
Nontraditional Roles of Magnesium Ions in Modulating Sav2152: Insight from a Haloacid Dehalogenase-like Superfamily Phosphatase from Staphylococcus aureus

 , , , and
, , , and
Abstract
1. Introduction
2. Results
2.1. Oligomeric State of Sav2152
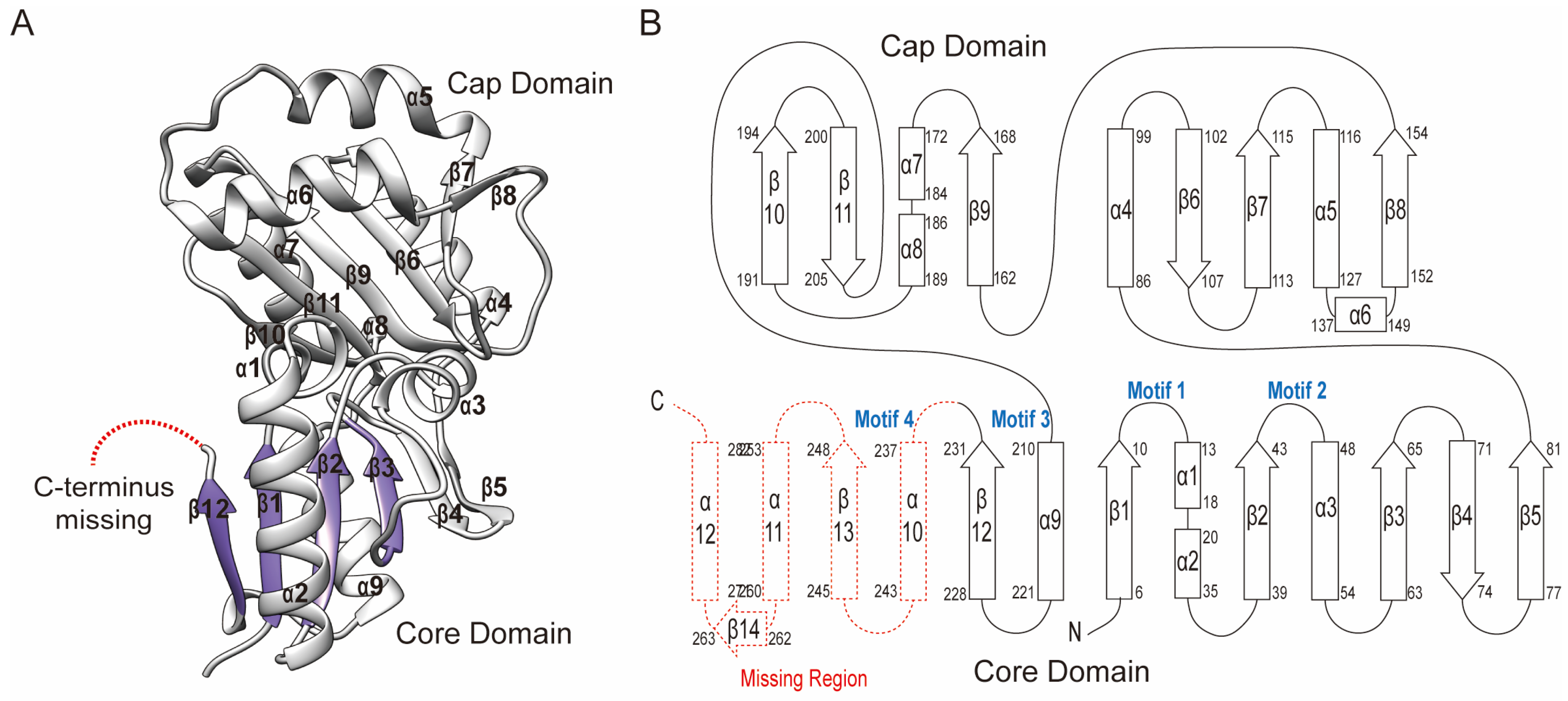
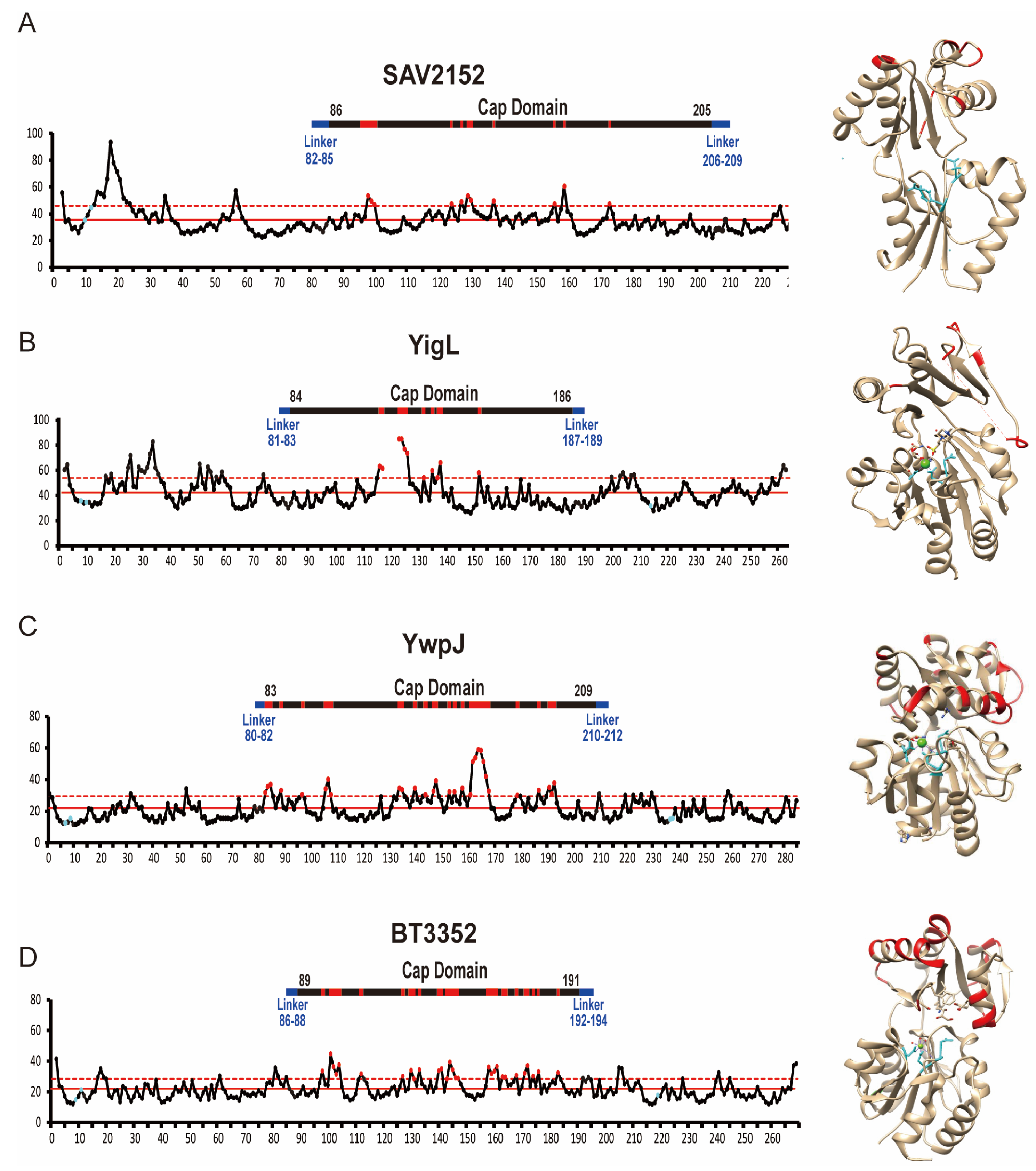
2.2. Overall Crystal Structure of Sav2152
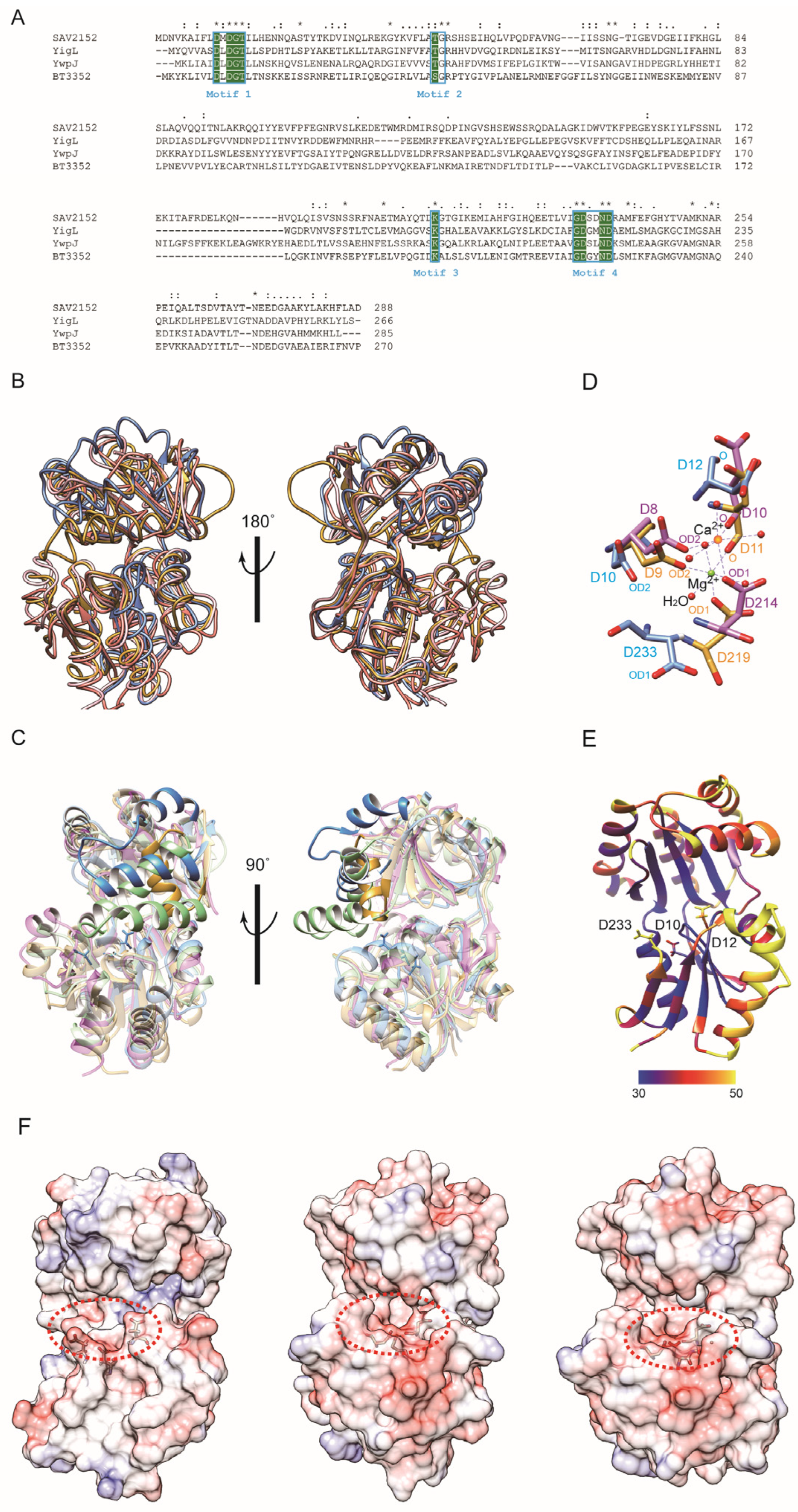
2.3. Structural Comparison with Homology
2.4. Enzyme Activity of Sav2152
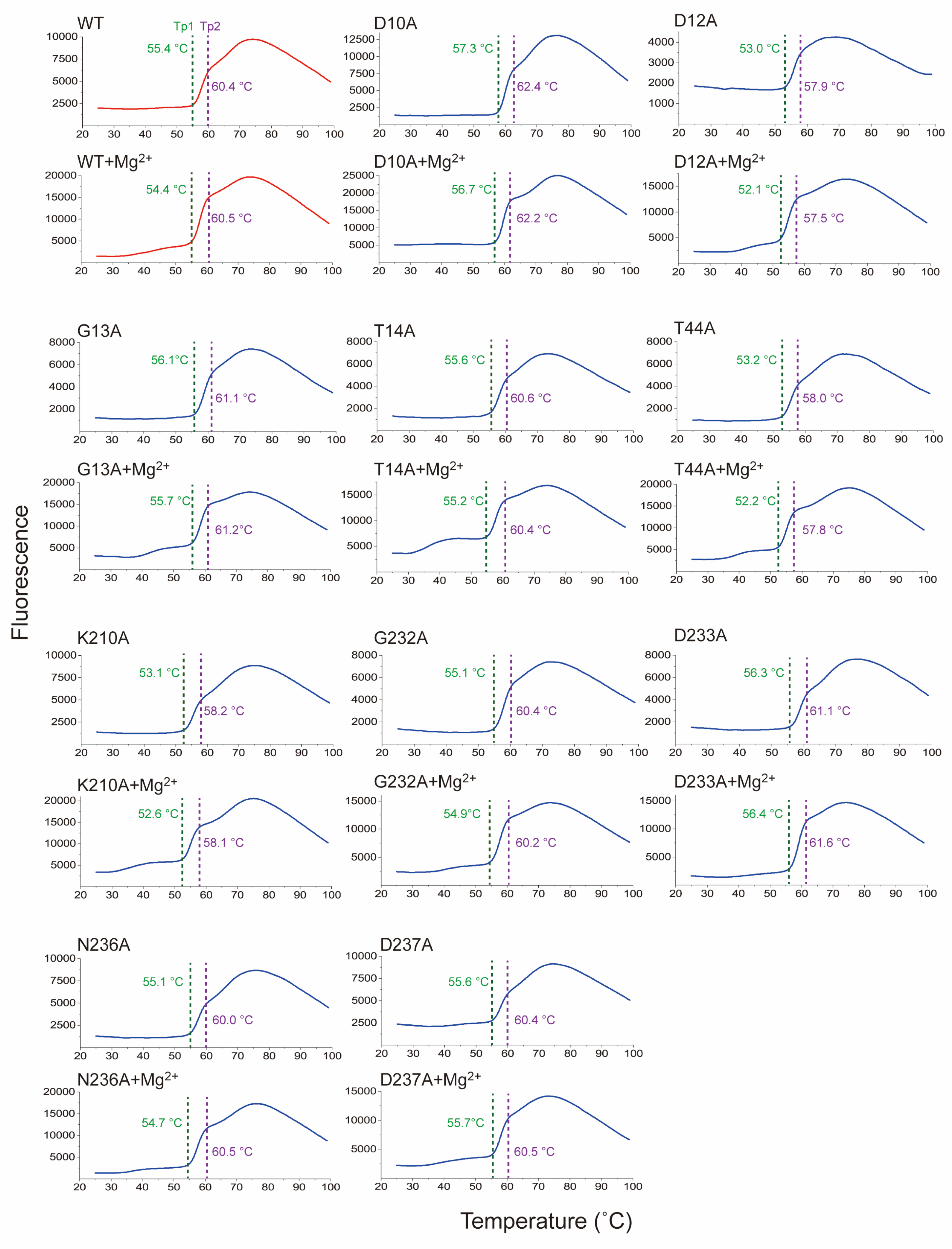
2.5. Thermal Stability of Sav2152
3. Discussion
4. Materials and Methods
4.1. Protein Preparation
4.2. Mutagenesis in Sav2152
4.3. Multiple-Angle Light Scattering
4.4. Crystallization and Data Collection
4.5. Structure Determination
4.6. Phosphatase Activity Measurement
4.7. Melting Temperature Scanning
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Newsom, S.W. Ogston’s coccus. J. Hosp. Infect. 2008, 70, 369–372. [Google Scholar] [CrossRef] [PubMed]
- Lowy, F.D. Staphylococcus aureus infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar] [CrossRef] [PubMed]
- Monecke, S.; Coombs, G.; Shore, A.C.; Coleman, D.C.; Akpaka, P.; Borg, M.; Chow, H.; Ip, M.; Jatzwauk, L.; Jonas, D.; et al. A field guide to pandemic, epidemic and sporadic clones of methicillin-resistant Staphylococcus aureus. PLoS ONE 2011, 6, e17936. [Google Scholar] [CrossRef] [PubMed]
