
Role of Calcitriol and Vitamin D Receptor (VDR) Gene Polymorphisms in Alzheimer’s Disease



Abstract
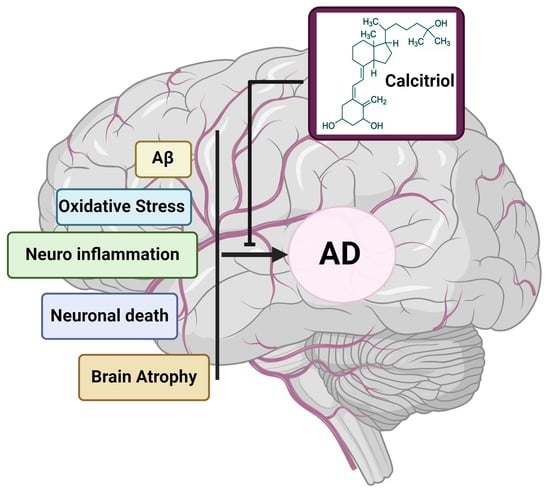
1. Introduction
2. Methods
3. Synthesis and Activation of Vitamin D
4. Transport of Vitamin D
5. Distribution of VDR in Brain
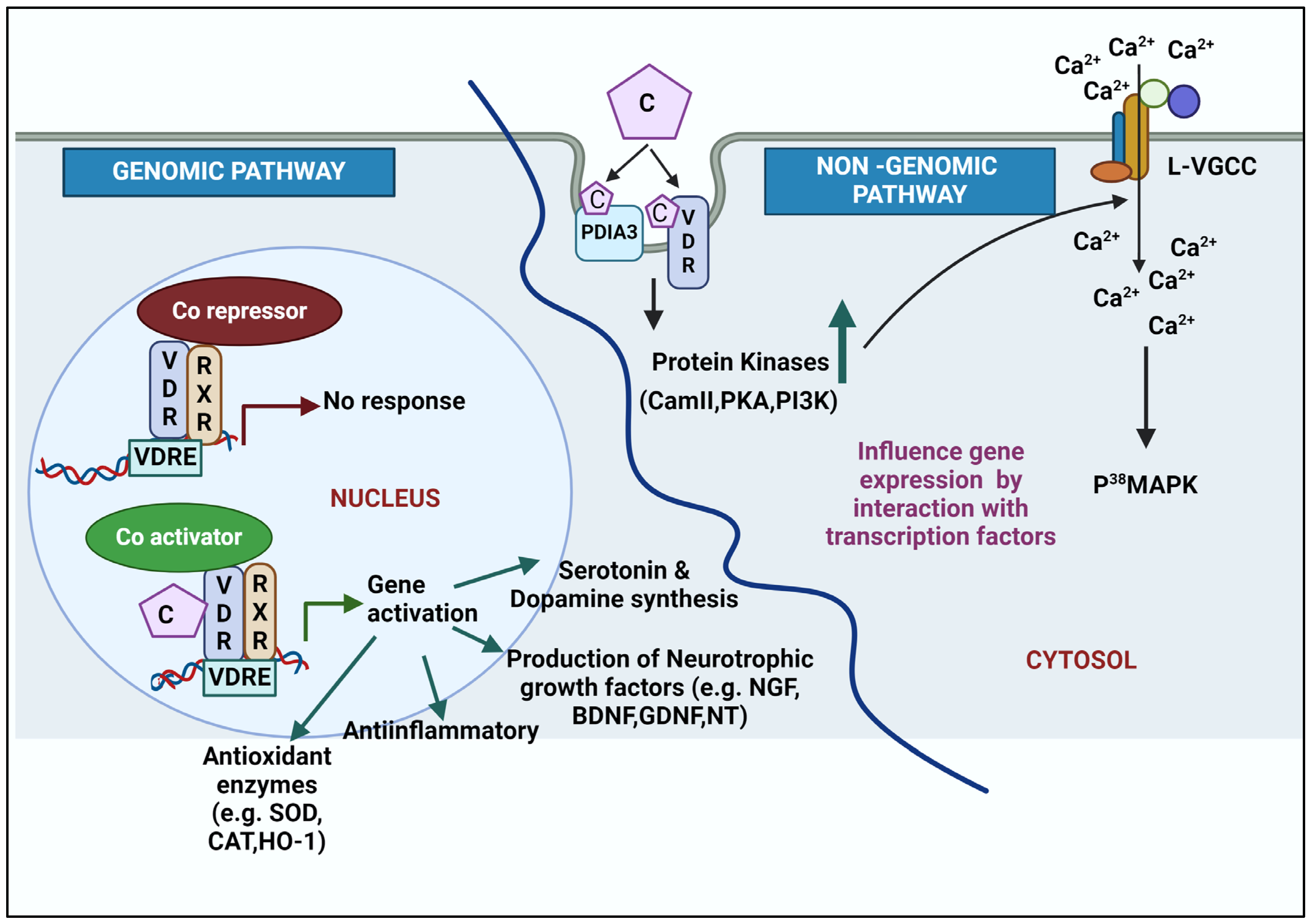
6. Molecular Mechanism of Action of Calcitriol
6.1. Genomic Action of Calcitriol
6.2. Non-Genomic Action of Calcitriol
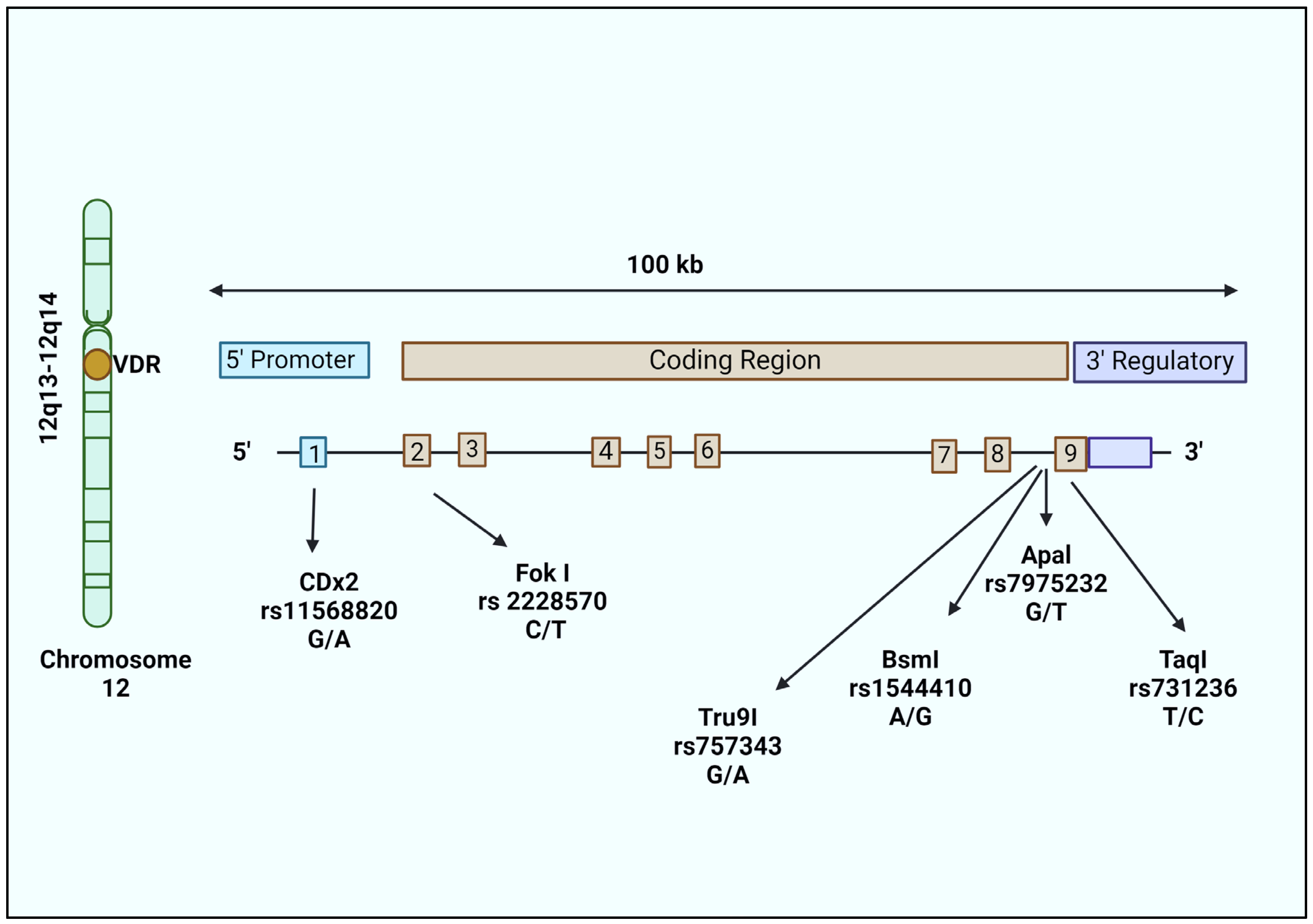
7. Genetic Variants in VDR and Risk of Late-Onset AD (LOAD)
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Gustavsson, A.; Norton, N.; Fast, T.; Frölich, L.; Georges, J.; Holzapfel, D.; Kirabali, T.; Krolak-Salmon, P.; Rossini, P.M.; Ferretti, M.T. Global estimates on the number of persons across the Alzheimer’s disease continuum. Alzheimer’s Dement. 2023, 19, 658–670. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.; Zhang, D.; Zeng, Y.; Huang, T.Y.; Xu, H.; Zhao, Y. Molecular and cellular mechanisms underlying the pathogenesis of Alzheimer’s disease. Mol. Neurodegener. 2020, 15, 40. [Google Scholar] [CrossRef] [PubMed]
- Tönnies, E.; Trushina, E. Oxidative stress, synaptic dysfunction, and Alzheimer’s disease. J. Alzheimer’s Dis. 2017, 57, 1105–1121. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.A.; Sharma, N.; An, S.S.A. Phyto-carbazole alkaloids from the rutaceae family as potential protective agents against neurodegenerative diseases. Antioxidants 2022, 11, 493. [Google Scholar] [CrossRef] [PubMed]
- Jeon, S.-M.; Shin, E.-A. Exploring vitamin D metabolism and function in cancer. Exp. Mol. Med. 2018, 50, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Sîrbe, C.; Rednic, S.; Grama, A.; Pop, T.L. An Update on the Effects of Vitamin D on the Immune System and Autoimmune Diseases. Int. J. Mol. Sci. 2022, 23, 9784. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Liu, M.; Wang, C.; Xiao, Y.; An, T.; Zou, M.; Cheng, G. Association between vitamin D status and asthma control: A meta-analysis of randomized trials. Respir. Med. 2019, 150, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Demir, M.; Demir, F.; Aygun, H. Vitamin D deficiency is associated with COVID-19 positivity and severity of the disease. J. Med. Virol. 2021, 93, 2992–2999. [Google Scholar] [CrossRef] [PubMed]
- Mercola, J.; Grant, W.B.; Wagner, C.L. Evidence regarding vitamin D and risk of COVID-19 and its severity. Nutrients 2020, 12, 3361. [Google Scholar] [CrossRef]
- Dietary Supplement Fact Sheets-Vitamin D. Available online: https://ods.od.nih.gov/factsheets/VitaminD-HealthProfessional/ (accessed on 5 January 2024).
