462477. [Google Scholar] [CrossRef]
- Choi, Y.H.; Kim, Y.S.; Yeo, S.J.; Roh, S.H.; Jeong, Y.C.; Kang, J.S.; Ryu, S.Y. Ameliorating effect of balloon flower saponin on the ethanol-induced memory impairment in mice. Phytother. Res. 2008, 22, 973–976. [Google Scholar] [CrossRef]
- Yoo, D.S.; Choi, Y.H.; Cha, M.R.; Lee, B.H.; Kim, S.J.; Yon, G.H.; Hong, K.S.; Jang, Y.S.; Lee, H.S.; Kim, Y.S.; et al. HPLC-ELSD analysis of 18 platycosides from balloon flower roots (Platycodi Radix) sourced from various regions in Korea and geographical clustering of the cultivation areas. Food Chem. 2011, 129, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Vincken, J.P.; Heng, L.; de Groot, A.; Gruppen, H. Saponins, classification and occurrence in the plant kingdom. Phytochemistry 2007, 68, 275–297. [Google Scholar] [CrossRef] [PubMed]
- Kalinowska, M.; Zimowski, J.; Pączkowski, C.; Wojciechowski, Z.A. The Formation of Sugar Chains in Triterpenoid Saponins and Glycoalkaloids. Phytochem. Rev. 2005, 4, 237–257. [Google Scholar] [CrossRef]
- Paquette, S.; Moller, B.L.; Bak, S. On the origin of family 1 plant glycosyltransferases. Phytochemistry 2003, 62, 399–413. [Google Scholar] [CrossRef]
- Song, C.; Hartl, K.; McGraphery, K.; Hoffmann, T.; Schwab, W. Attractive but Toxic: Emerging Roles of Glycosidically Bound Volatiles and Glycosyltransferases Involved in Their Formation. Mol. Plant 2018, 11, 1225–1236. [Google Scholar] [CrossRef]
- Speeckaert, N.; Adamou, N.M.; Hassane, H.A.; Baldacci-Cresp, F.; Mol, A.; Goeminne, G.; Boerjan, W.; Duez, P.; Hawkins, S.; Neutelings, G.; et al. Characterization of the UDP-glycosyltransferase UGT72 Family in Poplar and Identification of Genes Involved in the Glycosylation of Monolignols. Int. J. Mol. Sci. 2020, 21, 5018. [Google Scholar] [CrossRef]
- Meesapyodsuk, D.; Balsevich, J.; Reed, D.W.; Covello, P.S. Saponin biosynthesis in Saponaria vaccaria. cDNAs encoding beta-amyrin synthase and a triterpene carboxylic acid glucosyltransferase. Plant Physiol. 2007, 143, 959–969. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.Y.; Chen, G.; Song, W.L.; Fan, W.; Wei, K.H.; He, S.M.; Zhang, G.H.; Tang, J.R.; Li, Y.; Lin, Y.; et al. Transcriptome analysis of Panax zingiberensis identifies genes encoding oleanolic acid glucuronosyltransferase involved in the biosynthesis of oleanane-type ginsenosides. Planta 2019, 249, 393–406. [Google Scholar] [CrossRef]
- Zhang, H.; Hua, X.; Zheng, D.; Wu, H.; Li, C.; Rao, P.; Wen, M.; Choi, Y.E.; Xue, Z.; Wang, Y.; et al. De Novo Biosynthesis of Oleanane-Type Ginsenosides in Saccharomyces cerevisiae Using Two Types of Glycosyltransferases from Panax ginseng. J. Agric. Food Chem. 2022, 70, 2231–2240. [Google Scholar] [CrossRef]
- Tang, J.R.; Chen, G.; Lu, Y.C.; Tang, Q.Y.; Song, W.L.; Lin, Y.; Li, Y.; Peng, S.F.; Yang, S.C.; Zhang, G.H.; et al. Identification of two UDP-glycosyltransferases involved in the main oleanane-type ginsenosides in Panax japonicus var. major. Planta 2021, 253, 91. [Google Scholar] [CrossRef]
