
Transmembrane Protein TMEM230, Regulator of Glial Cell Vascular Mimicry and Endothelial Cell Angiogenesis in High-Grade Heterogeneous Infiltrating Gliomas and Glioblastoma
 , , , , , , , , ,
, , , , , , , , ,  ,
,  , ,
, ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
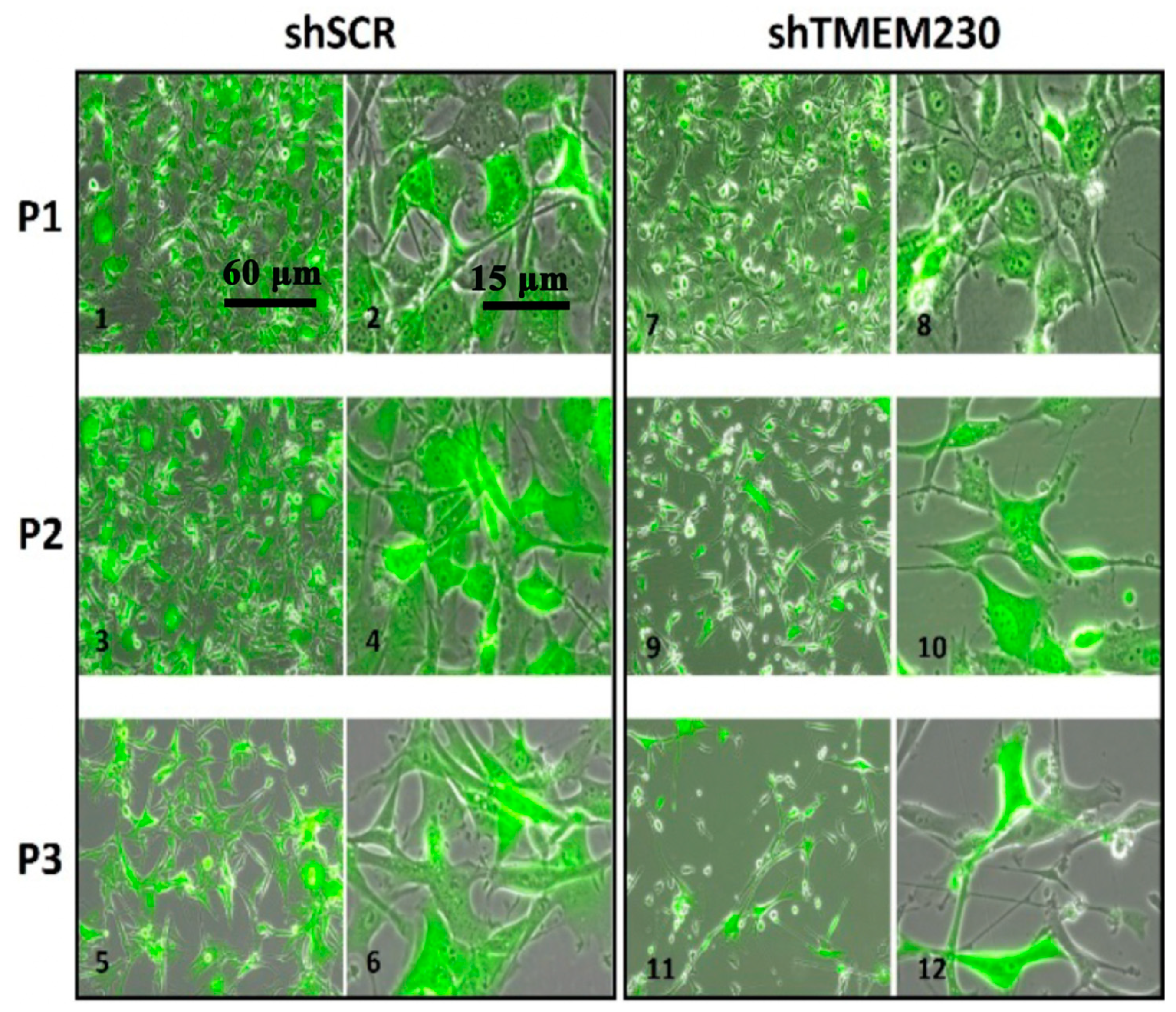
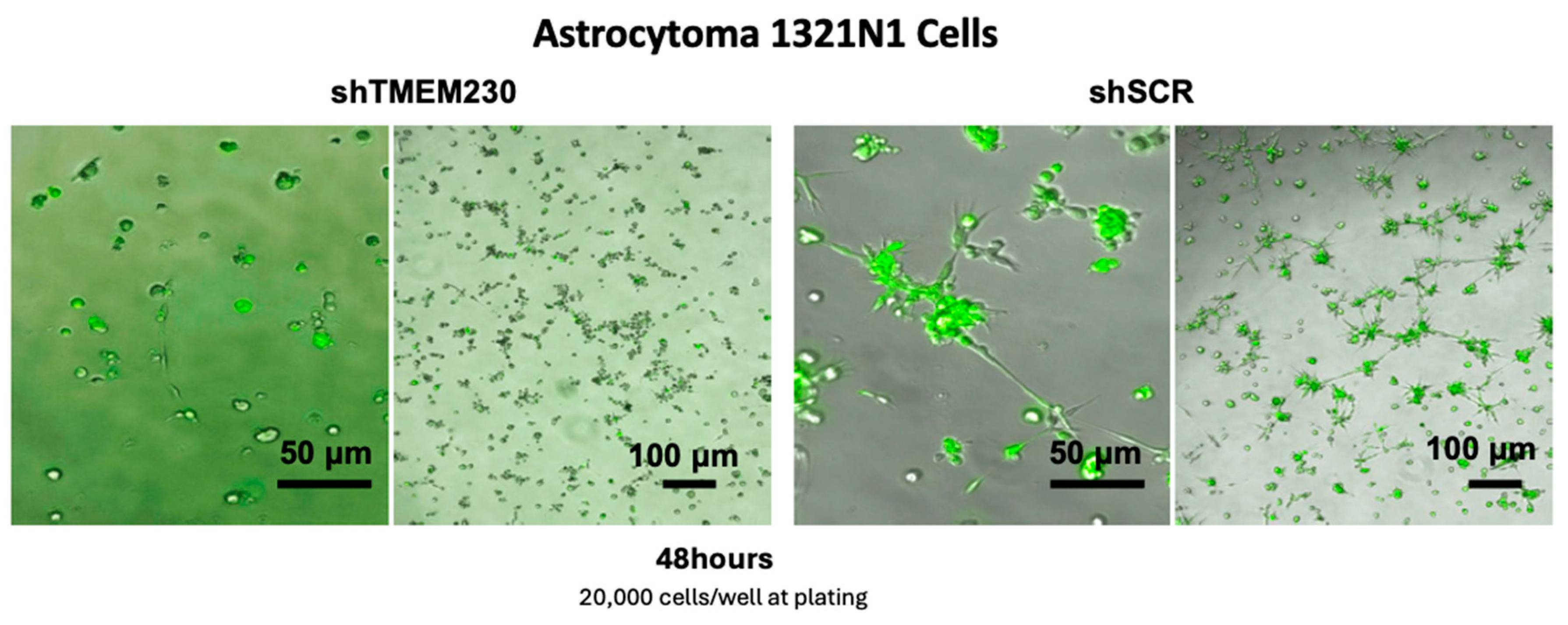
2.1. Transmembrane Protein TMEM230 Expression Is Necessary for Endothelial and Glial Cell Adhesion, Sprouting, Migration, and Infiltration
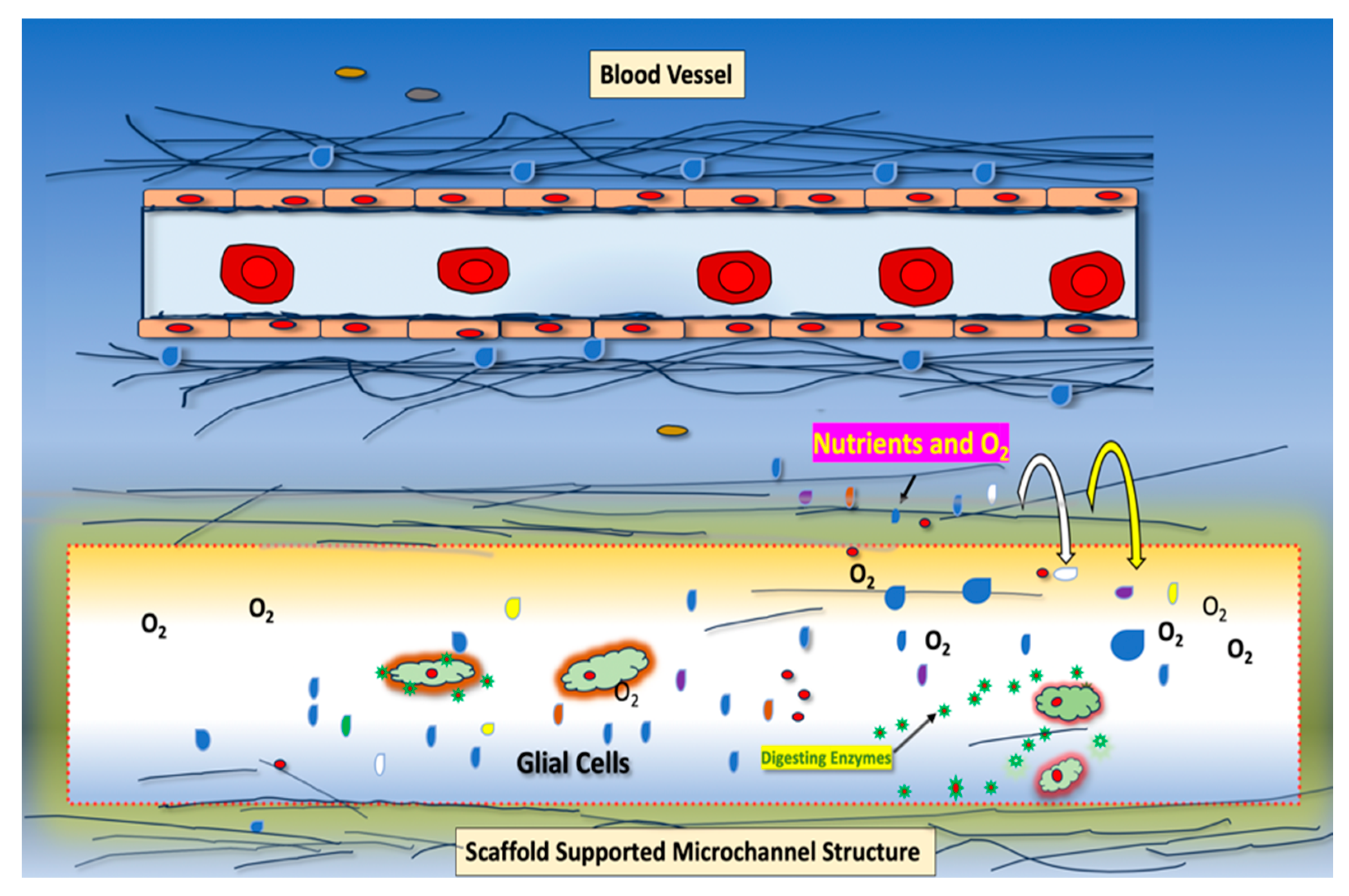
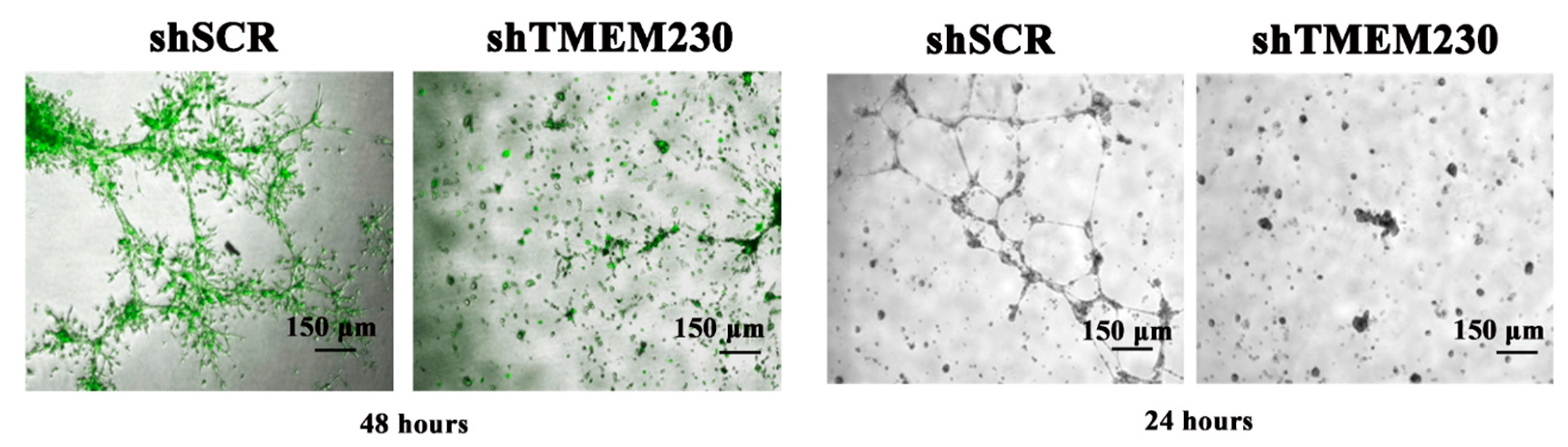
2.2. Endogenous Expression of TMEM230 Promotes “Tunneling”, Microchannel Formation, Extracellular Matrix Infiltration, and Vascular Mimicry
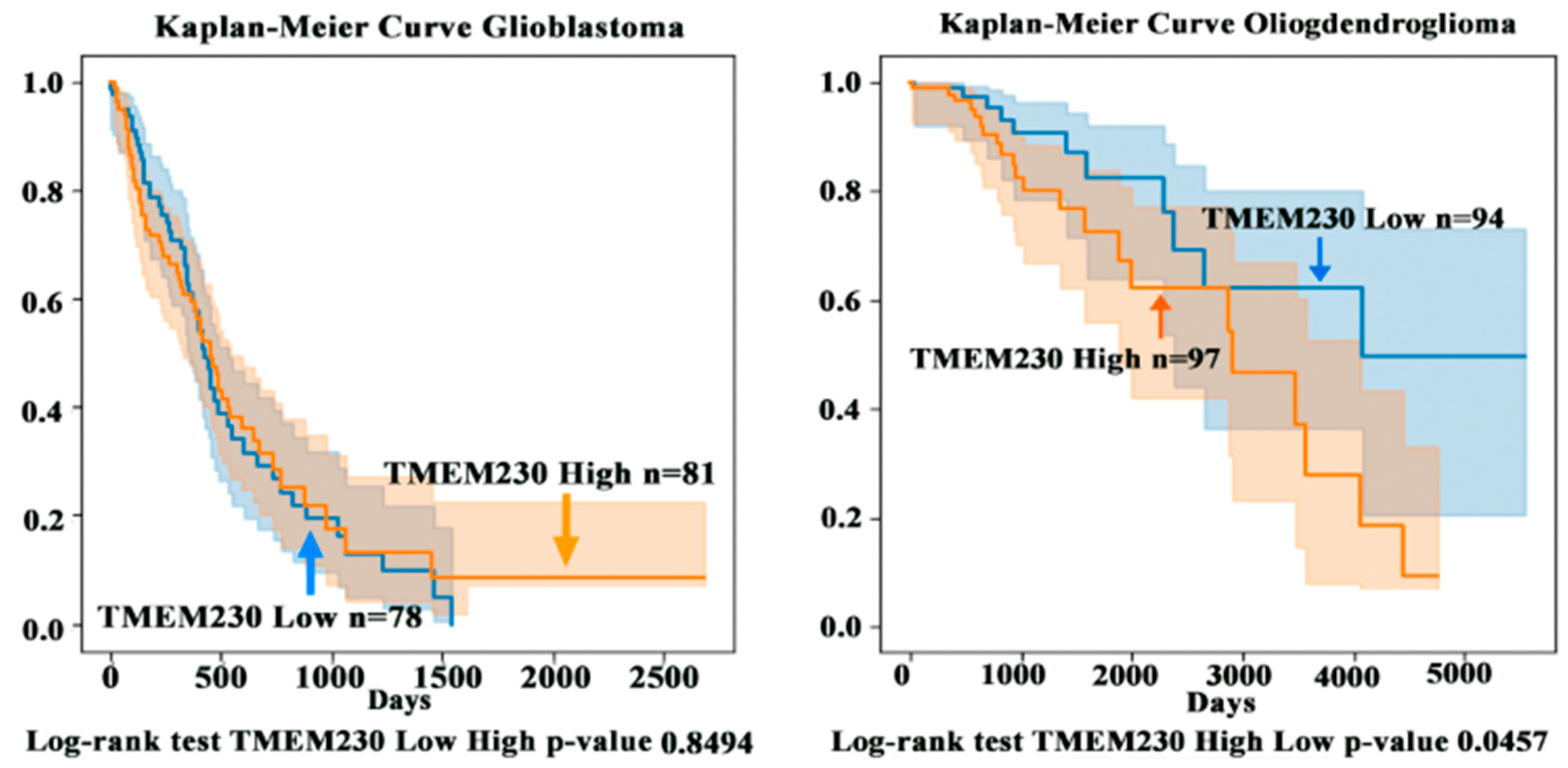
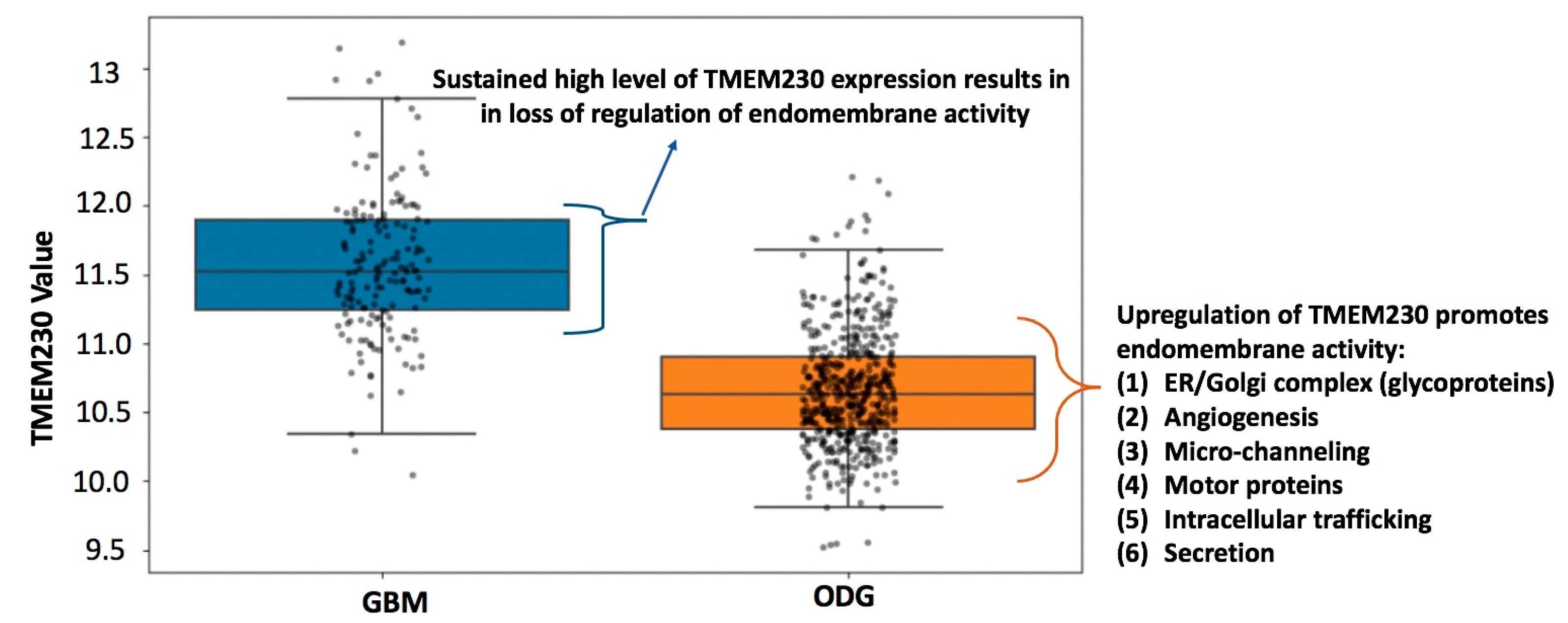
2.3. High Levels of Transmembrane Protein TMEM230 Are Associated with Lower Survivability in Patients with High-Grade Oligodendroglioma
2.4. Candidate Pathways Regulated by TMEM230 in HGG and GBM
2.5. Astrocytoma Patient Data Set Analysis
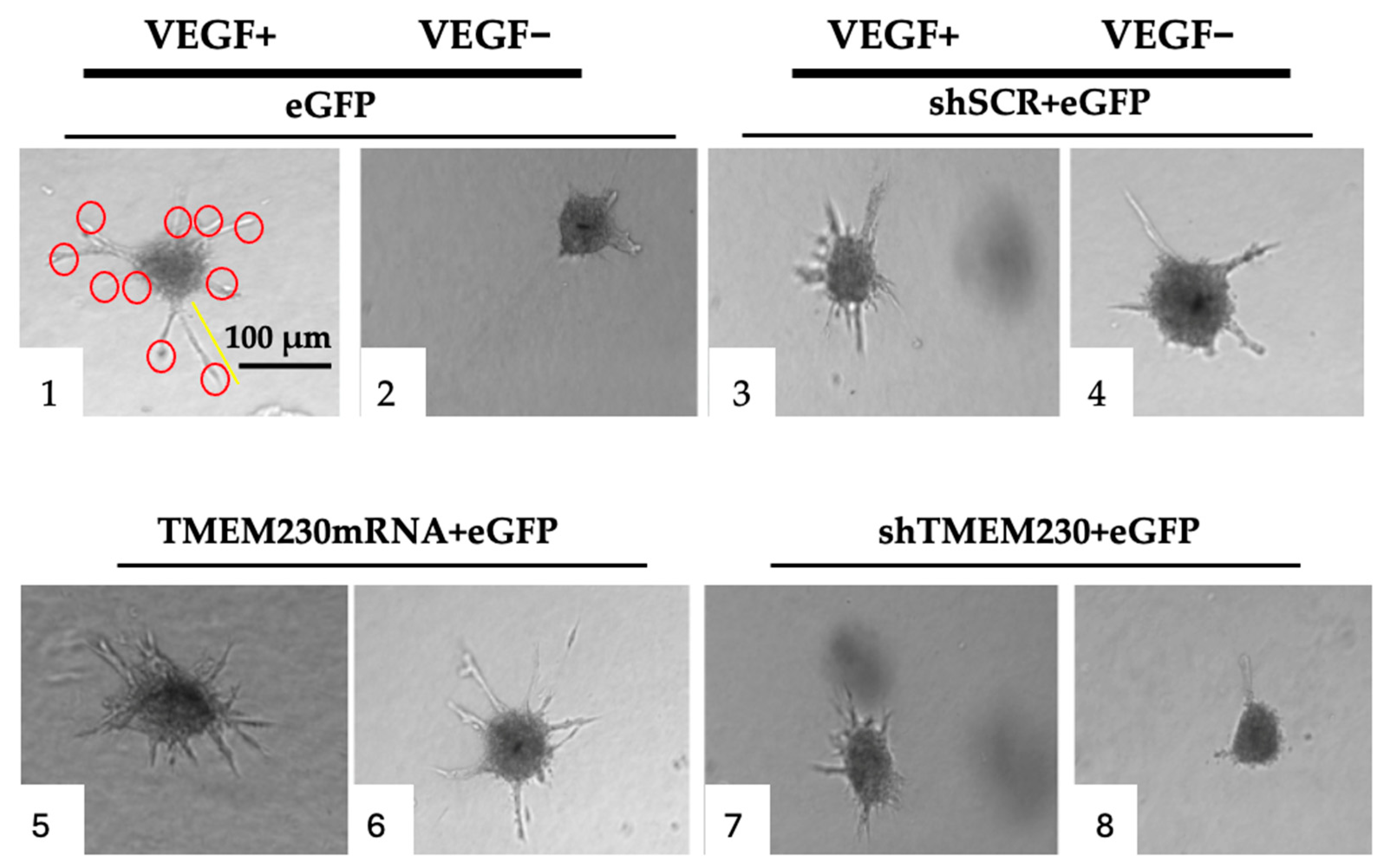
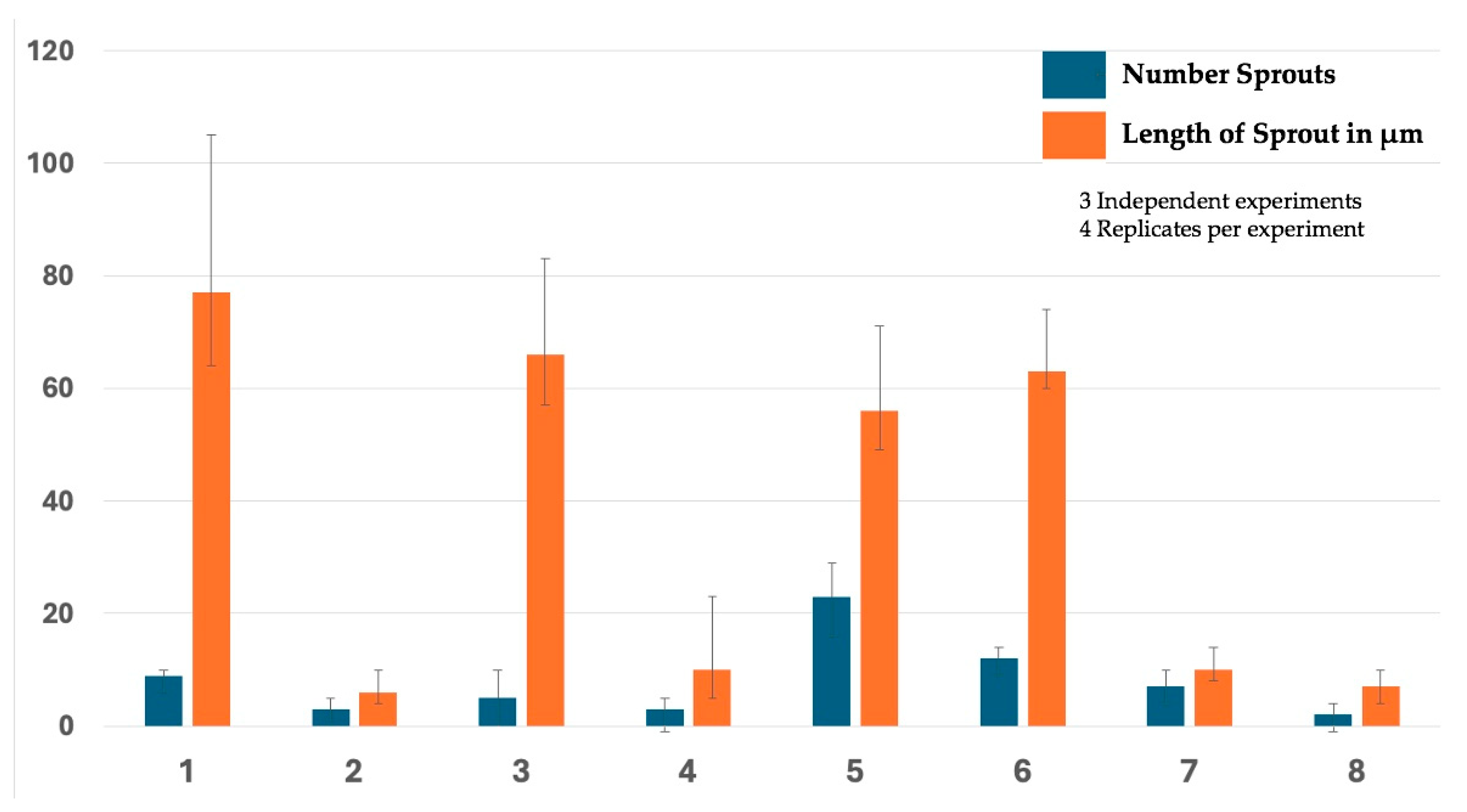
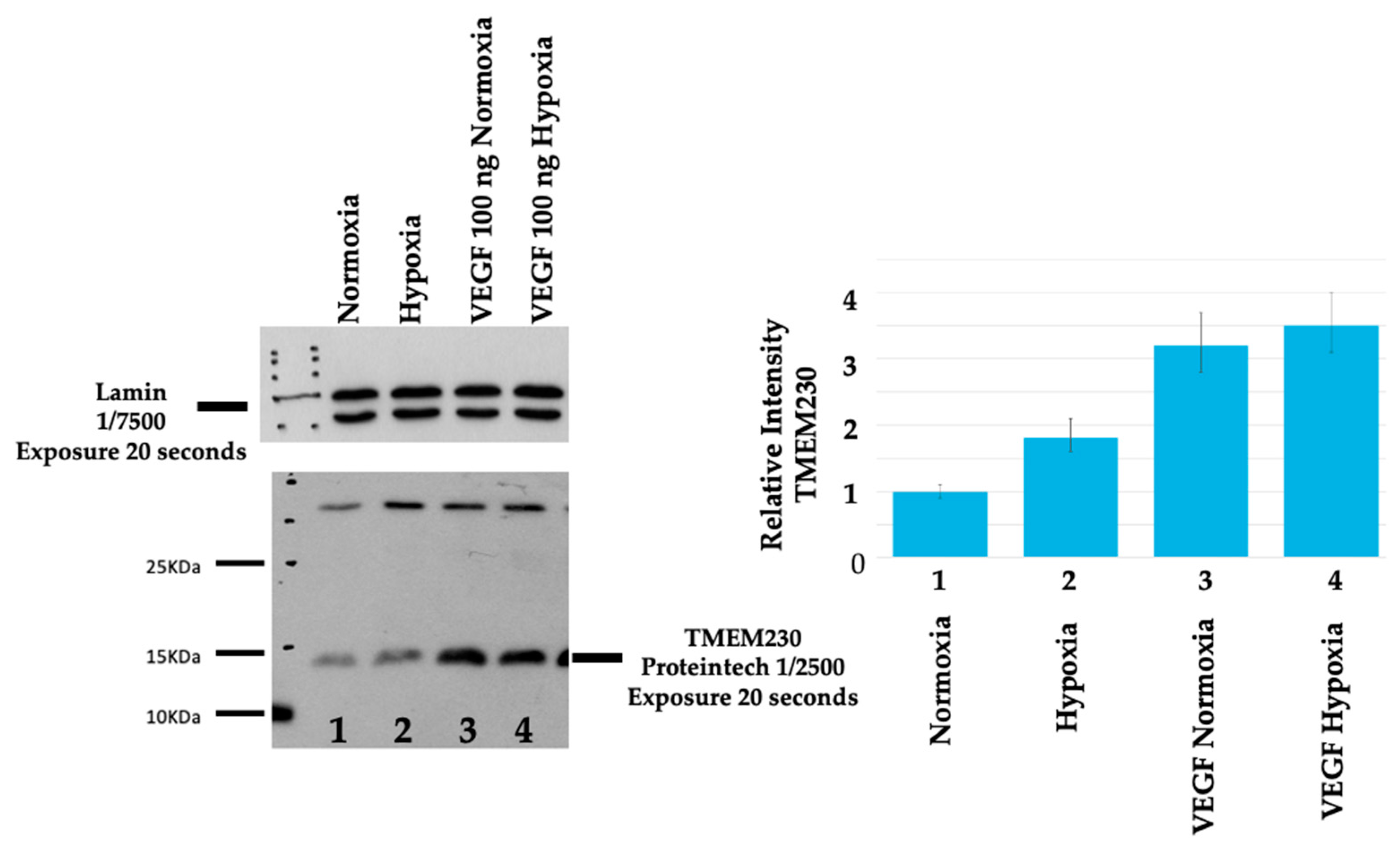
2.6. VEGF Analysis
2.7. Hypoxia Analysis
3. Discussion
4. Materials and Methods
4.1. Patient Data Collection
4.2. Patient RNA-Seq Gene Expression Analysis
4.3. Cloning of Lentiviral-System-Based Construct for Inhibiting TMEM230 Protein Expression
4.4. Generation and Cloning of the Endogenous TMEM230 Variant 2 (ISOFORM 2) Transcript
