
Phytochelatins and Cadmium Mitigation: Harnessing Genetic Avenues for Plant Functional Manipulation


Abstract
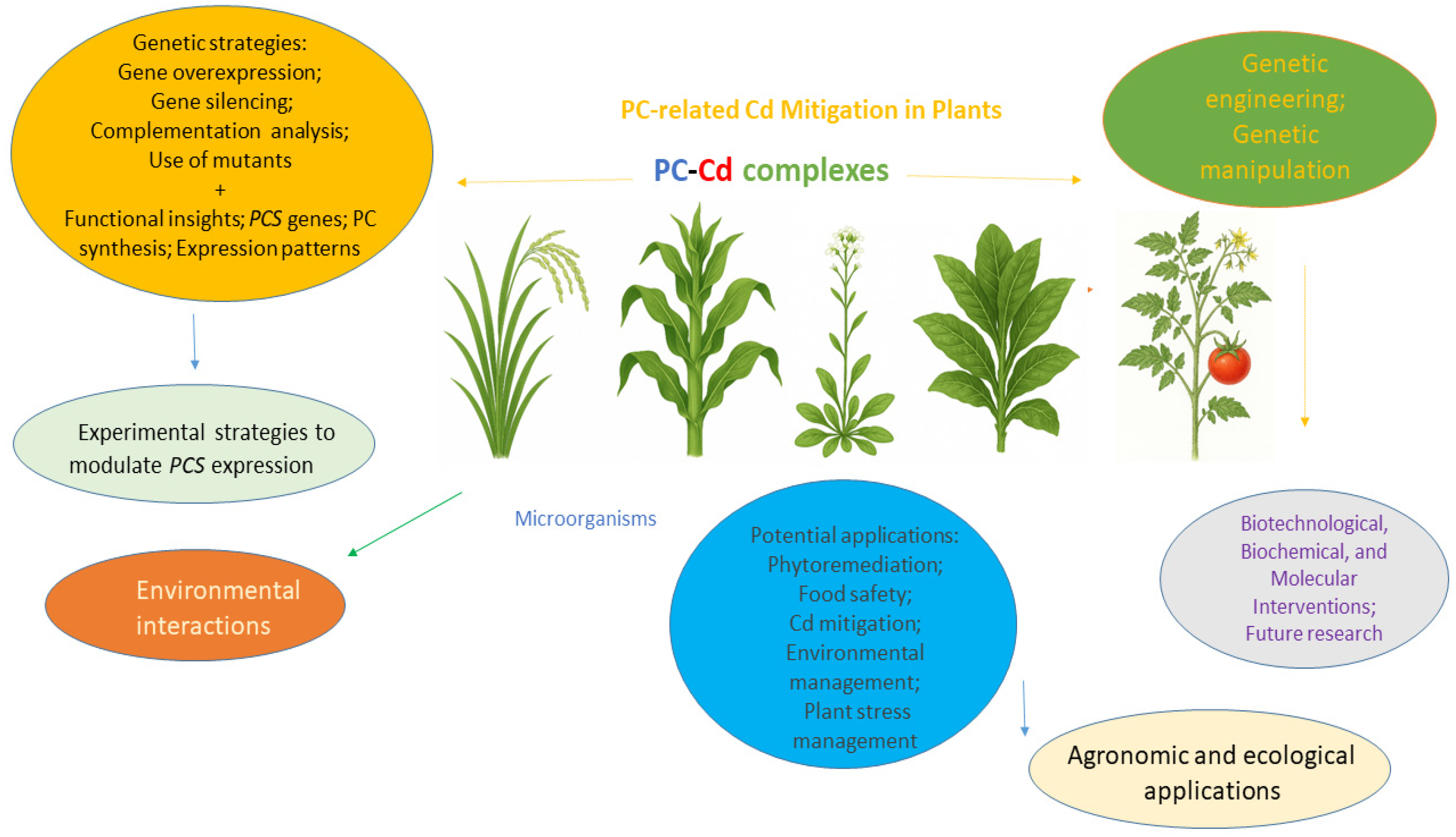
1. Cadmium and Phytochelatins as Strategic Targets
2. Advances in Phytochelatin Synthase Research and Plant Genetic Manipulation
3. Research Avenues and Approaches in PC-Related Genetic Manipulation
4. Concluding Remarks and Additional Future Directions
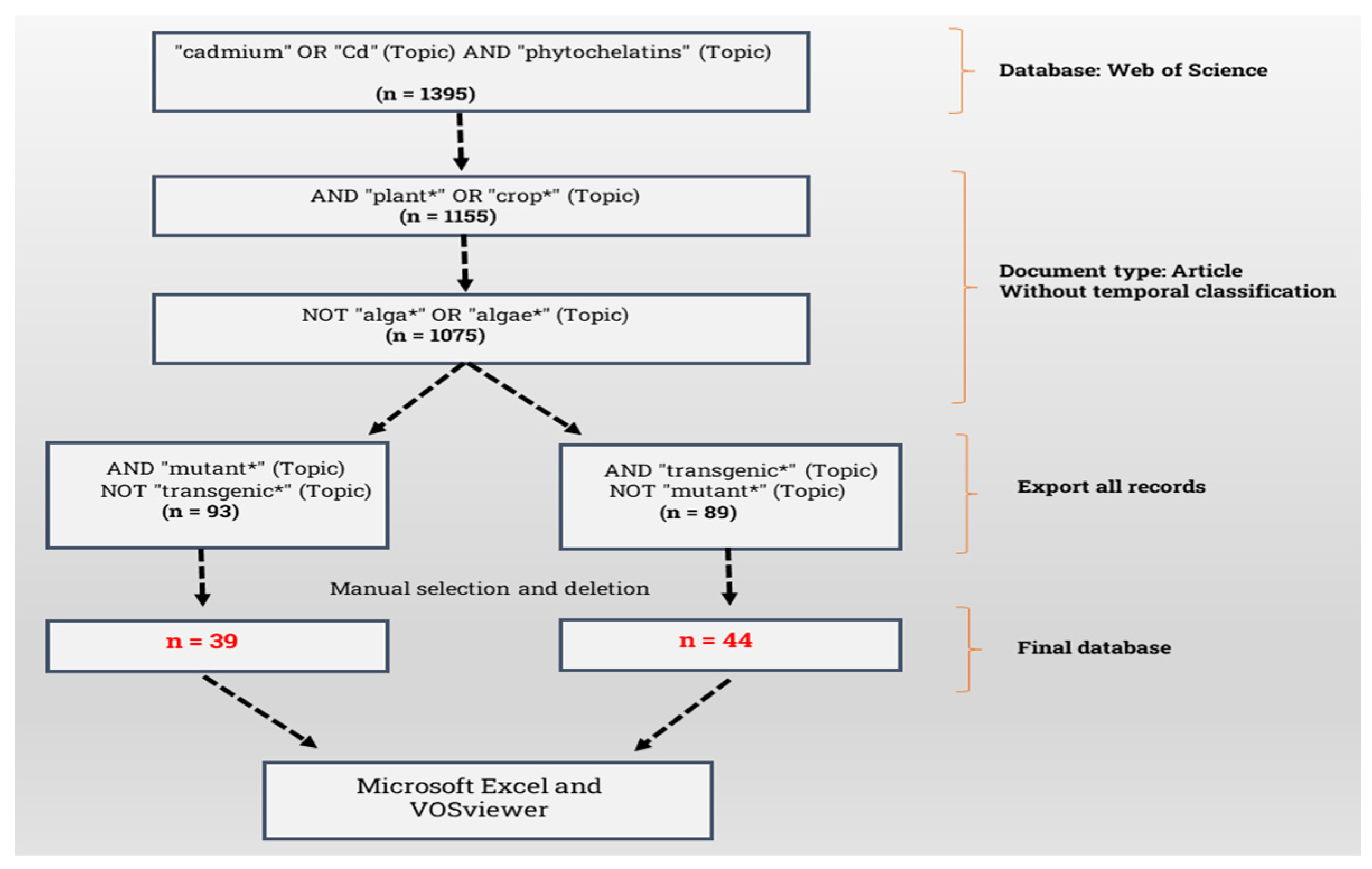
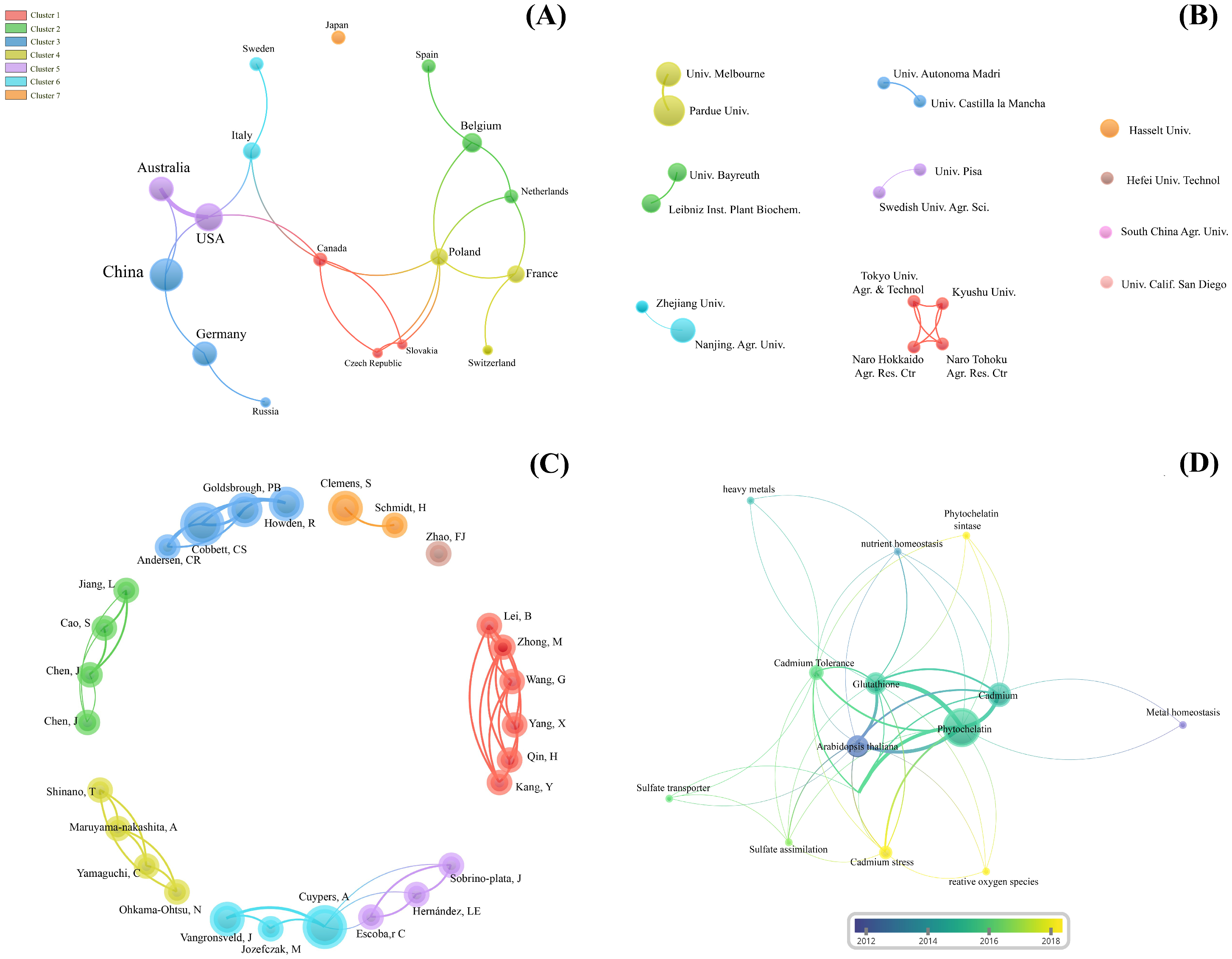
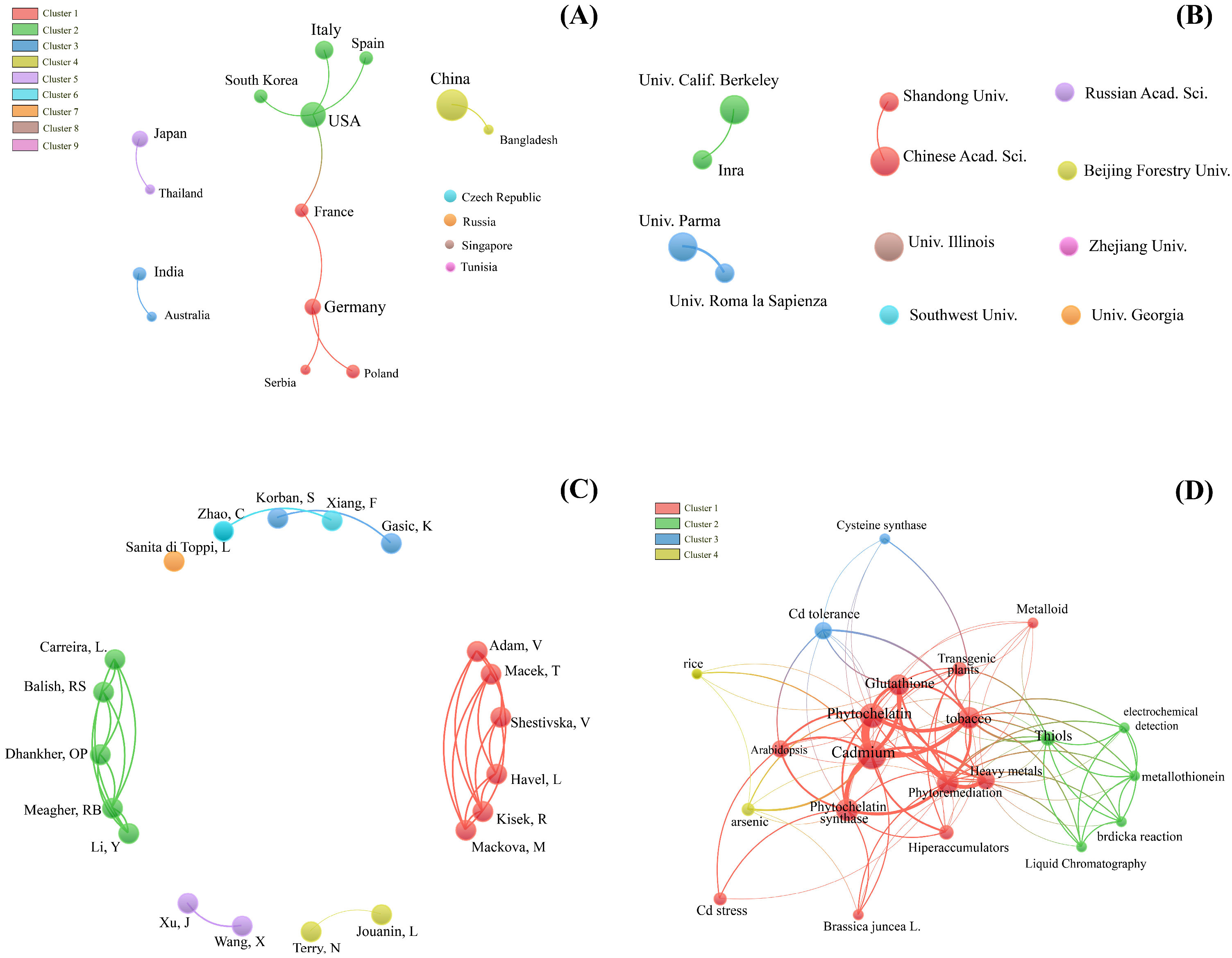
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Marques, D.N.; Mason, C.; Stolze, S.C.; Harzen, A.; Nakagami, H.; Skirycz, A.; Piotto, F.A.; Azevedo, R.A. Grafting systems for plant cadmium research: Insights for basic plant physiology and applied mitigation. Sci. Total Environ. 2023, 892, 164610. [Google Scholar] [CrossRef] [PubMed]
- Marques, D.N.; Nogueira, M.L.; Gaziola, S.A.; Batagin-Piotto, K.D.; Freitas, N.C.; Alcantara, B.K.; Paiva, L.V.; Mason, C.; Piotto, F.A.; Azevedo, R.A. New insights into cadmium tolerance and accumulation in tomato: Dissecting root and shoot responses using cross-genotype grafting. Environ. Res. 2023, 216, 114577. [Google Scholar] [CrossRef]
- Seregin, I.V.; Kozhevnikova, A.D. Phytochelatins: Sulfur-Containing Metal(loid)-Chelating Ligands in Plants. Int. J. Mol. Sci. 2023, 24, 2430. [Google Scholar] [CrossRef]
- Faizan, M.; Alam, P.; Hussain, A.; Karabulut, F.; Haque Tonny, S.; Cheng, S.H.; Yusuf, M.; Adil, M.F.; Sehar, S.; Alomrani, S.O.; et al. Phytochelatins: Key Regulator Against Heavy Metal Toxicity in Plants. Plant Stress 2024, 11, 10035. [Google Scholar] [CrossRef]
- Marques, D.N.; Gaziola, S.A.; Piotto, F.A.; Azevedo, R.A. Phytochelatins: Advances in Tomato Research. Agronomy 2025, 15, 80. [Google Scholar] [CrossRef]
- Howden, R.; Cobbett, C.S. Cadmium-sensitive mutants of Arabidopsis thaliana. Plant Physiol. 1992, 100, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Howden, R.; Goldsbrough, P.B.; Andersen, C.R.; Cobbett, C.S. Cadmium-Sensitive, cad1 Mutants of Arabidopsis thaliana Are Phytochelatin Deficient. Plant Physiol. 1995, 107, 1059–1066. [Google Scholar] [CrossRef]
- Ha, S.-B.; Smith, A.P.; Howden, R.; Dietrich, W.M.; Bugg, S.; O’Connell, M.J.; Goldsbrough, P.B.; Cobbett, C.S. Phytochelatin Synthase Genes from Arabidopsis and the Yeast Schizosaccharomyces pombe. Plant Cell 1999, 11, 1153–1163. [Google Scholar] [CrossRef]
- Cazalé, A.C.; Clemens, S. Arabidopsis thaliana expresses a second functional phytochelatin synthase. FEBS Lett. 2001, 507, 215–219. [Google Scholar] [CrossRef]
- Larsson, E.H.; Asp, H.; Bornman, J.F. Influence of prior Cd2+ exposure on the uptake of Cd2+ and other elements in the phytochelatin-deficient mutant, cad1-3, of Arabidopsis thaliana. J. Exp. Bot. 2002, 53, 447–453. [Google Scholar] [CrossRef]
- Lee, S.; Moon, J.S.; Domier, L.L.; Korban, S.S. Molecular characterization of phytochelatin synthase expression in transgenic Arabidopsis. Plant Physiol. Biochem. 2002, 40, 727–733. [Google Scholar] [CrossRef]
- Gisbert, C.; Ros, R.; De Haro, A.; Walker, D.J.; Bernal, M.P.; Serrano, R.; Navarro-Aviñó, J. A plant genetically modified that accumulates Pb is especially promising for phytoremediation. Biochem. Biophys. Res. Commun. 2003, 303, 440–445. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.-M.; Lee, D.A.; Schroeder, J.I. Long-distance root-to-shoot transport of phytochelatins and cadmium in Arabidopsis. Proc. Natl. Acad. Sci. USA 2003, 100, 10118–10123. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Petros, D.; Moon, J.S.; Ko, T.S.; Goldsbrough, P.B.; Korban, S.S. Higher levels of ectopic expression of Arabidopsis phytochelatin synthase do not lead to increased cadmium tolerance and accumulation. Plant Physiol. Biochem. 2003, 41, 903–910. [Google Scholar] [CrossRef]
- Lee, S.; Moon, J.S.; Ko, T.S.; Petros, D.; Goldsbrough, P.B.; Korban, S.S. Overexpression of Arabidopsis phytochelatin synthase paradoxically leads to hypersensitivity to cadmium stress. Plant Physiol. 2003, 131, 656–663. [Google Scholar] [CrossRef]
- Li, Y.; Dhankher, O.P.; Carreira, L.; Lee, D.; Chen, A.; Schroeder, J.I.; Balish, R.S.; Meagher, R.B. Overexpression of Phytochelatin Synthase in Arabidopsis Leads to Enhanced Arsenic Tolerance and Cadmium Hypersensitivity. Plant Cell Physiol. 2004, 45, 1787–1797. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kang, B.S. Expression of Arabidopsis phytochelatin synthase 2 is too low to complement an AtPCS1-defective Cad1-3 mutant. Mol. Cells 2005, 19, 81–87. [Google Scholar] [CrossRef]
- Bovet, L.; Eggmann, T.; Meylan-Bettex, M.; Polier, J.; Kammer, P.; Marin, E.; Feller, U.; Martinoia, E. Transcript levels of AtMRPs after cadmium treatment: Induction of AtMRP3. Plant Cell Environ. 2003, 26, 371–381. [Google Scholar] [CrossRef]
- Lee, S.; Kang, B.S. Phytochelatin Is Not a Primary Factor in Determining Copper Tolerance. J. Plant Biol. 2005, 48, 32–38. [Google Scholar] [CrossRef]
- Wong, C.K.E.; Cobbett, C.S. HMA P-type ATPases are the major mechanism for root-to-shoot Cd translocation in Arabidopsis thaliana. New Phytol. 2009, 181, 71–78. [Google Scholar] [CrossRef]
- Guo, W.J.; Meetam, M.; Goldsbrough, P.B. Examining the specific contributions of individual Arabidopsis metallothioneins to copper distribution and metal tolerance. Plant Physiol. 2008, 146, 1697–1706. [Google Scholar] [CrossRef] [PubMed]
- Tennstedt, P.; Peisker, D.; Böttcher, C.; Trampczynska, A.; Clemens, S. Phytochelatin synthesis is essential for the detoxification of excess zinc and contributes significantly to the accumulation of zinc. Plant Physiol. 2009, 149, 938–948. [Google Scholar] [CrossRef]
- Park, J.; Song, W.Y.; Ko, D.; Eom, Y.; Hansen, T.H.; Schiller, M.; Lee, T.G.; Martinoia, E.; Lee, Y. The phytochelatin transporters AtABCC1 and AtABCC2 mediate tolerance to cadmium and mercury. Plant J. 2012, 69, 278–288. [Google Scholar] [CrossRef] [PubMed]
- Molins, H.; Michelet, L.; Lanquar, V.; Agorio, A.; Giraudat, J.; Roach, T.; Krieger-Liszkay, A.; Thomine, S. Mutants impaired in vacuolar metal mobilization identify chloroplasts as a target for cadmium hypersensitivity in Arabidopsis thaliana. Plant Cell Environ. 2013, 36, 804–817. [Google Scholar] [CrossRef] [PubMed]