- Pinho, M.G.; de Lencastre, H.; Tomasz, A. An acquired and a native penicillin-binding protein cooperate in building the cell wall of drug-resistant staphylococci. Proc. Natl. Acad. Sci. USA 2001, 98, 10886–10891. [Google Scholar] [CrossRef] [PubMed]
- Fridkin, S.K.; Hageman, J.C.; Morrison, M.; Sanza, L.T.; Como-Sabetti, K.; Jernigan, J.A.; Harriman, K.; Harrison, L.H.; Lynfield, R.; Farley, M.M. Methicillin-resistant Staphylococcus aureus disease in three communities. N. Engl. J. Med. 2005, 352, 1436–1444. [Google Scholar] [CrossRef] [PubMed]
- Gillet, Y.; Vanhems, P.; Lina, G.; Bes, M.; Vandenesch, F.; Floret, D.; Etienne, J. Factors predicting mortality in necrotizing community-acquired pneumonia caused by Staphylococcus aureus containing Panton-Valentine leukocidin. Clin. Infect. Dis. 2007, 45, 315–321. [Google Scholar] [CrossRef]
- Moran, G.J.; Krishnadasan, A.; Gorwitz, R.J.; Fosheim, G.E.; McDougal, L.K.; Carey, R.B.; Talan, D.A. Methicillin-resistant S. aureus infections among patients in the emergency department. N. Engl. J. Med. 2006, 355, 666–674. [Google Scholar] [CrossRef] [PubMed]
- Pallin, D.J.; Egan, D.J.; Pelletier, A.J.; Espinola, J.A.; Hooper, D.C.; Camargo, C.A., Jr. Increased US emergency department visits for skin and soft tissue infections, and changes in antibiotic choices, during the emergence of community-associated methicillin-resistant Staphylococcus aureus. Ann. Emerg. Med. 