
Overexpression of GmPAP4 Enhances Symbiotic Nitrogen Fixation and Seed Yield in Soybean under Phosphorus-Deficient Condition

 ,
,
Abstract
1. Introduction
2. Results
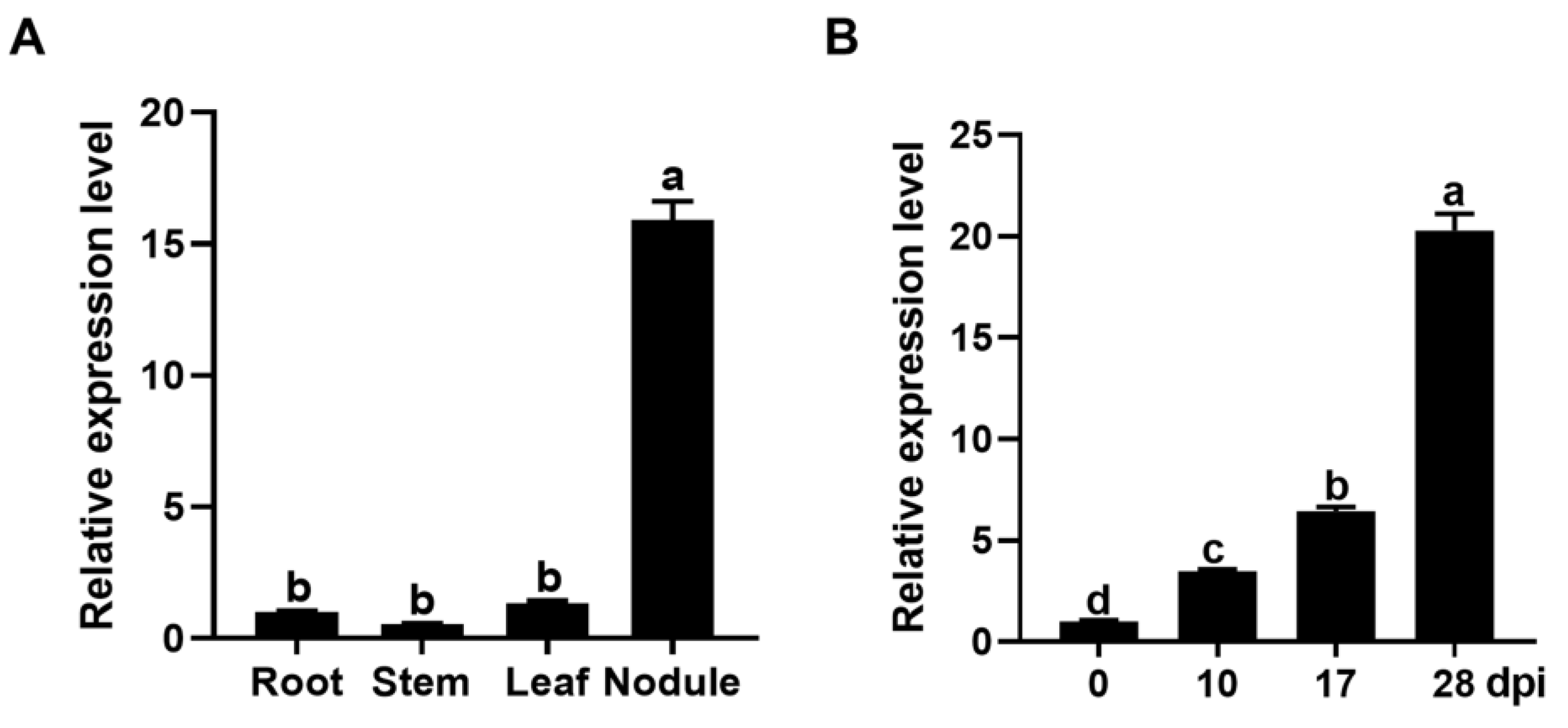
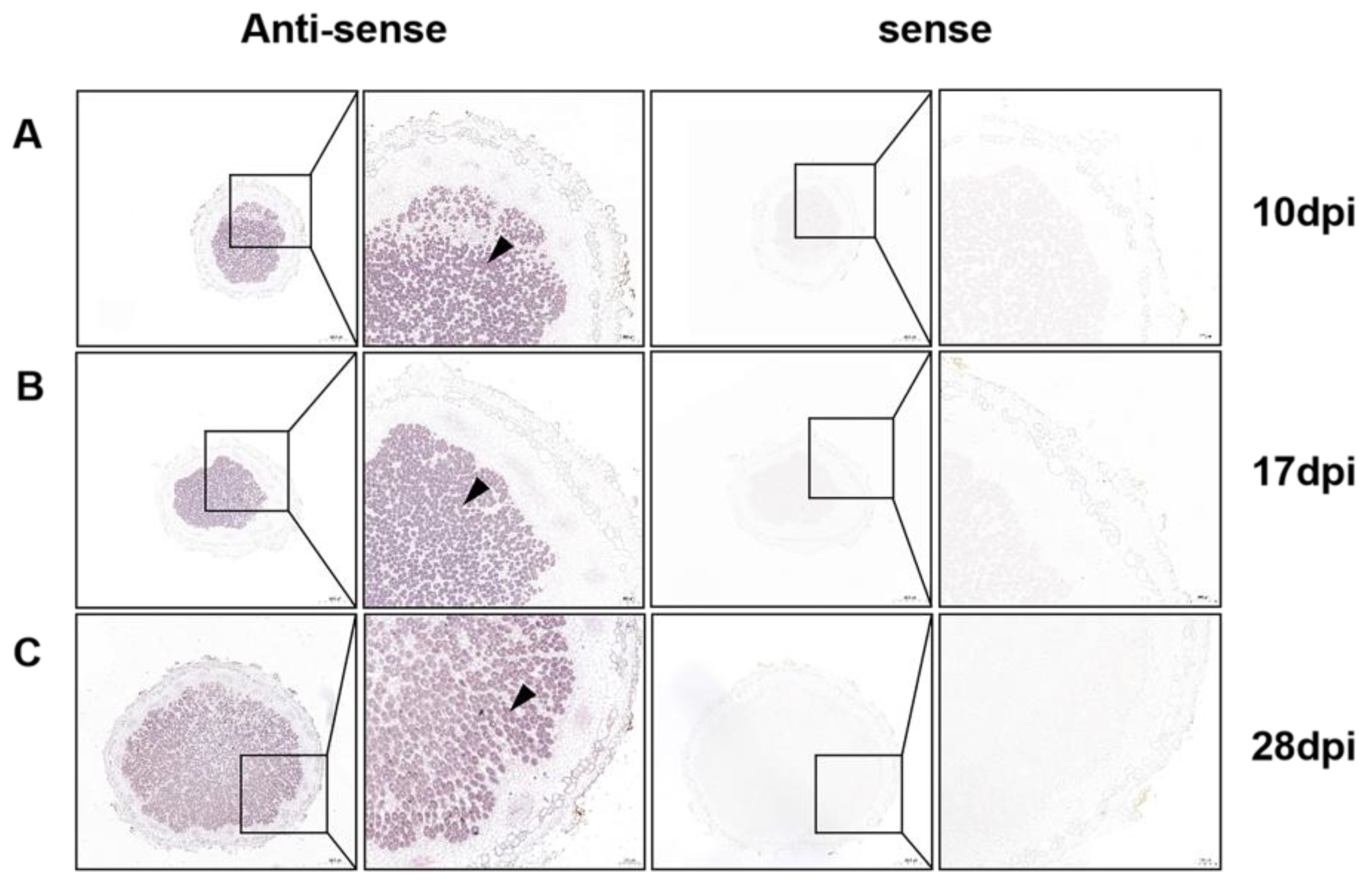
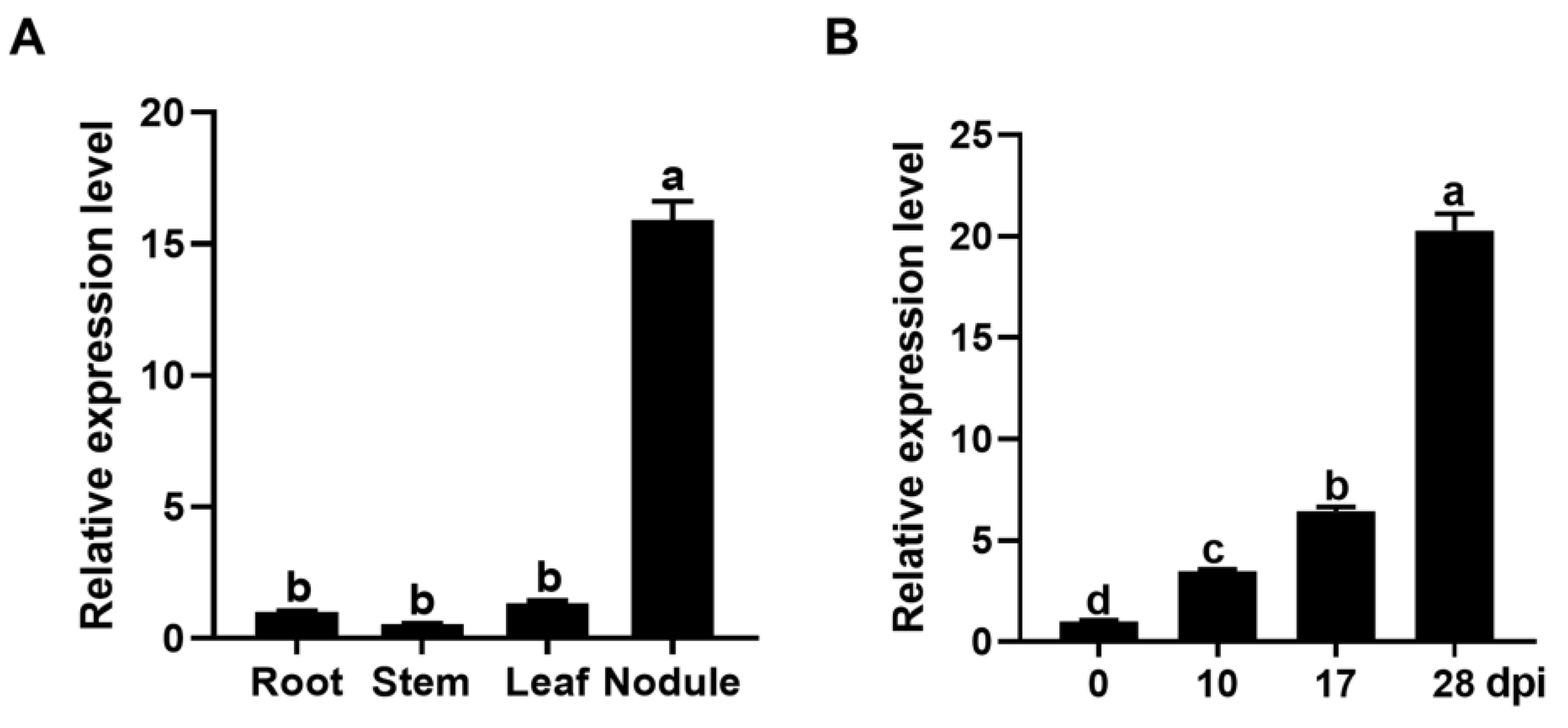
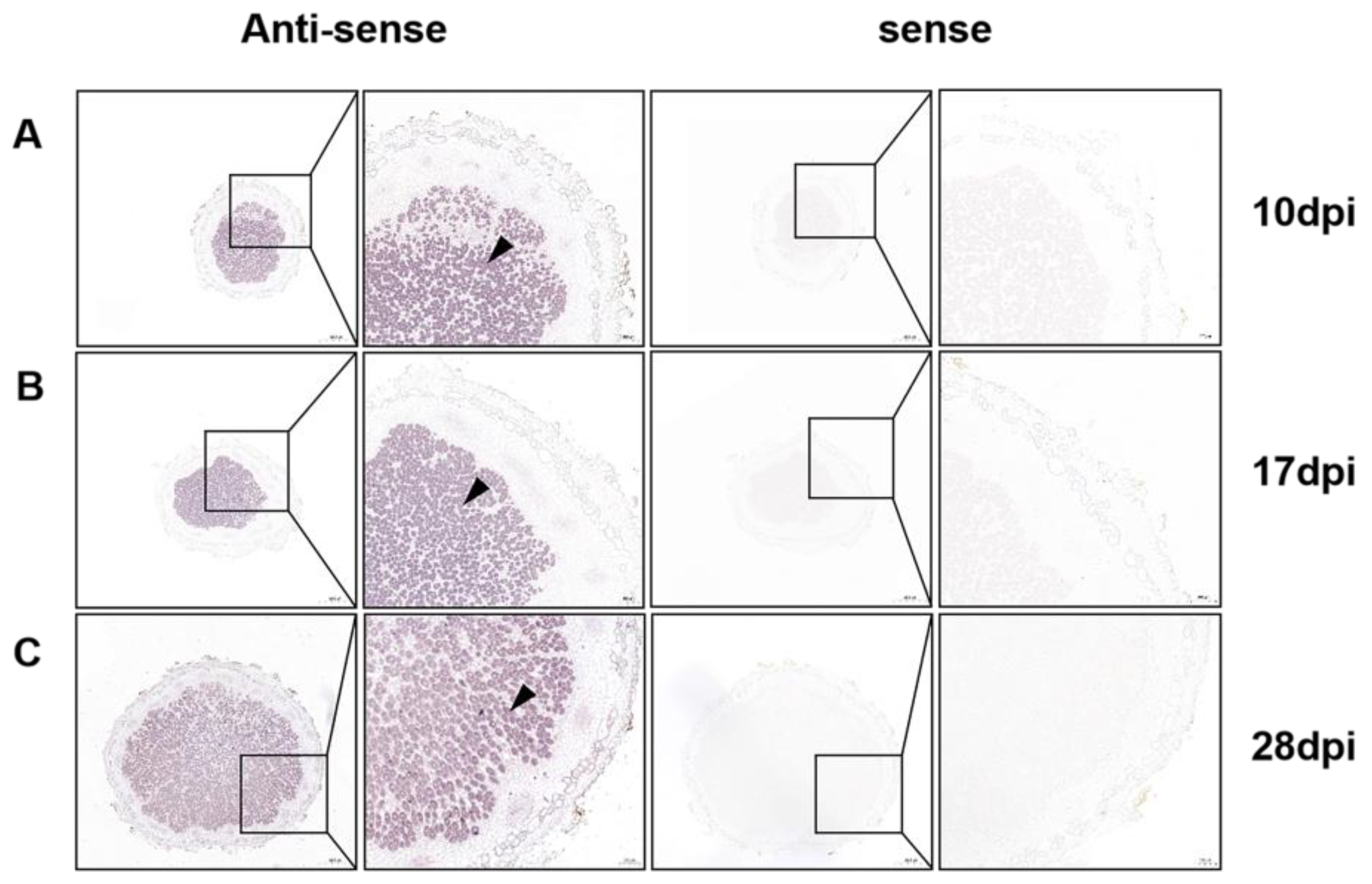
2.1. GmPAP4 Was Preferentially Expressed in Infection Zones of Nodules in Soybean
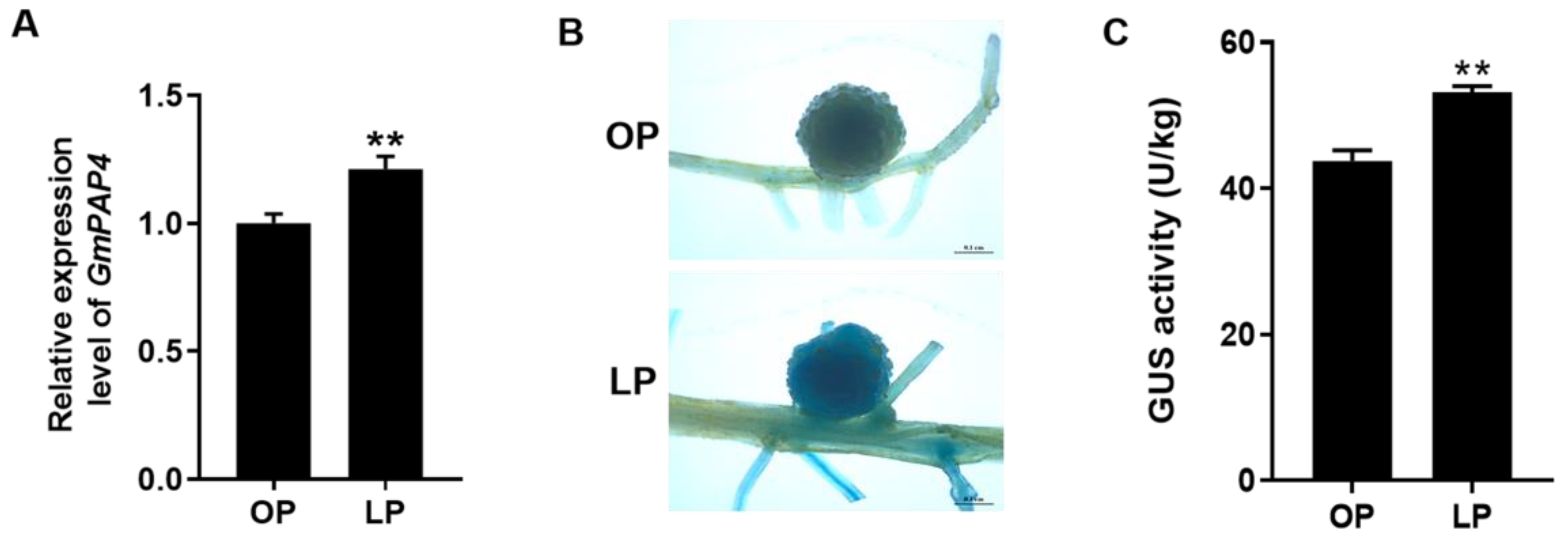
2.2. GmPAP4 Was Highly Induced by Low Phosphorus Stress in Soybean Nodules
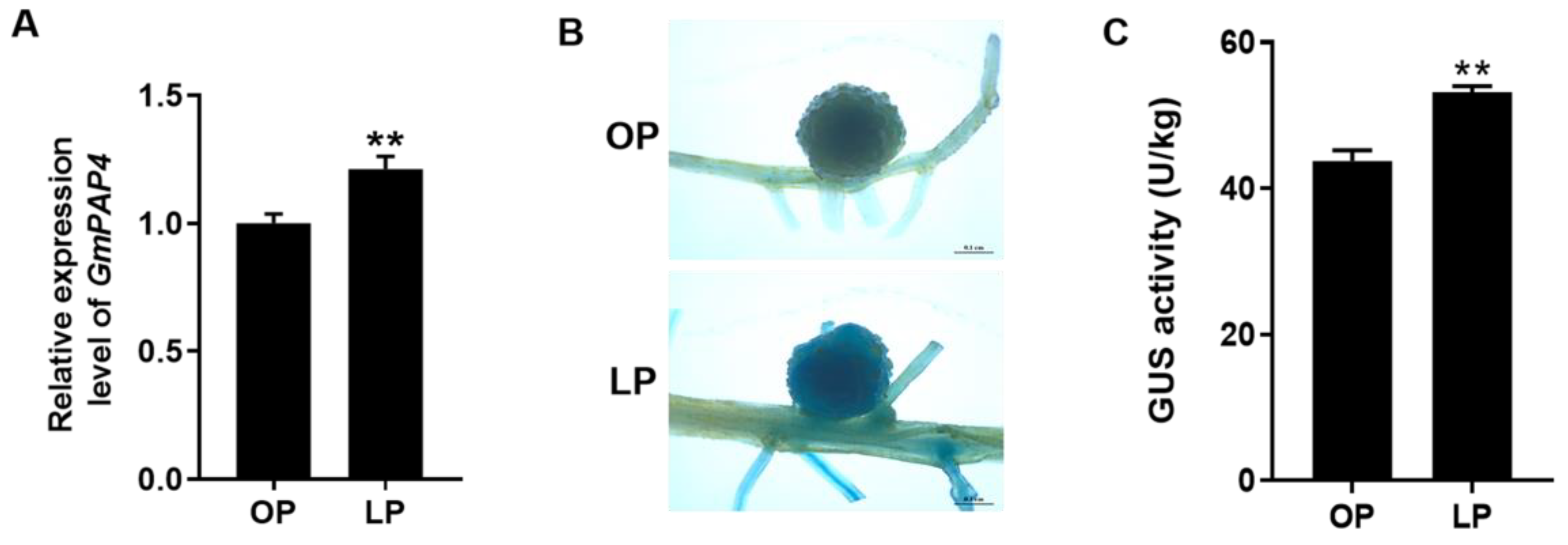
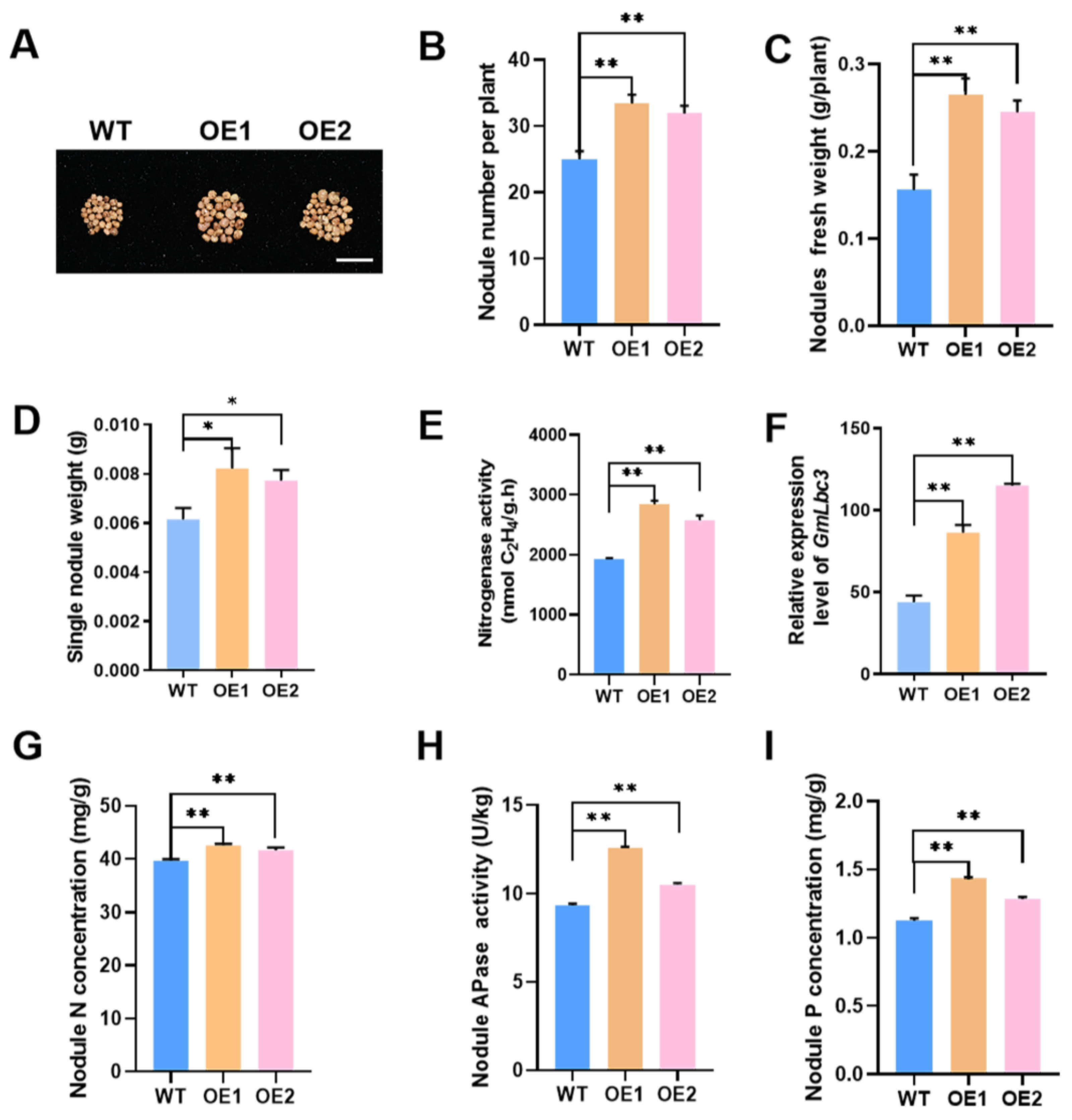
2.3. GmPAP4 Expression Was Positively Associated with Nodulation and Nitrogen Fixation under LP Stress
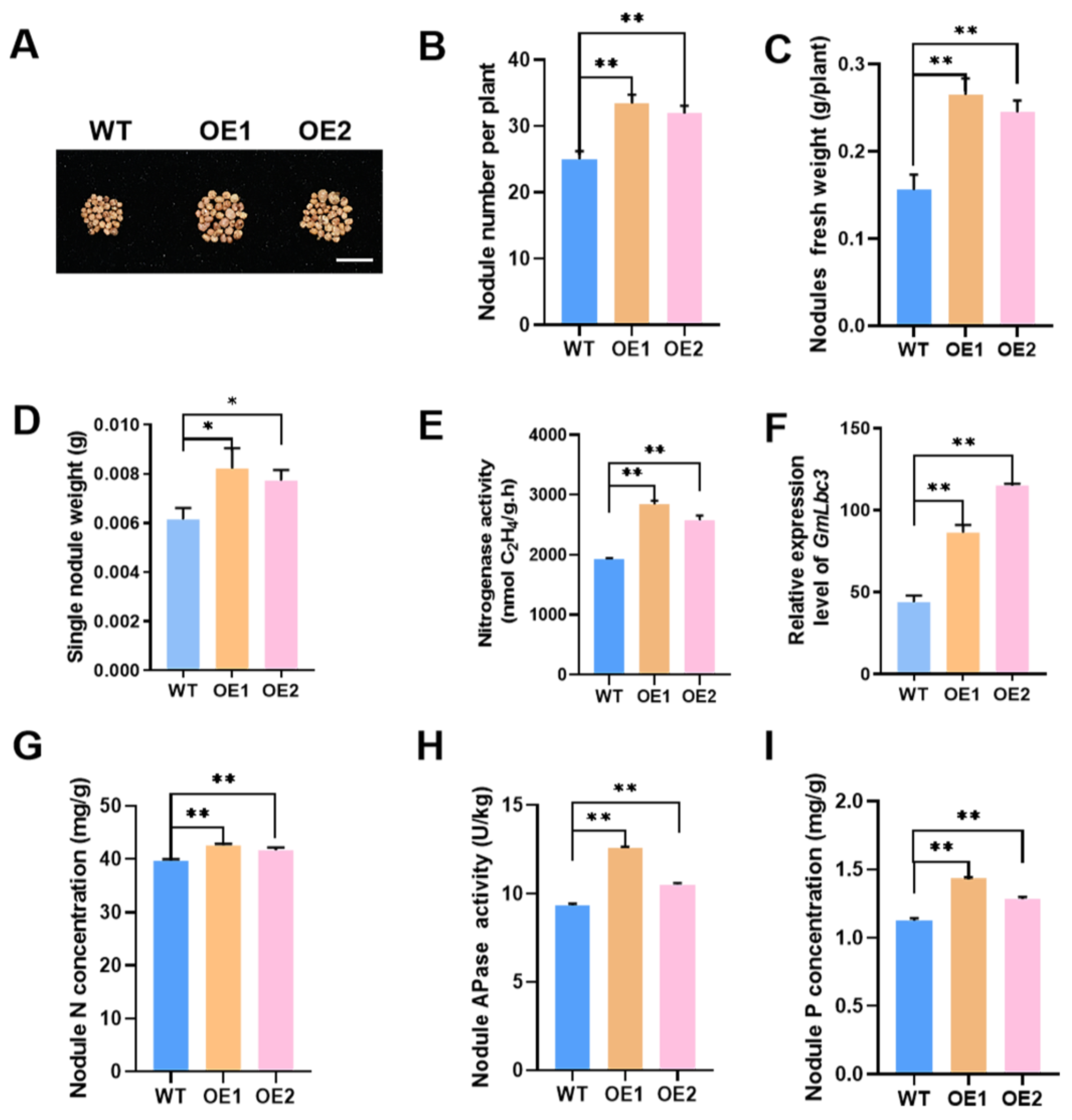
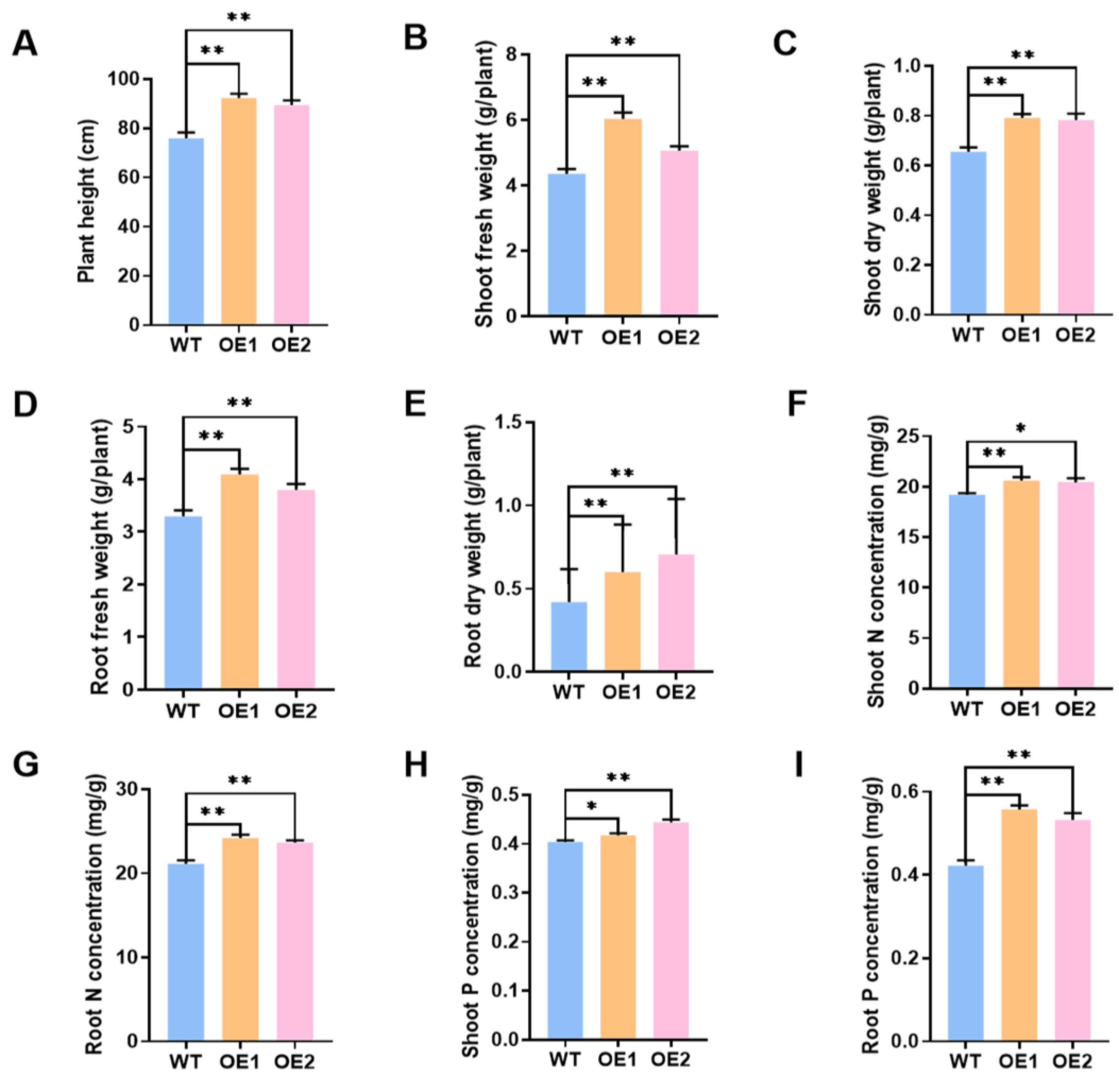
2.4. Overexpression of GmPAP4-Enhanced Soybean Plant Growth under P-Deficient Conditions
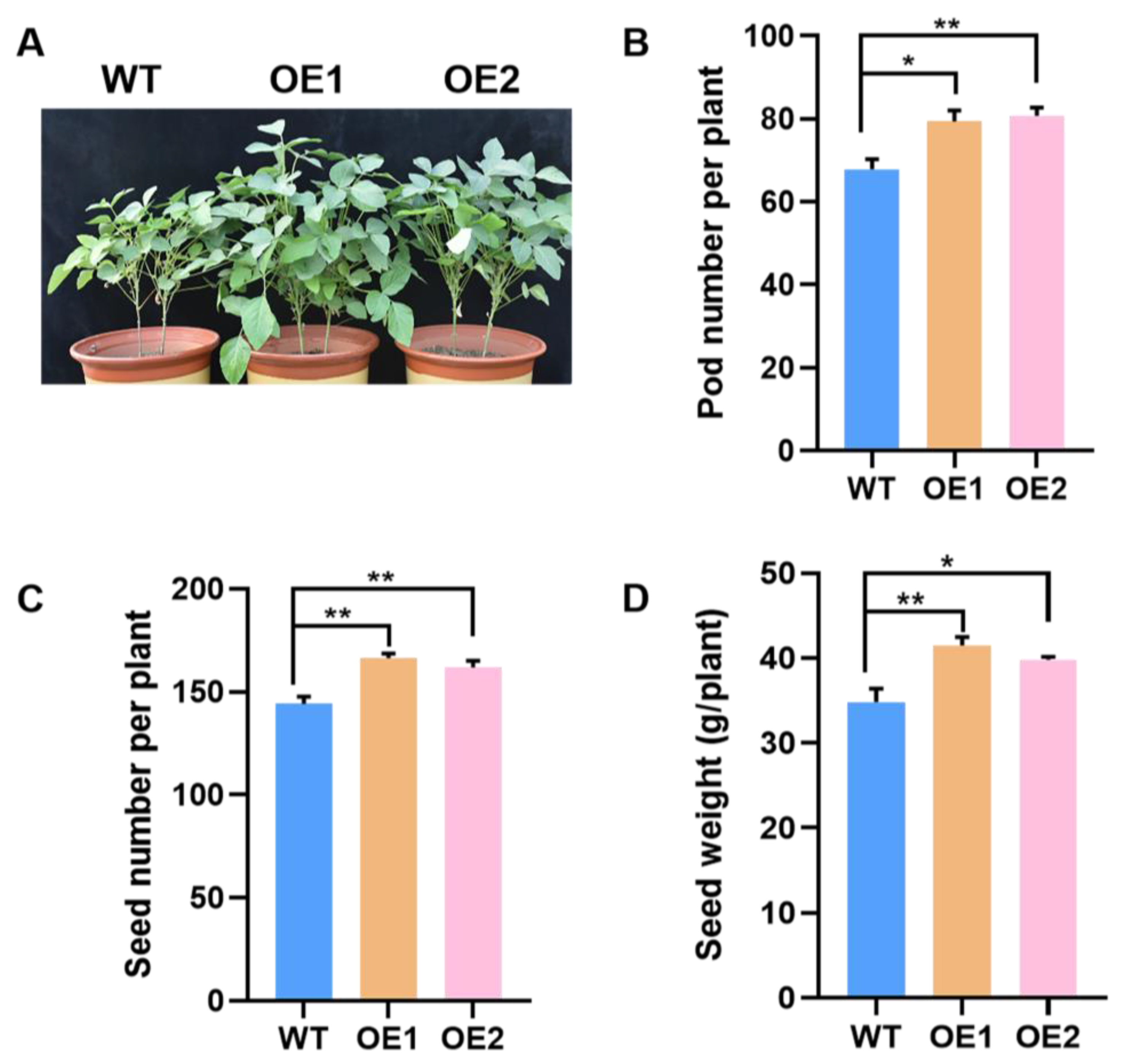
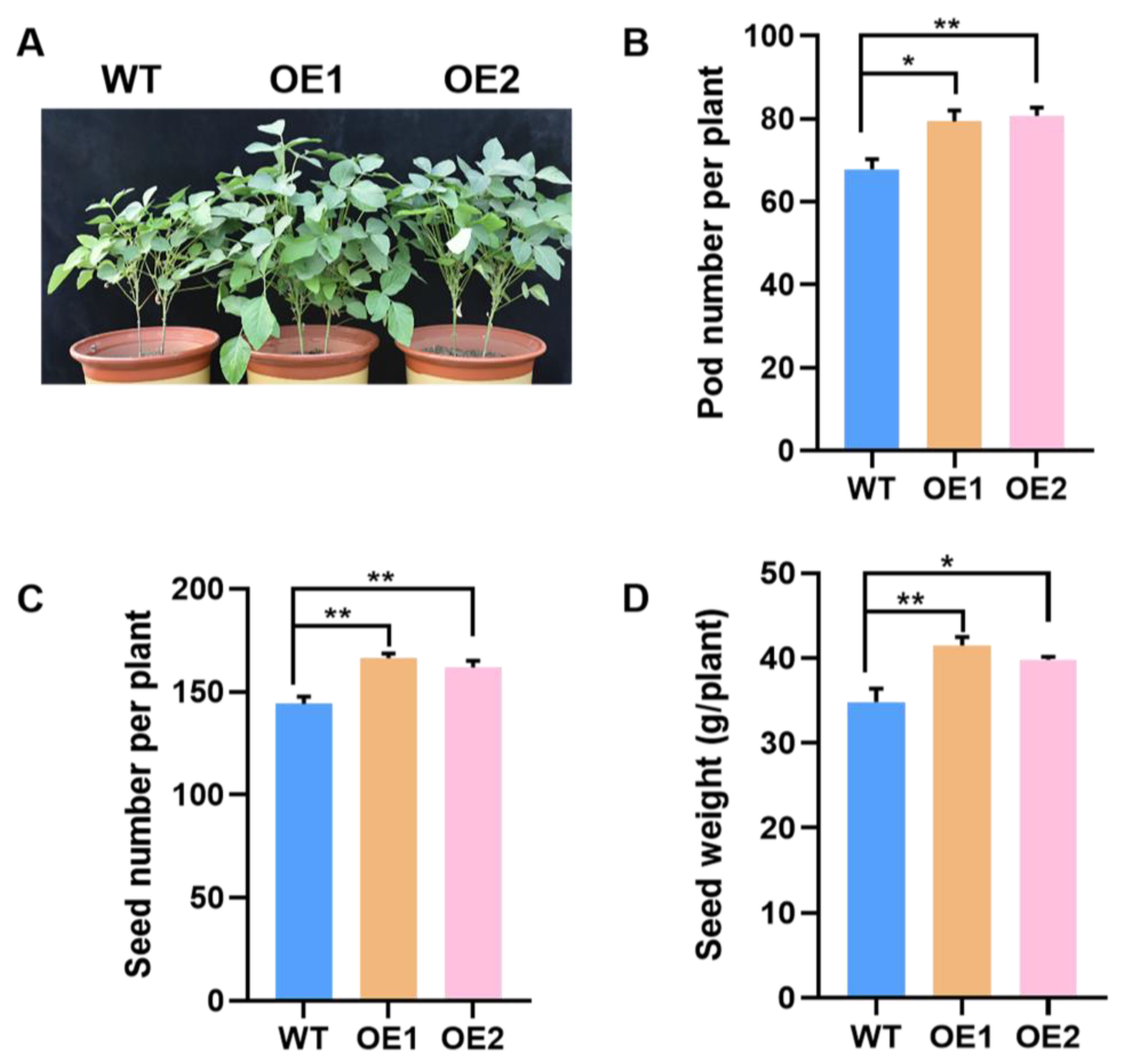
2.5. Overexpression of GmPAP4 Enhanced Soybean Yield under P-Deficient Conditions
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Plant mRNA Isolation and Quantitative Real-Time PCR (qRT-PCR)
4.3. Histochemical GUS Staining Analysis
4.4. Generation of Stable Transgenic Soybean Plants
4.5. RNA In Situ Hybridization Analysis
4.6. Toluidine Blue Staining
4.7. Measurement of N and P Contents