4.5. Adherent Cell Cultures
4.6. Adherent Co-Cultures of Human Umbilical Vein Endothelial Cells and U87-MG Cells
4.7. Microchannel Formation Assay
4.8. 1321-N1 Microchannel Formation Assay
4.9. Angiogenesis Assays
4.10. Hypoxia Assay
4.11. Western Blotting
4.12. Data Collection and Statistical Analysis
4.13. Microcopy and Imaging
4.14. Supplementary Tables (Excel Files)
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rigby, M.J.; Gomez, T.M.; Puglielli, L. Glial Cell-Axonal Growth Cone Interactions in Neurodevelopment and Regeneration. Front. Neurosci. 2020, 14, 203. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Geng, Z.; Wang, S.; Yu, Z.; Liu, T.; Guan, S.; Du, S.; Zhu, C. The driving mechanism and targeting value of mimicry between vascular endothelial cells and tumor cells in tumor progression. Biomed. Pharmacother. 2023, 165, 115029. [Google Scholar] [CrossRef]
- Quan, L.; Uyeda, A.; Muramatsu, R. Central nervous system regeneration: The roles of glial cells in the potential molecular mechanism underlying remyelination. Inflamm. Regen. 2022, 42, 7. [Google Scholar] [CrossRef]
- Zhou, S.; Xie, M.; Su, J.; Cai, B.; Li, J.; Zhang, K. New insights into balancing wound healing and scarless skin repair. J. Tissue Eng. 2023, 14, 20417314231185848. [Google Scholar] [CrossRef]
- Cocola, C.; Magnaghi, V.; Abeni, E.; Pelucchi, P.; Martino, V.; Vilardo, L.; Piscitelli, E.; Consiglio, A.; Grillo, G.; Mosca, E.; et al. Transmembrane Protein TMEM230, a Target of Glioblastoma Therapy. Front. Cell. Neurosci. 2021, 15, 703431. [Google Scholar] [CrossRef]