2008, 51, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Malachowa, N.; DeLeo, F.R. Mobile genetic elements of Staphylococcus aureus. Cell. Mol. Life Sci. 2010, 67, 3057–3071. [Google Scholar] [CrossRef]
- Enright, M.C.; Day, N.P.; Davies, C.E.; Peacock, S.J.; Spratt, B.G. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J. Clin. Microbiol. 2000, 38, 1008–1015. [Google Scholar] [CrossRef]
- O’Hara, F.P.; Suaya, J.A.; Ray, G.T.; Baxter, R.; Brown, M.L.; Mera, R.M.; Close, N.M.; Thomas, E.; Amrine-Madsen, H. spa Typing and Multilocus Sequence Typing Show Comparable Performance in a Macroepidemiologic Study of Staphylococcus aureus in the United States. Microb. Drug Resist. 2016, 22, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Nicholas, K.R.; Modepalli, V.; Watt, A.P.; Hinds, L.A.; Kumar, A.; Lefevre, C.; Sharp, J.A. Guiding Development of the Neonate: Lessons from Mammalia. Nestle Nutr. Inst. Workshop Ser. 2019, 90, 203–215. [Google Scholar] [PubMed]
- Enany, S.; Yoshida, Y.; Magdeldin, S.; Zhang, Y.; Bo, X.; Yamamoto, T. Extensive proteomic profiling of the secretome of European community acquired methicillin resistant Staphylococcus aureus clone. Peptides 2012, 37, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Grabowski, M.; Chruszcz, M.; Zimmerman, M.D.; Kirillova, O.; Minor, W. Benefits of structural genomics for drug discovery research. Infect. Disord. Drug Targets 2009, 9, 459–474. [Google Scholar] [CrossRef] [PubMed]