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Masson-Boivin, C.; Sachs, J.L. Symbiotic nitrogen fixation by rhizobia—The roots of a success story. Curr. Opin. Plant Biol. 2018, 44, 7–15. [Google Scholar] [CrossRef]
- Werner, G.D.A.; Cornwell, W.K.; Sprent, J.I.; Kattge, J.; Kiers, E.T. A single evolutionary innovation drives the deep evolution of symbiotic N-fixation in angiosperms. Nat. Commun. 2014, 5, 4087. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, B.J.; Indrasumunar, A.; Hayashi, S.; Lin, M.H.; Lin, Y.H.; Reid, D.E.; Gresshoff, P.M. Molecular analysis of legume nodule development and autoregulation. J. Integr. Plant Biol. 2010, 52, 61–76. [Google Scholar] [CrossRef]
- Oldroyd, G.E.; Downie, J.A. Coordinating nodule morphogenesis with rhizobial infection in legumes. Annu. Rev. Plant Biol. 2008, 59, 519–546. [Google Scholar] [CrossRef] [PubMed]
- Collier, R.; Tegeder, M. Soybean ureide transporters play a critical role in nodule development, function and nitrogen export. Plant J. 2012, 72, 355–367. [Google Scholar] [CrossRef]
- Marx, H.; Minogue, C.E.; Jayaraman, D.; Richards, A.L.; Kwiecien, N.W.; Siahpirani, A.F.; Rajasekar, S.; Maeda, J.; Garcia, K.; Del Valle-Echevarria, A.R.; et al. A proteomic atlas of the legume Medicago truncatula and its nitrogen-fixing endosymbiont Sinorhizobium meliloti. Nat. Biotechnol. 2016, 34, 1198–1205. [Google Scholar] [CrossRef] [PubMed]