- Su, X.; Meng, F.; Liu, Y.; Jiang, W.; Wang, Z.; Wu, L.; Guo, X.; Yao, X.; Wu, J.; Sun, Z.; et al. Molecular Cloning and Functional Characterization of a beta-Glucosidase Gene to Produce Platycodin D in Platycodon grandiflorus. Front. Plant Sci. 2022, 13, 955628. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Shi, J.J.; Liu, W.; Lu, X.; Li, B. MALDI Imaging Assisted Discovery of a Di-O-glycosyltransferase from Platycodon grandiflorum Root. Angew. Chem. Int. Ed. Engl. 2023, 62, e202301309. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Liu, Y.; Han, L.; Wang, Z.; Cao, M.; Wu, L.; Jiang, W.; Meng, F.; Guo, X.; Yu, N.; et al. A candidate gene identified in converting platycoside E to platycodin D from Platycodon grandiflorus by transcriptome and main metabolites analysis. Sci. Rep. 2021, 11, 9810. [Google Scholar] [CrossRef]
- Rahimi, S.; Kim, J.; Mijakovic, I.; Jung, K.H.; Choi, G.; Kim, S.C.; Kim, Y.J. Triterpenoid-biosynthetic UDP-glycosyltransferases from plants. Biotechnol. Adv. 2019, 37, 107394. [Google Scholar] [CrossRef]
- Singh, S.; Apoorva Saha, P.; Rai, N.; Kumari, S.; Pandey-Rai, S. Unravelling triterpenoid biosynthesis in plants for applications in bioengineering and large-scale sustainable production. Ind. Crops Prod. 2023, 199, 116789. [Google Scholar] [CrossRef]
- Bowles, D.; Lim, E.K.; Poppenberger, B.; Vaistij, F.E. Glycosyltransferases of lipophilic small molecules. Annu. Rev. Plant Biol. 2006, 57, 567–597. [Google Scholar] [CrossRef]
- Xu, J.; Chu, Y.; Liao, B.; Xiao, S.; Yin, Q.; Bai, R.; Su, H.; Dong, L.; Li, X.; Qian, J.; et al. Panax ginseng genome examination for ginsenoside biosynthesis. Gigascience 2017, 6, gix093. [Google Scholar] [CrossRef] [PubMed]
- Khorolragchaa, A.; Kim, Y.J.; Rahimi, S.; Sukweenadhi, J.; Jang, M.G.; Yang, D.C. Grouping and characterization of putative glycosyltransferase genes from Panax ginseng Meyer. Gene 2014, 536, 186–192. [Google Scholar] [CrossRef]
- Augustin, J.M.; Fau, D.S.; Shinoda, T.; Fau, S.T.; Sanmiya, K.; Fau, S.K.; Nielsen, J.K.; Fau, N.J.; Khakimov, B.; Fau, K.B.; et al. UDP-glycosyltransferases from the UGT73C subfamily in Barbarea vulgaris catalyze sapogenin 3-O-glucosylation in saponin-mediated insect resistance. Plant Physiol. 2012, 160, 1881–1895. [Google Scholar] [CrossRef]
- Erthmann, P.Ø.; Agerbirk, N.; Bak, S. A tandem array of UDP-glycosyltransferases from the UGT73C subfamily glycosylate sapogenins, forming a spectrum of mono- and bisdesmosidic saponins. Plant Mol. Biol. 2018, 97, 37–55. [Google Scholar] [CrossRef]
- Yano, R.; Takagi, K.; Tochigi, S.; Fujisawa, Y.; Nomura, Y.; Tsuchinaga, H.; Takahashi, Y.; Takada, Y.; Kaga, A.; Anai, T.; et al. Isolation and Characterization of the Soybean Sg-3 Gene that is Involved in Genetic Variation in Sugar Chain Composition at the C-3 Position in Soyasaponins. Plant Cell Physiol. 2018, 59, 792–805. [Google Scholar] [CrossRef]
