


Identification of Kunitz-Type Inhibitor Gene Family of Populus yunnanensis Reveals a Stress Tolerance Function in Inverted Cuttings
 and
and
Abstract
1. Introduction
2. Results
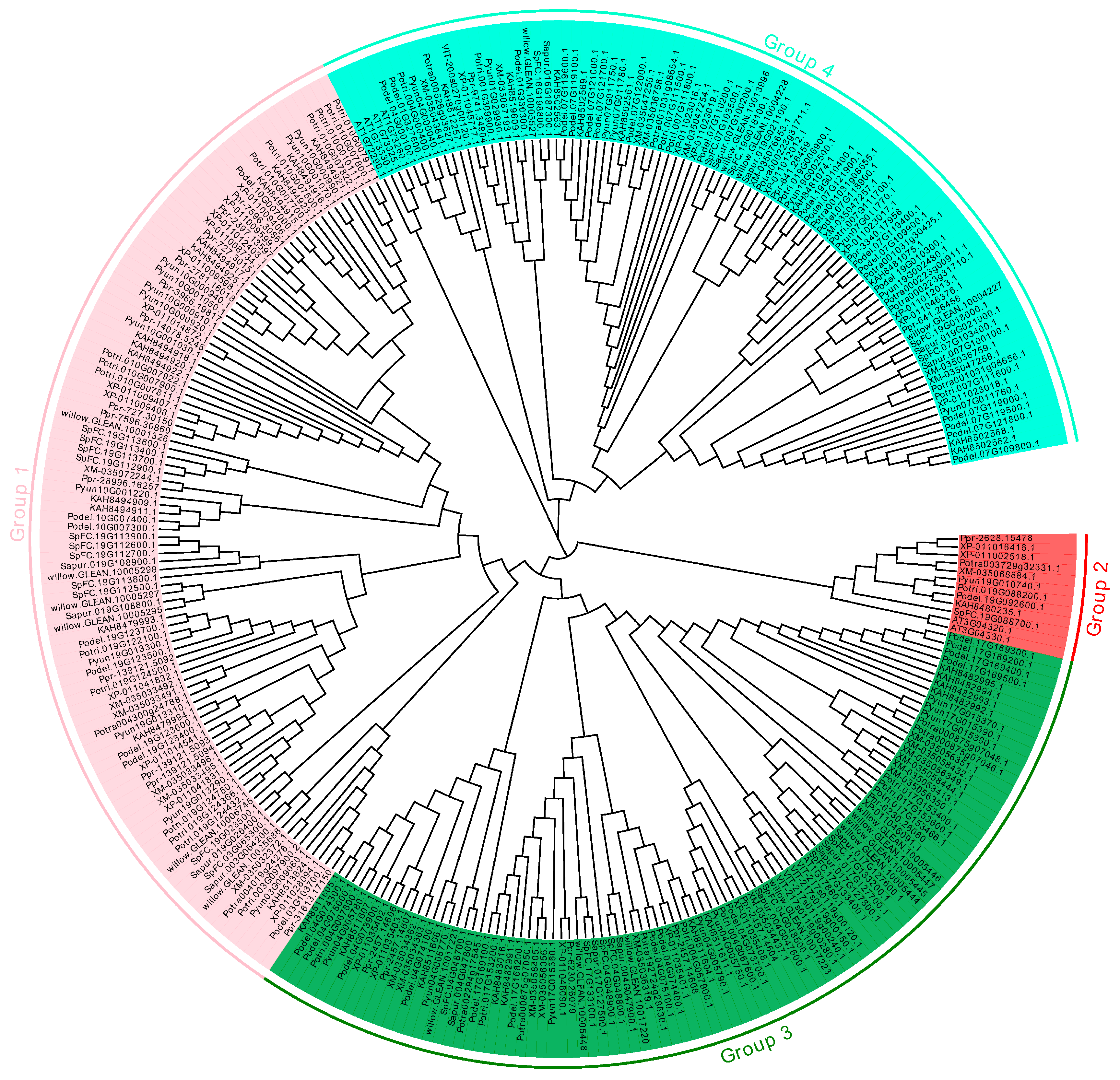
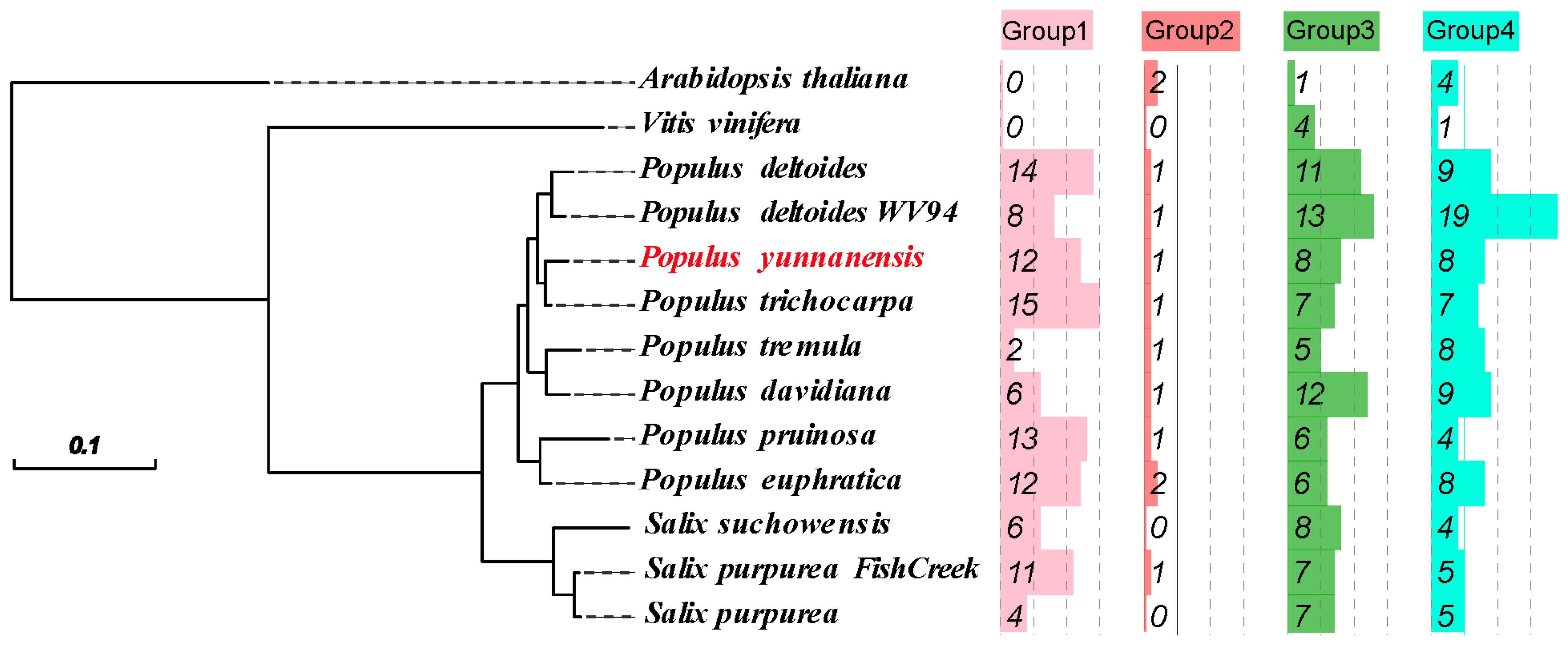
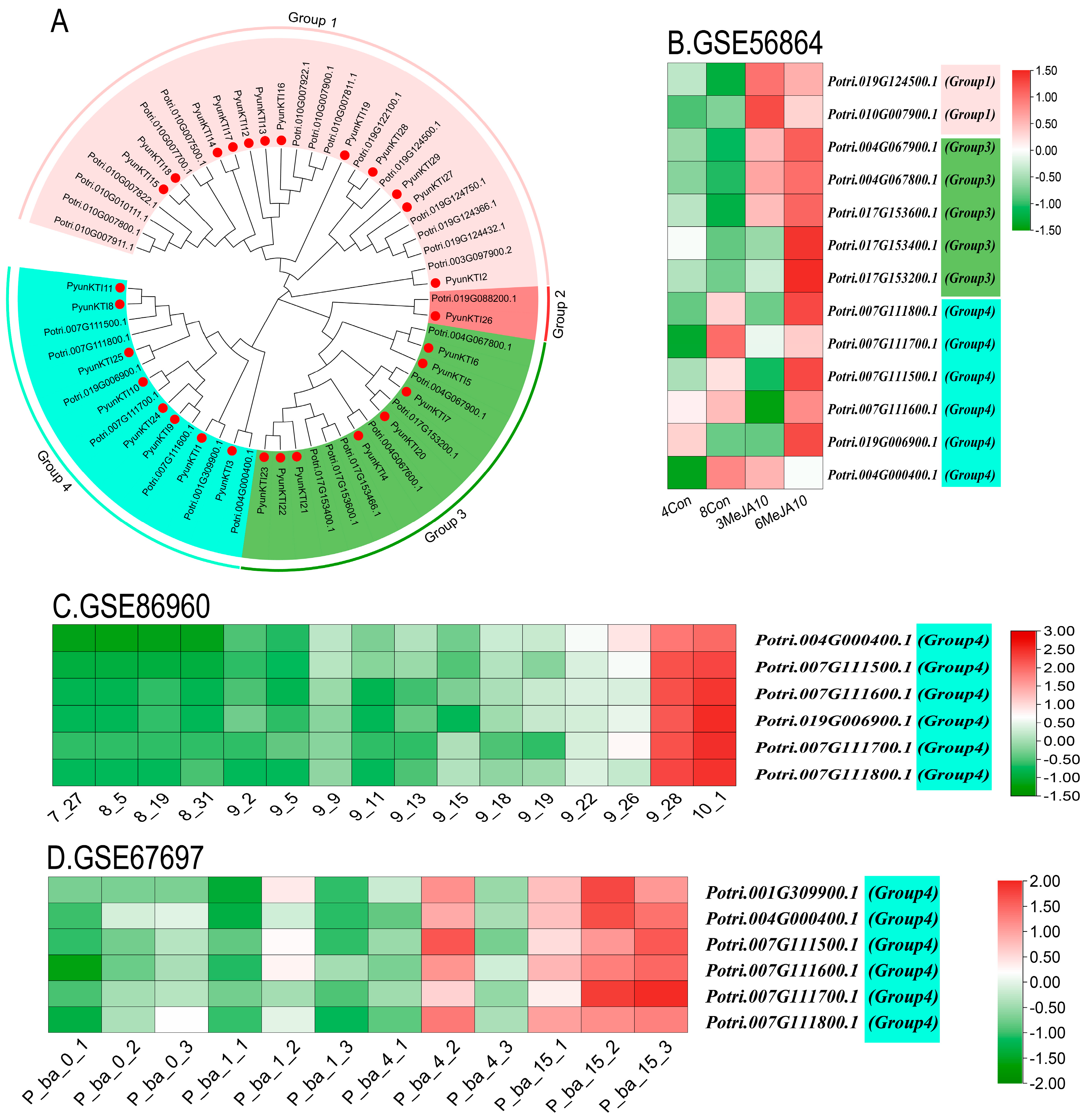
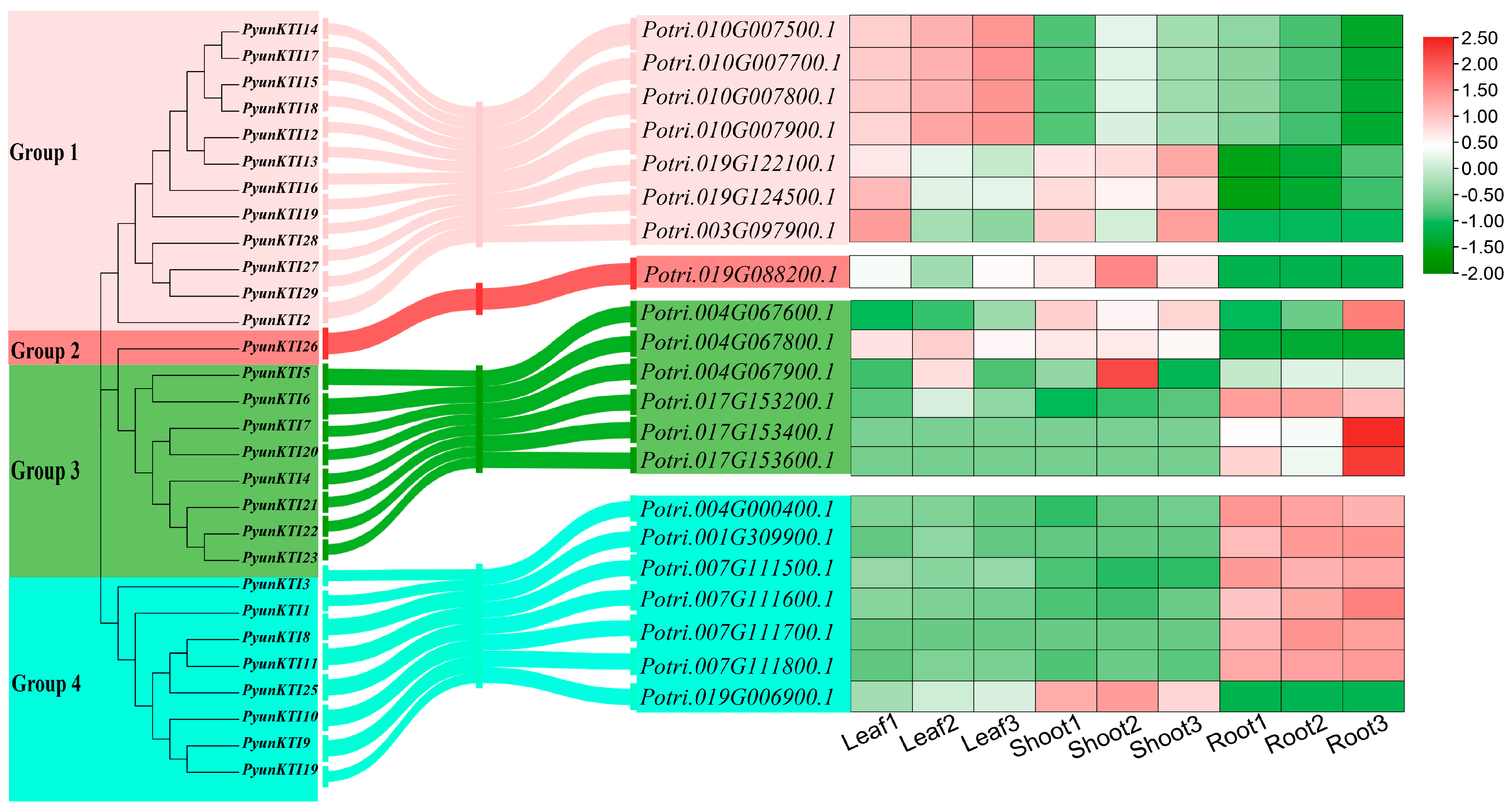
2.1. Identification and Phylogenetic Tree of the PyunKTI Genes Family in Populus
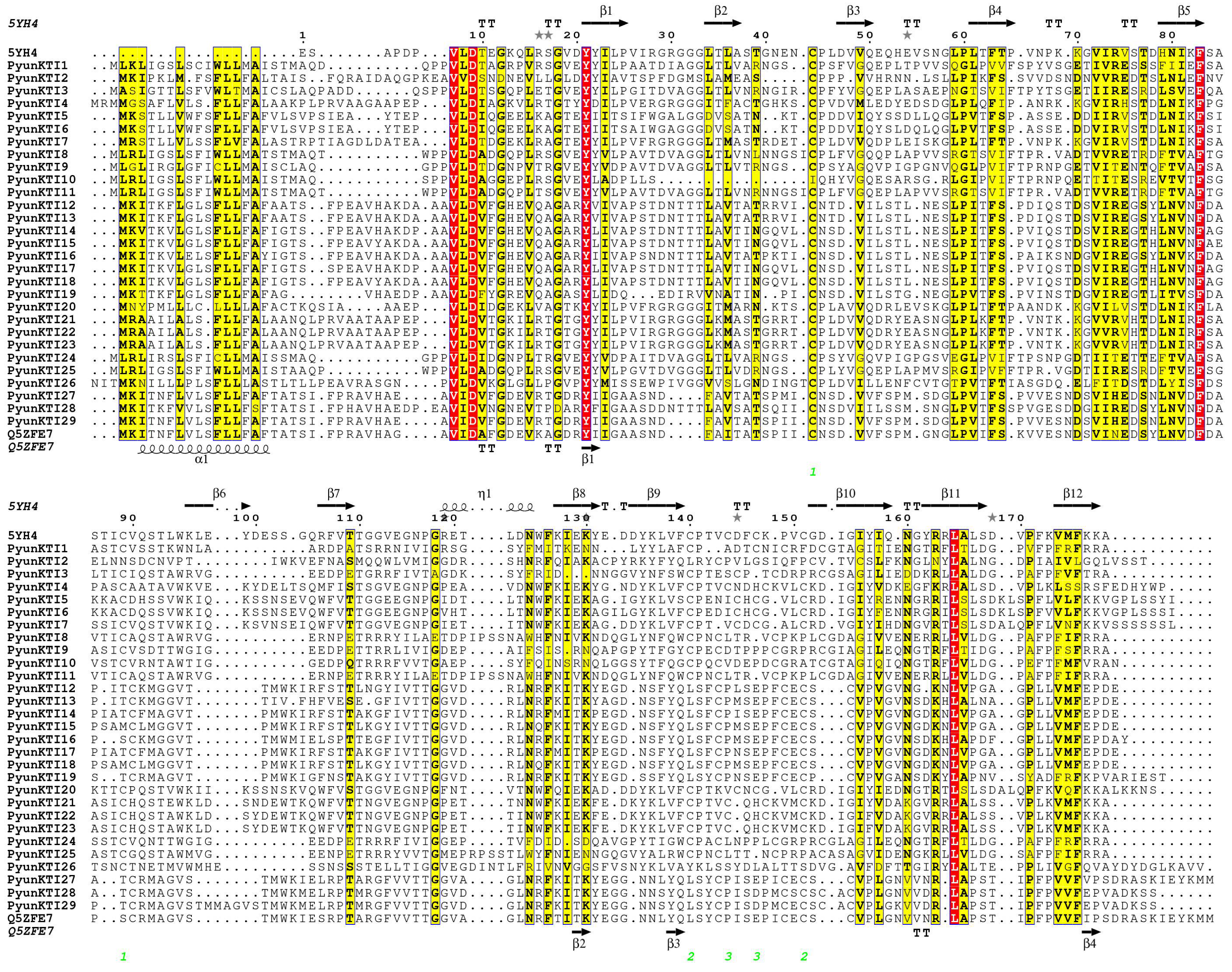
2.2. Characterization of the PyunKTIs Gene Family in P. yunnanensis
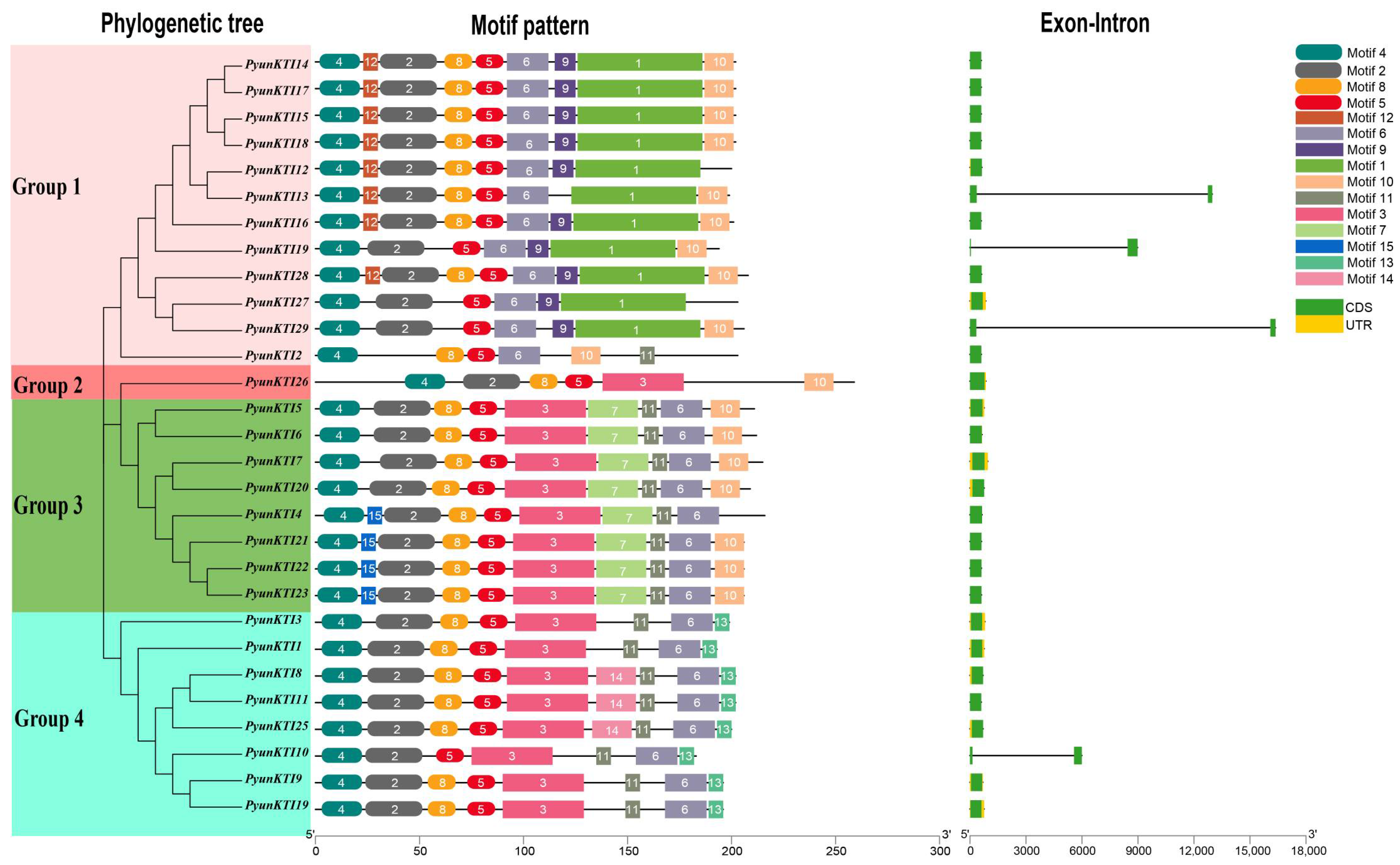
2.3. Gene Structures and Conserved Motifs of the PyunKTI Genes Family
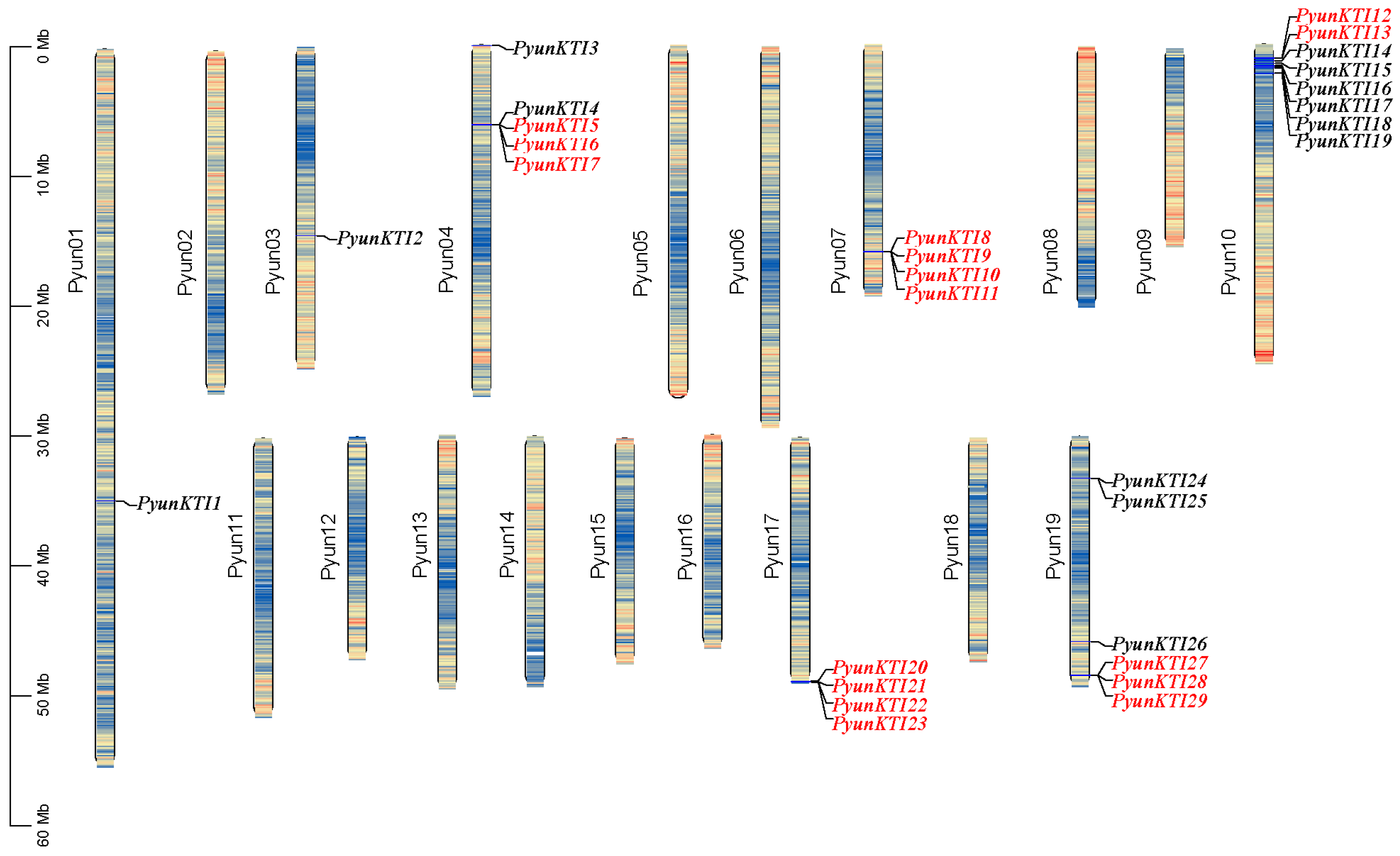
2.4. Chromosomal Location and Duplication of the PyunKTI Gene Family
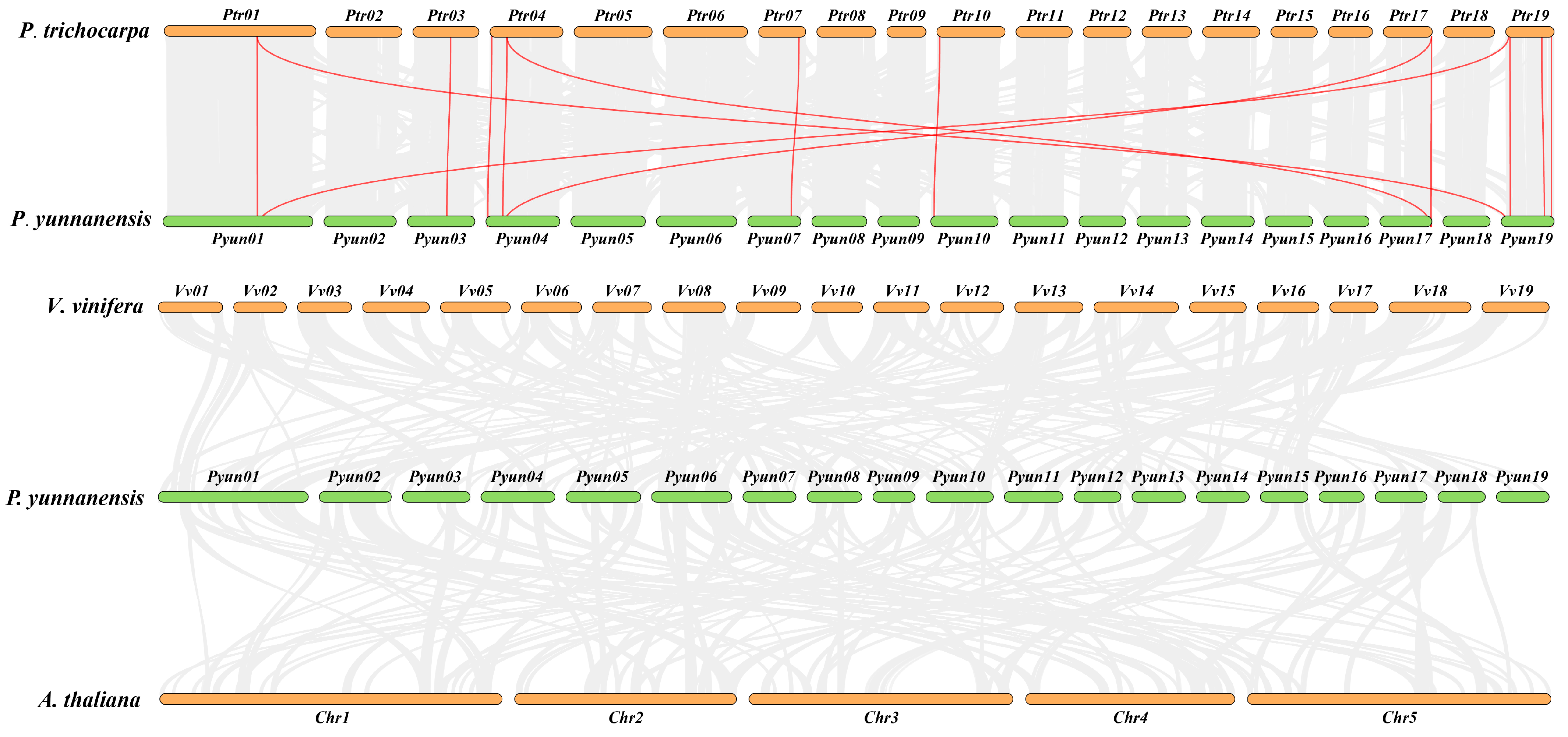
2.5. Tandem Repeat and Synteny Gene Pairs
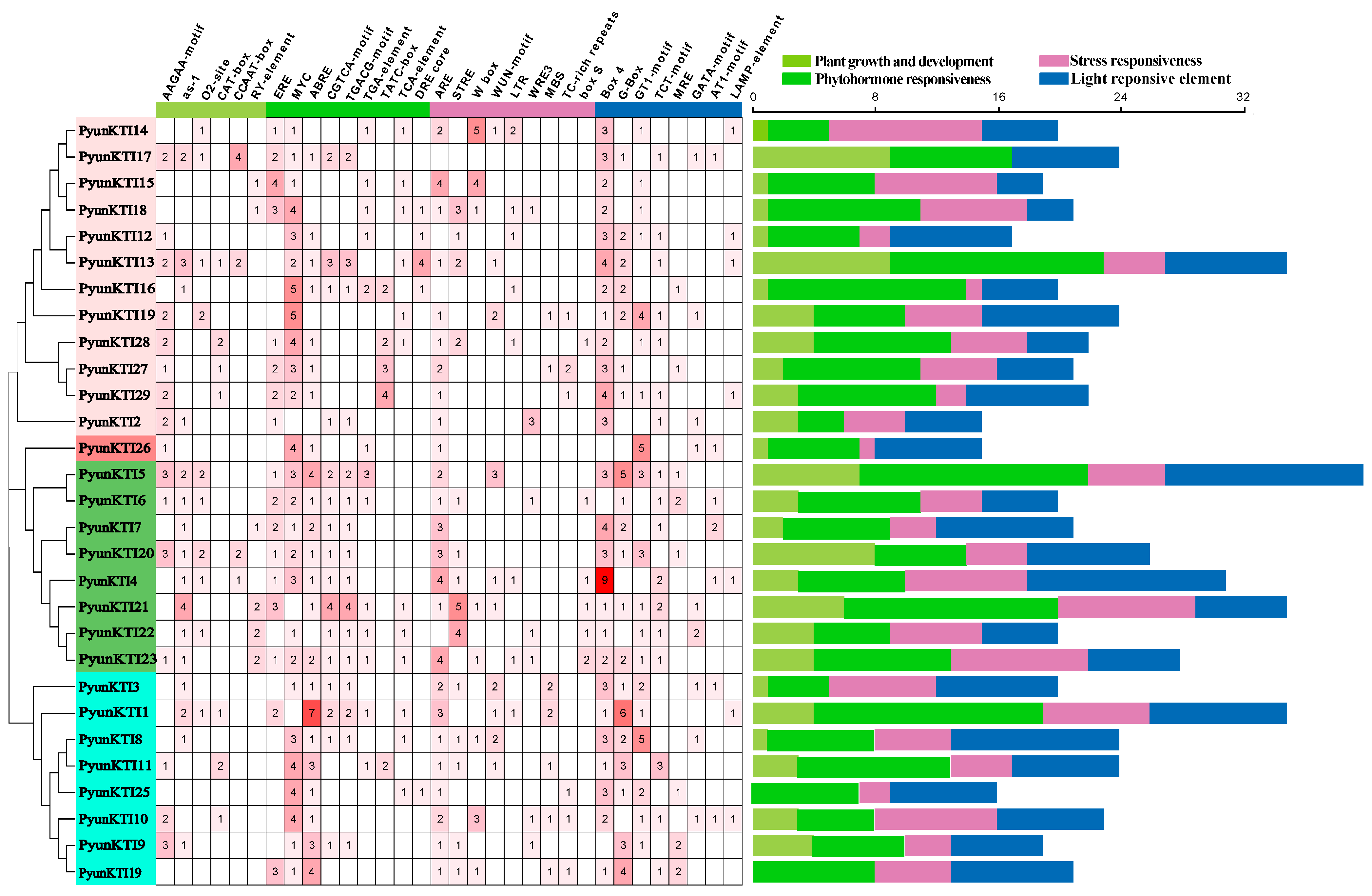
2.6. Promoter Analysis of the PyunKTIs Genes
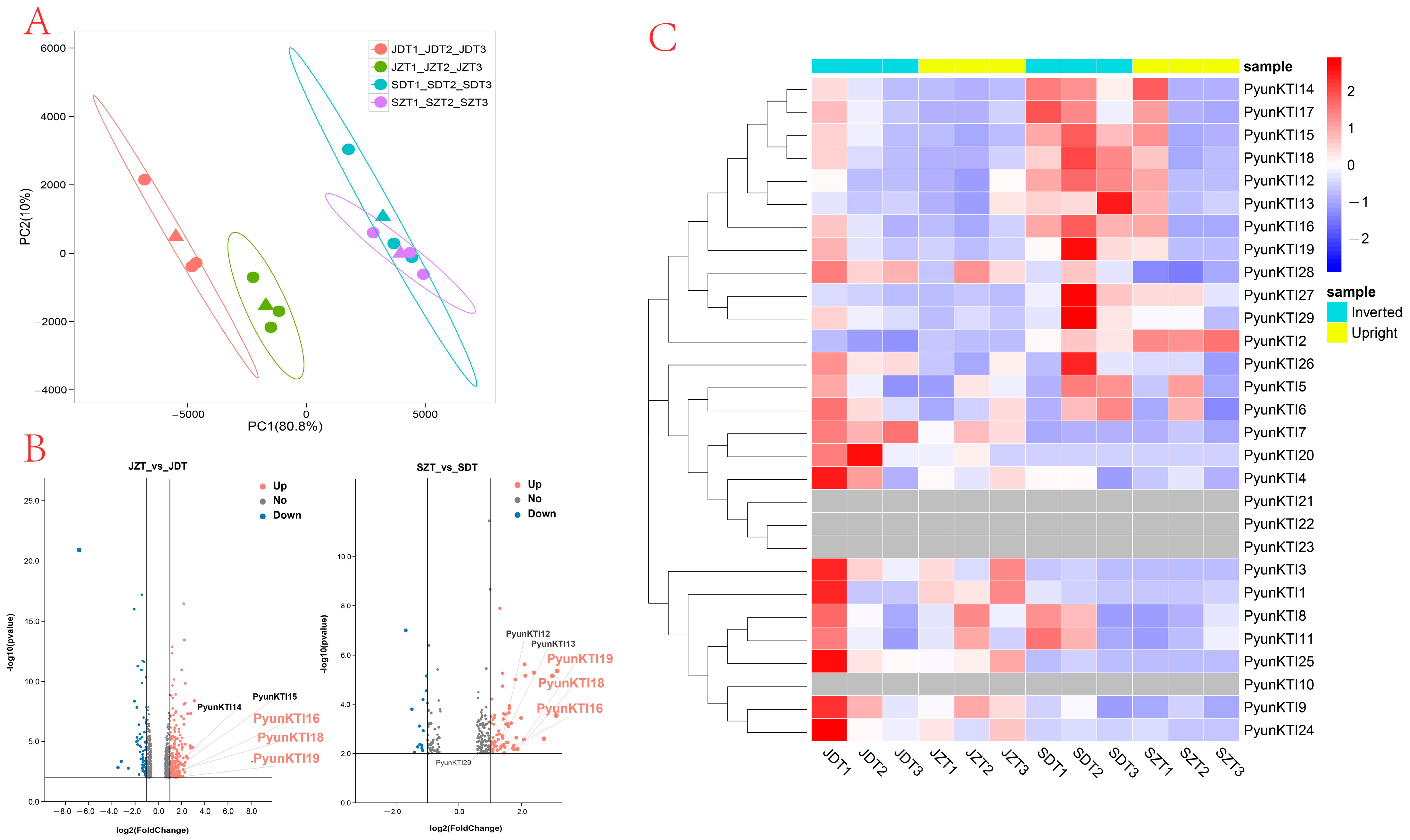
2.7. Expression Profile of PyunKTI Genes from P. yunnanensis with Upright and Inverted Cuttings
2.8. RT-qPCR Analysis of the PyunKTI Genes Expression Level
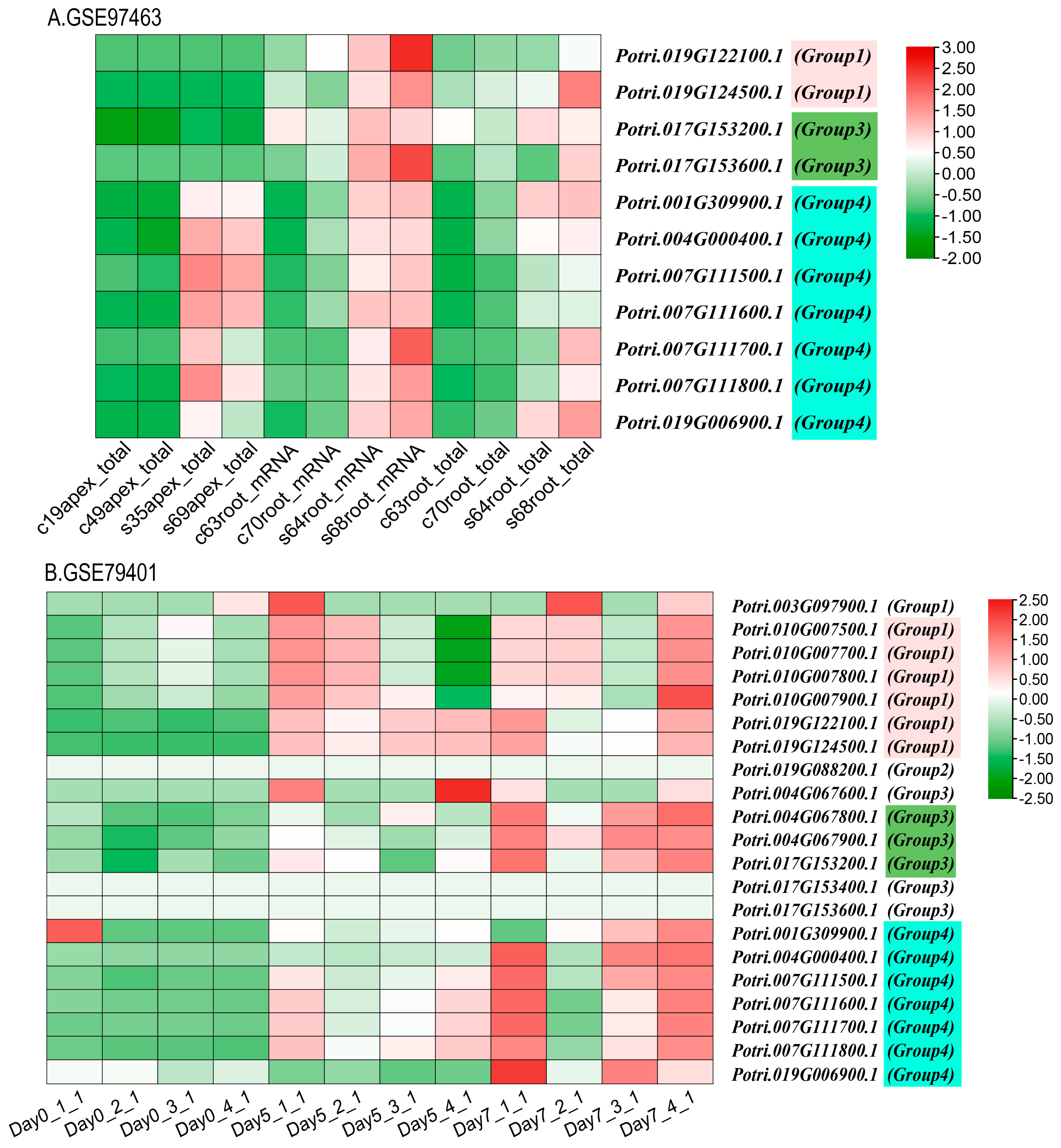
2.9. Transcriptome Analysis in GEO Database of PyunKTI Genes
3. Discussion
3.1. Conserved Fundamental Features and Diverse Expression Patterns of KTIs
3.2. Involved in Plant Growth and Development
3.3. Involvement in Biological and Environmental Stresses
4. Materials and Methods
4.1. Identification and Characterization of the PyunKTI Gene Family
4.2. Analyses of Phylogenetics, Gene Structure, Motif Composition, Chromosomal Distribution, Gene Duplication, and Synteny
4.3. Identification of Cis-Regulatory Elements (CREs) in PyunKTIs Gene Promoters
4.4. Transcriptome Analysis About PyunKTI Genes
4.5. The Expression Profiles of PyunKTI Genes Were Analyzed Using the Available Transcriptome Datasets
4.6. RT-qPCR Analysis of the PyunKTI Genes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bateman, K.S.; Michael, N.G.J. Plant Protein Proteinase Inhibitors: Structure and Mechanism of Inhibition. Curr. Protein Pept. Sci. 2011, 12, 341–347. [Google Scholar] [CrossRef]
- Turk, B. Targeting proteases: Successes, failures and future prospects. Nat. Rev. Drug Discov. 2006, 5, 785–799. [Google Scholar] [CrossRef] [PubMed]
- Rawlings, N.D.; Tolle, D.P.; Barrett, A.J. Evolutionary families of peptidase inhibitors. Biochem. J. 2004, 378, 705–716. [Google Scholar] [CrossRef]