- Nahar, N.; Rahman, A.; Moś, M.; Warzecha, T.; Ghosh, S.; Hossain, K.; Nawani, N.N.; Mandal, A. In silico and in vivo studies of molecular structures and mechanisms of AtPCS1 protein involved in binding arsenite and/or cadmium in plant cells. J. Mol. Model. 2014, 20, 2104. [Google Scholar] [CrossRef]
- Sobrino-Plata, J.; Carrasco-Gil, S.; Abadía, J.; Escobar, C.; Álvarez-Fernández, A.; Hernández, L.E. The role of glutathione in mercury tolerance resembles its function under cadmium stress in Arabidopsis. Metallomics 2014, 6, 356–366. [Google Scholar] [CrossRef]
- Brunetti, P.; Zanella, L.; De Paolis, A.; Di Litta, D.; Cecchetti, V.; Falasca, G.; Barbieri, M.; Altamura, M.M.; Costantino, P.; Cardarelli, M. Cadmium-inducible expression of the ABC-type transporter AtABCC3 increases phytochelatin-mediated cadmium tolerance in Arabidopsis. J. Exp. Bot. 2015, 66, 3815–3829. [Google Scholar] [CrossRef]
- Cahoon, R.E.; Lutke, W.K.; Cameron, J.C.; Chen, S.; Lee, S.G.; Rivard, R.S.; Rea, P.A.; Jez, J.M. Adaptive Engineering of Phytochelatin-based Heavy Metal Tolerance. J. Biol. Chem. 2015, 290, 17321–17330. [Google Scholar] [CrossRef]
- Kühnlenz, T.; Hofmann, C.; Uraguchi, S.; Schmidt, H.; Schempp, S.; Weber, M.; Lahner, B.; Salt, D.E.; Clemens, S. Phytochelatin Synthesis Promotes Leaf Zn Accumulation of Arabidopsis thaliana Plants Grown in Soil with Adequate Zn Supply and is Essential for Survival on Zn-Contaminated Soil. Plant Cell Physiol. 2016, 57, 2342–2352. [Google Scholar] [CrossRef]
- Gielen, H.; Vangronsveld, J.; Cuypers, A. Cd-induced Cu deficiency responses in Arabidopsis thaliana: Are phytochelatins involved? Plant Cell Environ. 2017, 40, 390–400. [Google Scholar] [CrossRef]
- De Benedictis, M.; Brunetti, C.; Brauer, E.K.; Andreucci, A.; Popescu, S.C.; Commisso, M.; Guzzo, F.; Sofo, A.; Ruffini Castiglione, M.; Vatamaniuk, O.K.; et al. The Arabidopsis thaliana Knockout Mutant for Phytochelatin Synthase1 (cad1-3) Is Defective in Callose Deposition, Bacterial Pathogen Defense and Auxin Content, But Shows an Increased Stem Lignification. Front. Plant Sci. 2018, 9, 19. [Google Scholar] [CrossRef]
- Chen, J.; Wang, X.; Zhang, W.; Zhang, S.; Zhao, F.J. Protein phosphatase 2A alleviates cadmium toxicity by modulating ethylene production in Arabidopsis thaliana. Plant Cell Environ. 2020, 43, 1008–1022. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Huang, S.B.; Wang, X.; Huang, L.; Gao, C.; Huang, X.Y.; Zhao, F.J. IAR4 mutation enhances cadmium toxicity by disturbing auxin homeostasis in Arabidopsis thaliana. J. Exp. Bot. 2024, 75, 438–453. [Google Scholar] [CrossRef] [PubMed]
- Dutilleul, C.; Jourdain, A.; Bourguignon, J.; Hugouvieux, V. The Arabidopsis Putative Selenium-Binding Protein Family: Expression Study and Characterization of SBP1 as a Potential New Player in Cadmium Detoxification Processes. Plant Physiol. 2008, 147, 239–251. [Google Scholar] [CrossRef] [PubMed]
- Picault, N.; Cazalé, A.C.; Beyly, A.; Cuiné, S.; Carrier, P.; Luu, D.T.; Forestier, C.; Peltier, G. Chloroplast targeting of phytochelatin synthase in Arabidopsis: Effects on heavy metal tolerance and accumulation. Biochimie 2006, 88, 1743–1750. [Google Scholar] [CrossRef]
- Pomponi, M.; Censi, V.; Di Girolamo, V.; De Paolis, A.; di Toppi, L.S.; Aromolo, R.; Costantino, P.; Cardarelli, M. Overexpression of Arabidopsis phytochelatin synthase in tobacco plants enhances Cd2+ tolerance and accumulation but not translocation to the shoot. Planta 2006, 223, 180–190. [Google Scholar] [CrossRef]
- Wawrzynski, A.; Kopera, E.; Wawrzynska, A.; Kaminska, J.; Bal, W.; Sirko, A. Effects of simultaneous expression of heterologous genes involved in phytochelatin biosynthesis on thiol content and cadmium accumulation in tobacco plants. J. Exp. Bot. 2006, 57, 2173–2182. [Google Scholar] [CrossRef]
- Li, J.; Guo, J.; Xu, W.; Ma, M. Enhanced cadmium accumulation in transgenic tobacco expressing the phytochelatin synthase gene of Cynodon dactylon L. J. Integr. Plant Biol. 2006, 48, 928–937. [Google Scholar] [CrossRef]
- Kim, J.H.; Lee, S. Overexpression of Arabidopsis phytochelatin synthase (atpcsi) does not change the maximum capacity for non-protein thiol production induced by Cadmium. J. Plant Biol. 2007, 50, 220–223. [Google Scholar] [CrossRef]
- Wojas, S.; Clemens, S.; Hennig, J.; Sklodowska, A.; Kopera, E.; Schat, H.; Bal, W.; Antosiewicz, D.M. Overexpression of phytochelatin synthase in tobacco: Distinctive effects of AtPCS1 and CePCS genes on plant response to cadmium. J. Exp. Bot. 2008, 59, 2205–2219. [Google Scholar] [CrossRef]
- Guo, J.; Dai, X.; Xu, W.; Ma, M. Overexpressing GSH1 and AsPCS1 simultaneously increases the tolerance and accumulation of cadmium and arsenic in Arabidopsis thaliana. Chemosphere 2008, 72, 1020–1026. [Google Scholar] [CrossRef] [PubMed]
- Wojas, S.; Ruszczyńska, A.; Bulska, E.; Clemens, S.; Antosiewicz, D.M. The role of subcellular distribution of cadmium and phytochelatins in the generation of distinct phenotypes of AtPCS1- and CePCS3-expressing tobacco. J. Plant Physiol. 2010, 167, 981–988. [Google Scholar]
- Brunetti, P.; Zanella, L.; Proia, A.; De Paolis, A.; Falasca, G.; Altamura, M.M.; Sanità di Toppi, L.; Costantino, P.; Cardarelli, M. Cadmium tolerance and phytochelatin content of Arabidopsis seedlings over-expressing the phytochelatin synthase gene AtPCS1. J. Exp. Bot. 2011, 62, 5509–5519. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.Y.; Zhang, Y.X.; Chai, T.Y. Phytochelatin synthase of Thlaspi caerulescens enhanced tolerance and accumulation of heavy metals when expressed in yeast and tobacco. Plant Cell Rep. 2011, 30, 1067–1076. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Xu, W.; Ma, M. The assembly of metals chelation by thiols and vacuolar compartmentalization conferred increased tolerance to and accumulation of cadmium and arsenic in transgenic Arabidopsis thaliana. J. Hazard. Mater. 2012, 199, 309–313. [Google Scholar] [CrossRef]