- Yates, R.J.; Howieson, J.G.; Reeve, W.G.; O’Hara, G.W. A re-appraisal of the biology and terminology describing rhizobial strain success in nodule occupancy of legumes in agriculture. Plant Soil 2011, 348, 255–267. [Google Scholar] [CrossRef]
- Makoudi, B.; Kabbadj, A.; Mouradi, M.; Amenc, L.; Domergue, O.; Blair, M.; Drevon, J.J.; Ghoulam, C. Phosphorus deficiency increases nodule phytase activity of faba bean–rhizobia symbiosis. Acta Physiol. Plant. 2018, 40, 63. [Google Scholar] [CrossRef]
- Valentine, A.J.; Kleinert, A.; Benedito, V.A. Adaptive strategies for nitrogen metabolism in phosphate deficient legume nodules. Plant Sci. 2017, 256, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Cabeza, R.A.; Liese, R.; Lingner, A.; von Stieglitz, I.; Neumann, J.; Salinas-Riester, G.; Pommerenke, C.; Dittert, K.; Schulze, J. RNA-seq transcriptome profiling reveals that Medicago truncatula nodules acclimate N-2 fixation before emerging P deficiency reaches the nodules. J. Exp. Bot. 2014, 65, 6035–6048. [Google Scholar] [CrossRef]
- Chen, L.; Qin, L.; Zhou, L.; Li, X.; Chen, Z.; Sun, L.; Wang, W.; Lin, Z.; Zhao, J.; Yamaji, N.; et al. A nodule-localized phosphate transporter GmPT7 plays an important role in enhancing symbiotic N2 fixation and yield in soybean. New Phytol. 2019, 221, 2013–2025. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhao, J.; Tan, Z.; Zeng, R.; Liao, H. GmEXPB2, a Cell Wall beta-Expansin, Affects Soybean Nodulation through Modifying Root Architecture and Promoting Nodule Formation and Development. Plant Physiol. 2015, 169, 2640–2653. [Google Scholar] [PubMed]