- Carra, S.; Sangiorgio, L.; Pelucchi, P.; Cermenati, S.; Mezzelani, A.; Martino, V.; Palizban, M.; Albertini, A.; Götte, M.; Kehler, J.; et al. Zebrafish Tmem230a cooperates with the Delta/Notch signaling pathway to modulate endothelial cell number in angiogenic vessels. J. Cell. Physiol. 2017, 233, 1455–1467. [Google Scholar] [CrossRef] [PubMed]
- Pellerino, A.; Bruno, F.; Soffietti, R.; Rudà, R. Antiangiogenic Therapy for Malignant Brain Tumors: Does It Still Matter? Curr. Oncol. Rep. 2023, 25, 777–785. [Google Scholar] [CrossRef] [PubMed]
- Alafandi, A.; Tbalvandany, S.S.; Arzanforoosh, F.; van Der Voort, S.R.; Incekara, F.; Verhoef, L.; Warnert, E.A.H.; Kruizinga, P.; Smits, M. Probing the glioma microvasculature: A case series of the comparison between perfusion MRI and intraoperative high-frame-rate ultrafast Doppler ultrasound. Eur. Radiol. Exp. 2024, 8, 13. [Google Scholar] [CrossRef]
- Shah, M.; Bin Anis, S.; Yousaf, I.; Bajwa, M.H.; Ali, S.A.; Jamshed, A.; Khattak, S.; Sultan, F.; Yousaf, M.; Siddiqui, N.; et al. Survival Analysis and Correlates with Molecular Epidemiology: 10-year Retrospective Series of High-Grade Glioma in Pakistan. J. Cancer Allied Spéc. 2023, 10, 565. [Google Scholar] [CrossRef]