- Liu, Z.; Gu, C.; Chen, F.; Yang, D.; Wu, K.; Chen, S.; Jiang, J.; Zhang, Z. Heterologous expression of a Nelumbo nucifera phytochelatin synthase gene enhances cadmium tolerance in Arabidopsis thaliana. Appl. Biochem. Biotechnol. 2012, 166, 722–734. [Google Scholar] [CrossRef]
- Shukla, D.; Kesari, R.; Tiwari, M.; Dwivedi, S.; Tripathi, R.D.; Nath, P.; Trivedi, P.K. Expression of Ceratophyllum demersum phytochelatin synthase, CdPCS1, in Escherichia coli and Arabidopsis enhances heavy metal(loid)s accumulation. Protoplasma 2013, 250, 1263–1272. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, Y.; Ding, Y.; Wang, X.; Xu, J. Overexpression of PtPCS enhances cadmium tolerance and cadmium accumulation in tobacco. Plant Cell Tiss. Organ Cult. 2015, 121, 389–396. [Google Scholar] [CrossRef]
- Lee, B.D.; Hwang, S. Tobacco phytochelatin synthase (NtPCS1) plays important roles in cadmium and arsenic tolerance and in early plant development in tobacco. Plant Biotechnol. Rep. 2015, 9, 107–114. [Google Scholar] [CrossRef]
- Zanella, L.; Fattorini, L.; Brunetti, P.; Roccotiello, E.; Cornara, L.; D’Angeli, S.; Della Rovere, F.; Cardarelli, M.; Barbieri, M.; Sanità di Toppi, L.; et al. Overexpression of AtPCS1 in tobacco increases arsenic and arsenic plus cadmium accumulation and detoxification. Planta 2016, 243, 605–622. [Google Scholar] [CrossRef]
- Fan, W.; Guo, Q.; Liu, C.; Liu, X.; Zhang, M.; Long, D.; Xiang, Z.; Zhao, A. Two mulberry phytochelatin synthase genes confer zinc/cadmium tolerance and accumulation in transgenic Arabidopsis and tobacco. Gene 2018, 645, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Stragliati, L.; Bellini, E.; Ricci, A.; Saba, A.; Sanità di Toppi, L.; Varotto, C. Evolution and functional differentiation of recently diverged phytochelatin synthase genes from Arundo donax L. J. Exp. Bot. 2019, 70, 5391–5405. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Zou, T.; Lin, R.; Zheng, J.; Jian, S.; Zhang, M. Characterization of a phytochelatin synthase gene from Ipomoea pes-caprae involved in cadmium tolerance and accumulation in yeast and plants. Plant Physiol. Biochem. 2020, 155, 743–755. [Google Scholar] [CrossRef]
- Zhu, S.; Shi, W.; Jie, Y. Overexpression of BnPCS1, a Novel Phytochelatin Synthase Gene From Ramie (Boehmeria nivea), Enhanced Cd Tolerance, Accumulation, and Translocation in Arabidopsis thaliana. Front. Plant Sci. 2021, 12, 639189. [Google Scholar] [CrossRef]
- Peterson, A.G.; Oliver, D.J. Leaf-targeted phytochelatin synthase in Arabidopsis thaliana. Plant Physiol. Biochem. 2006, 44, 885–892. [Google Scholar] [CrossRef]
- Sadi, B.B.; Vonderheide, A.P.; Gong, J.M.; Schroeder, J.I.; Shann, J.R.; Caruso, J.A. An HPLC-ICP-MS technique for determination of cadmium-phytochelatins in genetically modified Arabidopsis thaliana. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2008, 861, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Shukla, D.; Kesari, R.; Mishra, S.; Dwivedi, S.; Tripathi, R.D.; Nath, P.; Trivedi, P.K. Expression of phytochelatin synthase from aquatic macrophyte Ceratophyllum demersum L. enhances cadmium and arsenic accumulation in tobacco. Plant Cell Rep. 2012, 31, 1687–1699. [Google Scholar] [CrossRef]
- Shukla, D.; Tiwari, M.; Tripathi, R.D.; Nath, P.; Trivedi, P.K. Synthetic phytochelatins complement a phytochelatin-deficient Arabidopsis mutant and enhance the accumulation of heavy metal(loid)s. Biochem. Biophys. Res. Commun. 2013, 434, 664–669. [Google Scholar] [CrossRef]
- Kühnlenz, T.; Schmidt, H.; Uraguchi, S.; Clemens, S. Arabidopsis thaliana phytochelatin synthase 2 is constitutively active in vivo and can rescue the growth defect of the PCS1-deficient cad1-3 mutant on Cd-contaminated soil. J. Exp. Bot. 2014, 65, 4241–4253. [Google Scholar] [CrossRef]
- Kühnlenz, T.; Westphal, L.; Schmidt, H.; Scheel, D.; Clemens, S. Expression of Caenorhabditis elegans PCS in the AtPCS1-deficient Arabidopsis thaliana cad1-3 mutant separates the metal tolerance and non-host resistance functions of phytochelatin synthases. Plant Cell Environ. 2015, 38, 2239–2247. [Google Scholar] [CrossRef]
- Zhang, X.; Rui, H.; Zhang, F.; Hu, Z.; Xia, Y.; Shen, Z. Overexpression of a Functional Vicia sativa PCS1 Homolog Increases Cadmium Tolerance and Phytochelatins Synthesis in Arabidopsis. Front Plant Sci. 2018, 9, 107. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Wang, X.; Wang, R.; Wang, J.; Le, S.; Zhao, Y. Overexpression of Three Duplicated BnPCS Genes Enhanced Cd Accumulation and Translocation in Arabidopsis thaliana Mutant cad1-3. Bull. Environ. Contam. Toxicol. 2019, 102, 146–152. [Google Scholar] [CrossRef]
- Jin, D.; Zhang, Q.; Liu, Y.; Liang, M.; Li, A.; Wu, J. Overexpression of the maize phytochelatin synthase gene (ZmPCS1) enhances Cd tolerance in plants. Acta Physiol. Plant 2022, 44, 114. [Google Scholar] [CrossRef]
- Peng, J.S.; Ding, G.; Yi, H.Y.; Gong, J.M. Cloning and functional analysis of phytochelatin synthase gene from Sedum plumbizincicola. Plant Physiol. J. 2014, 50, 625–633. [Google Scholar]
- Li, J.; Guo, J.; Xu, W.; Ma, M. RNA Interference-mediated Silencing of Phytochelatin Synthase Gene Reduce Cadmium Accumulation in Rice Seeds. J. Integr. Plant Biol. 2007, 49, 1032–1037. [Google Scholar] [CrossRef]
- Gasic, K.; Korban, S.S. Expression of Arabidopsis phytochelatin synthase in Indian mustard (Brassica juncea) plants enhances tolerance for Cd and Zn. Planta 2007, 225, 1277–1285. [Google Scholar] [CrossRef]
- Gasic, K.; Korban, S.S. Transgenic Indian mustard (Brassica juncea) plants expressing an Arabidopsis phytochelatin synthase (AtPCS1) exhibit enhanced As and Cd tolerance. Plant Mol. Biol. 2007, 64, 361–369. [Google Scholar] [CrossRef]
- Venkataramaiah, N.; Ramakrishna, S.V.; Sreevathsa, R. Overexpression of phytochelatin synthase (AtPCS) in rice for tolerance to cadmium stress. Biologia 2011, 66, 1060–1073. [Google Scholar] [CrossRef]
- Wang, F.; Wang, Z.; Zhu, C. Heteroexpression of the wheat phytochelatin synthase gene (TaPCS1) in rice enhances cadmium sensitivity. Acta Biochim. Biophys. Sin. 2012, 44, 886–893. [Google Scholar] [CrossRef]
- Zhao, C.; Xu, J.; Li, Q.; Li, S.; Wang, P.; Xiang, F. Cloning and characterization of a Phragmites australis phytochelatin synthase (PaPCS) and achieving Cd tolerance in tall fescue. PLoS ONE 2014, 9, e103771. [Google Scholar] [CrossRef]