- Allen, K.N.; Dunaway-Mariano, D. Markers of fitness in a successful enzyme superfamily. Curr. Opin. Struct. Biol. 2009, 19, 658–665. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Yakunin, A.F.; Kuznetsova, E.; Xu, X.; Pennycooke, M.; Gu, J.; Cheung, F.; Proudfoot, M.; Arrowsmith, C.H.; Joachimiak, A.; et al. Structure- and function-based characterization of a new phosphoglycolate phosphatase from Thermoplasma acidophilum. J. Biol. Chem. 2004, 279, 517–526. [Google Scholar] [CrossRef]
- Yamamoto, H.; Takio, K.; Sugahara, M.; Kunishima, N. Structure of a haloacid dehalogenase superfamily phosphatase PH1421 from Pyrococcus horikoshii OT3: Oligomeric state and thermoadaptation mechanism. Acta Crystallogr. D Biol. Crystallogr. 2008, 64, 1068–1077. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera--a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Holm, L.; Laiho, A.; Toronen, P.; Salgado, M. DALI shines a light on remote homologs: One hundred discoveries. Protein Sci. 2023, 32, e4519. [Google Scholar] [CrossRef]
- Lahiri, S.D.; Zhang, G.; Dunaway-Mariano, D.; Allen, K.N. Diversification of function in the haloacid dehalogenase enzyme superfamily: The role of the cap domain in hydrolytic phosphoruscarbon bond cleavage. Bioorg Chem. 2006, 34, 394–409. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Armitano, J.; Redder, P.; Guimaraes, V.A.; Linder, P. An Essential Factor for High Mg(2+) Tolerance of Staphylococcus aureus. Front. Microbiol. 2016, 7, 1888. [Google Scholar] [CrossRef] [PubMed]