- Rawlings, N.D.; Barrett, A.J.; Alex, B. MEROPS: The database of proteolytic enzymes, their substrates and inhibitors. Nucleic Acids Res. 2012, 40, D343–D350. [Google Scholar] [CrossRef]
- Rawlings, N.D.; Waller, M.; Barrett, A.J.; Bateman, A. MEROPS: The database of proteolytic enzymes, their substrates and inhibitors. Nucleic Acids Res. 2013, 42, D503–D509. [Google Scholar] [CrossRef] [PubMed]
- Kunitz, M. Crystallization of a Trypsin Inhibitor from Soybean. Science 1945, 101, 668–669. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.X.; Ng, T.B. Concurrent isolation of a Kunitz-type trypsin inhibitor with antifungal activity and a novel lectin from Pseudostellaria heterophylla roots. Biochem. Biophys. Res. Commun. 2006, 342, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Bártová, V.; Bárta, J.; Jarošová, M. Antifungal and antimicrobial proteins and peptides of potato (Solanum tuberosum L.) tubers and their applications. Appl. Microbiol. Biotechnol. 2019, 103, 5533–5547. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Brader, G.; Palva, E.T. Kunitz Trypsin Inhibitor: An Antagonist of Cell Death Triggered by Phytopathogens and Fumonisin B1 in Arabidopsis. Mol. Plant 2008, 1, 482–495. [Google Scholar] [CrossRef]
- Huang, H.; Qi, S.D.; Qi, F.; Wu, C.A.; Zheng, C.C. NtKTI1, a Kunitz trypsin inhibitor with antifungal activity from Nicotiana tabacum, plays an important role in tobacco’s defense response. FEBS J. 2010, 277, 4076–4088. [Google Scholar] [CrossRef]
- Ana, A.; Lucia, T.M.; Pablo, G.M.; Manuel, M.; Isabel, D.; Santamaria, M.E. Arabidopsis Kunitz Trypsin Inhibitors in Defense Against Spider Mites. Front. Plant Sci. 2018, 9, 986. [Google Scholar]
- Botelho-Júnior, S.; Machado, O.L.T.; Fernandes, K.V.S.; Lemos, F.J.A.; Perdizio, V.A.; Oliveira, A.N.E.A.; Monteiro, L.R.; Filho, M.L.; Jacinto, T. Defense response in non-genomic model species: Methyl jasmonate exposure reveals the passion fruit leaves’ ability to assemble a cocktail of functionally diversified Kunitz-type trypsin inhibitors and recruit two of them against papain. Planta 2014, 240, 345–356. [Google Scholar] [CrossRef]
- Jiang, H.; Korpelainen, H.; Li, C. Populus yunnanensis males adopt more efficient protective strategies than females to cope with excess zinc and acid rain. Chemosphere 2013, 91, 1213–1220. [Google Scholar] [CrossRef]