- Das, N.; Bhattacharya, S.; Bhattacharyya, S.; Maiti, M.K. Identification of alternatively spliced transcripts of rice phytochelatin synthase 2 gene OsPCS2 involved in mitigation of cadmium and arsenic stresses. Plant Mol. Biol. 2017, 94, 167–183. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, S.; Kuramata, M.; Abe, T.; Takagi, H.; Ozawa, K.; Ishikawa, S. Phytochelatin synthase OsPCS1 plays a crucial role in reducing arsenic levels in rice grains. Plant J. 2017, 91, 840–848. [Google Scholar] [CrossRef] [PubMed]
- Uraguchi, S.; Tanaka, N.; Hofmann, C.; Abiko, K.; Ohkama-Ohtsu, N.; Weber, M.; Kamiya, T.; Sone, Y.; Nakamura, R.; Takanezawa, Y.; et al. Phytochelatin Synthase has Contrasting Effects on Cadmium and Arsenic Accumulation in Rice Grains. Plant Cell Physiol. 2017, 58, 1730–1742. [Google Scholar] [CrossRef]
- Yamazaki, S.; Ueda, Y.; Mukai, A.; Ochiai, K.; Matoh, T. Rice phytochelatin synthases OsPCS1 and OsPCS2 make different contributions to cadmium and arsenic tolerance. Plant Direct 2018, 2, e00034. [Google Scholar] [CrossRef] [PubMed]
- Park, H.C.; Hwang, J.E.; Jiang, Y.; Kim, Y.J.; Kim, S.H.; Nguyen, X.C.; Kim, C.Y.; Chung, W.S. Functional characterisation of two phytochelatin synthases in rice (Oryza sativa cv. Milyang 117) that respond to cadmium stress. Plant Biol. 2019, 21, 854–861. [Google Scholar] [CrossRef]
- Li, M.; Barbaro, E.; Bellini, E.; Saba, A.; Sanità di Toppi, L.; Varotto, C. Ancestral function of the phytochelatin synthase C-terminal domain in inhibition of heavy metal-mediated enzyme overactivation. J. Exp. Bot. 2020, 71, 6655–6669. [Google Scholar] [CrossRef]
- Li, M.; Leso, M.; Buti, M.; Bellini, E.; Bertoldi, D.; Saba, A.; Larcher, R.; Sanità di Toppi, L.; Varotto, C. Phytochelatin synthase de-regulation in Marchantia polymorpha indicates cadmium detoxification as its primary ancestral function in land plants and provides a novel visual bioindicator for detection of this metal. J. Hazard. Mater. 2022, 440, 129844. [Google Scholar] [CrossRef]
- Gui, Y.; Teo, J.; Tian, D.; Yin, Z. Genetic engineering low-arsenic and low-cadmium rice grain. J. Exp. Bot. 2024, 75, 2143–2155. [Google Scholar] [CrossRef]
- Pandey, N.; Rai, K.K.; Rai, S.K.; Pandey-Rai, S. Heterologous expression of cyanobacterial PCS confers augmented arsenic and cadmium stress tolerance and higher artemisinin in Artemisia annua hairy roots. Plant Biotechnol. Rep. 2021, 15, 317–334. [Google Scholar] [CrossRef]
- Wang, P.; Yang, N.; Zhang, A.; Tangnver, S.; Li, S.; Gao, C. Cloning ThPCS1 gene of Tamarix hispida to improve cadmium tolerance. J. Nanjing For. Univ. 2021, 45, 71–78. [Google Scholar]
- Xing, Q.; Hasan, M.K.; Li, Z.; Yang, T.; Jin, W.; Qi, Z.; Yang, P.; Wang, G.; Ahammed, G.J.; Zhou, J. Melatonin-induced plant adaptation to cadmium stress involves enhanced phytochelatin synthesis and nutrient homeostasis in Solanum lycopersicum L. J. Hazard. Mater. 2023, 456, 131670. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Komives, E.A.; Schroeder, J.I. An Improved Grafting Technique for Mature Arabidopsis Plants Demonstrates Long-Distance Shoot-to-Root Transport of Phytochelatins in Arabidopsis. Plant Physiol. 2006, 141, 108–120. [Google Scholar] [CrossRef] [PubMed]
- Postrigan, B.N.; Knyazev, A.B.; Kuluev, B.R.; Yakhin, O.I.; Chemeris, A.V. Expression of the Synthetic Phytochelatin Gene in Tobacco. Russ. J. Plant Physiol. 2012, 59, 275–280. [Google Scholar] [CrossRef]
- Zheng, Y.; Li, M.; Liu, B.; Qin, Y.; Li, J.; Pan, Y.; Zhang, X. Effects of Phytochelatin-like Gene on the Resistance and Enrichment of Cd²⁺ in Tobacco. Int. J. Mol. Sci. 2022, 23, 16167. [Google Scholar] [CrossRef]
- Vershinina, Z.R.; Maslennikova, D.R.; Chubukova, O.V.; Khakimova, L.R.; Mikhaylova, E.V. Synthetic Pph6his Gene Confers Resistance to Cadmium Stress in Transgenic Tobacco. Russ. J. Plant Physiol. 2023, 70, 83. [Google Scholar] [CrossRef]
- Li, F.; Deng, Y.; Liu, Y.; Mai, C.; Xu, Y.; Wu, J.; Zheng, X.; Liang, C.; Wang, J. Arabidopsis transcription factor WRKY45 confers cadmium tolerance via activating PCS1 and PCS2 expression. J. Hazard. Mater. 2023, 460, 132496. [Google Scholar] [CrossRef]
- Ding, G.; Peng, J.; Zhang, G.; Yi, H.; Fu, Y.; Gong, J. Regulation of the Phytochelatin Synthase Gene AtPCS2 in Arabidopsis thaliana. Sci. Sin. Vitae 2013, 43, 1112–1118. [Google Scholar] [CrossRef]
- Marques, D.N.; Piotto, F.A.; Azevedo, R.A. Phosphoproteomics: Advances in Research on Cadmium-Exposed Plants. Int. J. Mol. Sci. 2024, 25, 12431. [Google Scholar] [CrossRef]
- Marques, D.N.; Gaziola, S.A.; Azevedo, R.A. Phytochelatins and their relationship with modulation of cadmium tolerance in plants. In Handbook of Bioremediation: Physiological, Molecular and Biotechnological Interventions; Hasanuzzaman, M., Prasad, M.N.V., Eds.; Elsevier: Cambridge, MA, USA, 2021; pp. 91–113. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Overall Experimental Genetic Approach | Genes; Locus; Allelic Series | Examples of Main Findings Related to PCs in Cd-Exposed Plants | Citation |
|---|---|---|---|
| Isolation and characterization of Cd-sensitive mutants | CAD1 locus (cad1-1, cad1-2) | Mutants show reduced Cd sequestration, but BSO (PC biosynthesis inhibitor) further increases sensitivity non-additively, suggesting PCs are still produced and cad1 affects a related detoxification aspect. | [6] |
| Isolation and characterization of Cd-sensitive cad1 mutants | CAD1 locus (cad1-1, cad1-2, cad1-3, cad1-4, cad1-5) | Mutants show deficient PC accumulation and PCS activity correlating with increased Cd sensitivity; cad1-3 (most sensitive) shows no detectable PCs; BSO increases wild-type Cd sensitivity but has little effect on cad1-3, suggesting a primary role for PCs in Cd detoxification. | [7] |
| Positional cloning and characterization of cad1 mutants; functional complementation; heterologous expression in E. coli | CAD1 gene (MRH10.11) | cad1 mutants are Cd-sensitive and PC-deficient with lack of PC synthase activity; CAD1 encodes a GSH-dependent, heavy metal-activated PC synthase; CAD1 mRNA expression is not induced by Cd. | [8] |