- Sulieman, S.; Tran, L.S. Phosphorus homeostasis in legume nodules as an adaptive strategy to phosphorus deficiency. Plant Sci. 2015, 239, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, Z.; Kong, Y.; Li, X.; Li, W.; Du, H.; Zhang, C. GmPAP12 Is Required for Nodule Development and Nitrogen Fixation under Phosphorus Starvation in Soybean. Front. Plant Sci. 2020, 11, 450. [Google Scholar] [CrossRef] [PubMed]
- Tran, H.T.; Qian, W.Q.; Hurley, B.A.; She, Y.M.; Wang, D.W.; Plaxton, W.C. Biochemical and molecular characterization of AtPAP12 and AtPAP26: The predominant purple acid phosphatase isozymes secreted by phosphate-starved Arabidopsis thaliana. Plant Cell Environ. 2010, 33, 1789–1803. [Google Scholar] [CrossRef] [PubMed]
- Hur, Y.J.; Jin, B.R.; Nam, J.; Chung, Y.S.; Lee, J.H.; Choi, H.K.; Yun, D.J.; Yi, G.; Kim, Y.H.; Kim, D.H. Molecular characterization of OsPAP2: Transgenic expression of a purple acid phosphatase up-regulated in phosphate-deprived rice suspension cells. Biotechnol. Lett. 2010, 32, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.; Li, X.; Ma, J.; Li, W.; Yan, G.; Zhang, C. GmPAP4, a novel purple acid phosphatase gene isolated from soybean (Glycine max), enhanced extracellular phytate utilization in Arabidopsis thaliana. Plant Cell Rep. 2014, 33, 655–667. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Tian, J.; Lam, H.M.; Lim, B.L.; Yan, X.; Liao, H. Biochemical and molecular characterization of PvPAP3, a novel purple acid phosphatase isolated from common bean enhancing extracellular ATP utilization. Plant Physiol. 2010, 152, 854–865. [Google Scholar] [CrossRef]
- Zhu, S.; Chen, M.; Liang, C.; Xue, Y.; Lin, S.; Tian, J. Characterization of Purple Acid Phosphatase Family and Functional Analysis of GmPAP7a/7b Involved in Extracellular ATP Utilization in Soybean. Front. Plant Sci. 2020, 11, 661. [Google Scholar] [CrossRef]
- Xu, H.Q.; Zhang, H.Y.; Fan, Y.K.; Wang, R.Y.; Cui, R.F.; Liu, X.Q.; Chu, S.S.; Jiao, Y.Q.; Zhang, X.G.; Zhang, D. The purple acid phosphatase GmPAP17 predominantly enhances phosphorus use efficiency in soybean. Plant Sci. 2022, 320, 111283. [Google Scholar] [CrossRef]
- Roussis, A.; Flemetakis, E.; Dimou, M.; Kavroulakis, N.; Venieraki, A.; Aivalakis, G.; Katinakis, P. Nodulin PvNOD33, a putative phosphatase whose expression is induced during Phaseolus vulgaris nodule development. Plant Physiol. Biochem. 2003, 41, 719–725. [Google Scholar] [CrossRef]