- Toader, C.; Eva, L.; Costea, D.; Corlatescu, A.D.; Covache-Busuioc, R.-A.; Bratu, B.-G.; Glavan, L.A.; Costin, H.P.; Popa, A.A.; Ciurea, A.V. Low-Grade Gliomas: Histological Subtypes, Molecular Mechanisms, and Treatment Strategies. Brain Sci. 2023, 13, 1700. [Google Scholar] [CrossRef]
- Buonfiglioli, A.; Hambardzumyan, D. Macrophages and microglia: The cerberus of glioblastoma. Acta Neuropathol. Commun. 2021, 9, 54. [Google Scholar] [CrossRef] [PubMed]
- Nóbrega, A.H.L.; Pimentel, R.S.; Prado, A.P.; Garcia, J.; Frozza, R.L.; Bernardi, A. Neuroinflammation in Glioblastoma: The Role of the Microenvironment in Tumour Progression. Curr. Cancer Drug Targets 2024, 24, 1–16. [Google Scholar] [CrossRef]
- Hu, L.S.; Hawkins-Daarud, A.; Wang, L.; Li, J.; Swanson, K.R. Imaging of intratumoral heterogeneity in high-grade glioma. Cancer Lett. 2020, 477, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Nale, V.; Chiodi, A.; Di Nanni, N.; Cifola, I.; Moscatelli, M.; Cocola, C.; Gnocchi, M.; Piscitelli, E.; Sula, A.; Zucchi, I.; et al. scMuffin: An R package to disentangle solid tumor heterogeneity by single-cell gene expression analysis. BMC Bioinform. 2023, 24, 1–18. [Google Scholar] [CrossRef]
- Abdel-Rahman, S.A.; Gabr, M. Small Molecule Immunomodulators as Next-Generation Therapeutics for Glioblastoma. Cancers 2024, 16, 435. [Google Scholar] [CrossRef]
- Abdullaeva, S.; Chubarev, V.; Valeeva, A.; Preferanskaya, N.; Neganova, M.; Smolyarchuk, E.; Liu, J.; Sukocheva, O.; Samsonov, M.; Alyautdin, R. Analysis of Clinical Success and Molecular Mechanisms of Action of Novel Anti-glioblastoma Drugs: A Review. Curr. Med. Chem. 2024. [Google Scholar] [CrossRef]