- Collet, J.F.; Stroobant, V.; Van Schaftingen, E. Mechanistic studies of phosphoserine phosphatase, an enzyme related to P-type ATPases. J. Biol. Chem. 1999, 274, 33985–33990. [Google Scholar] [CrossRef] [PubMed]
- Rinaldo-Matthis, A.; Rampazzo, C.; Reichard, P.; Bianchi, V.; Nordlund, P. Crystal structure of a human mitochondrial deoxyribonucleotidase. Nat. Struct. Biol. 2002, 9, 779–787. [Google Scholar] [CrossRef] [PubMed]
- Kuznetsova, E.; Nocek, B.; Brown, G.; Makarova, K.S.; Flick, R.; Wolf, Y.I.; Khusnutdinova, A.; Evdokimova, E.; Jin, K.; Tan, K.; et al. Functional Diversity of Haloacid Dehalogenase Superfamily Phosphatases from Saccharomyces cerevisiae: BIOCHEMICAL, STRUCTURAL, AND EVOLUTIONARY INSIGHTS. J. Biol. Chem. 2015, 290, 18678–18698. [Google Scholar] [CrossRef] [PubMed]
- Kabsch, W. Integration, scaling, space-group assignment and post-refinement. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Liebschner, D.; Afonine, P.V.; Baker, M.L.; Bunkoczi, G.; Chen, V.B.; Croll, T.I.; Hintze, B.; Hung, L.W.; Jain, S.; McCoy, A.J.; et al. Macromolecular structure determination using X-rays, neutrons and electrons: Recent developments in Phenix. Acta Crystallogr. D Struct. Biol. 2019, 75, 861–877. [Google Scholar] [CrossRef]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Crystallogr. D Biol. Crystallogr. 