- Li, C.; Li, C.; Zhang, H.; Liao, H.; Wang, X. The purple acid phosphatase GmPAP21 enhances internal phosphorus utilization and possibly plays a role in symbiosis with rhizobia in soybean. Physiol. Plant 2017, 159, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Ott, T.; van Dongen, J.T.; Gunther, C.; Krusell, L.; Desbrosses, G.; Vigeolas, H.; Bock, V.; Czechowski, T.; Geigenberger, P.; Udvardi, M.K. Symbiotic leghemoglobins are crucial for nitrogen fixation in legume root nodules but not for general plant growth and development. Curr. Biol. 2005, 15, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Oldroyd, G.E.; Murray, J.D.; Poole, P.S.; Downie, J.A. The rules of engagement in the legume-rhizobial symbiosis. Annu. Rev. Genet. 2011, 45, 119–144. [Google Scholar] [CrossRef] [PubMed]
- Sulieman, S.; Ha, C.V.; Schulze, J.; Tran, L.S. Growth and nodulation of symbiotic Medicago truncatula at different levels of phosphorus availability. J. Exp. Bot. 2013, 64, 2701–2712. [Google Scholar] [CrossRef] [PubMed]
- Thuynsma, R.; Valentine, A.; Kleinert, A. Phosphorus deficiency affects the allocation of below-ground resources to combined cluster roots and nodules in Lupinus albus. J. Plant Physiol. 2014, 171, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Li, C.C.; Gui, S.H.; Yang, T.; Walk, T.; Wang, X.R.; Liao, H. Identification of soybean purple acid phosphatase genes and their expression responses to phosphorus availability and symbiosis. Ann. Bot. 2012, 109, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Zhao, J.; Tian, J.; Chen, L.; Sun, Z.; Guo, Y.; Lu, X.; Gu, M.; Xu, G.; Liao, H. The high-affinity phosphate transporter GmPT5 regulates phosphate transport to nodules and nodulation in soybean. Plant Physiol. 