- Olszewska, A.M.; Sieradzan, A.K.; Bednarczyk, P.; Szewczyk, A.; Żmijewski, M.A. Mitochondrial potassium channels: A novel calcitriol target. Cell. Mol. Biol. Lett. 2022, 27, 3. [Google Scholar] [CrossRef]
- Simpson, R.U.; Weishaar, R. Newly discovered activities for calcitriol (1,25-dihydroxyvitamin D3): Implications for future pharmacological use. Bioessays 1986, 4, 65–70. [Google Scholar] [CrossRef]
- Eyles, D.W.; Smith, S.; Kinobe, R.; Hewison, M.; McGrath, J.J. Distribution of the vitamin D receptor and 1α-hydroxylase in human brain. J. Chem. Neuroanat. 2005, 29, 21–30. [Google Scholar] [CrossRef]
- Cannell, J.J. Autism and vitamin D. Med. Hypotheses 2008, 70, 750–759. [Google Scholar] [CrossRef]
- McGrath, J. Hypothesis: Is low prenatal vitamin D a risk-modifying factor for schizophrenia? Schizophr. Res. 1999, 40, 173–177. [Google Scholar] [CrossRef]
- Darwish, H.; Haddad, R.; Osman, S.; Ghassan, S.; Yamout, B.; Tamim, H.; Khoury, S. Effect of Vitamin D Replacement on Cognition in Multiple Sclerosis Patients. Sci. Rep. 2017, 7, 45926. [Google Scholar] [CrossRef]
- Hongell, K.; Silva, D.G.; Ritter, S.; Meier, D.P.; Soilu-Hänninen, M. Efficacy and safety outcomes in vitamin D supplement users in the fingolimod phase 3 trials. J. Neurol. 2018, 265, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Nakajima-Ohyama, K.C.; Tansho, K.; Tanimukai, H. An alarm on vitamin D therapy for Alzheimer’s disease patients: A case with Alzheimer’s disease whose symptoms were exacerbated under chronic use of eldecalcitol. Psychogeriatrics 2022, 22, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Bivona, G.; Agnello, L.; Bellia, C.; Iacolino, G.; Scazzone, C.; Lo Sasso, B.; Ciaccio, M. Non-skeletal activities of vitamin D: From physiology to brain pathology. Medicina 2019, 55, 341. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Pelekanos, M.; Liu, P.-Y.; Burne, T.; McGrath, J.; Eyles, D. The vitamin D receptor in dopamine neurons; its presence in human substantia nigra and its ontogenesis in rat midbrain. Neuroscience 2013, 236, 77–87. [Google Scholar] [CrossRef]
- Annweiler, C. Vitamin D in dementia prevention. Ann. N. Y. Acad. Sci. 2016, 1367, 57–63. [Google Scholar] [CrossRef]
- Di Somma, C.; Scarano, E.; Barrea, L.; Zhukouskaya, V.V.; Savastano, S.; Mele, C.; Scacchi, M.; Aimaretti, G.; Colao, A.; Marzullo, P. Vitamin D and Neurological Diseases: An Endocrine View. Int. J. Mol. Sci. 2017, 18, 2482. [Google Scholar] [CrossRef]
- Calvello, R.; Cianciulli, A.; Nicolardi, G.; De Nuccio, F.; Giannotti, L.; Salvatore, R.; Porro, C.; Trotta, T.; Panaro, M.A.; Lofrumento, D.D. Vitamin D Treatment Attenuates Neuroinflammation and Dopaminergic Neurodegeneration in an Animal Model of Parkinson’s Disease, Shifting M1 to M2 Microglia Responses. J. Neuroimmune Pharmacol. 2017, 12, 327–339. [Google Scholar] [CrossRef]
- De Oliveira, L.R.C.; Mimura, L.A.N.; Fraga-Silva, T.F.D.C.; Ishikawa, L.L.W.; Fernandes, A.A.H.; Zorzella-Pezavento, S.F.G.; Sartori, A. Calcitriol prevents neuroinflammation and reduces blood-brain barrier disruption and local macrophage/microglia activation. Front. Pharmacol. 2020, 11, 161. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Park, M.; Lee, E.; Jung, J.; Kim, T. The Role of Vitamin D in Alzheimer’s Disease: A Transcriptional Regulator of Amyloidopathy and Gliopathy. Biomedicines 2022, 10, 1824. [Google Scholar] [CrossRef]
- Sadeghian, N.; Shadman, J.; Moradi, A.; ghasem Golmohammadi, M.; Panahpour, H. Calcitriol protects the Blood-Brain Barrier integrity against ischemic stroke and reduces vasogenic brain edema via antioxidant and antiapoptotic actions in rats. Brain Res. Bull. 2019, 150, 281–289. [Google Scholar] [CrossRef]
- Scandinavian Society for Clinical Chemistry, Clinical Physiology. Scandinavian Journal of Clinical & Laboratory Investigation; Blackwell Scientific Publications: Hoboken, NJ, USA, 1987; Volume 47. [Google Scholar]
- Hollis, B.W. Comparison of equilibrium and disequilibrium assay conditions for ergocalciferol, cholecalciferol and their major metabolites. J. Steroid Biochem. 1984, 21, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Afzal, S.; Bojesen, S.E.; Nordestgaard, B.G. Reduced 25-hydroxyvitamin D and risk of Alzheimer’s disease and vascular dementia. Alzheimer’s Dement. 2014, 10, 296–302. [Google Scholar] [CrossRef]