| Functional characterization and expression analysis of a second PCS gene in Arabidopsis thaliana through heterologous expression in yeast and comparison with cad1 mutants | AtPCS2, AtPCS1 (CAD1) | AtPCS2 encodes a functional PC synthase in yeast, but cad1 mutants (lacking functional AtPCS1) show no detectable PCs upon Cd exposure in Arabidopsis, indicating non-redundancy and that AtPCS1 is likely the primary Cd-inducible PC synthase in Arabidopsis. | [9] |
| Comparison of Cd uptake and accumulation in PC-deficient cad1-3 mutant and wild type Arabidopsis with and without prior Cd exposure. | cad1-3 (deficient in PC synthase activity) | The PC-deficient mutant accumulated less Cd than the wild type at higher growth Cd concentrations, with lower Cd translocation to the shoot, suggesting PCs enhance Cd uptake and translocation. | [10] |
| Characterization of AtPCS1 promoter activity using GUS reporter gene in transgenic Arabidopsis; analysis of AtPCS1 protein accumulation using FLAG-tagged constructs; investigation of Cd’s effect on AtPCS1 protein levels; comparison of expression using genomic DNA vs. cDNA of AtPCS1. | AtPCS1 | AtPCS1 promoter shows differential spatial and temporal activity, including strong expression in leaf trichomes; AtPCS1 protein levels correlate with promoter activity; AtPCS1 protein expression is not post-transcriptionally regulated by Cd; introns enhance AtPCS1 mRNA and protein accumulation. | [11] |
| Overexpression of wheat PCS gene (TaPCS1) in Nicotiana glauca | TaPCS1 | Transgenic N. glauca seedlings showed increased tolerance to Cd (longer roots and greener leaves) compared to wild-type, suggesting enhanced PC activity due to TaPCS1 overexpression. | [12] |
| Expression of wheat TaPCS1 cDNA in Arabidopsis cad1-3 mutant (root-specific and ectopic promoters); analysis of Cd transport and PC levels | Wheat TaPCS1, Arabidopsis AtPCS1 (cad1-3 locus) | Root-specific TaPCS1 expression leads to PC transport from roots to shoots, enhanced root-to-shoot Cd transport, and reduced Cd accumulation in roots, all of which are PC-dependent. | [13] |
| Generation and characterization of transgenic Arabidopsis overexpressing AtPCS1 under the CaMV 35S promoter; analysis of Cd tolerance, accumulation, and PC production | AtPCS1 | High levels of AtPCS1 overexpression led to increased PC production but did not necessarily enhance Cd tolerance or accumulation, and in some cases caused increased Cd sensitivity; moderately overexpressing lines showed increased tolerance and accumulation. | [14] |
| Overexpression of AtPCS1 in transgenic Arabidopsis under the control of the AtPCS1 promoter | AtPCS1 | Overexpression led to increased AtPCS1 mRNA and PC production under Cd stress but paradoxically resulted in hypersensitivity to Cd and Zn (not Cu), possibly due to toxicity of PCs at supraoptimal levels compared to GSH. | [15] |
| Overexpression of AtPCS1 in Arabidopsis thaliana | AtPCS1 | Overexpression led to increased PC levels upon Cd exposure but surprisingly resulted in Cd hypersensitivity compared to wild-type plants. | [16] |
| Expression analysis and complementation studies of AtPCS2 in the AtPCS1-deficient cad1-3 mutant using various promoters | AtPCS2, AtPCS1 (CAD1), cad1-3 | AtPCS2 expression is weak compared to AtPCS1 and cannot fully complement the cad1-3 mutant’s Cd sensitivity even when overexpressed, suggesting low expression levels limit its role in PC production in this context. | [17] |
| Transcript level analysis of AtMRPs under Cd treatment using wild-type and mutant plants | AtMRP3, cad2 (GSH deficient), cad1-3 (PC deficient) | AtMRP3-related induction by Cd is likely independent of GSH and PC biosynthesis/levels. | [18] |
| Analysis of Cd sensitivity in PC-deficient Arabidopsis mutants | cad1-3 (PCS deficient) | Deficiency in PC production leads to increased sensitivity to Cd. | [19] |
| Analysis of Cd sensitivity and root-to-shoot translocation using hma2, hma4, and cad1 single and multiple mutants | HMA2, HMA4, CAD1; hma2-4, hma4-2, cad1-3 | PC deficiency resulted in increased shoot Cd and percentage translocation, but had a minor effect compared to hma2 and hma4 loss. | [20] |
| Analysis of Cd tolerance in metallothionein and PC-deficient Arabidopsis mutants | cad1-3, mt1a-2, mt2b-1, mt1a-2, mt2b-1, cad1-3 | Combined deficiency of PCs and MTs (MT1a, MT2b) leads to increased sensitivity to Cd. | [21] |
| Analysis of Cd sensitivity in Arabidopsis PC-deficient mutants | AtPCS1; cad1-3, cad1-6 | PC deficiency leads to pronounced hypersensitivity to Cd. | [22] |
| Characterization of knockout mutants (single and double) and overexpression lines in Arabidopsis; heterologous expression in yeast. | AtABCC1 (At1g30400); AtABCC2 (At2g34660); atabcc1, atabcc2, atabcc1 atabcc2 (knockouts); AtABCC1 (overexpression) | AtABCC1 and AtABCC2 are important for vacuolar sequestration of PC–Cd(II), conferring tolerance; atabcc1 atabcc2 shows reduced vacuolar Cd. | [23] |
| Characterization of a loss-of-function mutant | PCS1 (CAD1); cad1-3 | Loss of PCS1 function leads to Cd hypersensitivity due to the inability to synthesize PCs under Cd stress. | [24] |
| Characterization of a T-DNA tagged knockout mutant in Arabidopsis; in silico 3D structure prediction and in vivo metal accumulation analysis. | PCS1 (At5g44070); SAIL_650_C12 (homozygous T-DNA insertion mutant) | The pcs1 mutant (SAIL_650_C12) shows higher Cd accumulation in shoots and increased Cd sensitivity due to the inability to synthesize PCs. | [25] |
| Characterization of knockout mutants | PCS (non-functional in cad1-3); cad2-1, rax1-1, cad1-3 | cad1-3 mutant deficient in PCS shows no PC accumulation under Cd exposure, confirming its role in PC synthesis; Cd is a stronger inducer of PCs than Hg. | [26] |
| Characterization of knockout mutant (abcc3) and overexpression lines (AtABCC3ox) in Arabidopsis; complementation assay in atabcc1 atabcc2 mutant; pharmacological inhibition of PC synthesis (BSO); analysis of Cd localization using fluorescent dyes. | AtABCC3 (AtMRP3); abcc3 (knockout); AtABCC3ox (overexpression); cad1-3 (PCS deficient); atabcc1 atabcc2 (double knockout) | AtABCC3 is involved in vacuolar transport of PC–Cd complexes, contributing to Cd tolerance; abcc3 mutants are Cd-sensitive with reduced vacuolar Cd; AtABCC3ox lines show increased Cd tolerance and vacuolar Cd; AtABCC3 function in Cd tolerance is PC-dependent and can complement atabcc1 atabcc2 in the presence of PCs. | [27] |