2012, 159, 1634–1643. [Google Scholar] [CrossRef]
- Zhang, J.; Zhou, X.; Xu, Y.; Yao, M.; Xie, F.; Gai, J.; Li, Y.; Yang, S. Soybean SPX1 is an important component of the response to phosphate deficiency for phosphorus homeostasis. Plant Sci. 2016, 248, 82–91. [Google Scholar] [CrossRef]
- Araújo, A.P.; Plassard, C.; Drevon, J.J. Phosphatase and phytase activities in nodules of common bean genotypes at different levels of phosphorus supply. Plant Soil 2008, 312, 129–138. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Yang, Z.W.; Du, H.; Xing, X.Z.; Li, W.L.; Kong, Y.B.; Li, X.H.; Zhang, C.Y. A small heat shock protein, GmHSP17.9, from nodule confers symbiotic nitrogen fixation and seed yield in soybean. Plant Biotechnol. J. 2021, 20, 103–115. [Google Scholar] [CrossRef]
- Li, X.; Zheng, J.; Yang, Y.; Liao, H. INCREASING NODULE SIZE1 Expression Is Required for Normal Rhizobial Symbiosis and Nodule Development. Plant Physiol. 2018, 178, 1233–1248. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′ to 3′) |
|---|---|
| GmPAP4-RT-F1 | GAGCCGTTTGCGAGTACAAG |
| GmPAP4-RT-R1 | TCTGCATAGGAGCCAAGCAT |
| GmActin11-F | ATCTTGACTGAGCGTGGTTATTCC |
| GmActin11-R | GCTGGTCCTGGCTGTCTCC |