- Lu, Y.; Li, J.; Hu, T.; Huang, G. Serum 25-hydroxy vitamin D level is associated with cognitive impairment in people aged 65 years and older. Ann. Palliat. Med. 2021, 10, 7479–7485. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Xue, W.; Li, J.; Fu, K.; Shi, H.; Zhang, B.; Teng, W.; Tian, L. 25-Hydroxyvitamin D Levels and the Risk of Dementia and Alzheimer’s Disease: A Dose-Response Meta-Analysis. Front. Aging Neurosci. 2018, 10, 368. [Google Scholar] [CrossRef]
- Jayedi, A.; Rashidy-Pour, A.; Shab-Bidar, S. Vitamin D status and risk of dementia and Alzheimer’s disease: A meta-analysis of dose-response. Nutr. Neurosci. 2019, 22, 750–759. [Google Scholar] [CrossRef]
- Shen, L.; Ji, H.F. Vitamin D deficiency is associated with increased risk of Alzheimer’s disease and dementia: Evidence from meta-analysis. Nutr. J. 2015, 14, 76. [Google Scholar] [CrossRef] [PubMed]
- Sommer, I.; Griebler, U.; Kien, C.; Auer, S.; Klerings, I.; Hammer, R.; Holzer, P.; Gartlehner, G. Vitamin D deficiency as a risk factor for dementia: A systematic review and meta-analysis. BMC Geriatr. 2017, 17, 16. [Google Scholar] [CrossRef] [PubMed]
- Acar, S.; Özkan, B. Vitamin D Metabolism; Özdemir, Ö., Ed.; IntechOpen: London, UK, 2021; Volume 2021, pp. 1–23. [Google Scholar]
- Bikle, D.D.; Malmstroem, S.; Schwartz, J. Current controversies: Are free vitamin metabolite levels a more accurate assessment of vitamin D status than total levels? Endocrinol. Metab. Clin. 2017, 46, 901–918. [Google Scholar] [CrossRef] [PubMed]
- Soeters, P.B.; de Leeuw, P.W. Chapter 16—Vitamin D in health and disease. In Reciprocal Translation Between Pathophysiology and Practice in Health and Disease; Soeters, P.B., de Leeuw, P.W., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 201–219. [Google Scholar] [CrossRef]
- Zhang, G.; Li, L.; Kong, Y.; Xu, D.; Bao, Y.; Zhang, Z.; Liao, Z.; Jiao, J.; Fan, D.; Long, X.; et al. Vitamin D-binding protein in plasma microglia-derived extracellular vesicles as a potential biomarker for major depressive disorder. Genes Dis. 2024, 11, 1009–1021. [Google Scholar] [CrossRef] [PubMed]
- Bikle, D.; Halloran, B.; Gee, E.; Ryzen, E.; Haddad, J. Free 25-hydroxyvitamin D levels are normal in subjects with liver disease and reduced total 25-hydroxyvitamin D levels. J. Clin. Investig. 1986, 78, 748–752. [Google Scholar] [CrossRef] [PubMed]
- Maghbooli, Z.; Omidifar, A.; Varzandi, T.; Salehnezhad, T.; Sahraian, M.A. Reduction in circulating vitamin D binding protein in patients with multiple sclerosis. BMC Neurol. 2021, 21, 168. [Google Scholar] [CrossRef] [PubMed]
- Bikle, D.D.; Schwartz, J. Vitamin D binding protein, total and free vitamin D levels in different physiological and pathophysiological conditions. Front. Endocrinol. 2019, 10, 317. [Google Scholar] [CrossRef] [PubMed]
- Gezen-Ak, D.; Alaylıoğlu, M.; Genç, G.; Gündüz, A.; Candaş, E.; Bilgiç, B.; Atasoy, I.L.; Apaydın, H.; Kızıltan, G.; Gürvit, H. GC and VDR SNPs and vitamin D levels in Parkinson’s disease: The relevance to clinical features. Neuromolecular Med. 2017, 19, 24–40. [Google Scholar] [CrossRef] [PubMed]
- Vargas, T.; Bullido, M.J.; Martinez-Garcia, A.; Antequera, D.; Clarimon, J.; Rosich-Estrago, M.; Martin-Requero, A.; Mateo, I.; Rodriguez-Rodriguez, E.; Vilella-Cuadrada, E. A megalin polymorphism associated with promoter activity and Alzheimer’s disease risk. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2010, 153, 895–902. [Google Scholar] [CrossRef]
- Wang, Y.; Zhu, J.; DeLuca, H.F. Where is the vitamin D receptor? Arch. Biochem. Biophys. 2012, 523, 123–133. [Google Scholar] [CrossRef]
- Sutherland, M.K.; Somerville, M.J.; Yoong, L.K.; Bergeron, C.; Haussler, M.R.; McLachlan, D.R.C. Reduction of vitamin D hormone receptor mRNA levels in Alzheimer as compared to Huntington hippocampus: Correlation with calbindin-28k mRNA levels. Mol. Brain Res. 1992, 13, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Moore, T.B.; Koeffler, H.P.; Yamashiro, J.M.; Wada, R.K. Vitamin D 3 analogs inhibit growth and induce differentiation in LA-N-5 human neuroblastoma cells. Clin. Exp. Metastasis 1996, 14, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Burkert, R.; McGrath, J.; Eyles, D. Vitamin D receptor expression in the embryonic rat brain. Neurosci. Res. Commun. 2003, 33, 63–71. [Google Scholar] [CrossRef]
- Musiol, I.; Stumpf, W.; Bidmon, H.-J.; Heiss, C.; Mayerhofer, A.; Bartke, A. Vitamin D nuclear binding to neurons of the septal, substriatal and amygdaloid area in the Siberian hamster (Phodopus sungorus) brain. Neuroscience 1992, 48, 841–848. [Google Scholar] [CrossRef] [PubMed]