| Directed evolution of Arabidopsis thaliana PCS (AtPCS1); heterologous expression of wild-type and mutant AtPCS1 in yeast (Saccharomyces cerevisiae); ectopic expression of wild-type and mutant AtPCS1 in Arabidopsis thaliana and Brassica juncea. | AtPCS1 (GenBankTMAF085230); various mutant variants (e.g., Y186C) | Specific AtPCS1 mutants with diminished catalytic activity conferred improved Cd tolerance and increased PC accumulation in yeast, Arabidopsis, and B. juncea. This was linked to the maintenance of glutathione and γ-glutamylcysteine precursor levels and redox homeostasis during Cd exposure, unlike the overexpression of wild-type AtPCS1, which could deplete these metabolites. | [28] |
| Characterization of Arabidopsis AtPCS1 knockout mutants (cad1-3 and cad1-6) grown on Cd-contaminated soil; heterologous expression of truncated AtPCS1 in yeast. | AtPCS1; cad1-3 (null mutant), cad1-6 (T-DNA insertion) | cad1-3 shows Cd hypersensitivity and no detectable PC accumulation; cad1-6 is more Cd tolerant with trace PCs in leaves; both mutants have reduced leaf Cd; truncated AtPCS1 can retain Cd-activated PC synthesis. | [29] |
| Characterization of a PC-deficient knockout mutant (cad1-3) in Arabidopsis; comparison with wild-type under Cd exposure (with/without supplemental Cu). | PCS1 (CAD1); cad1-3 | cad1-3 mutant lacking PCs is more sensitive to Cd; Cd induces a Cu deficiency response dependent on PCs (reduced in cad1-3); PCs likely complex both Cd and Cu; cad1-3 shows altered metal accumulation and reduced Cd translocation. | [30] |
| Comparison of Arabidopsis thaliana wild-type and the cad1-3 knockout mutant (deficient in AtPCS1) under control and Cd exposure conditions. | PCS1 (AtPCS1); cad1-3 | cad1-3 mutant shows defective callose deposition after Cd exposure and increased Cd sensitivity, suggesting AtPCS1 contributes to Cd tolerance not only through PC synthesis but also via callose deposition. | [31] |
| Forward genetics to isolate a Cd-hypersensitive mutant in PC-deficient (cad1-3) background; characterization of the identified PP2A-4C mutant and its interaction with ethylene production under Cd stress. | PCS1 (CAD1); cad1-3 (PC-deficient mutant); PP2A-4C; cdsr1 (allele isolated in cad1-3 background); pp2a-4c-1 (T-DNA insertion mutant for PP2A-4C). | The study used a PC-deficient mutant (cad1-3) to explore Cd toxicity responses beyond the PC pathway. The identified PP2A-4C mutation exacerbated Cd sensitivity in the cad1-3 background, indicating a PC-independent mechanism of Cd tolerance modulated by PP2A via ethylene production. | [32] |
| Forward genetic screen for Cd-hypersensitive mutants in PC-deficient (cad1-3) background; identification and characterization of cdsr2 mutant with a mutation in IAR4; analysis of Cd accumulation and thiol compound biosynthesis (including PCs). | PCS1 (CAD1); cad1-3 (PC-deficient mutant); IAR4; cdsr2 (allele isolated in cad1-3 background); cdsr2 cad1-3 (double mutant); iar4-2, iar4-8 (T-DNA insertion mutants for IAR4). | The study used the PC-deficient cad1-3 mutant to isolate further Cd-sensitive mutants, revealing that IAR4 mutation exacerbates Cd sensitivity independently of PC synthesis. In the Col-0 background, IAR4 mutation led to increased Cys, GSH, PC2, and PC3 under Cd stress, suggesting a heightened stress response. As expected, PCs were undetectable in cad1-3 and cdsr2 cad1-3 under Cd stress due to a defective PCS1 gene. | [33] |
| Expression analysis (RT-PCR, promoter-LUC assays), protein analysis (Western blot), characterization of sbp1 null mutant and SBP1 overexpression lines in wild-type and PC-deficient (cad) Arabidopsis, and in ycf1 yeast mutant; in vitro Cd binding assays of SBP1. | SBP1, SBP2, SBP3; sbp1 (null mutant); SBP1 (overexpression lines); PCS1 (CAD1) (cad mutants lacking PCs); YCF1 (Saccharomyces cerevisiae mutant). | SBP1 expression is induced by Cd. The sbp1 mutant showed no clear phenotype, possibly due to SBP2 upregulation. Overexpression of SBP1 enhanced Cd accumulation in roots but reduced Cd sensitivity in wild-type and, notably, in cad mutants (lacking PCs), suggesting a PC-independent Cd detoxification role potentially through direct Cd binding. | [34] |
| Overexpression of AtPCS1 (cytosol and chloroplast targeting) | AtPCS1 from Arabidopsis thaliana, targeted to cytosol and chloroplast. | Increased PC production upon Cd exposure; cytosolic overexpression led to Cd hypersensitivity; plastidial overexpression showed no significant effect on Cd tolerance. | [35] |
| Overexpression of AtPCS1 in Nicotiana tabacum | AtPCS1 from Arabidopsis thaliana expressed in tobacco. | Increased PC production and Cd tolerance/accumulation in roots and shoots (GSH-dependent), but no enhanced Cd translocation to the shoot. | [36] |
| Overexpression of E. coli cysE, E. coli gshA, and S. pombe PCS | Genes involved in cysteine and glutathione biosynthesis from E. coli and PC synthase from Schizosaccharomyces pombe. | Increased non-protein thiols; higher Cd concentration in roots, no significant increase in shoots after Cd exposure. | [37] |
| Overexpression of CdPCS1 in tobacco | CdPCS1 from Cynodon dactylon expressed in tobacco. | Increased PC levels and enhanced Cd accumulation and tolerance in transgenic plants. | [38] |
| Overexpression of AtPCS1 in Arabidopsis thaliana | AtPCS1 from Arabidopsis thaliana (pcs9 line). | Overexpression did not increase the maximum capacity for non-protein thiol (NPT) production upon Cd exposure; transgenic line showed Cd hypersensitivity. | [39] |
| Overexpression of AtPCS1 in N. tabacum | AtPCS1 from Arabidopsis thaliana expressed in tobacco. | Increased PCS activity, moderate PC increase, strong γ-EC accumulation, GSH depletion in leaves led to Cd hypersensitivity. | [40] |