| Bar-F | CCATCGTCAACCACTACATCGAGACA |
| Bar-R | CTTCAGCAGGTGGGTGTAGAGCGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, X.; Zhang, H.; Yang, Z.; Xing, X.; Fu, Z.; Li, X.; Kong, Y.; Li, W.; Du, H.; Zhang, C. Overexpression of GmPAP4 Enhances Symbiotic Nitrogen Fixation and Seed Yield in Soybean under Phosphorus-Deficient Condition. Int. J. Mol. Sci. 2024, 25, 3649. https://doi.org/10.3390/ijms25073649
Sun X, Zhang H, Yang Z, Xing X, Fu Z, Li X, Kong Y, Li W, Du H, Zhang C. Overexpression of GmPAP4 Enhances Symbiotic Nitrogen Fixation and Seed Yield in Soybean under Phosphorus-Deficient Condition. International Journal of Molecular Sciences. 2024; 25(7):3649. https://doi.org/10.3390/ijms25073649
Chicago/Turabian StyleSun, Xi, Huantao Zhang, Zhanwu Yang, Xinzhu Xing, Zhao Fu, Xihuan Li, Youbin Kong, Wenlong Li, Hui Du, and Caiying Zhang. 2024. "Overexpression of GmPAP4 Enhances Symbiotic Nitrogen Fixation and Seed Yield in Soybean under Phosphorus-Deficient Condition" International Journal of Molecular Sciences 25, no. 7: 3649. https://doi.org/10.3390/ijms25073649
APA StyleSun, X., Zhang, H., Yang, Z., Xing, X., Fu, Z., Li, X., Kong, Y., Li, W., Du, H., & Zhang, C. (2024). Overexpression of GmPAP4 Enhances Symbiotic Nitrogen Fixation and Seed Yield in Soybean under Phosphorus-Deficient Condition. International Journal of Molecular Sciences, 25(7), 3649. https://doi.org/10.3390/ijms25073649