2010, 66, 486–501. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data Collection | |
| wavelength (Å) | 1.0000 |
| resolution (Å) | 27.76–1.92 (1.99–1.92) |
| space group | P 21 2 21 |
| unit cell dimensions | |
| a, b, c (Å) | 53.61, 63.06, 92.73 |
| α, β, γ (°) | 90, 90, 90 |
| No. of total reflections | 177,406 (17,205) |
| No. of unique reflections | 24,627 (2409) |
| completeness (%) | 99.83 (99.88) |
| Mean I/sigma (I) | 14.24 (4.99) |
| Rmerge a | 0.0823 (0.4843) |
| Rmeas | 0.08952 (0.5221) |
| Rpim b | 0.03321 (0.1922) |
| CC1/2 | 0.996 (0.958) |
| Refinement | |
| Reflections used in refinement | 24,638 (2408) |
| Reflections used for Rfree | 1231 (121) |
| Rwork/Rfree (%) c | 0.1957 (0.2243)/0.2318 (0.2994) |
| No. of Protein residues / B factors (Å2) | 231/35.90 |
| No. of Water / B factors (Å2) | 147/41.85 |
| Wilson B-factor | 29.10 |
| rmsd (bonds) | 0.013 |
| rmsd (angles) | 1.33 |
| Ramachandran favored (%) | 97.82 |
| Ramachandran allowed (%) | 1.75 |
| Ramachandran outliers (%) | 0.44 |
| Rotamer outliers (%) | 0.99 |
| Clashscore | 4.38 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bang, J.; Park, J.; Lee, S.-H.; Jang, J.; Hwang, J.; Kamarov, O.; Park, H.-J.; Lee, S.-J.; Seo, M.-D.; Won, H.-S.; et al. Nontraditional Roles of Magnesium Ions in Modulating Sav2152: Insight from a Haloacid Dehalogenase-like Superfamily Phosphatase from Staphylococcus aureus. Int. J. Mol. Sci. 2024, 25, 5021. https://doi.org/10.3390/ijms25095021