- Li, X.; Yang, Y.; Sun, X.; Lin, H.; Chen, J.; Ren, J.; Hu, X.; Yang, Y. Comparative Physiological and Proteomic Analyses of Poplar (Populus yunnanensis) Plantlets Exposed to High Temperature and Drought. PLoS ONE 2014, 9, e107605. [Google Scholar] [CrossRef] [PubMed]
- Zhou, A.-P.; Zong, D.; Gan, P.-H.; Zou, X.-L.; Fei, X.; Zhong, Y.-Y.; He, C.-Z. Physiological analysis and transcriptome profiling of inverted cuttings of Populus yunnanensis reveal that cell wall metabolism plays a crucial role in responding to inversion. Genes 2018, 9, 572. [Google Scholar] [CrossRef]
- Zhou, A.P.; Gan, P.H.; Zong, D.; Fei, X.; Zhong, Y.Y.; Li, S.Q.; Yu, J.-D.; He, C.Z. Bark tissue transcriptome analyses of inverted Populus yunnanensis cuttings reveal the crucial role of plant hormones in response to inversion. PeerJ 2019, 7, e7740. [Google Scholar] [CrossRef]
- Mbaye, M.N.; Hou, Q.; Basu, S.; Teheux, F.; Rooman, M. A Comprehensive Computational Study of Amino Acid Interactions in Membrane Proteins. Sci. Rep. 2019, 9, 12043. [Google Scholar] [CrossRef]
- Wang, P.; Moore, B.M.; Panchy, N.L.; Meng, F.; Lehti-Shiu, M.D.; Shiu, S.H. Factors Influencing Gene Family Size Variation Among Related Species in a Plant Family, Solanaceae. Genome Biol. Evol. 2018, 10, 2596–2613. [Google Scholar] [CrossRef] [PubMed]
- Kong, H.; Landherr, L.L.; Frohlich, M.W.; Leebens-Mack, J.; Depamphilis, C.W. Patterns of gene duplication in the plant SKP1 gene family in angiosperms: Evidence for multiple mechanisms of rapid gene birth. Plant J. 2010, 50, 873–885. [Google Scholar] [CrossRef] [PubMed]
- Bendre, A.D.; Ramasamy, S.; Suresh, C.G. Analysis of Kunitz inhibitors from plants for comprehensive structural and functional insights. Int. J. Biol. Macromol. 2018, 113, S0141813018303222. [Google Scholar] [CrossRef] [PubMed]
- Oliva, M.; Silva, M.; Sallai, R.C.; Brito, M.V.; Sampaio, M.U. A novel subclassification for Kunitz proteinase inhibitors from leguminous seeds. Biochimie 2010, 92, 1667–1673. [Google Scholar] [CrossRef]
- Kim, S.H.; Hara, S.; Hase, S.; Ikenaka, T.; Toda, H.; Kitamura, K.; Kaizuma, N. Comparative study on amino acid sequences of Kunitz-type soybean trypsin inhibitors, Tia, Tib, and Tic. J. Biochem. 1985, 98, 435–448. [Google Scholar] [CrossRef]
- Kobayashi, H. A Kunitz-type Protease Inhibitor, Bikunin, Inhibits Ovarian Cancer Cell Invasion by Blocking the Calcium-dependent Transforming Growth Factor-β1 Signaling Cascade. J. Biol. Chem. 2003, 278, 7790–7799. [Google Scholar] [CrossRef]