- O’Neill, K.; Syed, N.; Crook, T.; Dubey, S.; Potharaju, M.; Limaye, S.; Ranade, A.; Anichini, G.; Patil, D.; Datta, V.; et al. Profiling of circulating glial cells for accurate blood-based diagnosis of glial malignancies. Int. J. Cancer 2023, 154, 1298–1308. [Google Scholar] [CrossRef] [PubMed]
- Borenstein, J.T.; Tupper, M.M.; Mack, P.J.; Weinberg, E.J.; Khalil, A.S.; Hsiao, J.; García-Cardeña, G. Functional endothelialized microvascular networks with circular cross-sections in a tissue culture substrate. Biomed. Microdevices 2009, 12, 71–79. [Google Scholar] [CrossRef]
- Yang, K.; Wu, Z.; Zhang, H.; Zhang, N.; Wu, W.; Wang, Z.; Dai, Z.; Zhang, X.; Zhang, L.; Peng, Y.; et al. Glioma targeted therapy: Insight into future of molecular approaches. Mol. Cancer 2022, 21, 39. [Google Scholar] [CrossRef]
- Delobel, T.; Ayala-Hernández, L.E.; Bosque, J.J.; Pérez-Beteta, J.; Chulián, S.; García-Ferrer, M.; Piñero, P.; Schucht, P.; Murek, M.; Pérez-García, V.M. Overcoming chemotherapy resistance in low-grade gliomas: A computational approach. PLOS Comput. Biol. 2023, 19, e1011208. [Google Scholar] [CrossRef]
- Liu, Z.-L.; Chen, H.-H.; Zheng, L.-L.; Sun, L.-P.; Shi, L. Angiogenic signaling pathways and anti-angiogenic therapy for cancer. Signal Transduct. Target. Ther. 2023, 8, 198. [Google Scholar] [CrossRef] [PubMed]