- Prüfer, K.; Veenstra, T.D.; Jirikowski, G.F.; Kumar, R. Distribution of 1, 25-dihydroxyvitamin D3 receptor immunoreactivity in the rat brain and spinal cord. J. Chem. Neuroanat. 1999, 16, 135–145. [Google Scholar] [CrossRef] [PubMed]
- López-Vicente, M.; Sunyer, J.; Lertxundi, N.; González, L.; Rodríguez-Dehli, C.; Espada Sáenz-Torre, M.; Vrijheid, M.; Tardón, A.; Llop, S.; Torrent, M.; et al. Maternal circulating Vitamin D3 levels during pregnancy and behaviour across childhood. Sci. Rep. 2019, 9, 14792. [Google Scholar] [CrossRef]
- Mossin, M.H.; Aaby, J.B.; Dalgård, C.; Lykkedegn, S.; Christesen, H.T.; Bilenberg, N. Inverse associations between cord vitamin D and attention deficit hyperactivity disorder symptoms: A child cohort study. Aust. New Zealand J. Psychiatry 2017, 51, 703–710. [Google Scholar] [CrossRef]
- Schmidt, R.J. Gestational vitamin D and Autism Spectrum Disorder. Biol. Psychiatry 2021, 90, 738–741. [Google Scholar] [CrossRef] [PubMed]
- Windham, G.C.; Pearl, M.; Anderson, M.C.; Poon, V.; Eyles, D.; Jones, K.L.; Lyall, K.; Kharrazi, M.; Croen, L.A. Newborn vitamin D levels in relation to autism spectrum disorders and intellectual disability: A case–control study in California. Autism Res. 2019, 12, 989–998. [Google Scholar] [CrossRef]
- Harms, L.R.; Burne, T.H.J.; Eyles, D.W.; McGrath, J.J. Vitamin D and the brain. Best Pract. Res. Clin. Endocrinol. Metab. 2011, 25, 657–669. [Google Scholar] [CrossRef]
- Xue, Y.; He, X.; Li, H.-D.; Deng, Y.; Yan, M.; Cai, H.-L.; Tang, M.-M.; Dang, R.-L.; Jiang, P. Simultaneous quantification of 25-hydroxyvitamin D 3 and 24, 25-dihydroxyvitamin D 3 in rats shows strong correlations between serum and brain tissue levels. Int. J. Endocrinol. 2015, 2015, 296531. [Google Scholar] [CrossRef] [PubMed]
- Spach, K.M.; Hayes, C.E. Vitamin D3 confers protection from autoimmune encephalomyelitis only in female mice. J. Immunol. 2005, 175, 4119–4126. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Dolnikowski, G.G.; Patterson, W.B.; Dawson-Hughes, B.; Zheng, T.; Morris, M.C.; Holland, T.M.; Booth, S.L. Determination of vitamin D and its metabolites in human brain using an Ultra-Pressure LC–Tandem Mass Spectra method. Curr. Dev. Nutr. 2019, 3, nzz074. [Google Scholar] [CrossRef] [PubMed]
- Lou, Y.-R.; Molnár, F.; Peräkylä, M.; Qiao, S.; Kalueff, A.V.; St-Arnaud, R.; Carlberg, C.; Tuohimaa, P. 25-Hydroxyvitamin D3 is an agonistic vitamin D receptor ligand. J. Steroid Biochem. Mol. Biol. 2010, 118, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, F.; Mayer, E.; Norman, A.W. Biological activity assessment of the 26, 23-lactones of 1, 25-dihydroxyvitamin D3 and 25-hydroxyvitamin D3 and their binding properties to chick intestinal receptor and plasma vitamin D binding protein. Arch. Biochem. Biophys. 1984, 233, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Feldman, D.; Malloy, P.J.; Gross, C. Vitamin D: Biology, action, and clinical implications. In Osteoporosis; Elsevier: Amsterdam, The Netherlands, 2001; pp. 257–303. [Google Scholar]
- Morris, H.A. Vitamin D metabolism and molecular modes of action: New insights into vitamin D activities. Med. Res. J. 2014, 2, 1–5. [Google Scholar]
- Theodoropoulos, C.; Demers, C.; Delvin, E.; Ménard, D.; Gascon-Barré, M. Calcitriol regulates the expression of the genes encoding the three key vitamin D3 hydroxylases and the drug-metabolizing enzyme CYP3A4 in the human fetal intestine. Clin. Endocrinol. 2003, 58, 489–499. [Google Scholar] [CrossRef]
- Liu, H.L.; Chuang, H.Y.; Hsu, C.N.; Lee, S.S.; Yang, C.C.; Liu, K.T. Effects of Vitamin D Receptor, Metallothionein 1A, and 2A Gene Polymorphisms on Toxicity of the Peripheral Nervous System in Chronically Lead-Exposed Workers. Int. J. Env. Res. Public Health 2020, 17, 2909. [Google Scholar] [CrossRef] [PubMed]
- Warwick, T.; Schulz, M.H.; Günther, S.; Gilsbach, R.; Neme, A.; Carlberg, C.; Brandes, R.P.; Seuter, S. A hierarchical regulatory network analysis of the vitamin D induced transcriptome reveals novel regulators and complete VDR dependency in monocytes. Sci. Rep. 2021, 11, 6518. [Google Scholar] [CrossRef]
- Ideraabdullah, F.Y.; Belenchia, A.M.; Rosenfeld, C.S.; Kullman, S.W.; Knuth, M.; Mahapatra, D.; Bereman, M.; Levin, E.D.; Peterson, C.A. Maternal vitamin D deficiency and developmental origins of health and disease (DOHaD). J. Endocrinol. 2019, 241, R65–R80. [Google Scholar] [CrossRef]