- Zhang, X.; Liao, D.-Y.; Gou, X.-Z.; Zhou, J.-Y.; Liao, H. Structure and function of plant Kunitz-type protease inhibitors. Plant Physiol. J. 2018, 54, 1391–1400. [Google Scholar]
- Stegmaier, P.; Kel, A.E.; Wingender, E. Systematic DNA-binding domain classification of transcription factors. Genome Inform. Int. Conf. Genome Inform. 2004, 15, 276–286. [Google Scholar]
- Luciński, R.; Misztal, L.; Samardakiewicz, S.A.; Jackowski, G. Involvement of Deg5 protease in wounding-related disposal of PsbF apoprotein. Plant Physiol. Biochem. 2011, 49, 311–320. [Google Scholar] [CrossRef]
- Budi, M.K.; Saboti, J.; Megli, V.; Kos, J.; Kidri, M. Characterization of two novel subtilases from common bean (Phaseolus vulgaris L.) and their responses to drought. Plant Physiol. Biochem. 2013, 62, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Islam, A.; Leung, S.; Burgess, E.P.; Laing, W.A.; Richardson, K.A.; Hofmann, R.W.; Dijkwel, P.P.; McManus, M.T. Knock-down of transcript abundance of a family of Kunitz proteinase inhibitor genes in white clover (Trifolium repens) reveals a redundancy and diversity of gene function. New Phytol. 2015, 208, 1188–1201. [Google Scholar] [CrossRef]
- Boex-Fontvieille, E.; Rustgi, S.; Von Wettstein, D.; Pollmann, S.; Reinbothe, S.; Reinbothe, C. Corrigendum: An Ethylene-Protected Achilles’ Heel of Etiolated Seedlings for Arthropod Deterrence. Front. Plant Sci. 2018, 9, 1741. [Google Scholar] [CrossRef]
- Edouard, B.F.; Sachin, R.; Steffen, R.; Christiane, R. A Kunitz-type protease inhibitor regulates programmed cell death during flower development in Arabidopsis thaliana. J. Exp. Bot. 2015, 66, 6119–6135. [Google Scholar]
- Srinivasan, T.; Kumar, K.R.R.; Kirti, P.B. Constitutive Expression of a Trypsin Protease Inhibitor Confers Multiple Stress Tolerance in Transgenic Tobacco. Plant Cell Physiol. 2009, 50, 541–553. [Google Scholar] [CrossRef] [PubMed]
- Afsana, I.; Susanna, L.; Aluh, N.; Dijkwel, P.P.; Mcmanus, M.T. Kunitz Proteinase Inhibitors Limit Water Stress Responses in White Clover (Trifolium repens L.) Plants. Front. Plant Sci. 2017, 8, 1683. [Google Scholar]
- Komatsu, S.; Yamamoto, R.; Nanjo, Y.; Mikami, Y.; Yunokawa, H.; Sakata, K. A comprehensive analysis of the soybean genes and proteins expressed under flooding stress using transcriptome and proteome techniques. J. Proteome Res. 2009, 8, 4766–4778. [Google Scholar] [CrossRef]