- Silvestre, D.C.; Maccioni, H.J.; Caputto, B.L. Content of endoplasmic reticulum and Golgi complex membranes positively correlates with the proliferative status of brain cells. J. Neurosci. Res. 2009, 87, 857–865. [Google Scholar] [CrossRef] [PubMed]
- Hellicar, J.; Stevenson, N.L.; Stephens, D.J.; Lowe, M. Supply chain logistics—The role of the Golgi complex in extracellular matrix production and maintenance. J. Cell Sci. 2022, 135, jcs258879. [Google Scholar] [CrossRef] [PubMed]
- Spano, D.; Colanzi, A. Golgi Complex: A Signaling Hub in Cancer. Cells 2022, 11, 1990. [Google Scholar] [CrossRef] [PubMed]
- Mellman, I.; Nelson, W.J. Coordinated protein sorting, targeting and distribution in polarized cells. Nat. Rev. Mol. Cell Biol. 2008, 9, 833–845. [Google Scholar] [CrossRef] [PubMed]
- Gandalovičová, A.; Vomastek, T.; Rosel, D.; Brábek, J. Cell polarity signaling in the plasticity of cancer cell invasiveness. Oncotarget 2016, 7, 25022–25049. [Google Scholar] [CrossRef] [PubMed]
- Elgendy, M.; Tamada, H.; Taira, T.; Iio, Y.; Kawamura, A.; Kunogi, A.; Mizutani, Y.; Kiyama, H. Dynamic changes in endoplasmic reticulum morphology and its contact with the plasma membrane in motor neurons in response to nerve injury. Cell Tissue Res. 2024, 1–14. [Google Scholar] [CrossRef]
- Harvey, C.; Weinreich, M.; Lee, J.A.; Shaw, A.C.; Ferraiuolo, L.; Mortiboys, H.; Zhang, S.; Hop, P.J.; Zwamborn, R.A.; van Eijk, K.; et al. Rare and common genetic determinants of mitochondrial function determine severity but not risk of amyotrophic lateral sclerosis. Heliyon 2024, 10, e24975. [Google Scholar] [CrossRef] [PubMed]