- Pertile, R.A.N.; Kiltschewskij, D.; Geaghan, M.; Barnett, M.; Cui, X.; Cairns, M.J.; Eyles, D. Developmental vitamin D-deficiency increases the expression of microRNAs involved in dopamine neuron development. Brain Res. 2022, 1789, 147953. [Google Scholar] [CrossRef]
- Donati, S.; Palmini, G.; Aurilia, C.; Falsetti, I.; Marini, F.; Giusti, F.; Iantomasi, T.; Brandi, M.L. Calcifediol: Mechanisms of Action. Nutrients 2023, 15, 4409. [Google Scholar] [CrossRef]
- Wang, Y.; Chiang, Y.-H.; Su, T.-P.; Hayashi, T.; Morales, M.; Hoffer, B.; Lin, S.-Z. Vitamin D3 attenuates cortical infarction induced by middle cerebral arterial ligation in rats. Neuropharmacology 2000, 39, 873–880. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, B.; Lopez-Martin, E.; Segura, C.; Labandeira-Garcia, J.L.; Perez-Fernandez, R. 1,25-Dihydroxyvitamin D3 increases striatal GDNF mRNA and protein expression in adult rats. Mol. Brain Res. 2002, 108, 143–146. [Google Scholar] [CrossRef] [PubMed]
- Roussa, E.; Krieglstein, K. GDNF promotes neuronal differentiation and dopaminergic development of mouse mesencephalic neurospheres. Neurosci. Lett. 2004, 361, 52–55. [Google Scholar] [CrossRef]
- Cui, X.; Pertile, R.; Liu, P.; Eyles, D.W. Vitamin D regulates tyrosine hydroxylase expression: N-cadherin a possible mediator. Neuroscience 2015, 304, 90–100. [Google Scholar] [CrossRef]
- Cass, W.A.; Smith, M.P.; Peters, L.E. Calcitriol protects against the dopamine-and serotonin-depleting effects of neurotoxic doses of methamphetamine. Ann. New York Acad. Sci. 2006, 1074, 261–271. [Google Scholar] [CrossRef]
- Sabir, M.S.; Haussler, M.R.; Mallick, S.; Kaneko, I.; Lucas, D.A.; Haussler, C.A.; Whitfield, G.K.; Jurutka, P.W. Optimal vitamin D spurs serotonin: 1,25-dihydroxyvitamin D represses serotonin reuptake transport (SERT) and degradation (MAO-A) gene expression in cultured rat serotonergic neuronal cell lines. Genes Nutr. 2018, 13, 19. [Google Scholar] [CrossRef] [PubMed]
- Clarke, J.; Yaqubi, M.; Futhey, N.C.; Sedaghat, S.; Baufeld, C.; Blain, M.; Baranzini, S.; Butovsky, O.; Antel, J.; White, J.H. Vitamin D regulates MerTK-dependent phagocytosis in human myeloid cells. J. Immunol. 2020, 205, 398–406. [Google Scholar] [CrossRef]
- Gezen-Ak, D.; Dursun, E.; Yilmazer, S. The effects of vitamin D receptor silencing on the expression of LVSCC-A1C and LVSCC-A1D and the release of NGF in cortical neurons. PLoS ONE 2011, 6, e17553. [Google Scholar] [CrossRef]
- Lasoń, W.; Jantas, D.; Leśkiewicz, M.; Regulska, M.; Basta-Kaim, A. Vitamin D3 and Ischemic Stroke: A Narrative Review. Antioxidants 2022, 11, 2120. [Google Scholar] [CrossRef] [PubMed]
- Mirarchi, A.; Albi, E.; Beccari, T.; Arcuri, C. Microglia and Brain Disorders: The Role of Vitamin D and Its Receptor. Int. J. Mol. Sci. 2023, 24, 11892. [Google Scholar] [CrossRef] [PubMed]
- Lasoń, W.; Jantas, D.; Leśkiewicz, M.; Regulska, M.; Basta-Kaim, A. The Vitamin D Receptor as a Potential Target for the Treatment of Age-Related Neurodegenerative Diseases Such as Alzheimer’s and Parkinson’s Diseases: A Narrative Review. Cells 2023, 12, 660. [Google Scholar] [CrossRef] [PubMed]
- Dulla, Y.A.T.; Kurauchi, Y.; Hisatsune, A.; Seki, T.; Shudo, K.; Katsuki, H. Regulatory mechanisms of vitamin D 3 on production of nitric oxide and pro-inflammatory cytokines in microglial BV-2 cells. Neurochem. Res. 2016, 41, 2848–2858. [Google Scholar] [CrossRef] [PubMed]
- Boontanrart, M.; Hall, S.D.; Spanier, J.A.; Hayes, C.E.; Olson, J.K. Vitamin D3 alters microglia immune activation by an IL-10 dependent SOCS3 mechanism. J. Neuroimmunol. 2016, 292, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yang, R.; Qiao, W.; Zhang, W.; Chen, J.; Mao, L.; Goltzman, D.; Miao, D. 1,25-Dihydroxyvitamin D exerts an antiaging role by activation of Nrf2-antioxidant signaling and inactivation of p16/p53-senescence signaling. Aging Cell 2019, 18, e12951. [Google Scholar] [CrossRef] [PubMed]
- Ward, A.; Crean, S.; Mercaldi, C.J.; Collins, J.M.; Boyd, D.; Cook, M.N.; Arrighi, H.M. Prevalence of apolipoprotein E4 genotype and homozygotes (APOE e4/4) among patients diagnosed with Alzheimer’s disease: A systematic review and meta-analysis. Neuroepidemiology 2012, 38, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Bien-Ly, N.; Andrews-Zwilling, Y.; Xu, Q.; Bernardo, A.; Ring, K.; Halabisky, B.; Deng, C.; Mahley, R.W.; Huang, Y. GABAergic interneuron dysfunction impairs hippocampal neurogenesis in adult apolipoprotein E4 knockin mice. Cell Stem Cell 2009, 5, 634–645. [Google Scholar] [CrossRef] [PubMed]