- Kang, S.G.; Choi, J.H.; Suh, S.G. A Leaf-specific 27 kDa Protein of Potato Kunitz-Type Proteinase Inhibitor Is Induced in Response to Abscisic Acid, Ethylene, Methyl Jasmonate, and Water Deficit. Mol. Cells 2002, 13, 144–147. [Google Scholar] [CrossRef]
- Sheshukova, E.V.; Komarova, T.V.; Ershova, N.M.; Shindyapina, A.V.; Dorokhov, Y.L. An Alternative Nested Reading Frame May Participate in the Stress-Dependent Expression of a Plant Gene. Front. Plant Sci. 2017, 8, 2137. [Google Scholar] [CrossRef]
- Wang, H.; Xu, C.; Zhang, Y.; Yan, X.; Zheng, B. PtKTI12 genes influence wobble uridine modifications and drought stress tolerance in hybrid poplar. Tree Physiol. 2020, 40, 1778–1791. [Google Scholar] [CrossRef]
- Liu, Z.; Zhu, Q.; Li, Y.; Yu, J.; Wang, W.; Tan, R.; Zhou, J.; Liao, H. Isolation and in silico characterization of a shikimate kinase from Cassia obtusifolia. Acta Physiol. Plant. 2015, 37, 85. [Google Scholar] [CrossRef]
- Sara, E.G.; Jaina, M.; Alex, B.; Eddy, S.R.; Aurélien, L.; Potter, S.C.; Matloob, Q.; Richardson, L.J.; Salazar, G.A.; Alfredo, S. The Pfam protein families database in 2019. Nuclc Acids Res. 2018, 47, D427–D432. [Google Scholar]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, 29–37. [Google Scholar] [CrossRef]
- Aron, M.B.; Bo, Y.; Han, L.; He, J.; Lanczycki, C.J.; Lu, S.; Farideh, C.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R. CDD/SPARCLE: Functional classification of proteins via subfamily domain architectures. Nucleic Acids Res. 2017, 45, D200–D203. [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Multiple Sequence Alignment Using ClustalW and ClustalX. Curr. Protoc. Bioinform. 2003, 1, 2–3. [Google Scholar] [CrossRef]
- Koichiro, T.; Glen, S.; Sudhir, K. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Subramanian, B.; Gao, S.; Lercher, M.J.; Hu, S.; Chen, W.H. Evolview v3: A webserver for visualization, annotation, and management of phylogenetic trees. Nuclc Acids Res. 2019, 47, W270–W275. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Mikael, B.; Buske, F.A.; Martin, F.; Grant, C.E.; Luca, C.; Jingyuan, R.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 9. [Google Scholar] [CrossRef]
- Yupeng, W.; Haibao, T.; Debarry, J.D.; Xu, T.; Jingping, L.; Xiyin, W.; Tae-Ho, L.; Huizhe, J.; Barry, M.; Hui, G. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar]