| Simultaneous overexpression of AsPCS1 and GSH1 in A. thaliana | AsPCS1 (Arsenate reductase-coupled PCS 1) and GSH1 (Glutathione synthetase) from an unspecified source, co-expressed in Arabidopsis. | Dual-gene transgenic lines showed higher Cd accumulation but not the highest PC content under Cd exposure compared to single-gene overexpressing lines and wild-type. | [41] |
| Overexpression of AtPCS1 in Nicotiana tabacum (lines PaII12) | AtPCS1 from Arabidopsis thaliana expressed in specific transgenic lines of tobacco. | Decreased cytosolic and vacuolar PC levels in Cd-exposed plants led to Cd hypersensitivity despite no significant change in vacuolar Cd accumulation compared to wild-type. | [42] |
| Overexpression of AtPCS1 in Arabidopsis | AtPCS1 from Arabidopsis thaliana expressed in Arabidopsis. | Increased PC content in AtPCSox lines upon Cd exposure; lower Cd tolerance at low Cd but higher tolerance at high Cd compared to wild type; exogenous GSH did not increase Cd tolerance in AtPCSox lines. | [43] |
| Overexpression of TcPCS1 in N. tabacum | TcPCS1 from Thlaspi caerulescens expressed in tobacco. | Enhanced PC production and increased Cd accumulation in roots and shoots; longer root length under Cd stress. | [44] |
| Simultaneous overexpression of AsPCS1 and YCF1 in A. thaliana | AsPCS1 (Arsenate reductase-coupled PCS 1) and YCF1 (vacuolar membrane protein involved in heavy metal tolerance) from unspecified sources, co-expressed in Arabidopsis. | Dual-gene lines showed higher PC content after Cd exposure and higher Cd accumulation than single-gene lines and wild type, indicating increased tolerance. | [45] |
| Heterologous overexpression of NnPCS1 in A. thaliana | NnPCS1 from Nelumbo nucifera expressed in Arabidopsis thaliana. | Increased PC content and enhanced Cd accumulation in transgenic Arabidopsis plants. | [46] |
| Heterologous overexpression of CdPCS1 in Arabidopsis thaliana | CdPCS1 from Cynodon dactylon expressed in Arabidopsis thaliana. | Enhanced accumulation of heavy metal(loid)s including Cd in aerial parts without significant growth difference compared to wild type, suggesting potential for phytoremediation. | [47] |
| Overexpression of PtPCS in N. tabacum | PtPCS from an unspecified plant source expressed in tobacco. | Transgenic plants showed enhanced Cd tolerance and higher Cd accumulation in roots and leaves upon Cd exposure compared to wild type, but a lower transfer coefficient. | [48] |
| Overexpression of NtPCS1 in Nicotiana tabacum (sense lines) | NtPCS1 from Nicotiana tabacum overexpressed in tobacco. | Enhanced Cd tolerance but no significant change in Cd accumulation was observed compared to control plants. | [49] |
| Expression of NtPCS1 in Nicotiana tabacum (antisense lines) | NtPCS1 from Nicotiana tabacum underexpressed in tobacco. | Showed growth retardation in the early stage suggesting a role in plant development; PC levels under Cd exposure were not explicitly detailed. | [49] |
| Overexpression of AtPCS1 in Nicotiana tabacum (rolB-AtPCS1) | AtPCS1 from Arabidopsis thaliana expressed in tobacco lines with the rolB gene. | Transgenic plants showed higher PC levels and increased Cd accumulation in roots when exposed to Cd, but inhibited Cd extrusion from leaves. | [50] |
| Overexpression of MnPCS1 and MnPCS2 in Arabidopsis thaliana | MnPCS1 and MnPCS2 from Mimulus guttatus expressed in Arabidopsis thaliana. | Transgenic Arabidopsis lines showed higher Cd accumulation in shoots and roots compared to wild type when grown in Cd-containing medium, suggesting enhanced PC activity led to increased metal uptake and sequestration. | [51] |
| Overexpression of AdPCS2 or AdPCS3 in Arabidopsis thaliana | AdPCS2 and AdPCS3 from Arundo donax expressed in Arabidopsis thaliana. | Transgenic Arabidopsis lines exhibited significant growth reduction and chlorosis when treated with Cd, suggesting that the overexpression of these A. donax PCS genes can lead to enhanced Cd sensitivity in A. thaliana. | [52] |
| Overexpression of truncated IpPCS1 in Arabidopsis | Truncated IpPCS1 from Ipomoea pes-caprae expressed in Arabidopsis. | Transgenic seedlings showed slightly improved Cd tolerance, but adult plants displayed no obvious effects on growth or Cd accumulation compared to the wild type under Cd stress. | [53] |
| Overexpression of BnPCS1 from Boehmeria nivea in Arabidopsis thaliana | BnPCS1 from ramie expressed in Arabidopsis thaliana. | Transgenic Arabidopsis showed enhanced Cd tolerance, higher Cd accumulation in shoots, and an increased translocation factor of Cd from roots to shoots compared to wild type under Cd stress, suggesting improved phytoremediation potential. | [54] |
| Transformation of cad1-3 mutant and wild type Arabidopsis with leaf-specifically expressed AtPCS1 | AtPCS1; cad1-3 | Leaf-specific AtPCS1 restored leaf PCS activity and PC production in cad1-3 without increasing total leaf PC levels compared to wild type, increased Cd tolerance but did not limit Cd accumulation to leaves | [55] |
| Analysis of cad1-3 mutant and transgenic Arabidopsis ectopically expressing wheat TaPCS1 | AtPCS1;cad1-3;TaPCS1 | Transgenic plants showed higher Cd-PC2 levels in roots and shoots than wild type; more Cd-PC2 found in shoots than roots in both, indicating PC contribution to shoot Cd accumulation | [56] |
| Complementation of Arabidopsis cad1-3 mutant and overexpression in transgenic tobacco with CdPCS1 from Ceratophyllum demersum | CdPCS1;AtPCS1(CAD1);cad1-3 | Transgenic tobacco showed increased PC content and enhanced Cd accumulation; CdPCS1 complemented cad1-3 Cd sensitivity. | [57] |
| Expression of synthetic PC (EC) genes in wild type and PC-deficient cad1-3 mutant Arabidopsis | EC14,EC16,EC20;AtPCS1(CAD1);cad1-3 | Expression of ECs complemented cad1-3 Cd sensitivity; transgenic plants showed enhanced heavy metal(loid) accumulation (including Cd) compared to controls. | [58] |
| Analysis of AtPCS2 overexpression in cad1-3 mutant to assess Cd tolerance and PC accumulation | AtPCS2 overexpression in AtPCS1 (CAD1) mutant cad1-3 | Overexpression led to constitutive PC2 production, partial rescue of cad1-3 Cd hypersensitivity on soil, and reduced PC3 levels upon Cd exposure compared to AtPCS1 overexpression | [59] |
| Expression of C. elegans CePCS in Arabidopsis AtPCS1-deficient cad1-3 mutant | AtPCS1 (CAD1); cad1-3; CePCS | CePCS expression in cad1-3 restored PC synthesis and Cd tolerance | [60] |