- Misra, A.; Chakrabarti, S.S.; Gambhir, I.S. New genetic players in late-onset Alzheimer’s disease: Findings of genome-wide association studies. Indian J. Med. Res. 2018, 148, 135–144. [Google Scholar] [CrossRef]
- Kuningas, M.; Mooijaart, S.P.; Jolles, J.; Slagboom, P.E.; Westendorp, R.G.; van Heemst, D. VDR gene variants associate with cognitive function and depressive symptoms in old age. Neurobiol. Aging 2009, 30, 466–473. [Google Scholar] [CrossRef]
- Beydoun, M.A.; Ding, E.L.; Beydoun, H.A.; Tanaka, T.; Ferrucci, L.; Zonderman, A.B. Vitamin D receptor and megalin gene polymorphisms and their associations with longitudinal cognitive change in US adults. Am. J. Clin. Nutr. 2012, 95, 163–178. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Zhang, T.; Ma, L.; Wei, W.; Li, Z.; Jiang, X.; Sun, J.; Pei, H.; Li, H. Vitamin D Receptor Gene Polymorphisms and Risk of Alzheimer Disease and Mild Cognitive Impairment: A Systematic Review and Meta-Analysis. Adv. Nutr. 2021, 12, 2255–2264. [Google Scholar] [CrossRef] [PubMed]
- Poduslo, S.; Yin, X. Chromosome 12 and late-onset Alzheimer’s disease. Neurosci. Lett. 2001, 310, 188–190. [Google Scholar] [CrossRef] [PubMed]
- Gezen-Ak, D.; Dursun, E.; Bilgic, B.; Hanagasi, H.; Ertan, T.; Gürvit, H.; Emre, M.; Eker, E.; Ulutin, T.; Uysal, Ö. Vitamin D receptor gene haplotype is associated with late-onset Alzheimer’s disease. Tohoku J. Exp. Med. 2012, 228, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Janjusevic, M.; Gagno, G.; Fluca, A.L.; Padoan, L.; Beltrami, A.P.; Sinagra, G.; Moretti, R.; Aleksova, A. The peculiar role of vitamin D in the pathophysiology of cardiovascular and neurodegenerative diseases. Life Sci. 2022, 289, 120193. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Li, S.; Yu, L.; Liu, J. Polymorphisms in Vitamin D Receptor Genes in Association with Childhood Autism Spectrum Disorder. Dis. Markers 2018, 2018, 7862892. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Lopez, E.; Jansen, T.; Ivaskevicius, V.; Kahles, H.; Klepzig, C.; Oldenburg, J.; Badenhoop, K. Protection from type 1 diabetes by vitamin D receptor haplotypes. Ann. New York Acad. Sci. 2006, 1079, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, D.J.; Refsum, H.; Warden, D.R.; Medway, C.; Wilcock, G.K.; Smith, A.D. The vitamin D receptor gene is associated with Alzheimer’s disease. Neurosci. Lett. 2011, 504, 79–82. [Google Scholar] [CrossRef] [PubMed]
- Łaczmański, Ł.; Jakubik, M.; Bednarek-Tupikowska, G.; Rymaszewska, J.; Słoka, N.; Lwow, F. Vitamin D receptor gene polymorphisms in Alzheimer’s disease patients. Exp. Gerontol. 2015, 69, 142–147. [Google Scholar] [CrossRef]
- Khorram Khorshid, H.R.; Gozalpour, E.; Saliminejad, K.; Karimloo, M.; Ohadi, M.; Kamali, K. Vitamin D Receptor (VDR) Polymorphisms and Late-Onset Alzheimer’s Disease: An Association Study. Iran. J. Public Health 2013, 42, 1253–1258. [Google Scholar]
- Arevalo, N.; Castillo, D.; Rogers, N.; Delgado, C.; Farías, G.; Behrens, M.I.; SanMartín, C.D. Apa I, Taq I and Fok I VDR polymorphisms: Functional effect on mRNA levels of amyloid beta transporters LRP1 and P-gp in lymphocytes of mild cognitive impairment patients: Genetics/genetic factors of Alzheimer’s disease. Alzheimer’s Dement. 2020, 16, e045564. [Google Scholar] [CrossRef]
- Dimitrakis, E.; Katsarou, M.-S.; Lagiou, M.; Papastefanopoulou, V.; Stanitsa, E.; Spandidos, D.A.; Tsatsakis, A.; Papageorgiou, S.; Moutsatsou, P.; Antoniou, K. Association of vitamin D receptor gene TaqI polymorphism with Alzheimer’s disease in a Southeastern European Caucasian population. Exp. Ther. Med. 2022, 23, 341. [Google Scholar] [CrossRef] [PubMed]
- Freitas, R.d.S.; Fratelli, C.F.; Silva, C.M.d.S.; Lima, L.R.d.; Stival, M.M.; Silva, I.C.R.d.; Funghetto, S.S. Association of vitamin D with the TaqI polymorphism of the VDR gene in older women attending the basic health unit of the Federal District, DF (Brazil). J. Aging Res. 2020, 2020, 7145193. [Google Scholar]