- Wang, D.; Zhang, Y.; Zhang, Z.; Zhu, J.; Yu, J. KaKs_Calculator 2.0: A toolkit incorporating gamma-series methods and sliding window strategies. Genom. Proteom. Bioinform. 2010, 8, 77–80. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Shi, R.; Wang, J.P.; Lin, Y.C.; Li, Q.; Sun, Y.H.; Chen, H.; Sederoff, R.R.; Chiang, V.L. Tissue and cell-type co-expression networks of transcription factors and wood component genes in Populus trichocarpa. Planta 2017, 245, 927–938. [Google Scholar] [CrossRef]
- Plett, J.M.; Daguerre, Y.; Wittulsky, S.; Vayssières, A.; Martin, F. Effector MiSSP7 of the mutualistic fungus Laccaria bicolor stabilizes the Populus JAZ6 protein and represses jasmonic acid (JA) responsive genes. Proc. Natl. Acad. Sci. USA 2014, 111, 8299–8304. [Google Scholar] [CrossRef] [PubMed]
- Edlund, E.; Novak, O.; Karady, M.; Ljung, K.; Jansson, S. Contrasting patterns of cytokinins between years in senescing aspen leaves. Plant Cell Environ. 2017, 40, 622. [Google Scholar] [CrossRef] [PubMed]
- Foster, A.J.; Gervais, P.; Philippe, T.; Armand, S.; Martina, S. Transcriptome Analysis of Poplar during Leaf Spot Infection with Sphaerulina spp. PLoS ONE 2015, 10, e0138162. [Google Scholar] [CrossRef]
- Yinan, Y.; Su, C. Widespread antisense transcription of Populus genome under drought. Mol. Genet. Genom. 2018, 293, 1017–1033. [Google Scholar]
- Li, S.; Lin, Y.C.J.; Wang, P.; Zhang, B.; Li, M.; Chen, S.; Shi, R.; Tunlaya-Anukit, S.; Liu, X.; Wang, Z.; et al. The AREB1 Transcription Factor Influences Histone Acetylation to Regulate Drought Responses and Tolerance in Populus trichocarpa. Plant Cell 2019, 31, 663–686. [Google Scholar] [CrossRef]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinform. 2012, 13, 134. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data using Real-Time Quantitative PCR. Methods 2002, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Genomics Position | Amino AcidsNumber(aa) | Molecular Formula | Isoelectric Point | Molecular Weight | Instability Index | Aliphatic Index | GRAVY | Subcellular Localization |
|---|---|---|---|---|---|---|---|---|---|---|
| PyunKTI1 | Pyun01G029930 | Chr01: 34845724-34846485 | 193 | C933H1468N248O273S11 | 8.18 | 20,880.08 | 61.68 | 89.95 | 0.25 | Vacuole. |
| PyunKTI2 | Pyun03G009060 | Chr03: 14540624-14541235 | 203 | C1020H1588N262O295S12 | 5.33 | 22,626.13 | 32.53 | 98.87 | 0.16 | Vacuole. |
| PyunKTI3 | Pyun04G000040 | Chr04: 55635-56439 | 199 | C958H1482N252O299S9 | 4.41 | 21,602.35 | 46.52 | 81.36 | −0.05 | Vacuole. |