- Zucchi, I.; Bini, L.; Valaperta, R.; Ginestra, A.; Albani, D.; Susani, L.; Sanchez, J.C.; Liberatori, S.; Magi, B.; Raggiaschi, R.; et al. Proteomic dissection of dome formation in a mammary cell line: Role of tropomyosin-5b and maspin. Proc. Natl. Acad. Sci. USA 2001, 98, 5608–5613. [Google Scholar] [CrossRef]
- Hey, S.; Linder, S. Matrix metalloproteinases at a glance. J. Cell Sci. 2024, 137, 4015–4016. [Google Scholar] [CrossRef]
- Winkler, J.; Abisoye-Ogunniyan, A.; Metcalf, K.J.; Werb, Z. Concepts of extracellular matrix remodelling in tumour progression and metastasis. Nat. Commun. 2020, 11, 5120. [Google Scholar] [CrossRef] [PubMed]
- Tewari, B.P.; Chaunsali, L.; Prim, C.E.; Sontheimer, H. A glial perspective on the extracellular matrix and perineuronal net remodeling in the central nervous system. Front. Cell. Neurosci. 2022, 16, 1022754. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.-W.; Allen, G.I.; Liu, Z. TCGA2STAT: Simple TCGA data access for integrated statistical analysis in R. Bioinformatics 2015, 32, 952–954. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cocola, C.; Abeni, E.; Martino, V.; Piscitelli, E.; Pelucchi, P.; Mosca, E.; Chiodi, A.; Mohamed, T.; Palizban, M.; Porta, G.; et al. Transmembrane Protein TMEM230, Regulator of Glial Cell Vascular Mimicry and Endothelial Cell Angiogenesis in High-Grade Heterogeneous Infiltrating Gliomas and Glioblastoma. Int. J. Mol. Sci. 2024, 25, 3967. https://doi.org/10.3390/ijms25073967