| Overexpression of Vicia sativa PCS1 homolog (VsPCS1) in wild-type and AtPCS1-deficient Arabidopsis (atpcs1) | VsPCS1;AtPCS1;atpcs1 | VsPCS1 overexpression increased PC synthesis and Cd tolerance; complemented atpcs1 PC deficiency and Cd sensitivity; led to higher-order PCs (PC4). | [61] |
| Overexpression of three duplicated BnPCS genes from Brassica napus in Arabidopsis thaliana cad1–3 mutant. | BnPCS (three duplicated genes); AtPCS1 (CAD1); cad1-3 | Transgenic lines showed higher PC content and enhanced Cd tolerance, accumulation, and translocation compared to cad1-3 under Cd stress. | [62] |
| Overexpression of maize ZmPCS1 in Arabidopsis and complementation of Arabidopsis atpcs1 mutant | ZmPCS1; AtPCS1; atpcs1 | Overexpression in Arabidopsis enhanced Cd tolerance and accumulation, along with increased GSH and PC contents; ZmPCS1 rescued the Cd-sensitive phenotype of the atpcs1 mutant. | [63] |
| Isolation and functional characterization of SepPCS from Sedum plumbizincicola by complementation in yeast and Arabidopsis cad1-3 mutant | SepPCS (from S. plumbizincicola); AtPCS1 (CAD1); cad1-3 | SepPCS expression restored PCs biosynthesis and Cd tolerance in mutants; PCs’ role in S. plumbizincicola Cd tolerance might increase with elevated Cd. | [64] |
| RNAi silencing of PCS in rice seeds. | OsPCS1 (GenBank AF439787); ZMM1 (seed-specific promoter). | OsPCS1 silencing in seeds reduced Cd accumulation by ~50%. | [65] |
| Overexpression of Arabidopsis PCS in Indian mustard. | AtPCS1 cDNA under the 35S promoter. | Moderate AtPCS1 overexpression increased PCs and Cd tolerance but decreased Cd accumulation. | [66] |
| Overexpression of Arabidopsis PCS in Indian mustard. | AtPCS1 gDNA under its native promoter. | Higher PC levels under Cd stress and lower Cd accumulation in shoots with enhanced Cd tolerance. | [67] |
| Overexpression of Arabidopsis PCS in rice. | AtPCS gene; hpt II marker gene; Oryza sativa genotype Azucena. | Enhanced Cd tolerance, increased PC content, reduced GSH depletion, higher root Cd. | [68] |
| Heteroexpression of wheat PCS in rice. | TaPCS1 cDNA under the 35S promoter; Oryza sativa L., cv. Zhonghua 11. | Enhanced Cd sensitivity, increased shoot Cd, higher PC and NPT in shoots under Cd. | [69] |
| Heterologous expression of Phragmites australis PCS in tall fescue. | PaPCS cDNA under the 35S promoter. | Higher PC levels, enhanced Cd tolerance, and increased Cd accumulation. | [70] |
| Seed-specific RNAi to silence OsPCS genes. | OsPCS1; OsPCS2; endosperm-specific GluC promoter. | OsPCS1 and OsPCS2 silencing in rice grains led to ~51% reduction in Cd content. | [71] |
| Characterization of a PCS mutant. | OsPCS1 (Os05g0415200); has2 (three-base deletion in catalytic domain). | has2 mutant (impaired OsPCS1) showed similar Cd tolerance and grain Cd content to wild type. | [72] |
| Characterization of OsPCS1 loss-of-function mutants (T-DNA and Tos17 insertion lines) in rice. | OsPCS1 (Os06g0102300/LOC_Os06g01260); T-DNA insertion; Tos17 insertion (NG5045). | OsPCS1 mutants showed increased Cd sensitivity, decreased Cd in shoots/grains, reduced PCs under Cd. | [73] |
| RNAi-mediated silencing of OsPCS2 expression. | OsPCS2 gene; constitutive maize ubiquitin1 promoter. | OsPCS2-silenced plants showed reduced PC levels under Cd stress, but Cd tolerance was little affected. | [74] |
| Overexpression of two rice PCS genes in Arabidopsis thaliana. | OsPCS5; OsPCS15; under the CaMV 35S promoter. | Overexpression paradoxically increased sensitivity to Cd, possibly due to excess PC. | [75] |
| Functional characterization of the first PCS gene from a liverwort using yeast and Arabidopsis cad1-3 complementation. | MpPCS (Mapoly0046s0028.1); cad1-3 (Arabidopsis AtPCS1 null mutant). | MpPCS overexpression in cad1-3 complemented Cd sensitivity and restored PC2/PC3 production. | [76] |
| Generation and characterization of M. polymorpha knockout mutants and overexpressing lines for PCS. | MpPCS gene; CRISPR/Cas9 knockout (four alleles); overexpression lines (MpPCS-ox-3, MpPCS-ox-7). | Mppcs mutants Cd hypersensitive, no PC; overexpressors also Cd hypersensitive, reduced GSH/PCn, higher Cd accumulation. | [77] |
| Overexpression of a rice PCS gene. | OsPCS1 (Os05g0415200) cDNA under the rice OsActin1 promoter. | Increased PC, decreased grain Cd, but caused seedling Cd hypersensitivity by suppressing vacuolar Cd sequestration. | [78] |
| Heterologous expression of cyanobacterial PCS in Artemisia annua hairy roots. | anaPCS gene (from Anabaena PCC 7120) under CMV35S promoter. | Transformed hairy roots showed significantly higher PC content and greater Cd accumulation. | [79] |
| Overexpression of Tamarix hispida PCS gene. | ThPCS1 gene (full length, 1581 bp ORF) from Tamarix hispida; pROKII-ThPCS1 vector. | ThPCS1 overexpressors showed less reactive oxygen species and Cd ions under Cd stress. | [80] |
| VIGS of PCS in tomato. | PCS | PCS silencing aggravated Cd phytotoxicity; melatonin-induced Cd tolerance partly depends on PCs. | [81] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marques, D.N.; Thiengo, C.C.; Azevedo, R.A. Phytochelatins and Cadmium Mitigation: Harnessing Genetic Avenues for Plant Functional Manipulation. Int. J. Mol. Sci. 2025, 26, 4767. https://doi.org/10.3390/ijms26104767
Marques DN, Thiengo CC, Azevedo RA. Phytochelatins and Cadmium Mitigation: Harnessing Genetic Avenues for Plant Functional Manipulation. International Journal of Molecular Sciences. 2025; 26(10):4767. https://doi.org/10.3390/ijms26104767
Chicago/Turabian StyleMarques, Deyvid Novaes, Cássio Carlette Thiengo, and Ricardo Antunes Azevedo. 2025. "Phytochelatins and Cadmium Mitigation: Harnessing Genetic Avenues for Plant Functional Manipulation" International Journal of Molecular Sciences 26, no. 10: 4767. https://doi.org/10.3390/ijms26104767
APA StyleMarques, D. N., Thiengo, C. C., & Azevedo, R. A. (2025). Phytochelatins and Cadmium Mitigation: Harnessing Genetic Avenues for Plant Functional Manipulation. International Journal of Molecular Sciences, 26(10), 4767. https://doi.org/10.3390/ijms26104767