| PyunKTI4 | Pyun04G005750 | Chr04: 6150656-6151306 | 216 | C1075H1670N278O307S12 | 6.43 | 23,785.48 | 41.02 | 84.86 | −0.07 | Vacuole. |
| PyunKTI5 | Pyun04G005770 | Chr04: 6159791-6160553 | 211 | C1058H1648N262O315S7 | 5.09 | 23,302.71 | 42.68 | 98.34 | 0.08 | Vacuole. |
| PyunKTI6 | Pyun04G005780 | Chr04: 6172938-6173586 | 212 | C1057H1643N261O315S7 | 4.96 | 23,271.65 | 42.54 | 96.93 | 0.11 | Vacuole. |
| PyunKTI7 | Pyun04G005790 | Chr04: 6182198-6183153 | 215 | C1039H1659N271O318S8 | 5.48 | 23,291.71 | 35.00 | 95.58 | 0.09 | Vacuole. |
| PyunKTI8 | Pyun07G011750 | Chr07: 15976464-15977166 | 202 | C1006H1578N276O282S9 | 8.27 | 22,339.82 | 43.52 | 95.50 | 0.13 | Vacuole. |
| PyunKTI9 | Pyun07G011760 | Chr07: 15991434-15992125 | 196 | C920H1436N246O278S10 | 4.58 | 20,711.60 | 40.41 | 83.06 | 0.09 | Vacuole. |
| PyunKTI10 | Pyun07G011770 | Chr07: 15993379-15999370 | 183 | C879H1380N246O268S9 | 5.41 | 19,970.65 | 48.54 | 79.34 | −0.09 | Vacuole. |
| PyunKTI11 | Pyun07G011780 | Chr07: 16001371-16001979 | 202 | C1004H1573N275O284S9 | 7.58 | 22,328.75 | 41.01 | 93.56 | 0.11 | Vacuole. |
| PyunKTI12 | Pyun10G000910 | Chr10: 1058936-1059577 | 200 | C975H1523N247O291S10 | 5.15 | 21,681.90 | 30.23 | 89.65 | 0.23 | Vacuole. |
| PyunKTI13 | Pyun10G000920 | Chr10: 1070920-1083898 | 199 | C969H1504N244O291S10 | 4.86 | 21,548.66 | 36.54 | 90.60 | 0.30 | Vacuole. |
| PyunKTI14 | Pyun10G000940 | Chr10: 1273093-1273701 | 202 | C975H1525N245O287S11 | 4.94 | 21,623.96 | 26.31 | 93.07 | 0.35 | Vacuole. |
| PyunKTI15 | Pyun10G000990 | Chr10: 1485338-1485946 | 202 | C980H1532N242O293S12 | 4.72 | 21,777.11 | 27.63 | 93.07 | 0.30 | Vacuole. |
| PyunKTI16 | Pyun10G001030 | Chr10: 1649193-1649798 | 201 | C982H1523N241O296S12 | 4.79 | 21,826.05 | 38.14 | 86.32 | 0.17 | Vacuole. |
| PyunKTI17 | Pyun10G001050 | Chr10: 1730499-1731107 | 202 | C968H1518N246O289S11 | 4.94 | 21,578.84 | 28.91 | 91.63 | 0.30 | Vacuole. |
| PyunKTI18 | Pyun10G001070 | Chr10: 1821744-1822352 | 202 | C977H1528N242O291S12 | 4.81 | 21,705.05 | 26.67 | 93.07 | 0.31 | Vacuole. |
| PyunKTI19 | Pyun10G001220 | Chr10: 2244406-2253386 | 194 | C946H1458N242O287S9 | 5.00 | 21,101.97 | 30.27 | 81.39 | 0.06 | Vacuole. |
| PyunKTI20 | Pyun17G015360 | Chr17: 18745402-18746164 | 209 | C1033H1660N268O295S11 | 9.17 | 22,906.82 | 36.39 | 95.60 | 0.06 | Vacuole. |
| PyunKTI21 | Pyun17G015370 | Chr17: 18797532-18798152 | 206 | C1027H1641N283O285S10 | 9.59 | 22,833.65 | 27.43 | 88.01 | −0.10 | Vacuole. |