- Shibuya, M.; Nishimura, K.; Yasuyama, N.; Ebizuka, Y. Identification and characterization of glycosyltransferases involved in the biosynthesis of soyasaponin I in Glycine max. FEBS Lett. 2010, 584, 2258–2264. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.S.; Li, C.J.; Wei, W.; Wei, Y.J.; Liu, Q.F.; Zhao, G.P.; Yue, J.M.; Yan, X.; Wang, P.P.; Zhou, Z.H. The unprecedented diversity of UGT94-family UDP-glycosyltransferases in Panax plants and their contribution to ginsenoside biosynthesis. Sci. Rep. 2020, 10, 15394. [Google Scholar] [CrossRef] [PubMed]
- Takagi, K.; Yano, R.; Tochigi, S.; Fujisawa, Y.; Tsuchinaga, H.; Takahashi, Y.; Takada, Y.; Kaga, A.; Anai, T.; Tsukamoto, C.; et al. Genetic and functional characterization of Sg-4 glycosyltransferase involved in the formation of sugar chain structure at the C-3 position of soybean saponins. Phytochemistry 2018, 156, 96–105. [Google Scholar] [CrossRef]
- Barvkar, V.T.; Pardeshi, V.C.; Kale, S.M.; Kadoo, N.Y.; Gupta, V.S. Phylogenomic analysis of UDP glycosyltransferase 1 multigene family in Linum usitatissimum identified genes with varied expression patterns. BMC Genom. 2012, 13, 175. [Google Scholar] [CrossRef]
- Li, Y.; Li, P.; Wang, Y.; Dong, R.; Yu, H.; Hou, B. Genome-wide identification and phylogenetic analysis of Family-1 UDP glycosyltransferases in maize (Zea mays). Planta 2014, 239, 1265–1279. [Google Scholar] [CrossRef]
- Cui, L.; Yao, S.; Dai, X.; Yin, Q.; Liu, Y.; Jiang, X.; Wu, Y.; Qian, Y.; Pang, Y.; Gao, L.; et al. Identification of UDP-glycosyltransferases involved in the biosynthesis of astringent taste compounds in tea (Camellia sinensis). J. Exp. Bot. 2016, 67, 2285–2297. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Gu, J.; Luo, Y.; Wang, Y.; Pang, Y.; Shen, G.; Guo, B. Genome-wide analysis of UGT gene family identified key gene for the biosynthesis of bioactive flavonol glycosides in Epimedium pubescens Maxim. Synth. Syst. Biotechnol. 2022, 7, 1095–1107. [Google Scholar] [CrossRef] [PubMed]
- Li, G.X.; Li, J.Y.; Qin, G.H.; Liu, C.Y.; Liu, X.; Cao, Z.; Jia, B.T.; Zhang, H.P. Characterization and Expression Analysis of the UDP Glycosyltransferase Family in Pomegranate (Punica granatum L.). Horticulturae 2023, 9, 119. [Google Scholar] [CrossRef]
- Li, L.; Modolo, L.V.; Escamilla-Trevino, L.L.; Achnine, L.; Dixon, R.A.; Wang, X. Crystal structure of Medicago truncatula UGT85H2—Insights into the structural basis of a multifunctional (iso) flavonoid glycosyltransferase. J. Mol. Biol. 2007, 370, 951–963. [Google Scholar] [CrossRef] [PubMed]
- Radominska-Pandya, A.; Bratton, S.M.; Redinbo, M.R.; Miley, M.J. The crystal structure of human UDP-glucuronosyltransferase 2B7 C-terminal end is the first mammalian UGT target to be revealed: The significance for human UGTs from both the 1A and 2B families. Drug Metab. Rev. 2010, 42, 133–144. [Google Scholar] [CrossRef]
- Dai, L.H.; Qin, L.J.; Hu, Y.M.; Huang, J.W.; Hu, Z.Y.; Min, J.; Sun, Y.X.; Guo, R.T. Structural dissection of unnatural ginsenoside-biosynthetic UDP-glycosyltransferase Bs-YjiC from Bacillus subtilis for substrate promiscuity. Biochem. Biophys. Res. Commun. 2021, 534, 73–78. [Google Scholar] [CrossRef]
- Chang, A.; Singh, S.; Phillips, G.N.; Thorson, J.S. Glycosyltransferase structural biology and its role in the design of catalysts for glycosylation. Curr. Opin. Biotech. 2011, 22, 800–808. [Google Scholar] [CrossRef]