Cocola C, Abeni E, Martino V, Piscitelli E, Pelucchi P, Mosca E, Chiodi A, Mohamed T, Palizban M, Porta G, et al. Transmembrane Protein TMEM230, Regulator of Glial Cell Vascular Mimicry and Endothelial Cell Angiogenesis in High-Grade Heterogeneous Infiltrating Gliomas and Glioblastoma. International Journal of Molecular Sciences. 2024; 25(7):3967. https://doi.org/10.3390/ijms25073967
Chicago/Turabian StyleCocola, Cinzia, Edoardo Abeni, Valentina Martino, Eleonora Piscitelli, Paride Pelucchi, Ettore Mosca, Alice Chiodi, Tasnim Mohamed, Mira Palizban, Giovanni Porta, and et al. 2024. "Transmembrane Protein TMEM230, Regulator of Glial Cell Vascular Mimicry and Endothelial Cell Angiogenesis in High-Grade Heterogeneous Infiltrating Gliomas and Glioblastoma" International Journal of Molecular Sciences 25, no. 7: 3967. https://doi.org/10.3390/ijms25073967
APA StyleCocola, C., Abeni, E., Martino, V., Piscitelli, E., Pelucchi, P., Mosca, E., Chiodi, A., Mohamed, T., Palizban, M., Porta, G., Palizban, H., Nano, G., Acquati, F., Bruno, A., Greve, B., Gerovska, D., Magnaghi, V., Mazzaccaro, D., Bertalot, G., ... Reinbold, R. A. (2024). Transmembrane Protein TMEM230, Regulator of Glial Cell Vascular Mimicry and Endothelial Cell Angiogenesis in High-Grade Heterogeneous Infiltrating Gliomas and Glioblastoma. International Journal of Molecular Sciences, 25(7), 3967. https://doi.org/10.3390/ijms25073967