| PyunKTI22 | Pyun17G015380 | Chr17: 18822863-18823483 | 206 | C1032H1647N281O281S10 | 9.67 | 22,807.74 | 29.10 | 90.39 | 0.00 | Vacuole. |
| PyunKTI23 | Pyun17G015390 | Chr17: 18827204-18827824 | 206 | C1032H1647N281O281S10 | 9.67 | 22,807.74 | 29.10 | 90.39 | 0.00 | Vacuole. |
| PyunKTI24 | Pyun19G002480 | Chr19: 3226565-3227324 | 196 | C931H1460N248O279S10 | 4.60 | 20,911.93 | 48.91 | 88.52 | 0.15 | Vacuole. |
| PyunKTI25 | Pyun19G002500 | Chr19: 3248588-3249293 | 200 | C979H1519N267O283S11 | 5.53 | 21,910.11 | 57.69 | 83.80 | 0.01 | Vacuole. |
| PyunKTI26 | Pyun19G010740 | Chr19: 15803407-15804259 | 259 | C1262H1993N309O384S9 | 4.71 | 27,927.09 | 32.92 | 107.92 | 0.33 | Vacuole. |
| PyunKTI27 | Pyun19G013290 | Chr19: 18393576-18394424 | 203 | C989H1543N253O292S13 | 4.99 | 22,066.43 | 39.30 | 90.69 | 0.31 | Vacuole. |
| PyunKTI28 | Pyun19G013300 | Chr19: 18397269-18397895 | 208 | C984H1537N253O310S14 | 4.54 | 22,320.38 | 33.11 | 83.37 | 0.18 | Vacuole. |
| PyunKTI29 | Pyun19G013310 | Chr19: 18402299-18418665 | 206 | C987H1539N251O299S16 | 4.64 | 22,218.54 | 35.10 | 84.61 | 0.29 | Vacuole. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, H.; Ma, S.; Zhang, X.; Xu, R.; Wang, C.; Zhang, S.; Zhao, L.; Li, D.; Zong, D. Identification of Kunitz-Type Inhibitor Gene Family of Populus yunnanensis Reveals a Stress Tolerance Function in Inverted Cuttings. Int. J. Mol. Sci. 2025, 26, 188. https://doi.org/10.3390/ijms26010188
Guo H, Ma S, Zhang X, Xu R, Wang C, Zhang S, Zhao L, Li D, Zong D. Identification of Kunitz-Type Inhibitor Gene Family of Populus yunnanensis Reveals a Stress Tolerance Function in Inverted Cuttings. International Journal of Molecular Sciences. 2025; 26(1):188. https://doi.org/10.3390/ijms26010188
Chicago/Turabian StyleGuo, Haiyang, Shaojie Ma, Xiaolin Zhang, Rong Xu, Cai Wang, Shihai Zhang, Lihong Zhao, Dan Li, and Dan Zong. 2025. "Identification of Kunitz-Type Inhibitor Gene Family of Populus yunnanensis Reveals a Stress Tolerance Function in Inverted Cuttings" International Journal of Molecular Sciences 26, no. 1: 188. https://doi.org/10.3390/ijms26010188
APA StyleGuo, H., Ma, S., Zhang, X., Xu, R., Wang, C., Zhang, S., Zhao, L., Li, D., & Zong, D. (2025). Identification of Kunitz-Type Inhibitor Gene Family of Populus yunnanensis Reveals a Stress Tolerance Function in Inverted Cuttings. International Journal of Molecular Sciences, 26(1), 188. https://doi.org/10.3390/ijms26010188