- Jia, Y.; Chen, S.; Chen, W.; Zhang, P.; Su, Z.; Zhang, L.; Xu, M.; Guo, L. A Chromosome-Level Reference Genome of Chinese Balloon Flower (Platycodon grandiflorus). Front. Genet. 2022, 13, 869784. [Google Scholar] [CrossRef]
- Alzohairy, A. BioEdit: An important software for molecular biology. GERF Bull. Biosci. 2011, 2, 60–61. [Google Scholar]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef]
- Savojardo, C.; Martelli, P.L.; Fariselli, P.; Profiti, G.; Casadio, R. BUSCA: An integrative web server to predict subcellular localization of proteins. Nucleic Acids Res. 2018, 46, W459–W466. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed]
- Chao, J.; Li, Z.; Sun, Y.; Aluko, O.O.; Wu, X.; Wang, Q.; Liu, G. MG2C: A user-friendly online tool for drawing genetic maps. Mol. Hortic 2021, 1, 16. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Nystrom, S.L.; McKay, D.J. Memes: A motif analysis environment in R using tools from the MEME Suite. PLoS Comput. Biol. 2021, 17, e1008991. [Google Scholar] [CrossRef]
- Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Gonzales, N.R.; Gwadz, M.; Lu, S.; Marchler, G.H.; Song, J.S.; Thanki, N.; Yamashita, R.A.; et al. The conserved domain database in 2023. Nucleic Acids Res. 2023, 51, D384–D388. [Google Scholar] [CrossRef]
- Yang, J.; Anishchenko, I.; Park, H.; Peng, Z.; Ovchinnikov, S.; Baker, D. Improved protein structure prediction using predicted interresidue orientations. Proc. Natl. Acad. Sci. USA 2020, 117, 1496–1503. [Google Scholar] [CrossRef]
- Morris, G.M.; Goodsell, D.S.; Halliday, R.S.; Huey, R.; Hart, W.E.; Belew, R.K.; Olson, A.J. Automated docking using a Lamarckian genetic algorithm and an empirical binding free energy function. J. Comput. Chem. 1998, 19, 1639–1662. [Google Scholar] [CrossRef]
- DeLano, W.L. PyMOL: An Open-Source Molecular Graphics Tool. CCP4 Newsl. Protein Crystallogr. 2002, 40, 82–92. [Google Scholar]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, J.; Rao, W.; Peng, D.; Wei, T.; Xing, S. Genome-Wide Identification of UGT Gene Family and Functional Analysis of PgUGT29 in Platycodon grandiflorus. Int. J. Mol. Sci. 2025, 26, 4832. https://doi.org/10.3390/ijms26104832
Fan J, Rao W, Peng D, Wei T, Xing S. Genome-Wide Identification of UGT Gene Family and Functional Analysis of PgUGT29 in Platycodon grandiflorus. International Journal of Molecular Sciences. 2025; 26(10):4832. https://doi.org/10.3390/ijms26104832
Chicago/Turabian StyleFan, Jizhou, Weiyi Rao, Daiyin Peng, Tao Wei, and Shihai Xing. 2025. "Genome-Wide Identification of UGT Gene Family and Functional Analysis of PgUGT29 in Platycodon grandiflorus" International Journal of Molecular Sciences 26, no. 10: 4832. https://doi.org/10.3390/ijms26104832
APA StyleFan, J., Rao, W., Peng, D., Wei, T., & Xing, S. (2025). Genome-Wide Identification of UGT Gene Family and Functional Analysis of PgUGT29 in Platycodon grandiflorus. International Journal of Molecular Sciences, 26(10), 4832. https://doi.org/10.3390/ijms26104832