- Wang, L.; Hara, K.; Van Baaren, J.M.; Price, J.C.; Beecham, G.W.; Gallins, P.J.; Whitehead, P.L.; Wang, G.; Lu, C.; Slifer, M.A.; et al. Vitamin D receptor and Alzheimer’s disease: A genetic and functional study. Neurobiol. Aging 2012, 33, e1841–e1844. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Qiao, Y.; Zhang, H.; Zhang, Y.; Hua, J.; Jin, S.; Liu, G. Circulating Vitamin D Levels and Alzheimer’s Disease: A Mendelian Randomization Study in the IGAP and UK Biobank. J. Alzheimer’s Dis. 2020, 73, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Lu’o’ng, K.V.Q.; Nguyễn, L.T.H. The Beneficial Role of Vitamin D in Alzheimer’s Disease. Am. J. Alzheimer’s Dis. Other Dement.® 2011, 26, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Mizwicki, M.T.; Menegaz, D.; Zhang, J.; Barrientos-Durán, A.; Tse, S.; Cashman, J.R.; Griffin, P.R.; Fiala, M. Genomic and nongenomic signaling induced by 1α, 25(OH)2-vitamin D3 promotes the recovery of amyloid-β phagocytosis by Alzheimer’s disease macrophages. J. Alzheimer’s Dis. 2012, 29, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Lai, R.H.; Hsu, C.C.; Yu, B.H.; Lo, Y.R.; Hsu, Y.Y.; Chen, M.H.; Juang, J.L. Vitamin D supplementation worsens Alzheimer’s progression: Animal model and human cohort studies. Aging Cell 2022, 21, e13670. [Google Scholar] [CrossRef]
- Landel, V.; Annweiler, C.; Millet, P.; Morello, M.; Féron, F. Vitamin D, cognition and Alzheimer’s disease: The therapeutic benefit is in the D-tails. J. Alzheimer’s Dis. 2016, 53, 419–444. [Google Scholar] [CrossRef] [PubMed]
- Bivona, G.; Lo Sasso, B.; Gambino, C.M.; Giglio, R.V.; Scazzone, C.; Agnello, L.; Ciaccio, M. The Role of Vitamin D as a Biomarker in Alzheimer’s Disease. Brain Sci. 2021, 11, 334. [Google Scholar] [CrossRef]
- Institute of Medicine (US) Committee to Review Dietary Reference Intakes for Vitamin D and Calcium. Dietary Reference Intakes for Calcium and Vitamin D; National Academies Press: Washington, DC, USA, 2011.
- Beauchet, O.; Cooper-Brown, L.A.; Allali, G. Vitamin D supplementation and cognition in adults: A systematic review of randomized controlled trials. CNS Drugs 2021, 35, 1249–1264. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Liang, F.; Zhang, L.; Liu, J.; Dou, H. Vitamin D Supplement for Prevention of Alzheimer’s Disease: A Systematic Review and Meta-Analysis. Am. J. Ther. 2021, 28, e638–e648. [Google Scholar] [CrossRef] [PubMed]
- Jorde, R.; Kubiak, J.; Svartberg, J.; Fuskevåg, O.M.; Figenschau, Y.; Martinaityte, I.; Grimnes, G. Vitamin D supplementation has no effect on cognitive performance after four months in mid-aged and older subjects. J. Neurol. Sci. 2019, 396, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Jia, J.; Zhang, Y.; Miao, R.; Huo, X.; Ma, F. Effects of vitamin D3 supplementation on cognition and blood lipids: A 12-month randomised, double-blind, placebo-controlled trial. J. Neurol. Neurosurg. Psychiatry 2018, 89, 1341–1347. [Google Scholar] [CrossRef] [PubMed]
- Stein, M.S.; Scherer, S.C.; Ladd, K.S.; Harrison, L.C. A randomized controlled trial of high-dose vitamin D2 followed by intranasal insulin in Alzheimer’s disease. J. Alzheimer’s Dis. 2011, 26, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Tripkovic, L.; Lambert, H.; Hart, K.; Smith, C.P.; Bucca, G.; Penson, S.; Chope, G.; Hyppönen, E.; Berry, J.; Vieth, R. Comparison of vitamin D2 and vitamin D3 supplementation in raising serum 25-hydroxyvitamin D status: A systematic review and meta-analysis. Am. J. Clin. Nutr. 2012, 95, 1357–1364. [Google Scholar] [CrossRef]
- Norman, A. Intestinal calcium absorption: A vitamin D-hormone-mediated adaptive response. Am. J. Clin. Nutr. 1990, 51, 290–300. [Google Scholar] [CrossRef]
- Usategui-Martín, R.; De Luis-Román, D.-A.; Fernández-Gómez, J.M.; Ruiz-Mambrilla, M.; Pérez-Castrillón, J.-L. Vitamin D receptor (VDR) gene polymorphisms modify the response to vitamin D supplementation: A systematic review and meta-analysis. Nutrients 2022, 14, 360. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, S.P.; Sharma, N.; An, S.S.A. Role of Calcitriol and Vitamin D Receptor (VDR) Gene Polymorphisms in Alzheimer’s Disease. Int. J. Mol. Sci. 2024, 25, 4806. https://doi.org/10.3390/ijms25094806
Jeong SP, Sharma N, An SSA. Role of Calcitriol and Vitamin D Receptor (VDR) Gene Polymorphisms in Alzheimer’s Disease. International Journal of Molecular Sciences. 2024; 25(9):4806. https://doi.org/10.3390/ijms25094806
Chicago/Turabian StyleJeong, Soon Pyo, Niti Sharma, and Seong Soo A. An. 2024. "Role of Calcitriol and Vitamin D Receptor (VDR) Gene Polymorphisms in Alzheimer’s Disease" International Journal of Molecular Sciences 25, no. 9: 4806. https://doi.org/10.3390/ijms25094806
APA StyleJeong, S. P., Sharma, N., & An, S. S. A. (2024). Role of Calcitriol and Vitamin D Receptor (VDR) Gene Polymorphisms in Alzheimer’s Disease. International Journal of Molecular Sciences, 25(9), 4806. https://doi.org/10.3390/ijms25094806