Bang J, Park J, Lee S-H, Jang J, Hwang J, Kamarov O, Park H-J, Lee S-J, Seo M-D, Won H-S, et al. Nontraditional Roles of Magnesium Ions in Modulating Sav2152: Insight from a Haloacid Dehalogenase-like Superfamily Phosphatase from Staphylococcus aureus. International Journal of Molecular Sciences. 2024; 25(9):5021. https://doi.org/10.3390/ijms25095021
Chicago/Turabian StyleBang, Jaeseok, Jaehui Park, Sung-Hee Lee, Jinhwa Jang, Junwoo Hwang, Otabek Kamarov, Hae-Joon Park, Soo-Jae Lee, Min-Duk Seo, Hyung-Sik Won, and et al. 2024. "Nontraditional Roles of Magnesium Ions in Modulating Sav2152: Insight from a Haloacid Dehalogenase-like Superfamily Phosphatase from Staphylococcus aureus" International Journal of Molecular Sciences 25, no. 9: 5021. https://doi.org/10.3390/ijms25095021
APA StyleBang, J., Park, J., Lee, S.-H., Jang, J., Hwang, J., Kamarov, O., Park, H.-J., Lee, S.-J., Seo, M.-D., Won, H.-S., Seok, S.-H., & Kim, J.-H. (2024). Nontraditional Roles of Magnesium Ions in Modulating Sav2152: Insight from a Haloacid Dehalogenase-like Superfamily Phosphatase from Staphylococcus aureus. International Journal of Molecular Sciences, 25(9), 5021. https://doi.org/10.3390/ijms25095021